國立臺灣大學生物資源暨農學院農藝學系 博士論文
Department of Agronomy
College of Bioresources and Agriculture National Taiwan University
Doctoral Dissertation
非生物逆境下亞托敏及硒處理對小麥幼苗生理之影響
Physiological Effects of Azoxytrobin and Selenium on Wheat Seedlings under abiotic Stresses
藍秋月 Chiu-Yueh Lan
指導教授: 黃文達 博士 陳昶璋 博士 Advisors: Wen-Dar Huang, Ph.D.
Chang-Chang Chen, Ph.D.
中華民國 109 年 6 月
June, 2020
誌謝
本論文的完成,首先要感謝兩位指導教授 黃文達老師及衛生福利部國家中醫 藥研究所助理研究員 陳昶璋老師在實驗架構及論文撰寫上的指導,同時也感謝中 國文化大學園藝暨生物技術學系 林冠宏老師協助文章修改及投稿事宜,使投搞文 章在短期內得以順利被接受。論文文稿初成,復蒙中央研究院生物多樣性研究中心 副研究員 楊棋明老師、台灣中油股份有限公司煉製研究所研究員 許明晃博士、中 國文化大學園藝暨生物技術學系助理教授 黃盟元老師、行政院農業委員會桃園區 農業改良場副研究員 楊志維博士及台中鴻禧太平高爾夫球場執行副總理 陳宏銘 副總提供諸多寶貴意見,使論文更臻嚴謹及完善,特誌卷首,謹致由衷之謝忱。
與台大農藝系的緣份起始於 101 年 3 月份我參與了台灣大學進修推廣部農業 技術學分班第 3 期的學習。感謝當時所有的授課老師:王裕文老師、王淑珍老師、
林彥蓉老師、胡凱康老師、黃文達老師、常玉強老師、張孟基老師、陳凱儀老師、
陳尊賢老師、郭華仁老師、彭雲明老師、廖振鐸老師及劉力瑜老師引領我進入農藝 的殿堂,也開啟了黃文達老師與我截至目前為止 8 年多的師生之緣。若非黃文達 老師這一路以來的鼓勵,引領及提攜,我不可能進入台大農藝系修讀博班,不可能 在台大完成教育學程的學業,不可能堅持到最後一刻終於有機會寫到博士論文的 誌謝這一頁。再多的感謝也無法傳達的謝意,只願未來的我能將我在黃老師身上所 學到的精神繼續傳遞下去。薪火相傳,願這份精神能永存人間,綿延不盡。而我們 的師生之緣也還會一直持續下去,未來還請黃老師再繼續多多指教。
在 308 的這段日子,感謝黃秀鳳技士在生活上及實驗上的照顧,也感謝實驗室 歷年助理,研究生與大學部的研究夥伴:陳傑君、林盈如、朱衍臻、歐陽志翔、蔡 沛穎、林家伃、鄭瓊如、蔡顓均、邱昱潔、游筑涵,有了你們的協助與幫忙,我的 論文才能得以順利完成,非常感謝你們!
最後,感謝在我背後默默支持我的家人們,你們的支持是我最強大的後盾。也 感謝你們的包容,允許我以自己的步伐,找到我自己,也成為我自己。謝謝你們!
願你們永遠健康、幸福、快樂!
中文摘要
本研究的目的主要在評估亞托敏(azoxystrobin, AZ)和硒(Selenium, Se)
處理對小麥品種台中選 2 號(Triticum aestivum L. variety Taichung No. 2, TCS2)
在高溫和鹽逆境下的作用機制。在亞托敏實驗中,探討殺真菌劑亞托敏前處理對 小麥幼苗於突然來襲的熱逆境下(Heat stress, HT, 46℃)的生理機制影響。本試 驗分別以 0.4, 4, 40, 80 和 120 mg L-1的 AZ 預處理 4 天,再針對有經過 AZ 預處 理和未處理 AZ 的幼苗施以 46 oC 1 小時的高溫逆境,緊接著進行 1000 μmol m-2 s-1 之高光處理 20 分鐘。 高溫誘導的氧化逆境導致未處理亞托敏幼苗葉片中的
還原力降低及丙二醛含量增加,抗壞血酸過氧化物酶(ascorbate peroxidase, APX)
和過氧化氫酶(catalase, CAT)酶活性增強。然而在高溫逆境下的 AZ 預處理卻 導致葉綠素螢光,APX 和 CAT 活性以及清除 DPPH 能力的降低,並且隨著 AZ 處理濃度的增加,熱逆境所引起的生理損傷亦加劇。AZ 預處理雖可增加光合色 素的含量,然而綜合上述生理指標的結果表明 AZ 無法為小麥台中選 2 號在熱逆 境下提供保護作用。在硒與鹽害實驗中,主要探討22 μM 之硒處理在不同濃度
(0, 100, 200, 300 和 400 mM NaCl)的鹽逆境下對台中選 2 號幼苗的保護作用。
研究結果顯示,經過硒處理之小麥在葉綠素螢光參數,抗氧化酵素 (過氧化氫酶 CAT),抗氧化物質(總酚,類黃酮及黃青素含量),抗氧化能力 (清除 DPPH 自由 基能力、還原力分析),光合作用色素 (總葉綠素及類胡蘿蔔素),株高及根長皆 有較佳之表現,顯示22 μM 的硒處理確實對台中選 2 號幼苗在鹽脅迫下具有保
護效果。在硒及鹽害下之滲透逆境及離子逆境實驗之研究中,旨在區分鹽逆境引 起的滲透和離子脅迫對台中選2 號的影響,並評估 22 μM Se 處理對此二種脅迫
之調節效果。我們假設耐鹽性較強的台中選 2 號較易受到滲透脅迫的影響,而
22 μM 的 Se 處理對鹽害逆境所導致之滲透脅迫及離子脅迫皆發揮保護作用。為 了驗證此一假設,本實驗將聚乙二醇(PEG)和鹽 (NaCl) 調成 3 種等滲濃度,
最終滲透勢分別為 -1.05 MPa(24%(w / v)PEG 和 200 mM NaCl ), - 1.33 MPa(26.5%(w / v)PEG 和 250 mM NaCl)和-1.57 MPa(29%(w / v)PEG 和 300 mM NaCl)。研究結果顯示,相較於 PEG 所造成之滲透逆境,鹽處理所造成 的之離子逆境可使葉綠素螢光(ChlF)參數有較佳之表現,光合色素(總葉綠素 和類胡蘿蔔素) 之降解速度較慢,抗壞血酸過氧化物酶(APX)活性較好,且 丙二醛 (MDA)累積也較少。顯示在鹽害逆境下的滲透脅迫比離子脅迫對台中 選二號造成更大之生理影響。同時,我們觀察到22 μM Se 處理對鹽害逆境下之 滲透脅迫及離子脅迫均無法提供保護效果,顯示硒的有效處理濃度還需要更進 一步的實驗加以闡明。整體而言,本研究結果分別為亞托敏及硒處理下對台中選 2 號之小麥品種在熱逆境和鹽脅迫下之生理機制提供了更好的理解。
關鍵詞:小麥、台中選二號、亞托敏、硒、高溫逆境、鹽逆境、滲透逆境、離子 逆境、聚乙二醇
Abstract
The objective of this study was to evaluate the effect of azoxystrobin (AZ) and
Selenium (Se) on wheat (Triticum aestivum L.) variety Taichung No. 2 'Taichung SEL.2'
(TCS2) in high temperature and salt stress. In the AZ exp., the physiological mechanism
of the fungicide AZ in protecting against heat (HT, 46 °C) stress in wheat TCS2 seedlings
was investigated. Seedlings were pretreated with 0.4, 4, 40, 80, and 120 mg L-1 of AZ for
4 d. Next, AZ-pretreated and untreated seedlings were subjected to HT for 1 h followed
by 1000 μmol m-2 s-1 lighting for 20 min. HT induced oxidant stress which resulted in
a decrease in the reducing power, an increase in malondialdehyde, and enhanced enzyme
activities of ascorbate peroxidase (APX) and catalase (CAT) in leaves of untreated
seedlings. However, AZ-pretreated seedlings under HT displayed reductions in
chlorophyll fluorescence, APX and CAT activities, and the 1,1-diphenyl-2-picryl-
hydrazyl scavenging capacity. Physiological damage caused by HT was aggravated by an
increase in the AZ concentration. In addition, increased photosynthetic pigments were
also observed in leaves of AZ-pretreated and HT-exposed seedlings. The results suggest
that AZ does not provide a protective effect against HT stress. In the Se & salt exp., the
mitigative effects of 22 μM Se on TCS2 were investigated under different salt stress levels
(0, 100, 200, 300, and 400 mM NaCl). Results of the antioxidative capacity showed that
catalase (CAT) activity, non-enzymatic antioxidants (total phenols, total flavonoids, and
anthocyanins), 1,1-Diphenyl-2-Picryl-Hydrazyl (DPPH) radical-scavenging activity, and
the reducing power of Se-treated seedlings were enhanced under saline conditions. The
more-stabilized chlorophyll fluorescence parameters (maximal quantum yield of
photosystem II (Fv/Fm), minimal chlorophyll fluorescence (F0), effective quantum yield
of photosystem II (ΦPSII), quantum yield of regulated energy dissipation of photosystem
II (Y(NPQ)), and quantum yield of non-regulated energy dissipation of photosystem II
(Y(NO)) and the less-extensive degradation of photosynthetic pigments (total chlorophyll
and carotenoids) in Se-treated seedlings were also observed under salt stress. The
elongation of shoots and roots of Se-treated seedling was also preserved under salt stress.
