行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※
※
※
Dipyr idamole 抑制人類腹膜表面細胞增生之作用機轉
※
※
※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:■個別型計畫 □整合型計畫
計畫編號:NSC89-2314-B-002-059-
執行期間:88 年 8 月 1 日至 89 年 7 月 31 日
計畫主持人:蔡敦仁
共同主持人:洪冠予
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立臺台灣大學醫學院內科
中 華 民 國 89 年 10 月 20 日
行政院國家科學委員會專題研究計畫成果報告
計畫編號 : NSC89-2314-B-002-059 執行期限:88 年 8 月 1 日 至 89 年 7 月 31 日 主持人:蔡敦仁 執行機構及單位名稱: 國立臺台灣大學醫學院內科 共同主持人:洪冠予 執行機構及單位名稱: 國立臺台灣大學醫學院內科 一. 中文摘要 腹膜硬化症(PF)是腹膜透析(CAPD)病人長期嚴重的合併症之一。PF 的成因目前認 為和人類腹膜表面細胞(HPMC)過度增生伴隨纖維蛋白合成增加有關。然而,HPMC 增 生的調控機轉從未有人報告過。吾人曾經報告 dipyridamole 具有抑制細胞增生及預防纖 維化的作用。因此,本計畫擬進一步探究 dipyridamole 調控 MPMC 增生之效果及其可 能機轉。 自手術取得的正常腹膜大網分離出 HPMC 進行培養。採用 MTT(methyltetra zolium assay)方法測量 HPMC 增生情形,並利用 ELISA 方式測量細胞內 cAMP 濃度。細胞週期 的分佈及成份蛋白的表現是採用流式細胞儀及西方點墨法完成。 結果顯示:dipyridamole 可以抑制 PDGF 刺激 HPMC 增生,使 HPMC 停留在 G1期, 此種抑制增生作用可能主要經由增加細胞內 cAMP 濃度有關。PDGF 使 cyclin D1濃度增 加,對 CDK4 則沒有影響。dipyridamole 不會干擾 PDGF 對上述兩者的作用。PDGF 會 使細胞內 p27kip減少,並促進 pRB 蛋白質活化;但是 dipyridamole 會抑制這兩種作用進 行。 結論:dipyridamole 會抑制 PDGF 刺激 HPMC 增生,此種作用的機轉主要透過增加 細胞內 cAMP,預防 HPMC 細胞內 p27kip1減少,和抑制 pRB 蛋白質活化所致。本計畫 成果可以提供作為 dipyridamole 預防 PF 發生的治療基礎。 (關鍵詞:腹膜透析,纖維化,細胞週期,dipyriamole ) 二. 英文摘要Proliferation of human peritoneal mesothelial cells (HPMC) accompanied by collagen synthesis is regarded as the main process predisposing to peritoneal fibrosis (PF) in patients of long-term continuous ambulatory peritoneal dialysis (CAPD). However, the precise molecular mechanism regulating HPMC proliferation has never been reported. We previously had reported that dipyridamole (DP) is potential as an antiproliferative and antifibrotic agent. We thus investigated the mechanism and effect of dipyridamole in regulation of HPMC proliferation.
HPMC was cultured from human omentum by an enzyme digestion method. Cell proliferation was measured by methyltetrazolium assay. Intracellular cAMP was measured using an enzyme immunoassay kit. Cell-cycle distribution of HPMC was analyzed by flow cytometry. Expressions of cell-cycle proteins (Cyclin D1, CDK4, pRB and p27kip1) were determined by Western blotting.
Addition of DP suppressed PDGF-stimulated HPMC proliferation by cell-cycle arrest at G1 phase. The antimitogenic effect of DP was mediated predominantly through the cAMP
pathway. PDGF induced elevated protein levels of cyclin D1 but the CDK4 protein level did not change. Dipyridamole and DBcAMP had no effect on levels of cyclin D1 and CDK4 in
PDGF-stimulated HPMC. PDGF decreased p27kip1 and induced pRB phosphorylation of HPMC. In contrast, dipyridamole attenuated PDGF-stimulated pRB phosphorylation by preventing p27kip1
Dipyridamole appears to inhibit PDGF-stimulated HPMC proliferation through increased cAMP, preservation of p27kip1
and decreased pRb phosphorylation. Our study of dipyridamole may provide a therapeutic basis for clinical applications in the prevention of PF.