Protection of these physiological traits in Se-treated seedlings might have contributed to
stable growth observed under salt stress. The present study showed the protective effect
of Se on the growth and physiological traits of wheat seedlings under salt stress. In the Se
and salt/PEG exp., the aim was to distinguish the effects of osmotic and ionic stress
induced by salt stress on TCS2 and evaluate the effect of 22 μM Se treatment on these
two stresses. We hypothesized that TCS2 is more susceptible to osmotic stress, and 22
μM Se treatment protects against osmotic stress and ionic stress caused by salt stress. In
order to verify this hypothesis, this experiment adjusted polyethylene glycol (PEG) and
salt (NaCl) to three isotonic concentrations, and the final osmotic potential was -1.05 MPa
(24% (w / v) PEG and 200 mM NaCl), - 1.33 MPa (26.5% (w / v) PEG and 250 mM
NaCl) and -1.57 MPa (29% (w / v) PEG and 300 mM NaCl). According to the results of
chlorophyll fluorescence parameters, APX activity, photosynthetic pigments (total
chlorophyll and carotenoids), malondialdehyde (MDA), and shoot height. The osmotic
stress caused by PEG was more harmful than the ionic stress caused by salt stress to TCS2.
Also, we observed that 22μM Se treatment could not protect against osmotic stress and
ionic stress under salt stress. The effective treatment concentration of Se required
furthering experimentation. Overall, the results of this study provide a better
understanding of the physiological mechanisms of TCS2 under thermal and salt stress
with the treatment of AZ and Se.
Key words: wheat; Taichung SEL.2; azoxystrobin; selenium; heat stress; salt stress;
osmotic stress; ionic stress; polyethylene glycol
Table of contents
中文摘要 ··· I Abstract ··· III List of Figure ···VIII List of Table··· XI List of Original Publications ··· XII
Chapter 1. Introduction ··· 1
Chapter 2. Physiological Effects of the Fungicide Azoxystrobin on Wheat Seedlings under Extreme Heat ··· 10
Abstract ··· 10
Introduction ··· 11
Materials and Methods ··· 14
Results and Discussion ··· 19
References ··· 27
Chapter 3. Protective Effects of Selenium on Wheat Seedlings under Salt Stress · 40 Abstract ··· 40
Introduction ··· 41
Materials and Methods ··· 44
Results ··· 50
Discussion ··· 54
Conclusions ··· 60
References ··· 61
Chapter 4. Comparison of the chlorophyll fluorescence and other physiological characteristics on Wheat seedlings influence by iso-osmotic stress of PEG and NaCl and their relative mitigation effect of Se ··· 76
Abstract ··· 76
Introduction ··· 78
Materials and methods ··· 82
Results ··· 86
Discussion ··· 91
Conclusion ··· 97
References ··· 98
Chapter 5. Conclusion ··· 118
List of Figure
Figure 2-1. Images of chlorophyll fluorescence in the maximum quantum yield of photosystem II (Fv/Fm). ... 35 Figure 2-2. The mean ± standard deviation (error bar) of the minimal (F0) and maximal (Fm) chlorophyll fluorescence, and the effective quantum yield of photosystem II (PSII) under illumination (ΦPSII) of leaves collected from wheat seedlings treated with azoxystrobin (AZ) and grown under the control (CK) or heated (HT) environment.. ... 36 Figure 2-3. The mean ± standard deviation (error bar) of non-photochemical quenching (NPQ), the quantum yield of regulated energy dissipation of photosystem II (PSII) (Y(NPQ)), and the quantum yield of non-regulated energy dissipation of PSII (Y(NO)) of leaves collected from wheat seedlings treated with azoxystrobin (AZ) and grown under the control (CK) or heated (HT) environment. ... 37 Figure2-4. The mean ± standard deviation (error bar) of ascorbate peroxidase (APX) and catalase (CAT) activities of leaves collected from wheat seedlings treated with azoxystrobin (AZ) and grown under the control (CK) or heated (HT) environment.
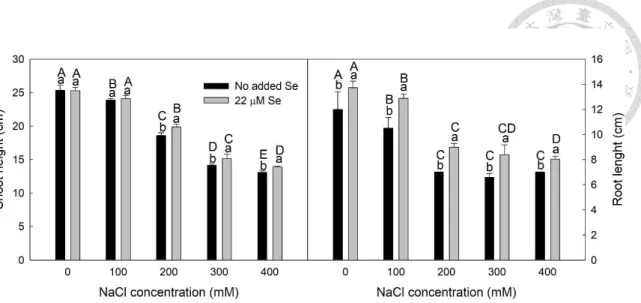
... 38 Figure 2-5. The mean ± standard deviation (error bar) of malondialdehyde (MDA) of leaves collected from wheat seedlings treated with azoxystrobin (AZ) and grown under the control (CK) or heated (HT) environment.. ... 39 Figure 3-1. Means ± SD (n = 5) of the shoot height and root length of untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under different NaCl concentrations. ... 72 Figure 3-2. Images and the mean ± SD (n = 4) of the maximum quantum yield of photosynthetic system II (Fv/Fm) and the minimal fluorescence of photosynthetic system II (F0) of leaves collected from untreated seedlings (No added Se) and Se- treated seedlings (22 μM Se) under different NaCl concentrations.. ... 73 Figure 3-3. Means ± SD (n = 4) of the effective quantum yield of photosystem II under illumination (ΦPSII), quantum yield of regulated energy dissipation of photosystem II (Y(NPQ)), and the quantum yield of non-regulated energy dissipation of photosystem II (Y(NO)) of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under different NaCl concentrations. ... 74 Figure 3-4. Means ± SD (n = 3) of Chl a, Chl b, Chl a+b, and carotenoids of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under different NaCl concentrations.. ... 75
Figure 4-1A. Means ± SD (n=5) of shoot height of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under three corresponding iso-osmotic potential creating by PEG & NaCl……….106 Figure 4-1B. Means ± SD (n=5) of shoot height of leaves under three corresponding iso-
osmotic potential creating by PEG & NaCl ... 107 Figure 4-2A. the mean ± SD (n=3) of the maximum quantum yield of photosynthetic system II (Fv/Fm) of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under three corresponding iso-osmotic potential creating by PEG & NaCl (-0.58 Mpa: 24% (w/v) PEG, 200mM NaCl; -1.33 Mpa:
26.5% (w/v) PEG, 250mM NaCl; -1.57 MPa: 29% (w/v) PEG, 300mM NaCl)……….108 Figure 4-2B. Images and the mean ± SD (n=3) of the maximum quantum yield of
photosynthetic system II (Fv/Fm) of leaves collected under three corresponding iso- osmotic potential creating by PEG & NaCl (-0.58 Mpa: 24% (w/v) PEG, 200mM NaCl; -1.33 Mpa: 26.5% (w/v) PEG, 250mM NaCl; -1.57 MPa: 29% (w/v) PEG, 300mM NaCl)………109 Figure 4-3A. Means ± SD (n=3) of the effective quantum yield of photosystem II under illumination (ΦPSII), non-photochemical quenching (NPQ) of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under three corresponding iso-osmotic potential creating by PEG & NaCl ... 110 Figure 4-3B. Means ± SD (n=3) of the effective quantum yield of photosystem II under illumination (ΦPSII), non-photochemical quenching (NPQ) of leaves ... 111 Figure 4-4A. Means ± SD (n=3) of APX activity of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under three corresponding iso-osmotic potential creating by PEG & NaCl ... 112 Figure 4-4B. Means ± SD (n=3) of APX activity of leaves collected from the seedlings treated with three corresponding iso-osmotic potential creating by PEG & NaCl113 Figure 4-5A. Means ± SD of chlorophyll (Chl a, Chl b, Chl a+b) and carotenoids of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under three corresponding iso-osmotic potential creating by PEG &
NaCl ... 114 Figure 4-5B. Means ± SD of chlorophyll (Chl a, Chl b, Chl a+b) and carotenoids of leaves collected from three corresponding iso-osmotic potential creating by PEG &
NaCl ... 115 Figure 4-6A. Means ± SD (n=3) of Malondialdehyde (MDA) concentration of leaves collected from untreated seedlings (No added Se) and Se-treated seedlings (22 μM Se) under three corresponding iso-osmotic potential creating by PEG &
Figure 4-6B. Means ± SD (n=3) of Malondialdehyde (MDA) concentration of leaves collected from the seedlings treated with three corresponding iso-osmotic potential creating by PEG & NaCl ………...……….…………117
List of Table
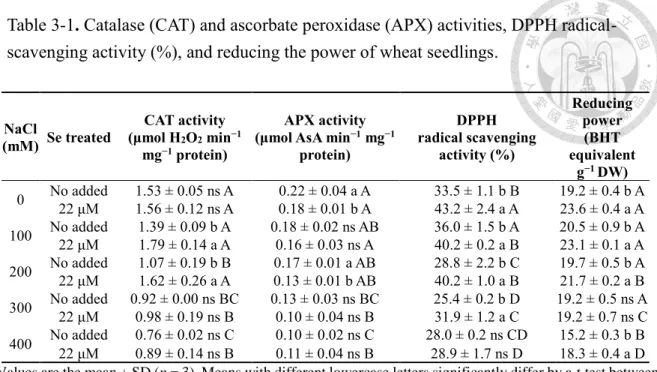
Table 2-1. Effect of azoxystrobin (AZ) on the level of the DPPH radical scavenging capacity (expressed in μg BHT equivalent g-1 dry weight (DW)) and reducing power (express in mg BHT equivalent g-1 DW) in seedlings leaves collected from the control (CK) or heated (HT) condition ... 33 Table 2-2. Effect of azoxystrobin (AZ) on the level of total chlorophyll (Chl; expressed in mg g-1 dry weight (DW)), the Chl a/b ratio, carotenoids (Car; expressed in mg g-
1 DW), and the Chl/Car ratio in seedling leaves collected from the control (CK) or heated (HT) condition. ... 34 Table 3-1. Catalase (CAT) and ascorbate peroxidase (APX) activities, DPPH radical-
scavenging activity (%), and reducing the power of wheat seedlings. ... 70 Table 3-2. Total phenols, total flavonoids, and anthocyanin concentrations of wheat seedlings. ... 71
List of Original Publications
This thesis is published on the following articles:
Lan CY, Lin KH, Huang WD, Chen CC (2019). Protective Effects of Selenium on Wheat Seedlings under Salt Stress. Agronomy, 9(6): 272.
Lan CY, Lin KH, Huang WD, Chen CC (2019). Physiological Effects of the Fungicide Azoxystrobin on Wheat Seedlings under Extreme Heat. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 47(3) 683-690.
Lan CY, Lin KH, Chen CL, Huang WD, Chen CC (2020). Comparisons of chlorophyll fluorescence and physiological characteristics of wheat
seedlings influenced by iso-osmotic stresses from polyethylene glycol and
sodium chloride. Agronomy, 10(3): 325
Introduction
Wheat (Triticum aestivum L.), the world’s third most important primary cereal with more than 600 million tons of global production, preceded only by corn and rice (Asseng et al., 2011), provides the main source of carbohydrates for 35% ~ 40% of the world’s population (Chen et al., 2017). However, under the influence of global warming, extreme environmental stresses such as heat, drought (Fábián et al., 2019), and salinity (Yadav et al., 2019) have impacted wheat (Triticum aestivum L.) production around the world with increasing frequency and intensity. Therefore, it is crucial to understand the mechanisms by which plants encounter abiotic stresses to promote crop performance under adverse conditions. Many protective chemicals have been used to mitigate the adverse effects caused by abiotic stresses, such as salicylic acid (Borsani et al., 2001; Lu
et al., 2009), caffeic acid (Bhardwaj and Ramandeep, 2017), NO (Song et al., 2013), H2O2
(Uchida et al., 2002), spermidine (Mostofa et al., 2014), and magnetized water (Hasan et
al., 2018). In addition, azoxystrobin (AZ) (Grossmann and Retzlaff, 1997) and selenium
(Se) (Djanaguiraman et al., 2005) are certified as agents for promoting yields and delaying senescence in some crops.
Azoxystrobin (AZ) is one of the widely used strobilurins used as systemic
fungicides (Giuliani et al., 2011). It is also certified as a chemical agent for promoting
yields and delaying senescence in some crops, such as rice and wheat (Grossmann and
Retzlaff, 1997; Wu and Tiedemann, 2001). Therefore, it is also classified as a plant
bioregulator (Rademacher, 2004). Previous studies report that strobilurins stimulate
nitrogen assimilation (Debona et al., 2016), mediate plant hormones, cause leaf aging
delays, increase chlorophyll concentrations (Grossmann and Retzlaff, 1997), mitigate
oxidative stress (Wu and von Tiedemann, 2001; Zhang et al., 2010), and improve plant
water use efficiency and stabilize yields during droughts (Giuliani et al., 2011; Cantore
et al., 2016). Furthermore, strobilurins also enhance the activity of nitrate reductase, a
key enzyme involved in plant nitrogen assimilation, while increasing nitrogen
assimilation (Debona et al., 2016). Conversely, Nason et al., (2007), Swoboda and
Pedersen (2009), and Amaro et al., (2018) found that strobilurins might not be able to
provide protective effects under stress conditions. The effects of strobilurins are complex
and require further clarification.
Selenium (Se) is considered a beneficial element to plants (Pilon-Smits et al.,
2009). Previous studies indicated that Se can delay senescence (Djanaguiraman et al.,
2005; Xue et. al., 2001) and promote the vegetative and reproductive growth of plants
(Hajiboland et al., 2012). Previous research reveals that the possible protective
mechanisms of Se in plants against stresses include the enhancement of antioxidant
enzyme activity (peroxidase (POD) and catalase (CAT), etc.) and increase the levels of
antioxidant compounds (anthocyanins, flavonoids, phenolic compounds, etc.), and these
antioxidant features thus reduce stress-induced oxidative situations (Chu et al., 2010). In
addition, Se can improve plant photosynthesis by increasing the efficiency of
photosystem II (PSII), enhancing chlorophyll fluorescence, and reducing the degradation
of chlorophyll concentration (Chu et al., 2010). Moreover, Se contributes to water status
regulation in plants by promoting water uptake efficiency in roots and reducing water loss
from tissues (Kuznetsov et al., 2003). In addition, Se stimulates plant growth by
promoting the integrity of the membrane system, which results in root and shoot
elongation and biomass accumulation (Djanaguiraman et al., 2005; Xiaoqin et al., 2009;
Hawrylak-Nowak et al., 2009; Hu et al., 2013; Sun et al., 2010). Research by Hawrylak-
Nowak (2009) shows that Se particularly supports root system development. However,
Se exerts a dual effect in plants (Djanaguiraman et al., 2005), in that while it can stimulate
plant growth and provide beneficial effects as a micronutrient at low concentrations, it is
harmful to plants at higher concentrations. The positive effects of Se depend on its form,
dose, and plant genotype (Sieprawska et al., 2015). Two forms of inorganic Se can be
found, depending on the pH and redox potential of the soil, one being selenite and the
other selenate, each of which exhibits different availabilities to and effects on plants. In
our study, wheat was treated with sodium selenite (Na2SeO3) in an acidic nutrient solution,
primarily as HSeO3- (Guerrero et al., 2014). The recommended Se doses for hydroponic
conditions are usually < 1 mg L−1 (29 μM) (Pilon-Smits et al., 2009). Conversely, toxic
effects can occur when the Se dosage is > 0.35 mg L−1 (10 μM). The effects and beneficial
dosages of Se on wheat (Triticum aestivum L.) cultivar ‘Taichung SEL.2’ (TCS2) need to
be explored.
In the present study, the functions and effects of AZ and Se on wheat (Triticum
aestivum L.) cultivar ‘Taichung SEL.2’ (TCS2) were investigated. In AZ experiments, the
effects of AZ applications on the ChlF and antioxidant activity of wheat seedlings grown
under high heat (HT) were evaluated. In Se and salt experiments, the growth and
physiological effects of Se on wheat subjected to salt stress were conducted. In Se and
salt/PEG experiments, the effects of applying Se to wheat under three iso-osmotic
concentrations of salt and PEG were evaluated.
References
Asseng S, Foster I, Turner NC (2011). The impact of temperature variability on wheat
yields. Global Change Biology, 17(2): 997-1012.
Bhardwaj RD, Ramandeep K (2017). Protective role of pre-treatment with different
phenolic acids in wheat seedlings against heat stress induced oxidative damage.
Indian Journal of Agricultural Biochemistry, 30(2): 147-155.
Borsani O, Valpuesta V, Botella MA (2001). Evidence for a role of salicylic acid in the
oxidative damage generated by NaCl and osmotic stress in Arabidopsis seedlings.
Plant Physiology, 126(3): 1024-1030.
Cantore V, Lechkar O, Karabulut E, Sellami MH, Albrizio R, Boari F and Todorovic M
(2016). Combined effect of deficit irrigation and strobilurin application on yield,
fruit quality and water use efficiency of“cherry” tomato (Solanum lycopersicum
L.). Agricultural Water Management, 167:53-61.
Chen YE, Cui JM, Su YQ, Zhang CM, Ma J, Zhang ZW, Yuan S (2017). Comparison of
phosphorylation and assembly of photosystem complexes and redox homeostasis
in two wheat cultivars with different drought resistance. Scientific Reports, 7(1):
12718.
Chu J, Yao X, Zhang Z (2010). Responses of wheat seedlings to exogenous selenium
supply under cold stress. Biological Trace Element Resarch, 136:355–363.
Debona D, Rodrigues FA (2016). A strobilurin fungicide relieves Bipolaris oryzae
induced oxidative stress in rice. Journal of Phytopathology, 164(9):571-581.
Djanaguiraman M, Devi DD, Shanker AK, Sheeba JA, Bangarusamy U (2005). Selenium
– an antioxidative protectant in soybean during senescence. Plant and Soil, 272(1):
77-86.
Fábián A, Sáfrán E, Szabó-Eitel G, Barnabás B, Jäger K (2019). Stigma functionality and
fertility are reduced by heat and drought co-stress in wheat. Frontiers in Plant
Science, 10: 244-244.
Giuliani MM, Nardella E, Gatta G, Caro AD, Quitadamo M (2011). Processing tomato
cultivated under water deficit conditions: the effect of azoxystrobin. Acta
Horticulturae, 914:287-294.
Grossmann K, & Retzlaff G (1997). Bioregulatory effects of the fungicidal strobilurin
kresoxim-methyl in wheat (Triticum aestivum). Pesticide Science, 50(1): 11-20.
Guerrero B, Llugany M, Palacios O, Valiente (2014). Dual effects of different selenium
species on wheat. Plant Physiology Biochemistry, 83:300–307.
Hajiboland R (2012). Selenium supplementation stimulates vegetative and reproductive
growth in canola (Brassica napus L.) plants. Acta Agriculturae Slovenica, 99:13.
Hasan MM, Alharby HF, Hajar AS, Hakeem KR, Alzahrani Y (2018). Effects of
magnetized water on phenolic compounds, lipid peroxidation and antioxidant
activity of Moringa species under drought stress. Journal of Animal and Plant
Sciences, 28(3): 803-810.
Hawrylak-Nowak B (2009). Beneficial Effects of Exogenous Selenium in Cucumber
Seedlings Subjected to Salt Stress. Biological Trace Element Research, 132:259–
269.
Hu K, Zhang L, Wang J, You Y (2013). Influence of selenium on growth, lipid
peroxidation and antioxidative enzyme activity in melon (Cucumis melo L.)
seedlings under salt stress. Acta Societatis Botanicorum Polloniae, 82:193–197.
Kuznetsov VV, Kholodova V Kuznetsov VV, Yagodin B (2003). In Selenium Regulates
the Water Status of Plants Exposed to Drought. Doklady Biological Sciences,
390:266–268.
Lu J, Zhang R, Zong X, Wang S, He G (2009). Effect of salicylic acid on heat resistance
of rice seedling under heat stress. Chinese Journal of Eco-Agriculture, 17(6):
1168-1171.
Mostofa MG, Yoshida N, Fujita M (2014). Spermidine pretreatment enhances heat
tolerance in rice seedlings through modulating antioxidative and glyoxalase
systems. Plant Growth Regulation, 73(1): 31-44.
Nason MA, Bartlett D, Farrar J (2007). Strobilurin fungicides induce changes in
photosynthetic gas exchange that do not improve water use efficiency of plants
grown under conditions of water stress. Pest Management Science, 63(12):1191-
1200.
Pilon-Smits EA, Quinn CF, Tapken W, Malagoli M, Schiavon M (2009). Physiological
functions of beneficial elements. Current Opinion in Plant Biology, 12:267–274.
Song L, Yue L, Zhao H, Hou M (2013). Protection effect of nitric oxide on photosynthesis
in rice under heat stress. Acta physiologiae plantarum, 35(12): 3323-3333.
Sieprawska A, Kornaś A, Filek M (2015). Involvement of selenium in protective
mechanisms of plants under environmental stress conditions–review. Acta
Biological Cracoviensia series Boanica, 57:9–20.
Sun HW, Ha J, Liang SX, Kang WJ (2010). Protective role of selenium on garlic growth
under cadmium stress. Communications in Soil Science and Plant Analysis,
41:1195–1204.
Swoboda C, Pedersen P (2009). Effect of fungicide on soybean growth and yield.
Agronomy Journal, 101(2):352-356.
Uchida A, Jagendorf AT, Hibino T, Takabe T, Takabe T (2002). Effects of hydrogen
peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant
Science, 163(3): 515-523.
Wu Y, von Tiedemann AV (2001). Physiological effects of azoxystrobin and
epoxiconazole on senescence and the oxidative status of wheat. Pesticide
Biochemistry and Physiology, 71(1):1-10.
Xiaoqin Y, Jianzhou C, Guangyin W (2009). Effects of drought stress and selenium supply
on growth and physiological characteristics of wheat seedlings. Acta Physiologiae
Plantarum, 31:1031–1036.
Xue T, Hartikainen H (2000). Association of antioxidative enzymes with the synergistic
effect of selenium and UV irradiation in enhancing plant growth. Agricultural and
Food Science in Finland, 9:177–186.
Yadav SP, Bharadwaj R, Nayak H, Mahto R, Singh RK, Prasad SK (2019). Impact of salt
stress on growth, productivity and physicochemical properties of plants: A Review.
International Journal of Chemical Studies, 7(2): 1793-1798
Zhang YJ, Zhou MG, Wang HC, Zhang X, Chen CJ (2010). Effects of fungicides JS399-
19, azoxystrobin, tebuconazloe, and carbendazim on the physiological and
biochemical indices and grain yield of winter wheat. Pesticide Biochemistry and
Physiology, 98(2):151-157.
Chapter 2. Physiological Effects of the Fungicide Azoxystrobin on Wheat Seedlings under Extreme Heat
Lan CY, Lin KH, Huang WD, Chen CC (2019). Physiological Effects of the Fungicide Azoxystrobin on Wheat Seedlings under Extreme Heat. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 47(3) 683-690.
Abstract
Azoxystrobin (AZ) is not only a fungicide used for disease control, but also a protective chemical for crops against specific stresses. The physiological mechanism of
the fungicide AZ in protecting against heat (HT, 46 °C) stress in wheat (Triticum aestivum
L.) seedlings was investigated. ‘Taichung SEL.2’ variety seedlings were pretreated with
0.4, 4, 40, 80, and 120 mg L-1 of AZ for 4 d. Next, AZ-pretreated and untreated seedlings were subjected to HT for 1 h followed by 1000 μmol m-2 s-1 lighting for 20 min. HT
induced oxidant stress which resulted in a decrease in the reducing power, an increase in
malondialdehyde, and enhanced enzyme activities of ascorbate peroxidase (APX) and
catalase (CAT) in leaves of untreated seedlings. However, AZ-pretreated seedlings under
HT displayed reductions in chlorophyll fluorescence, APX and CAT activities, and the
1,1-diphenyl-2-picryl-hydrazyl scavenging capacity. Physiological damage caused by HT
was aggravated by an increase in the AZ concentration. In addition, increased
photosynthetic pigments were also observed in leaves of AZ-pretreated and HT-exposed
seedlings. The results suggest that AZ does not provide a protective effect against HT
stress.
Keywords: antioxidant activity; azoxystrobin; chlorophyll fluorescence; heat stress;
wheat
Introduction
Under the influence of global warming, heat waves have impacted wheat production
around the world with increasing frequency and intensity (Teixeira et al., 2013; Asseng
et al., 2014). Heat is one of the main environmental factors influencing wheat yields and
physiological senescence, such as inhibiting antioxidant enzyme activities and enhancing
lipid peroxidation (Zhao et al., 2007). Maintaining wheat’s physiological functions under
sudden heat waves is an urgent issue, as well as preserving wheat yields and quality from
acute and unexpected heat waves. In order to promote crop performances under adverse
conditions, applying protective chemicals is one preventive strategy to protect against
stresses (Gondim et al., 2012).
Introduced in the 1990s, strobilurins are a group of chemicals used as systemic
fungicides, and one of the most widely used strobilurin is azoxystrobin (AZ) (Giuliani et
al., 2011). Strobilurins are also certified as chemical agents for promoting yields and
delaying senescence in some crops, such as rice and wheat (Grossmann and Retzlaff,
1997; Wu and Tiedemann, 2001). Therefore, strobilurins are also classified as plant
bioregulators (Rademacher, 2004). Previous studies reported that strobilurins stimulate
nitrogen assimilation (Debona et al., 2016), mediate plant hormones, cause leaf aging
delays, increase chlorophyll concentrations (Grossmann and Retzlaff, 1997), mitigate
oxidative stress (Wu and Tiedemann, 2001; Zhang et al., 2010), and improve plant water
use efficiency and stabilize yields during droughts (Giuliani et al., 2011; Cantore et al.,
2016). Furthermore, strobilurins also enhance the activity of nitrate reductase, a key
enzyme involved in plant nitrogen assimilation, while raising nitrogen assimilation
(Debona et al., 2016). Grossmann and Retzlaff (1997) demonstrated that kresoxim-
methyl, one type of strobilurin with auxin-like activity, induced plant morphogenesis and
differentiation, reduced the activity of aminocyclopropane-1-carboxylic acid synthase
which is involved in the ethylene synthesis pathway, and increased chlorophyll (Chl)
concentrations in leaves, suggesting that strobilurins might provide protective effects
against senescence. Moreover, application of AZ increased the antioxidant enzyme
activity with a reduction in free radicals in plants, and reduced protein concentrations and
electrolyte leakage from leaves (Wu and von Tiedemann, 2001; Zhang et al., 2010).
Cantore et al. (2016) and Giuliani et al. (2011) indicated that AZ and other strobilurin
fungicides reduced stomatal conductance and water evaporation, and promoted the water
use efficiency (WUE) in tomatoes cultivated in an arid environment. The effects of AZ
on the WUE of plants are related to an increase of endogenous abscisic acid and mediation
of stomatal closure (Venancio et al., 2003). However, Nason et al. (2007) observed that
the beneficial effects which strobilurins provided of alleviating the WUE and
evapotranspiration of crops grown in water deficient environments were extremely
limited, and this resulted in a reduction in the maximum quantum yield (Fv/Fm). Debona
et al. (2016) revealed that AZ mediated stomatal movements and inhibited the
photosynthesis capacity, but did not influence Chl fluorescence (ChlF) or levels of
photosynthetic pigments. The effects of strobilurins on stomata are complex and require
further clarification. In addition, Swoboda and Pedersen (2009) illustrated that
pyraclostrobin, a kind of strobilurin, did not promote the growth or yields of soybeans.
Amaro et al. (2018) observed that AZ blocked electron transfer of the cytochrome-bc1
complex in mitochondria and inhibited the respiration of plants, followed by a decrease
in adenosine triphosphate and a reduction in the osmotic potential of guard cells, which
was associated with the degree of stomatal opening. The main mechanism of AZ in
reducing pathogen-induced oxidative stress from pathogen infections was to limit the
expansion of the pathogen rather than increasing the antioxidant activity (Debona and
Rodrigues, 2016). Strobilurin-induced delay of senescence in plants is well described.
Unfortunately, strobilurin might not be able to provide protective effects when a plant is
under a water deficit or is infected with a pathogen. However, Pedersen (2016) found that
AZ promoted endogenous cytokinins and phenolic components of creeping bentgrass for
maintaining physiological functions under heat stress. Nevertheless, there is no study on
whether strobilurins can also provide crops with protective effects during heat stress. In
this study, the effects of AZ applications on the ChlF and antioxidant activities of wheat
seedlings grown under high heat (HT) were evaluated.
Materials and Methods
Plant and growth conditions
Wheat (Triticum aestivum L.) cultivar ‘Taichung SEL.2’ (TCS2), one of the most
widely grown wheat cultivars in Taiwan, was used in this study. The seeds were sterilized
with 1% hydrogen peroxide for 5 min, washed with distilled water, and germinated in
Petri dishes on wetted filter paper at 25 °C in the dark. After 24 h of incubation, uniformly
germinated seeds were selected and cultivated in 150 ml beakers containing one-fifth-
strength Hoagland nutrient solution (Hoagland and Arnon, 1950), and the solution was
replaced every 2 d. Hydroponically cultivated wheat seedlings were raised in growth
chambers with fluorescent lamp lighting at 30 and 25 °C at day and night, respectively,
under a 12-h photoperiod. The photosynthetic photon flux density (PPFD) was uniformly
set to 300 μmol m-2 s-1.
Experimental treatments
Hydroponically grown seedlings that had reached stage Z1.0 (Zadoks et al., 1974) on
day 4 were treated with the AZ fungicide (250 g AI L-1, AmistarR, Syngenta Limited,
Waterford, Ireland) at concentrations of 0.4, 4, 40, 80, and 120 mg L-1 for 4 d. AZ was
added to the nutrient solution according to the concentration of each AZ pretreatment.
After AZ treatment, these seedlings were placed in a high temperature of 46 °C for 1 h in
the dark as the HT condition. There were also a group of seedlings grown under HT
without AZ pretreatment. Untreated seedlings were used as a control (CK). The
experiment was independently performed three times for a randomized design of growth
conditions.
Measurements of chlorophyll fluorescence (ChlF)
The fluorescence parameters in seedling leaves were determined after 1 h of HT in
the dark. ChlF was measured in the middle portion of the first leaf of each seedling taken
at ambient temperature with Chl fluorimeter imaging-PAM (Walz, Effeltrich, Germany).
Actinic light and saturating light intensities were set to 500 and 7200 μmol m-2 s-1 of
photosynthetically active radiation (PAR), respectively, and then the effective quantum
yield of photosystem (PS)II under illumination (ΦPSII) in leaves was determined after
1000 μmol m-2 s-1 (300 μmol m-2 s-1 for CK) lighting for 20 min. The minimal (F0) and
maximal (Fm) ChlF, maximum quantum yield of PSII (Fv/Fm), ΦPSII, non-photochemical
quenching (NPQ), the quantum yield of regulated energy dissipation of PSII (Y(NPQ)),
the quantum yield of non-regulated energy dissipation of PSII (Y(NO)), and the relative
electron transfer rate (ETR) were measured and calculated according to previously
described methods (Vankooten and Snel, 1990; Kramer et al., 2004).
Measurement of ascorbate peroxidase (APX) and catalase (CAT) activity
APX activity was determined using the method of Nakano and Asada (1981). Briefly,
0.06 g of the latest newly expanded leaf was placed in 2 mL sodium phosphate buffer (50
mM, pH 6.8) in an ice bath for extraction and centrifuged at 4 °C and 12,000 g for 20 min.
The supernatant (0.1 mL) was collected, followed by the sequential addition of 2.7 mL of
potassium phosphate buffer (150 mM, pH 7.0), 0.4 mL of ethylenediaminetetraacetic acid
(EDTA, 0.75 mM), 0.5 ml of H2O2 (6 mM), 0.5 mL of H2O, and 0.5 mL of ascorbate (1.5
mM) and then mixed well. The absorbance at 290 nm (A290) of the sample solution was
determined every 15 s for 1 min using a spectrophotometer (Hitachi U3010, Tokyo,
Japan). The blank containing the same mixture with no enzyme extract was also measured.
CAT activity was measured based on the method of Kato and Shimizu (1987). Briefly,
0.03 g of the latest newly expanded leaf was placed in 2 mL of sodium phosphate buffer
(50 mM, pH 6.8) in an ice bath for extraction and centrifuged at 4 °C and 12,000 g for 20
min. The supernatant (0.2 mL) was collected, followed by adding 2.7 mL of sodium
phosphate buffer (100 mM, pH 7.0), 0.05 mL of H2O, and 0.05 ml of H2O2 (1 M), and
then mixing well. The absorbance of the sample solution at 240 nm (A240) was
determined every 15 s for 1 min. The blank containing the same mixture with no enzyme
extract was also measured.
Measurement of 1,1-diphenyl-2-picryl-hydrazyl (DPPH) scavenging capacity and the
reducing power
The DPPH scavenging capacity was determined using the method of Shimada et al.
(1992). Briefly, 160 μL of a methanol extract of the sample combined with methanol or
standard solution of butylated hydroxytoluene (BHT) was added to 40 μL of a freshly
prepared DPPH solution (1mM) to initiate the antioxidant-radical reaction at room
temperature. The control was 160 μL of sample extract, methanol, or BHT solution diluted
to 200 μL. The absorbance of the reaction mixture was determined at 517 nm during a
30-min reaction time. The DPPH scavenging capacity was calculated using a curve of
BHT standards. Results are expressed as μg BHT equivalent g-1 dry weight (DW).
The reducing power was determined using the method of Oyaizu (1986). Briefly, 0.3
mL of the methanol extract from a leaf was placed in 0.3 mL of sodium phosphate buffer
(0.2 M, pH 6.6) and 0.3 mL of 1% K3Fe (CN)6 in a water bath at 50 °C for 20 min,
immediately placed in 0.3 mL of 10% TCA in an ice bath, and then centrifuged at 9000
rpm for 10 min. The supernatant (0.5 mL) was mixed well with 0.5 mL distilled water
and 0.1 mL FeCl3·H2O (0.1%). The absorbance of the reaction mixture was determined
at 700 nm during the 10-min reaction. The reducing power was calculated using a curve
of BHT standards. Results are expressed as mg BHT equivalent g-1 DW.
Determination of the malondialdehyde (MDA) concentration
MDA was determined using a previously described method (Heath and Packer, 1968).
Briefly, lyophilized sample powder (0.03 g) was mixed with 1 mL of 5% TCA, and then
centrifuged at 10,000 rpm and 20 °C for 5 min. The supernatant (250 μL) was added to 1
mL of 0.5% thiobarbituric acid (TBA) which was made up with 20% TCA. The mixture
was placed in a water bath at 95 °C for 30 min, and then immediately cooled in an ice
bath. The reaction mixture was centrifuged at 3000 rpm and 20 °C for 10 min, and the
absorbance was determined at 532 and 600 nm. The blank was the same reaction mixture
with no sample extract.
Determination of the photosynthetic pigment concentrations
The photosynthetic pigment concentrations were determined using the method of
Yang et al. (1998). Briefly, 0.01 g of lyophilized sample powder was extracted with 10
ml of an 80% acetone solution, and then centrifuged at 4500 rpm for 5 min. The
supernatant of the sample extract was tested to determine the absorbance of Chl a, Chl b,
and carotenoids (Cars) in acetone at 663.6, 646.6, and 440.5 nm, respectively.
Statistical analyses
All measurements were evaluated for significance using an analysis of variance
(ANOVA) followed by a least significant difference (LSD) test at the p < 0.05 level. All
statistical analyses were conducted using R i386 3.5.1 software (https://cran.r
project.org/bin/windows).
Results and Discussion
Chlorophyll fluorescence (ChlF)
The response of ChlF can be used to evaluate the physiological condition of
photosynthetic tissues in plants. Our preliminary data (data not shown) indicated that the
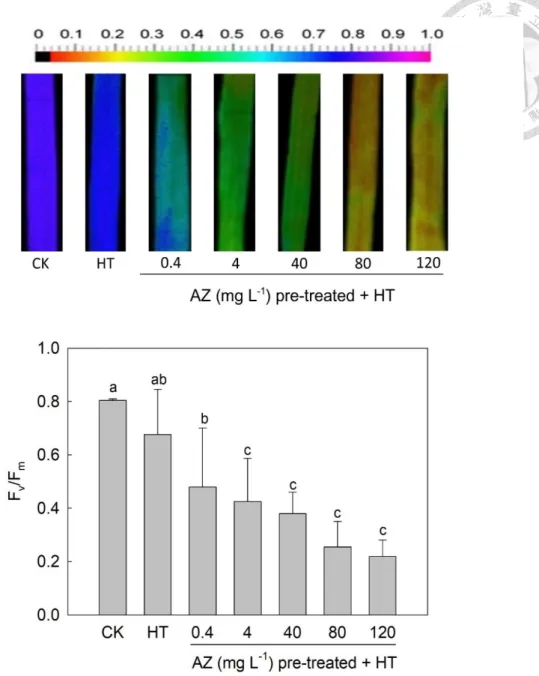
light saturation point in leaves of seedlings was at 500 μmol m-2 s-1. Fv/Fm in leaves was
determined after dark adaption with HT (CK at 30 °C) for 1 h. With the exception of ΦPSII,
other ChlF parameters were determined at 500 μmol m-2 s-1. ΦPSII was determined after
1000 μmol m-2 s-1 treatment (CK under 300 μmol m-2 s-1) for 20 min. Fv/Fm and ΦPSII are
widely used to estimate the status under heat stress (Sun et al., 2006), and Fv/Fm in CK
seedling leaves was 0.80. The difference in Fv/Fm of leaves of untreated seedlings grown
under HT and CK was insignificant. However, Fv/Fm values of AZ-pretreated seedlings
grown under HT were significant lower than that of CK, and continued to decrease from
0.48 to 0.22 with an increase in the pretreated AZ concentration from 0.4 to 120 mg L-1
(Fig. 2-1). A similar trend was observed in ΦPSII in leaves of heated seedlings after
exposure to light at 1000 μmol m-2 s-1, and CK seedlings exposed to light at 300 μmol m-
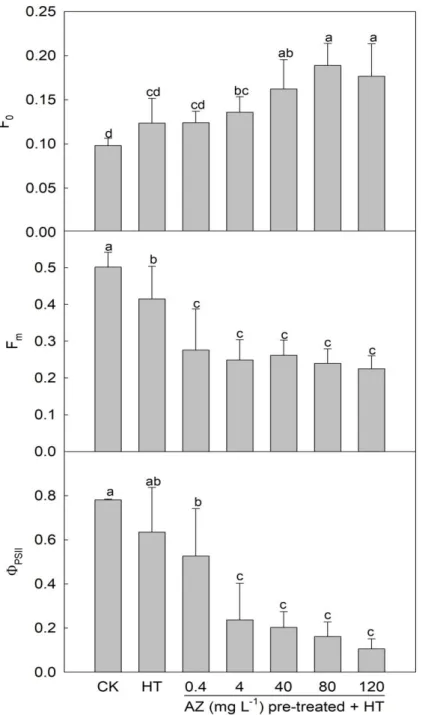
2 s-1 for 20 min (Fig. 2-2). Meanwhile, an increase in F0 and a decrease in Fm were
observed in leaves of AZ-pretreated seedlings. Furthermore, a rise in the AZ concentration
significantly influenced F0, but not Fm. F0 is a fluorescent signal when the PSII reaction
center is fully open (Sun et al., 2006), and an increase in F0 usually indicates that a plant
is under stress (Song et al., 2013). NPQ indicates the ability of plants to dissipate excess
light energy, and is one of protective mechanisms against high light stress, while Y(NPQ)
and Y(NO) are important indicators of photo-protection and photo-damage, respectively
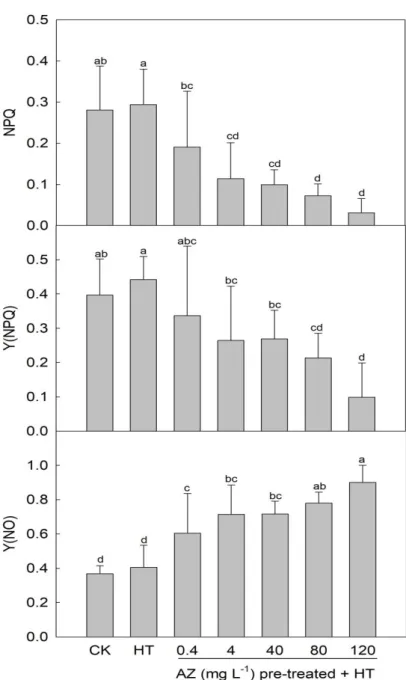
(Kramer et al., 2004). Lower NPQ levels were detected in leaves of seedlings pretreated
with higher AZ concentrations and exposed to HT (Fig. 2-3), and the dynamics of Y (NPQ)
also showed a similar pattern. On the other hand, Y(NO) in seedling leaves was enhanced
with an increase in the AZ concentration, indicating that AZ pretreatment provided no
protective effect for photosynthesis against HT. ChlF is an ideal tool for evaluating
damage to a plant’s photosynthetic tissues. In this study, decreases in Fv/Fm and ΦPSII in
seedling leaves with an increase in the AZ concentration suggest that the damage level of
the D1-protein was more severe at higher pretreatment AZ concentrations. On the other
hand, the increase in F0 also suggests that the light-harvesting complex had suffered
irreversible damage, and/or the ability to transmit light energy from the antenna system
to the PSII reaction center had degraded (Song et al., 2013). Meanwhile, the fall in NPQ
and Y (NPQ) and increase in Y (NO) in seedling leaves also indicate loss of photo-
protective ability and expansion of photo-damage (Kramer et al., 2004). These responses
of ChlF parameters are consistent with results of previous studies (Nason et al., 2007;
Debona et al., 2016) and suggest that an increase in the pretreated AZ concentration
caused greater physiological damage to the photosystem in leaves. These
disadvantageous effects of strobilurin on ChlF might have resulted from blockage of the
transmission of electrons between PSII and PSI because of the combination of strobilurin
and Qi in the cytochrome bf complex in chloroplasts (Nason et al., 2007). However, other
studies reported that the foliar application of AZ might inhibit stomatal movement rather
than ChlF, and would result in inefficient gas exchange (Debona et al., 2016; Amaro et
al., 2018). In this study, AZ was added to the hydroponic solution rather than being
applied to the foliage, and the impact of strobilurin on ChlF should be due to blockage of
the transmission of electrons between PSII and PSI.
Activities of APX and CAT, DPPH scavenging capacity, and reducing power
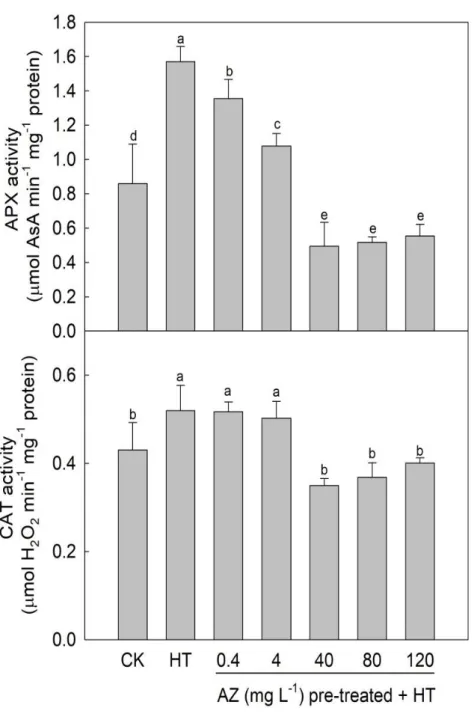
APX activity in leaves of untreated seedlings grown under HT improved 86% more
than that of CK leaves (Fig. 2-4), indicating that HT induced APX activity. However, a
continuous reduction in APX activity of AZ-pretreated and heated seedlings from 1.36 to
0.55 μmol ascorbate min-1 mg-1 protein was observed with an increase in the AZ
concentration up to 40 mg L-1. APX activities in leaves of seedlings subjected to
pretreatment with AZ at 40 mg L-1 or higher were significantly lower than that of CK
leaves. The dynamics of CAT activity in seedlings showed a different pattern from the
results of APX.
CAT activity of HT seedlings improved 24% more than that of CK seedlings (0.43
μmol H2O2 min-1 mg-1 protein), but did not increase as sharply as did APX. Furthermore,
the sudden reduction in CAT activity from 0.50 to 0.35 μmol H2O2 min-1 mg-1 protein was
determined in leaves of seedlings pretreated with AZ 40 mg L-1 and grown under HT,
implying that both APX and CAT activities in leaves of AZ pretreated and heated
seedlings were inhibited, and the critical concentration of AZ pretreatment was 40 mg L-
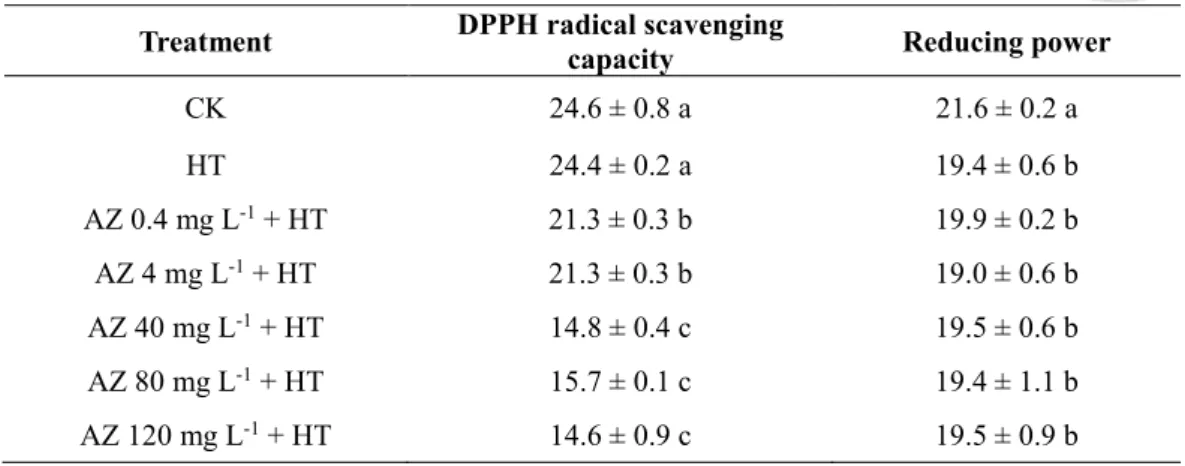
1. The difference in the DPPH scavenging capacity between CK (24.6 μg BHT equivalent
g-1 DW) and HT leaves of seedlings (24.4 μg BHT equivalent g-1 DW) was insignificant,
but a significant reduction in DPPH scavenging capacity from 24.4 to 14.6 μg BHT
equivalent g-1 DW was observed in AZ-pretreated and heated seedlings with an increase
in the AZ pretreatment concentration (Table 2-1). Nevertheless, the reducing power in
leaves of HT seedlings (19.4 mg BHT equivalent g-1 DW) was significant lower than that
of CK seedlings (21.6 mg BHT equivalent g-1 DW), but the difference in the reducing
power in leaves of seedlings among the AZ-pretreated and heated groups was
insignificant. APX and CAT are involved in the antioxidant system to protect against
stress-induced reactive oxygen species (ROS). Wu and Tiedemann (2001) showed that
AZ induced a delay in senescence which resulted from an increase in superoxide
dismutase activity. Zhang et al. (2010) also suggested that AZ enhanced the activity of
antioxidant enzymes, including CAT, and induced a delay in senescence. In our study, the
activities of APX and CAT in seedling leaves were enhanced after being exposed to HT
for 1 h, but suppression of enzyme activities was observed in AZ-pretreated seedlings.
Furthermore, AZ also reduced the DPPH radical scavenging capacity in leaves of
seedlings after HT exposure, suggesting that AZ was unable to provide a protective effect
against HT in seedlings, but in fact damaged the antioxidant system in seedling leaves.
These results are consistent with a previous study by Debona and Rodrigues (2016) who
observed that AZ suppressed stress induced activities of APX and CAT in rice leaves.
Moreover, Amaro et al. (2018) also reported that strobilurins, with the exception of AZ,
improved the activity of the antioxidative system.
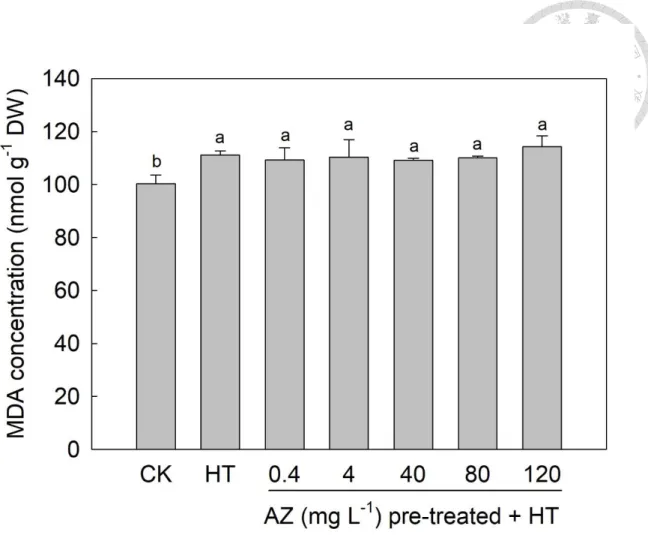
Malondialdehyde (MDA) concentration
In plants, the concentration of MDA, a product of lipid peroxidation, reflects the status
of heat-induced damage (Lu et al., 2009; Bhardwaj and Ramandeep, 2017). Fig. 2-5
shows that the level of MDA in leaves of HT seedlings (111 nmol g-1 DW) was higher
than that of CK seedlings (100 nmol g-1 DW). Meanwhile, a stable level (109~114 nmol
g-1 DW) of MDA in leaves of heated seedlings was observed among all AZ pretreatment
concentrations. Zhang et al. (2010) observed that a reduction in MDA was accompanied
by enhanced antioxidant enzyme activity. In our study, HT resulted in higher MDA levels
in seedling leaves, but AZ neither enhanced nor suppressed the level of MDA in leaves
of seedlings after exposure to HT. In addition, a similar trend was observed in the
reducing power in leaves of seedlings after HT treatment. Debona and Rodrigues (2016)
reported that glutathione, a chemical involved in the non-enzymatic system, inhibited
oxidative stress. There might be another antioxidant mechanism, which was undermined
in this study, in the AZ-treated seedlings against oxidant stress.
Photosynthetic pigments
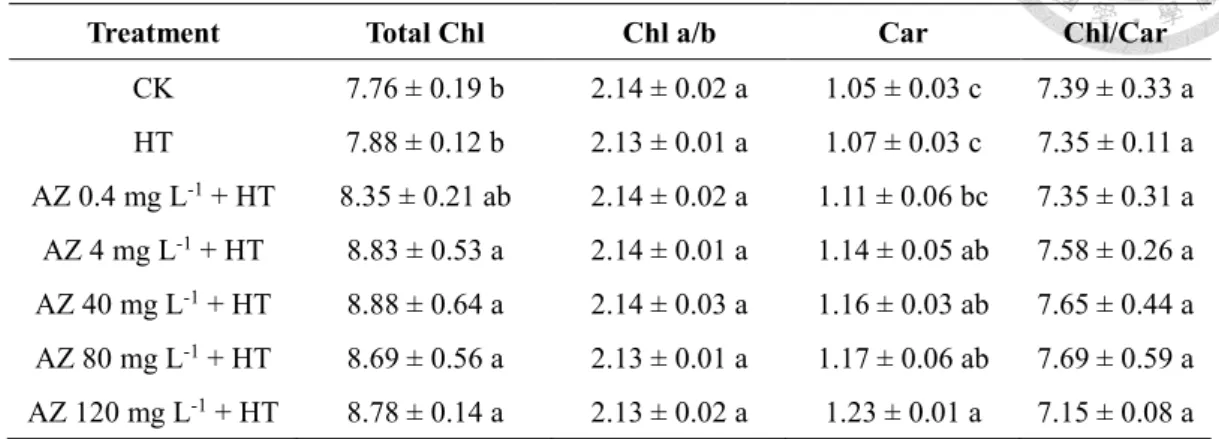
Both Chl and Cars are involved in the light reaction of photosynthesis. Levels of total
Chl (7.88 mg g-1 DW), which is the sum of Chl a and Chl b concentrations, and Cars (1.07
mg g-1 DW) in leaves of seedlings exposed to HT were consistent with those of CK
seedlings. In addition, total Chl and Car concentrations in leaves of AZ-pretreated and
heated seedlings exhibited a significant upward trend from 7.88 to 8.78 mg g-1 DW and
from 1.07 to 1.23 mg g-1 DW, respectively, with an increase in the AZ concentration
applied (Table 2-2). However, differences in Chl a/b ratios in leaves were insignificant
among all experimental treatments. Biotic and/or abiotic stresses usually lead to
reductions in the concentration of photosynthetic pigments (Ashraf and Harris, 2013;
Chen et al., 2016). The lower Chl concentration in leaves might result from an imbalance
between the biosynthetic and degradative pathways of Chl (Chen et al., 2015), but levels
of photosynthetic pigments in leaves of HT seedlings were consistent with those of CK
seedlings. Short-term exposure to HT probably did not effectively induce sharp
reductions in pigment levels in leaves. On the other hand, AZ increased the accumulation
of photosynthetic pigments in seedling leaves. A similar result was also presented in
previous studies (Grossmann and Retzlaff, 1997; Wu and Tiedemann, 2001), and the
phenomena, such as a reduction in Chl degradation or a delay in leaf yellowing, is called
a ‘greening effect’. Our results showed that even though AZ induced a greening effect, it
was obviously unable to protect against oxidant stress which was also caused by AZ. Song
et al. (2013) reported that an enhanced Chl/Car ratio could mitigate heat-induced
oxidative stress. In our study, a stable ratio of Chl/Car was observed in AZ-treated
seedling leaves, and this might be another reason that AZ was unable to provide a
protective effect against heat stress in wheat seedlings. Strobilurin is one of the most
important fungicides for plant disease control. In addition, strobilurin is also considered
a chemical to improve crop physiology. The effect of strobilurins on wheat grown under
a well-controlled environment without stress and applied during the later growth stages
induced a delay in senescence and promoted grain yields (Wu and Tiedemann, 2001;
Zhang et al., 2010). Nevertheless, each strobilurin produced a dynamic effect on the
plant’s physiology and growth (Amaro et al., 2018), and the responses of crops to
strobilurin are dramatically diverse at different growth stages (Zhang et al., 2010), and
some specific physiological effects are only fully presented with sufficient N fertilizer
(Ishikawa et al., 2012). A previous study reported that strobilurins alleviated paraquat-
induced stress (Wu and Tiedemann, 2001), but reduced photosynthesis during a drought
(Nason et al., 2007). The heat-induced oxidant stress decreased the Chl concentration and
CAT activity, increased the MDA concentration, and ultimately reduced wheat yields
(Zhao et al., 2007). In our study, the effect of AZ on the wheat seedlings exposed to
sudden HT was also observed. Further studies on the protective effect of strobilurins in
crops are needed.
Conclusions
This study explored the physiological mechanism of AZ in wheat seedlings exposed
to sudden HT. AZ treatment displayed an ability to increase the concentrations of
photosynthetic pigments in seedling leaves of wheat, but impacted ChlF, APX and CAT
activities. There were no trends observed in the responses of the reducing power or MDA
concentration in leaves to AZ concentrations. Therefore, AZ provided limited support for
the physiological functioning of wheat seedlings under HT stress.
References
Amaro ACE, Ramos ARP, Macedo AC, Ono EO, Rodrigues JD (2018). Effects of the
fungicides azoxystrobin, pyraclostrobin and boscalid on the physiology of Japanese
cucumber. Scientia Horticulturae, 228:66-75.
Ashraf M, Harris P (2013). Photosynthesis under stressful environments: an overview.
Photosynthetica, 51(2):163-190.
Asseng S, Ewert F, Martre P, Rotter RP, Lobell DB, Cammarano D, Zhu Y (2014). Rising
temperatures reduce global wheat production. Nature Climate Change, 5(2):143.
Bhardwaj RD, Ramandeep K (2017). Protective role of pre-treatment with different
phenolic acids in wheat seedlings against heat stress induced oxidative damage.
Indian Journal of Agricultural Biochemistry, 30(2):147-155.
Cantore V, Lechkar O, Karabulut E, Sellami MH, Albrizio R, Boari F, Todorovic M
(2016). Combined effect of deficit irrigation and strobilurin application on yield,
fruit quality and water use efficiency of“cherry” tomato (Solanum lycopersicum L.).
Agricultural Water Management, 167:53-61.
Chen YE, Cui JM, Su YQ, Yuan S, Yuan M, Zhang HY (2015). Influence of stripe rust
infection on the photosynthetic characteristics and antioxidant system of susceptible
and resistant wheat cultivars at the adult plant stage. Frontiers in Plant Science, 6:779.
Chen YE, Liu WJ, Su YQ, Cui JM, Zhang ZW, Yuan M, Yuan S (2016). Different response
of photosystem II to short and long-term drought stress in Arabidopsis thaliana.
Physiologia Plantarum, 158(2):225-235.
Choi CW, Kim SC, Hwang SS, Choi BK, Ahn HJ, Lee MY, Park SH, Kim SK (2002).
Antioxidant activity and free radical scavenging capacity between Korean medicinal
plants and flavonoids by assay-guided comparison. Plant science, 163: 1161–1168.
Debona D, Nascimento KJT, Gomes JGO, Aucique-Perez CE, Rodrigues FA (2016).
Physiological changes promoted by a strobilurin fungicide in the rice-Bipolaris
oryzae interaction. Pesticide Biochemistry and Physiology, 130:8-16.
Debona D, Rodrigues FA (2016). A strobilurin fungicide relieves Bipolaris oryzae
induced oxidative stress in rice. Journal of Phytopathology, 164(9):571-581.
Giuliani MM, Nardella E, Gatta G, Caro AD, Quitadamo M (2011). Processing tomato
cultivated under water deficit conditions: the effect of azoxystrobin. Acta
Horticulturae, 914:287-294.
Gondim FA, Gomes-Filho E, Costa JH, Alencar NLM, Prisco JT (2012). Catalase plays
a key role in salt stress acclimation induced by hydrogen peroxide pretreatment in
maize. Plant Physiology Biochemistry, 56:62-71.
Grossmann K, Retzlaff G (1997). Bioregulatory effects of the fungicidal strobilurin
kresoxim-methyl in wheat (Triticum aestivum). Pesticide Science, 50(1):11-20.
Heath RL, Packer L (1968). Photoperoxidation in isolated chloroplasts: I. Kinetics and
stoichiometry of fatty acid peroxidation. Archives of Biochemistry and Biophysics,
125(1):189-198.
Hoagland DR, Arnon DI (1950). The water-culture for growing plants without soil.
California Agricultural Experiment Station, pp 347.
Ishikawa S, Hare MC, Kettlewell PS (2012). Effects of strobilurin fungicide programmes
and fertilizer nitrogen rates on winter wheat: leaf area, dry matter yield and nitrogen
yield. The Journal of Agricultural Science, 150(4):427-441.
Kato M, Shimizu S (1987). Chlorophyll metabolism in higher plants. VII. Chlorophyll
degradation in senescing tobacco leaves; phenolic dependent peroxidative
degradation. Canadian Journal of Botany, 65(4):729-735.
Kramer DM, Johnson G, Kiirats O, Edwards GE (2004). New fluorescence parameters
for the determination of QA redox state and excitation energy fluxes. Photosynthesis
Research, 79(2):209.
Lu J, Zhang R, Zong X, Wang S, He G (2009). Effect of salicylic acid on heat resistance
of rice seedling under heat stress. Chinese Journal of Eco-Agriculture, 17(6):1168
1171.
Nakano Y, Asada K (1981). Hydrogen peroxide is scavenged by ascorbatespecific
peroxidase in spinach chloroplasts. Plant and Cell Physiology, 22(5):867-880.
Nason MA, Bartlett D, Farrar J (2007). Strobilurin fungicides induce changes in
photosynthetic gas exchange that do not improve water use efficiency of plants
grown under conditions of water stress. Pest Management Science, 63(12):1191-
1200.
Oyaizu M (1986). Studies on products of browning reaction. Antioxidative activities of
products of browning reaction prepared from glucosamine. Japanese Journal of
Nutrition, 44(6):307-315.
Pedersen MA (2016). Strobilurin fungicide secondary plant stress alleviation effects. PhD
Thesis, University of Nebraska.
Rademacher W (2004). Recent situation and trends in global plant bioregulator utilization.
Regulation of Plant Growth and Development, 39(1):142-151.
Shimada K, Fujikawa K, Yahara K, Nakamura T (1992). Antioxidative properties of
xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. Journal of
Agricultural and Food Chemistry, 40(6):945-948.
Song L, Yue L, Zhao H, Hou M (2013). Protection effect of nitric oxide on photosynthesis
in rice under heat stress. Acta Physiologiae Plantarum, 35(12):3323-3333.
Sun Y, Xu W, Fan A (2006). Effects of salicylic acid on chlorophyll fluorescence and
xanthophyll cycle in cucumber leaves under high temperature and strong light. The
Journal of Applied Ecology, 17(3):399-402.
Swoboda C, Pedersen P (2009). Effect of fungicide on soybean growth and yield.
Agronomy Journal, 101(2):352-356.
Teixeira EI, Fischer G, Van Velthuizen H, Walter C, Ewert F (2013). Global hot-spots of
heat stress on agricultural crops due to climate change. Agricultural and Forest
Meteorology, 170:206-215.
Vankooten O, Snel JFH (1990). The use of chlorophyll fluorescence nomenclature in
plant stress physiology. Photosynthesis Research, 25(3):147-150.
Venancio WS, Rodrigues MaT, Begliomini E, De Souza NL (2003). Physiological effects
of strobilurin fungicides on plants. Agrariase Engenharias, 9(3):59-68.
Wu Y, von Tiedemann AV (2001). Physiological effects of azoxystrobin and
epoxiconazole on senescence and the oxidative status of wheat. Pesticide
Biochemistry and Physiology, 71(1):1-10.
Yang CM, Chang KW, Yin MH, Huang HM (1998). Methods for the determination of the
chlorophylls and their derivatives. Taiwania, 43(2):116-122.
Zadoks JC, Chang TT, Konzak CF (1974). A decimal code for the growth stages of cereals.
Weed Research, 14(6):415-421.
Zhang YJ, Zhou MG, Wang HC, Zhang X, Chen CJ (2010). Effects of fungicides JS399-
19, azoxystrobin, tebuconazloe, and carbendazim on the physiological and
biochemical indices and grain yield of winter wheat. Pesticide Biochemistry and
Physiology, 98(2):151-157.
Zhao H, Dai T, Jing Q, Jiang D, Cao W (2007). Leaf senescence and grain filling affected
by post-anthesis high temperatures in two different wheat cultivars. Plant Growth
Regulation, 51(2):149-158.