(Keywords: continuous ambulatory peritoneal dialysis, fibrosis, cell-cycle, dipyridamole)
三. 緣由與目的
The successful use of peritoneal dialysis (PD) as a long-term replacement therapy depends on the sustained integrity of the peritoneal membrane. Peritoneal fibrosis (PF), a serious complication found occasionally in patients on long-term PD, has been demonstrated to compromise the peritoneal membrane to a severe degree [1]. Proliferation of human peritoneal mesothelial cells (HPMC) accompanied by collagen synthesis plays a fundamental role in the pathogenesis of PF. However, the precise cellular mechanism regulating HPMC proliferation has, to our knowledge, never been reported.
Platelet-derived growth factor (PDGF) has been reported to be elevated during peritoneal injury [2]. In rat pleural mesothelial cell culture, PDGF has been shown to stimulate PMC proliferation and collagen synthesis [3]. Dipyridamole [2,6-bis(diethanolamino)-4, 8-dipiperidinopyrimido-(5,4-d)-yrimi-dine] is a popular anti-platelet agent [4]. In addition to its anti-platelet activity, we [5] previously have demonstrated that dipyridamole, through increased intracellular cAMP, may inhibit rat mesangial cell proliferation and reduce their expression of collagen α1(I) mRNA. However, its mechanism remains undetermined.
Cell proliferation is governed by the cell-cycle machinery. In mammalian cells, progression through the cell-cycle is dependent on the balance of positive and negative regulatory cell-cycle proteins [6]. Cyclin D1 when bound to its catalytic partner, the cyclin
dependent kinase 4 (CDK4), may result in the formation of active Cyclin D1/CDK4 complex,
which is responsible for the phosphorylation of the retinoblastoma gene product (pRB). Unphosphorylated pRB negatively controls G1 progression, and the phosphorylated form of
pRB is essential and required for proliferation of normal cells. Cyclin-kinase inhibitors (CKI) negatively regulate the cell-cycle by inhibiting the Cyclin D1/CDK4 complex. p27
Kip1
is regarded as a major CKI of G1 phase and is most directly involved in cell cycle restriction
control [7]. Reduction of p27Kip1
expression by PDGF results in cell-cycle re-entry and proliferation of rat mesangial cells. While cyclins and CDKs have been well investigated in a wide range of mammalian cell types, there has been, to our knowledge, no report detailing their induction and regulation in HPMC.
四. 結果與討論
At first, we performed growth curve experiments to evaluate the effect of dipyridamole on PDGF-stimulated HPMC proliferation (Fig. 1). PDGF (25 ng/ml) resulted in a significant increase of HPMC proliferation. This effect was clearly reduced by pretreating the cells with dipyridamole before the addition of PDGF. In time-response experiments, dipyridamole resulted in a dose-dependent inhibition of HPMC. Dipyridamole, at concentrations of 6 µg/ml and 17 µg/ml, inhibited PDGF-stimulated HPMC proliferation by 40.1% and 57.0%, respectively.
To investigate the cellular mechanism of this inhibitory effect of dipyridamole on HPMC growth, cell cycle distribution was determined by flow cytometry (Fig. 2). After starvation, 79.6 ± 4.1% of HPMC were arrested in the G1 phase of the cell cycle. After treated with
PDGF (25 ng/ml) for 24 hours, 14.3 ± 5.2% and 26.1 ± 6.3% of the HPMC were distributed in the S and G2/M phase, respectively. In contrast, 81.5 ±4.5% of the HPMC were still in the G1
phase 24 hours after incubation with PDGF (25 ng/ml) in the presence of dipyridamole (17
plus IBMX (1 mM).
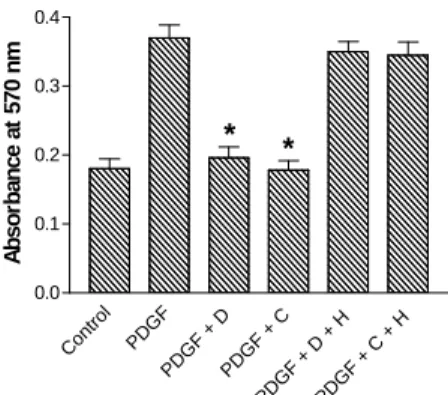
Dipyridamole is known to function as a phosphodiesterase inhibitor to increase intracellular cAMP levels [4]. Thus, we measured cAMP levels after incubation of HPMC with varying concentrations of dipyridamole (2, 6, and 17 µg/ml). In these experiments, IBMX (1 mM), a known phosphodiesterase inhibitor, was used as a positive control (Fig. 3). Intra-cellular cAMP levels were elevated in a dose-dependent manner by dipyridamole. To elucidate if the growth inhibitory actions of dipyridamole was through the increased cAMP, we next investigated the growth inhibitory effect of DBcAMP in cultured HPMC. The addition of DBcAMP (0.5 mM) resulted in an inhibitory effect on PDGF-stimulated HPMC proliferation. H-89, a cell-permeable PKA inhibitor, could overcome the growth inhibitory effects of dipyridamole and DBcAMP (Fig. 4).
As dipyridamole arrests HPMC in G1/S transition, we questioned whether this
inhibitory effect might be due to alterations of cell-cycle proteins. Western blotting analysis (Fig. 5) indicated that there was a low level of protein expression for cyclin D1 in quiescent (0
hr) HPMC. During PDGF stimulation, protein levels of cyclin D1 rose between 0-4 hours, and reached maximal levels around 8-12 hours after stimulation. The CDK4 protein level did not oscillate following the addition of PDGF. Dipyridamole had no effect on protein levels of cyclin D1 or CDK4 in PDGF-stimulated HPMC. However, the addition of dipyridamole
resulted in an attenuated pRB phosphorylation (Fig. 6) and prevents PDGF-stimulated degradation of p27kip1
in HPMC (Fig. 7).
It has been postulated that the relative levels of p27kip1
versus those of cyclinD1/CDK4 are
crucial in determining whether pRB is activated (phosphorylated) or not [6-7]. A role for cAMP in maintenance of p27kip1 by inhibiting its depletion has been suggested [10]. The possible mechanism for blockade of pRB phosphorylation may be through the antagonizing of cyclin D1/CDK4 activity by p27
kip1
. Our data supports the above notion. However, the mechanisms by which dipyridamole prevents the reduction of p27kip1 in PDGF-treated HPMC deserves further study.
In summary, we found in this work that dipyridamole arrests HPMC in G1/S transition by
preventing the depletion of p27kip1 and the modulation on pRB phosphorylation.
1 3 5 0.0 0.1 0.2 0.3 0.4 Control PDGF PDGF + DP2 PDGF + DP6 PDGF + DP17 * * ** ** ** Day A b s o rb a n c e a t 5 7 0 n m ** Control 0 2 6 17 0.0 0.1 0.2 0.3 0.4 0.5 PDGF (25 ng/ml) Dipyridamole (µg/ml) * ** ** A b s o rb a n c e a t 5 7 0 n m
Fig.1. Dipyridamole (DP) inhibits PDGF (25 ng/ml)-stimulated HPMC growth. (A) Time-response curve: Varying concentrations (2, 6, and 17µg/ml) of dipyridamole were added after overnight plating of HPMC and further incubated for 1 to 5 days. (B) Dose-response study: HPMC were incubated for 120 hours after addition of varying concentrations of dipyridamole and a fixed dosage (25 ng/ml) of PDGF. *p<0.05, **p<0.01, compared to PDGF. Each point represents the mean ± SEM of three experiments performed in triplicate.
A
B
(C) PDGF + Dipyr idamole (D) PDGF + IBMX
Fig. 2. Flow cytometry analysis for cell-cycle distribution. Subconfluent HPMC were growth arrested by 0.5% FCS for 48 hours (A), then changed into mediums in the presence of PDGF (25 ng/ml) alone (B), or (C) PDGF plus 17 µg/mL dipyridamole, or (D) PDGF plus IBMX (1 mM). The x axis shows DNA content; the y axis shows the number of cells. The experiments were repeated four times with similar results observed.
Control IBMX 17 6 2 0 5 0 1 00 1 50 2 00 2 50 3 00 * * * Dipyridamole ** c A M P p m o l/ m g p ro te in (µg/ml) Control PDGF PDGF + DPDGF + C PDGF + D + HPDGF + C + H 0.0 0.1 0.2 0.3 0.4 * * A b s o rb a n c e a t 5 7 0 n m
Fig.3. Effect of dipyridamole (unit: µg/ml) on intra-cellular cAMP levels (pmol/mg protein) as measured by a cAMP kit. Intra-cellular cAMP levels were elevated in a dose-dependent manner by dipyridamole. IBMX
(1 mM), a known phosphodiesterase
inhibitor, was used as a positive control. *p<0.01, **p<0.05, relative to control. All experiments were performed four times.
Fig. 4. Effect of dipyridamole(D,17µg/ml) and DBcAMP (C,0.5mM) on PDGF-stimulated HPMC proliferation. MTT assay was performed on the 5th day. H-89 (H, 3 µM) nearly completely reversed the inhibitory effect of dipyridamole or DBcAMP on
PDGF-stimulated HPMC proliferation.
*p<0.01 vs PDGF alone. Data represents the mean ± SEM of four experiments.
pRB-pp pRB p27
(Hr ) 0 4 8 12 24 4
8 12 24 Dipyr idamole - - - - - + + + + PDGF - + + + + + + + +
Fig. 5. Kinetics of expression of cyclin D1 (34KD), CDK4 (32KD), pRB and p27
kip1 proteins upon
stimulation with PDGF (25 ng/ml) and the effect of dipyridamole (DP, 17 µg/ml) upon their expression. Lysates (50 µg protein / lane) of HPMC were analyzed by using specific antibodies. Quiescent HPMC (0 hour) were incubated with PDGF (25 ng/ml) alone or in the presence of dipyridamole (17 µg/ml), and then were harvested at indicated time points for Western blotting analysis. Hyperphosphorylation of pRB slows electrophoretic mobility of the protein, creating the appearance of a double or widened band (gel shift). For evaluation of p27kip1, cellular extracts were electrophoresed on 15% SDS-PAGE then immunoblotted with a
polyclonal p27kip1 antibody. The data is representative of four such studies.
CDK4 Cyclin D1
五. 計畫成果自評
Our work implicated that dipyridamole may serve as a therapeutic agent for prevention or retardation of PF. The inhibitory effect of dipyridamole demonstrated here can provide basis for future studies. An animal in vivo study and possibly a human clinical trail are mandatory.
Both studies can be arranged to examine the role of dipyridamole in the treatment of PF.
六. 參考文獻
1. Dobbie JW, Anderson JD, Hind C. Long-term effects of peritoneal dialysis on peritoneal morphology. Perit Dial Int 14(Suppl 3): S16-S20, 1994
2. Topley N. The cytokine network controlling peritoneal inflammation. Perit Dial Int
15(suppl 7): S35-39, 1995
3. Owens MW, Milligan SA. Growth factor modulation of rat pleural mesothelial cell mitogenesis and collagen synthesis. Effects of epidermal growth factor and platelet-derived factor. Inflammation 18: 77-87, 1994
4. FitzGerald GA. Dipyridamole. N Engl J Med 316: 1247-1257, 1987
5. Tsai TJ, Lin RH, Chang CC, Ko FN, Teng CM. Vasodilator agents modulate rat glomerular mesangial cell growth and collagen synthesis. Nephron 70: 91-100, 1995
6. Sherr CJ. Mammalian G1 cyclins. Cell 73: 1059-1065, 1993
7. Shankland SJ, Pippin J, Flanagan M, Coats SR, Nangaku M, Gordon KL, Roberts JM, Couser WG, Johnson RJ. Mesangial cell proliferation mediated by PDGF and bFGF is determined by levels of the cyclin kinase inhibitor p27Kip1. Kidney Int 51: 1088-1099,
1997
8. Vadiveloo PK, Vairo G, Novak U, Royston AK, Whitty G, Filonzi FL, Cragoe EJ, Hamilton JA. Differential regulation of cell cycle machinery by various antiproliferative agents is linked to macrophage arrest at distinct checkpoints. Oncogene 13: 599-608,
1996
9. Williamson EA, Burgess GS, Eder P, Litz-Jackson S, Boswell HS. Cyclic AMP negatively controls c-myc transcription and G1 cell cycle progression in p210 BCR-ABL transformed cells: inhibitory activity exerted through cyclin D1 and cdk4. Leukemia 11:
73-85, 1997
10. Kato JY, Matsuoka M, Polyak K, Massagur J, Sherr CJ. Cyclic AMP-induced G1 phase arrest mediated by an inhibitor (p27Kip1) of cyclin-dependent kinase 4 activation. Cell 79: