Ferulic acid reverses the cognitive dysfunction caused by amyloid β peptide 1-40
through antioxidant activity and cholinergic activation in rats
Fan-Shiu Tsai†a, Lung-Yuan Wua, Shu-Er Yangb, Hao-Yuan Chengc, Chin-Chuan Tsaia, Chi-Rei Wu*d,
Li-Wei Lin*a
a. School of Chinese Medicines for Post-Baccal aureate, I-Shou University, Kaohsiung 82445, Taiwan b. Department of Beauty Science And Graduate Institute of Beauty Science Technology, Chienkuo Technology University, Changhua 500, Taiwan
c. Department of Nursing, Chung Jen College of Nursing, Health Sciences and Management, Chiayi 600, Taiwan
d. Department of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, College of Pharmacy, China Medical University, Taichung 402, Taiwan
Running title:
Ferulic acid reverses the cognitive dysfunction
Number of pages: 28 Number of figures: 10 Number of tables: 0
Correspondence to: Dr. Li-Wei Lin, School of Chinese Medicines for Post-Baccal aureate, I-Shou University, Kaohsiung 82445, Taiwan. Tel: 886-7-6151100 ext. 7079, E-mail: [email protected]; or Dr. Chi-Rei Wu, Department of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, College of Pharmacy, China Medical University, Taichung 402, Taiwan. E-mail: [email protected] These authors contributed equally to this work.
Abstract
Cholinergic dysfunction and oxidation stress are the dominant mechanism of memory deficit in Alzheimer’s disease (AD). This study describes how ferulic acid (FA) ameliorates cognitive deficits induced by mecamylamine (MECA), scopolamine (SCOP), central acetylcholinergic neurotoxin ethylcholine mustard aziridinium ion (AF64A) and amyloid β peptide (Aβ1-40). This study also
elucidates the role of antioxidant enzymes and cholinergic marker acetylcholinesterase (AChE) in the reversal of FA from Aβ1-40-induced cognitive deficits in rats. At 100 mg/kg, FA attenuated impairment
induced by MECA and SCOP plus MECA; however, this improvement was not blocked by the peripheral muscarinic receptor antagonist scopolamine methylbromide. At 100 and 300 mg/kg, FA also attenuated the impairment of inhibitory passive avoidance induced by AF64A. Further, FA attenuated the performance impairment and memory deficit induced by Aβ1-40 in rats, as did vitamin E/C. FA
reversed the deterioration of superoxide dismutase (SOD) and AChE activities, and the glutathione disulfide (GSSG) and glutathione (GSH) levels in the cortex and hippocampus. Vitamin E/C only selectively reversed deterioration in the hippocampus. We suggest that FA reduced the progression of cognitive deficits by activating central muscarinic and nicotinic receptors and antioxidant enzymes.
Introduction:
Alzheimer’s disease (AD) is characterized by neuropathological hallmarks such as senile plaque and neurofibrillary tangles caused by the amyloid β peptide (Aβ) (Nabeshima and Nitta, 1994; Yamada et
al., 1999). A 39-43 amino acid peptide, Aβ possesses neuronal toxic activity and the potential to cause
neuronal damage in the brain. The deposition of Aβ in the brain, is associated with cerebral neuronal loss, and oxidative stress, particularly degeneration of cholinergic neurons OR neuronal system deficits, oxidative stress, and the degeneration of cholinergic neurons (Nabeshima and Nitta, 1994; McGeer and McGeer, 1999; Tran et al., 2002; Heo, Hyon and Lee, 2013). In studies of intracisternal infusion of Aβ 1-40 into rats indicated that Aβ1-40 mainly deposited in the frontal cortex and hippocampus, causing
learning and memory deficits in the inhibitory avoidance task and Morris water maze (MWM) (Nabeshima and Nitta, 1994; Nitta et al.,1994). The cholinergic neuronal system plays an important role in learning and memory in humans and other animals (Gacar et al.,2011; Sutalangka et al., 2013). Cholinergic receptors were mainly grouped into muscarinic and nicotinic receptors two main subtypes, which also play important roles in learning and memory (Glick and Greenstein, 1972). In fact, some investigators found that patients with AD or dementia lacked the normal number of central muscarinic and nicotinic receptors OR patients with AD or dementia had deficits in the number of central muscarinic and nicotinic receptors OR patients with AD or dementia had few central muscarinic and nicotinic receptors(Glick and Greenstein, 1972). Muscarinic antagonist scopolamine (SCOP) impaired learning and memory in rodents and humans, especially learning acquisition and short-term memory (Tsai et al.,2007; Gacar et al., 2011). The nicotinic antagonist mecamylamine (MECA) also impaired cognitive performance in rodents, such as inhibitory avoidance or MWM performance (Hsieh et al., 1998; Gacar et al., 2011). Ethylcholine mustard aziridinium ion (AF64A), a neurotoxic derivative of choline, significantly decreases hippocampal acetylcholine contents by producing long-term presynaptic cholinergic deficits, impairing learning performance in rats (Sutalangka et al., 2013; Sutalangka et al., 2013). Therefore, SCOP, MECA and AF64A are useful models when screening anti-amnestic drugs.
Ferulic acid (FA), a phenolic compound and derivative of p-hydroxybenzyl alcohol, possesses anti-oxidant, anti-inflammatory and activity and promotes neuritogenesis (Wenk et al., 2004; Mamiya et al., 2008; Lin et al., 2012). Previous studies showed that p-hydroxybenzyl alcohol attenuated the
impairment of inhibitory passive avoidance caused by SCOP or cycloheximide (Wu et al., 1996; Hsieh
et al., 1998). Other researchers found that FA protects against a single intracerebroventricular injection
of Aβ1-42, which induces learning and memory impairment via astrocyte and microglial activation in
mice (Yan et al., 2004; Cho et al., 2005). Therefore, we hypothesize that FA has beneficial effects for AD patients. To address this hypothesis, this study determined whether FA counteracted the impairment of inhibitory passive avoidance caused by such cholinergic antagonists as SCOP and MECA and cholinergic neurotoxin AF64A and investigated the potential ameliorating effects of FA against intracisternal infusion of Aβ1-40 by behavioral measurements such as the inhibitory avoidance task and
MWM, and assessing levels of antioxidant markers and ccetylcholinesterase (AChE) activity in rats. Antioxidants such as α-tocopherol (vitamin E) and ascorbic acid (vitamin C) improved learning and memory deficits induced by Aβ in vitro and in vivo (Heo et al., 2013; Herrera et al., 2013; Dysken et
al., 2014; Kook et al., 2014). For this reason, we also demonstrated the role of cortical and hippocampal
antioxidant enzymes and AChE in the protective effect of FA against continuous intracisternal injection of Aβ1-40 in rats.
Materials and methods
Animals and Housing
Male Sprague-Dawley rats, weighing 250-300 gm, were obtained from the BioLASCO Taiwan Co. Ltd. and were used in experiments according to the Guiding Principles for the Care and Use of Laboratory Animals. They were randomly housed six per wire-mesh cage (39 26 21 cm) for at least 1 week before experiment start. Temperature was 231oC and humidity was 60% in this regulated
environment. Rats had free access to standard food pellets (Fwusow Industry Co. Ltd., Taiwan) and tap water. The light/dark cycle was 12 h (light, 08:00-20:00 h).
Each 10-12 rats were then randomized into groups of MECA, SCOP, M-SCOP, AF64A, Aβ1-40 and
FA treated for the inhibitory avoidance test, just only Aβ1-40 and the following drugs FA and vitamin E/C
were administered for MWM task and then determination of brain antioxidant markers and AChE activity. The following drugs were administered and behavioral tests were operated by double-blind method.
Chemicals
All drugs expcept synthesized human Aβ 1-40 (Aβ1-40) were purchased from the Sigma-Aldrich
Chemical Co. (USA). The FA, with a purity exceeding 98%, was dissolved with 0.5% carboxymethylcellulose. Vitamin E and vitamin C were dissolved with 0.5% Tween 80 at a ratio of 3:1 (Vitamin E/C). The SCOP, MECA, and scopolamine methylbromide (M-SCOP) were dissolved with saline. The AF64A was freshly prepared by dissolving acetylcholine mustard hydrochloride in normal saline according to the technique described by Fisher et al. (1982). The pH was adjusted to 7.4 with NaHCO3 and the solutions were maintained at room temperature for 1 h. The Aβ1-40 was purchased from
Tocris Cookson Ltd. (province, UK) and prepared as a stock solution at 250 pmol/μL in 35% acetonitrile/0.1% trifluoacetic acid. Aliquots were stored at -20 C until use.
Evaluation of cognitive function Inhibitory avoidance test
The apparatus (Coulbourn Instruments L.L.C., USA) consisted of two compartments (26 × 26 × 30 cm) with a grid floor made of steel rods (the 36 parallel steel rods were 0.15 cm in diameter and set 2 cm apart). One light compartment lighted with a 20W lamp 30 cm above the floor, and one dark compartment of the same size were connected by a guillotine door (5 × 5 cm). The room was dark during experiment, which were sessions that were conducted between 09:00 and 12:00 h.
During the training trial, the guillotine door between the light and dark compartments was closed. When each rat was placed in the light compartment with its back to the guillotine door, the door was opened and the step-through latency was measured with a stopwatch until the rat entered the dark compartment. After the rat entered the dark compartment, the door closed. An inescapable scrambled footshock (1 mA for 2 s) was delivered through the grid floor. The rat was removed from the dark compartment 5 s after the shock. The rat was then placed back into its home cage until its retention trial, which was conducted at 24 h after the training trial. During the retention trial, each rat was again placed in the light compartment and, as in the training trial, the guillotine door was opened and step-through latency was recorded and used as a measure of retention (McEntee and Crook, 1991; Hsieh et al., 1998). The upper cut-off time was 300 sec.
Morris water maze task
To evaluate spatial learning and memory function, rats were tested in an MWM The MWM was a black circular pool (diameter, 165 cm; height, 60 cm) filled with water at 23 ± 1 °C to a depth of 35 cm, which were sessions that were conducted between 09:00 and 12:00 h. The maze was divided geographically into four equal quadrants and each quadrant had a release point. Each rat’s position in the pool was recorded by a video camera and an automated video tracking system equipped with EthoVision XT software (Noldus Information Technology, Leesburg, VA, USA). The swim path, escape latency, and swimming speed were recorded during each trial.
To find the hidden platform submerged 1.0 cm below the water surface, each rat was given 4 trials/day for 4 consecutive days. A trial was initiated by placing the rat in the water facing the pool wall in one of the four quadrants. The daily choice of initial quadrant was randomized, such that all four quadrants were used once each day. For each trial, the rat was allowed to swim for a maximum of 120 sec to find the platform. When successful, the rat was allowed a 30-sec rest period on the platform. If unsuccessful, the rat was given a score of 120 sec and then physically placed on the platform and, like a successful rat, given a 30-sec rest period. In both cases, a rat was immediately given its next trial after the rest period. All swimming times during each trial were recorded. Escape latencies were compared between groups. At 4 h after the last spatial learning trial on the four consecutive days, the probe test was administered to measure reference memory. In this probe trial, the platform was removed from the pool to measure “spatial bias”, which was accomplished by measuring the time traveled in the target quadrant.
On the following day, the platform was again placed in the pool in the same position, but raised 1 cm above the water surface. Each rat was also given 4 trials to locate the platform visually. Latency, the time it took to find the platform, was measured as a comparison of visual acuity.
Models of amnesia and dementia in rats Cholinergic antagonists-induced amnesia
FA at 50 and 100 mg/kg was administered to rats treated with these below drugs, the training trial of inhibitory avoidance test was carried out 60 min after the treatment. SCOP (1 mg/kg, i.p.) or MECA (10 mg/kg, i.p.) were given to rats 30 min before the training trial of inhibitory avoidance test respectively
(Gacar et al., 2011). SCOP (0.3 mg/kg, i.p.) plus MECA (3 mg/kg, i.p.) was given to rats 30 min before the training trial of inhibitory avoidance test (Hsieh et al., 2000). SCOP (1 mg/kg, i.p.) plus M-SCOP (0.5 mg/kg, i.p.) was given to rats 30 min before the training trial of inhibitory avoidance test (Tsai et
al., 2007).
Intracisternal injection of cholinergic neurotoxin AF64A
All rats were anesthetized with 45 mg/kg sodium phentobarbital (i.p.), positioned in a Narishige stereotaxic instrument, and a cannula was implanted into the left cerebral ventricle (A 1.5, L 0.9, V 3.6). Cholinergic neurotoxin AF64A was intracisternally administrated at 3 nmol/3 μL/brain, and the training trial of the inhibitory avoidance test was carried out 10 days after the injection (Tsai et al., 2007). The control rats were injected with the vehicle only.
Intracisternal infusion of Aβ with Alzet osmotic pump
An Aβ infusion model in rats was used as an AD-like animal model and followed by Nabeshima (Nabeshima and Nitta, 1994). Again, rats anesthetized with 45 mg/kg sodium phentobarbital (i.p.), were positioned in a Narishige stereotaxic instrument, and a cannula was implanted into the left cerebral ventricle (A 1.5, L 0.9, V 3.6). Continuous infusion of the A1-40 (300 pmol/day) was maintained for at
least 2 weeks by attaching a cannula to a mini-osmotic pump (Alzet 2002; Alza, Palo Alto, CA, USA). The control rats were infused with the vehicle only (35% acetonitrile/0.1% trifluoacetic acid). Throughout the infusion period, 50 or 100 mg/kg FA or 150 mg/kg vitamin E/C was given daily.
Determination of brain antioxidant makers
The brain cortex and hippocampus were separated on ice and rapidly stored in liquid nitrogen until assays. Each cortex and hippocampus was weighed and homogenized in 9 vol 25 mM ice-cold phosphate buffered saline (pH 7.4). The levels of total glutathione (GSH) were determined according to the method by Baker, Cerniglia and Zaman. Briefly, lysates (20 μg/50 μL) and the glutathione standard were pipetted into each well of 96-well plates. Immediately, 0.1 mL of the reaction solution, which included of 660 μM 5,5’-Dithiobis(2-nitrobenzoic acid) (DTNB), 900 μM nicotinamide adenine dinucleotide phosphate (NADPH) and 4.5 U/mL glutathione reductase was added to each well, and then
was recorded kinetically at 405 nm for 5 min by microplate reader. The activity of superoxide dismutase (SOD) was measured spectrophotometrically by monitoring absorbance at 560 nm using nitroblue tetrazolium chloride as a chromogen. The SOD activities were expressed as units/mg protein. To estimate total SOD, 10 μM potassium cyanide (KCN) was added to the medium to inhibit cytochrome oxidase activity. To estimate Mn-SOD activity, 1 mM KCN was added to the incubation mixture to inhibit Cu,Zn-SOD activity (McCord and Fridovich, 1969). The activity of Cu,Zn-SOD was calculated by subtracting Mn-SOD activity from total SOD activity.
Determination of brain AChE activity
Acetylcholinesterase (AChE) activity was measured by the principle of the Ellman method (Ellman et
al., 1961). Brain homogenates were incubated for 150 sec at 37°C with 0.3 mM acetylthiocholine iodide
and 5,5'-dithiobis-(2-nitrobenzoic acid), for color development. Production of the yellow anion of 5-thio-2-nitrobenzoic acid was measured at 412 nm. AChE activity was expressed as U AChE /mg protein.
Statistical Analysis:
The data from the inhibitory avoidance task were analyzed using a Kruskal-Wallis non-parametric one-way analysis of variance (ANOVA), followed by the Mann-Whitney U-test. Statistical significance was p < 0.05 in all statistical evaluations. The data for spatial performance were subjected to one-way ANOVA while two-way ANOVA was used for data from the second experiments. These analyses were followed by a post hoc least significant difference (LSD) test for differences between groups. Statistical significance was p < 0.05. The data for reference memory, working memory and the concentration of cortical and hippocampal GSH and SOD were subjected to one-way ANOVA, followed by Scheffe’s test. Statistical significance was p < 0.05.
Result and Discussion
Evidently, the central cholinergic neuronal system plays an important role in learning and memory processes (Lee et al., 2014; Wattanathorn et al., 2014). Some investigators found that AD patients or dementia patients had a significant loss of central muscarinic and nicotinic receptors (Glick and
Greenstein, 1972). Notably, SCOP is a commonly used muscarinic cholinergic receptor antagonist and MECA is a nicotinic cholinergic receptor antagonist in vivo and in vitro (Fu and Jhamandas, 2003; Gacar et al., 2011). Inhibition of the muscarinic acetylcholine receptor by SCOP also contributes to cognitive and memory deficits that are characteristic of AD (Hsieh et al., 1998; Gacar et al., 2011). This study indicates that 50 and 100 mg/kg FA reversed the step-through latency shortened by SCOP, MECA and SCOP plus MECA (Fig. 1 and 2; p < 0.01, p < 0.001). The role of central or peripheral nervous system in the anti-amnesic effect of FA was then elucidated by combining SCOP with M-SCOP or an intracisternal injection of AF64A. The M-SCOP, a peripherally acting muscarinic receptor antagonist, can reduce the activity of the peripheral cholinergic neuronal system and attenuates the ameliorating effects of nootropics (Peng et al., 2007). In this study, 0.5 mg/kg M-SCOP did not shorten the step-through latency in the retention trial, and did not block FA-induced reversal of impairment of passive avoidance induced by SCOP (Fig. 2; p < 0.05). A neurotoxic derivative of choline, AF64A significantly decreases hippocampal acetylcholine contents by producing long-term presynaptic cholinergic deficits and oxidative stress resulting in impaired learning performance in rats (Thukham-Mee and Wattanathorn, 2012; Sutalangka et al., 2013; Wattanathorn et al., 2014). Experimental data also show that AF64A administered intracisternally caused impairment of inhibitory passive avoidance, and this impairment was then partially reversed by 100 and 300 mg/kg FA (Fig. 3; p < 0.01, p <0.001). From these analytical results, we suggest that FA attenuated impairment of inhibitory passive avoidance, which the effects were related to activating the central not the peripheral cholinergic neuronal system
via muscarinic and nicotinic receptors.
The pathogenic character of AD is defined by many senile plaques resulting from aggregation of Aβ, which caused oxidative stress, apoptosis and neurodegeneration in the brain (Heo et al., 2013; Mori et
al., 2013). Intracisternal injection of Aβ1-40 in rats impaired memory and produced cholinergic dysfunction (Terry and Buccafusco, 2003; Shinagawa and Shigeta, 2014; Wattanathorn et al., 2014). Consisten with those in previous reports(Yamada et al., 1999; Tran et al., 2002), data obtained by this study show that Aβ1-40 intracisternal injection caused behavioral deficits including impaired inhibitory
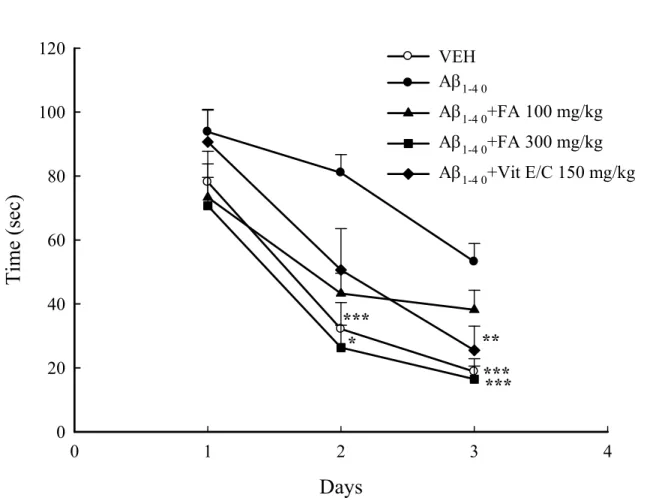
avoidance response and spatial performance in MWM (Fig. 4 and 5; p < 0.001). Additionally, data show that Aβ1-40 administered intracisternally impaired passive avoidance, which was partially reversed by FA
intracisternally impaired spatial performance, and was partially reversed by FA and vitamin E/C (Fig. 5; p < 0.05, p < 0.01, p < 0.001); these analytical results were similar to those for reference memory (Fig. 6; p < 0.01) and working memory (Fig. 7; p < 0.05). However, in velocity there are all non-significantly, show that Aβ or other treatment drugs did not affect the rats. Experimental results for rats are in agreement with those acquired by Yan for mice(Yan et al., 2001; Cho et al., 2005). Additionally, a previous study (Tsai et al., 2010) showed that FA is a potential anti-amnesic and anti-dementia phytochemical.
Although reports shown that FA acts against inflammatory response and memory impairment induced by intracisternal infusion of Aβ1-42 into mice(Yan et al., 2001; Cho et al., 2005), in this study
intracisternally injected of Aβ1-40 into rats and combined anticholinergic agents to identify the protective
effects of FA in Aβ1-40 induced memory deficit in the cholinergic system and oxidative stress. Vitamins
C and E eliminated oxidative stress and AChE activity against Aβ-induced toxicity in vitro and in vivo (McGeer and McGeer, 1999; Heo et al., 2013; Herrera et al., 2013; Dysken et al., 2014; Kook et al., 2014). Intracisternal infusion of Aβ1-40 in rats impaired learning and memory, both of which were
associated with increased of AChE activity (Nabeshima and Nitta, 1994; Nitta et al., 1994). Treating with Aβ1-40 caused cholinergic and other neurotransmitter neuronal systems dysfunction, evidenced by
the reduction in nicotine induced stimulation of acetylcholine in vivo (Fu and Jhamandas, 2003). Yan et
al. demonstrated that intracisternal infusion of Aβ1-42 in mice causes learning and memory deficits that are accompanied by increased immunoreactivities of astrocyte marker glial fibrillary acidic protein (GFAP), interleukin-1β (IL-1β), endothelial nitric oxide synthase (eNOS) and 3-nitrotyrosine (3-NT) in the hippocampus; these effects were also suppressed by pretreatment with FA (Yan et al., 2001; Cho et
al., 2005). A previous study found intracisternal injection of Aβ1-40 induced impairment of MWM spatial performance, reference memory, and passive avoidance behavior in rats that revered by luteolin (Tsai et
al., 2010).
Notably, Aβ induced oxidative stress and inflammation in the brain, which may play important roles in the pathogenesis of AD (McGeer and McGeer, 1999). Aβ in the brain, is associated with cerebral neuronal loss, and activate glial cells generate oxidative stress and proinflammatory cytokines, and particularly degeneration of the cholinergic neuronal system (Nabeshima and Nitta, 1994; Tran et al., 2002; Cho et al., 2005; Heo et al., 2013). Yan et al. determined that FA protects against intracisternal
injection of Aβ1-42 induced learning and memory deficits and microglial activation (Yan et al., 2001;
Cho et al., 2005) in mice. Furthermore, FA also protect against Aβ1-42-induced oxidative stress in vitro
and in vivo (Sultana et al., 2005; Joshi et al., 2006). A previous study demonstrated that continuous intracisternal infusion of Aβ1-40 in rats resulted in a significantly decreased protein expressions of
Mn-SOD, GSH, glutathione disulfide (GSSG) in the rat brain, suggesting that Aβ1-40 impairs antioxidant
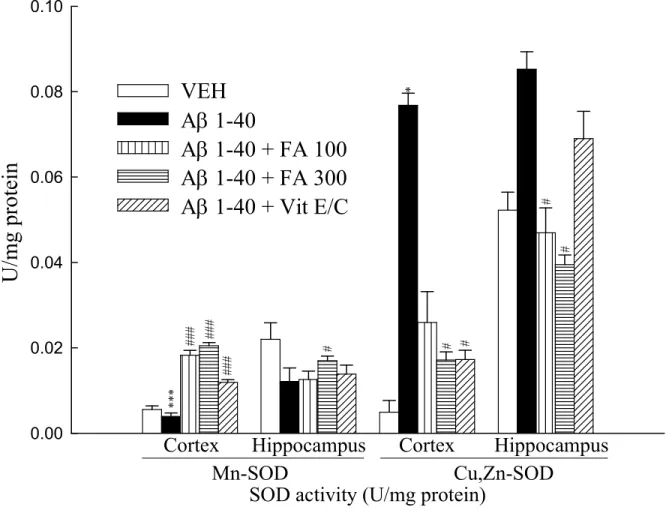
capacity(Tsai et al., 2010). Another study showed that SCOP induced changes in reactive oxygen species (ROS), malondialdehyde (MDA), and AChE activity in mice(Hou et al., 2014; Lee et al., 2014). Data show that intracisternal injection of Aβ1-40 infusion for 14 day decreased Mn-SOD expression and
increased Cu, Zn-SOD activity. Administering FA and vitamin E/C to rats attenuated Aβ1-40-indcuded
Mn-SOD and Cu, Zn-SOD activity in the cortex and hippocampus (Fig. 8; p < 0.05, p < 0.001). The GSH activity result indicates that administering FA and vitamin E/C also increased GSH activity induced by Aβ1-40 in the cortex. In hippocampus, only vitamin E/C reversed GSH activity induced by
Aβ1-40 (Fig. 9; p < 0.05, p < 0.001). These experimental data show that intracisternal infusion of Aβ1-40
caused oxidative stress in the cortex and hippocampus. Furthermore, in accord with the finding by Yan
et al. study for mice (Yan et al., 2001; Cho et al., 2005) and that in our previous study(Tsai et al., 2010),
FA ameliorated oxidative stress induced by Aβ.
Lesions of cholinergic neurons that decrease ACh released into the synaptic cleft results in learning and memory dysfunction (Shinagawa and Shigeta, 2014). The duration of ACh action is dependent upon the activity of AChE, which hydrolyzes ACh after its released (Shinagawa and Shigeta, 2014). Inhibition of AChE is a therapeutic approach for AD and other types of dementia (Shinagawa and Shigeta, 2014; Wattanathorn et al., 2014). Vitamin C and E eliminated oxidative stress and AChE activity against Aβ-induced toxicity in vitro and in vivo(Heo et al., 2013; Herrera et al., 2013; Dysken
et al., 2014; Kook et al., 2014). Evidently, the central cholinergic neuronal system plays an important
role in learning and memory processes (Lee et al., 2014; Wattanathorn et al., 2014). These findings are in accordance with data for AChE activity (Fig. 10) and behavior results obtained by this study; that is, FA and vitamin E/C reversed the AChE activity in the cortex and hippocampus after intracisternal injection of Aβ1-40 ( p < 0.05, p < 0.001). Therefore, we suggest that the neuroprotective effects of 300
including an increase in cortical and hippocampus acetylcholine levels and a decrease in acetylcholine utility via inhibition of AChE activity.
In conclusion, FA attenuated the MECA-, SCOP-, M-SCOP- AF64A- and Aβ1-40-induced impairment
of inhibitory passive avoidance and MWM spatial performance, and these effects were related to activating the central, not peripheral cholinergic neuronal system via muscarinic and nicotinic receptors, and the action mechanism may be related to antioxidant levels and inhibiting AChE activity. And the dose of FA difference between MECA-, SCOP-, M-SCOP- AF64A- and Aβ1-40-induced impairment that
may be Aβ1-40-induced impairment via multiple factors like oxidative stress, inflammatory response and
the central cholinergic neuronal system deficit et al., compared with MECA-, SCOP-, M-SCOP-AF64A-induced impairment via the central cholinergic neuronal system deficit in rats. In recent years, there were many studies of learning and memory focus on gene express. Our previoussent study showed that genes MAPT、PSEN2、ABCG1、APLP2 and APBA1 were associated with SCOP and M-SCOP induced impairment of inhibitory passive avoidance and MWM spatial performance (Hsieh et al., 2003). And Stepanichev et al. (2014) found that Aβ induced cholinergic locus genes(Stepanichev et al., 2014). In AF64A induced cognitive dysfunction, Park et al. (2012) found that it was associated with choline acetyltransferase (ChAT) gene (Park et al., 2012). According to this conclusion, we will commence gene affect to found the affected pathways of FA attenuated the MECA-, SCOP-, M-SCOP- AF64A-and Aβ 1-40 -induced impairment of inhibitory passive avoidance AF64A-and MWM spatial performance in the future.
Acknowledgments
This study is supported in part by the Ministry of Science and Technology, Taiwan (NSC 100-2320-B-214 -001) and ISU-103-01-E-02.
References
Cho, J.Y., H.S. Kim, D.H. Kim, J.J. Yan, H.W Suh. and D.K Song. Inhibitory effects of long-term administration of ferulic acid on astrocyte activation induced by intracerebroventricular injection of beta-amyloid peptide (1-42) in mice. Pro. Neuro-Psychoph. 29: 901-907, 2005.
Dysken, M.W., M. Sano, S. Asthana, J.E. Vertrees, M. Pallaki, M. Llorente, S. Love, G.D. Schellenberg, J.R. McCarten, J. Malphurs, S. Prieto, P. Chen, D.J. Loreck, G. Trapp, R.S. Bakshi, J.E. Mintzer, J.L. Heidebrink, A. Vidal-Cardona, L.M. Arroyo, A.R. Cruz, S. Zachariah, N.W. Kowall, M.P. Chopra, S. Craft, S. Thielke, C.L. Turvey, C. Woodman, K.A. Monnell, K. Gordon, J. Tomaska, Y. Segal, P.N. Peduzzi and P.D. Guarino. Effect of vitamin E and memantine on functional decline in Alzheimer disease: the TEAM-AD VA cooperative randomized trial. JAMA . 311: 33-44, 2014.
Fu, W. and J.H. Jhamandas. Beta-amyloid peptide activates non-alpha7 nicotinic acetylcholine receptors in rat basal forebrain neurons. J. Neurophysiol. 90: 3130-3136, 2003.
Gacar, N., O. Mutlu, T. Utkan, I. Komsuoglu Celikyurt, S.S. Gocmez and G. Ulak. Beneficial effects of resveratrol on scopolamine but not mecamylamine induced memory impairment in the passive avoidance and Morris water maze tests in rats. Pharmacol. Biochem. Be. 99: 316-323, 2011.
Glick, S.D. and S. Greenstein. Differential effects of scopolamine and mecamylamine on passive avoidance behavior. Life Sci. Pt 1: Physiology and pharmacology 11: 169-179, 1972.
Heo, J.H., L. Hyon and K.M. Lee. The possible role of antioxidant vitamin C in Alzheimer's disease treatment and prevention. Am. J. Alzheimers Dis. Other Demen. 28: 120-125, 2013.
Herrera, A., J. Pineda and M.T. Antonio. Toxic effects of perinatal arsenic exposure on the brain of developing rats and the beneficial role of natural antioxidants. Environ. Toxicol. Pharmacol. 36: 73-79, 2013.
Hou, X.Q., D.W. Wu, C.X. Zhang, R. Yan, C. Yang, C.P. Rong, L. Zhang, X. Chang, R.Y. Su, S.J. Zhang, W.Q. He, Z. Qu, S. Li, Z.R. Su, Y.B. Chen, Q Wang. and S.H Fang. BushenYizhi formula ameliorates cognition deficits and attenuates oxidative stressrelated neuronal apoptosis in scopolamineinduced senescence in mice. Int. J. Mol. Med. 2014.
Hsieh, M.T., Y.T. Lin, Y.H. Lin and C.R. Wu. Radix Angelica Sinensis extracts ameliorate scopolamine- and cycloheximide-induced amnesia, but not p-chloroamphetamine-induced amnesia in rats. Am. J. Chinese Med. 28: 263-272, 2000.
Hsieh, M.T., C.R. Wu and C.C. Hsieh. Ameliorating effect of p-hydroxybenzyl alcohol on cycloheximide-induced impairment of passive avoidance response in rats: interactions with compounds acting at 5-HT1A and 5-HT2 receptors. Pharmacol. Biochem. Be. 60: 337-343, 1998.
Hsieh, M.T., C.L. Hsieh, L.W. Lin, C.R. Wu and G.S. Huang. Differential gene expression of scopolamine-treated rat hippocampus-application of cDNA microarray technology. Life Sci. 73: 1007-1016, 2003.
Jhoo, J.H., H.C. Kim, T. Nabeshima, K. Yamada, E.J. Shin, W.K. Jhoo, W. Kim, K.S. Kang, S.A. Jo and J.I. Woo. Beta-amyloid (1-42)-induced learning and memory deficits in mice: involvement of oxidative burdens in the hippocampus and cerebral cortex. Behav. Brain Res. 155: 185-196, 2004.
Joshi, G., M. Perluigi, R. Sultana, R. Agrippino, V. Calabrese and D.A. Butterfield. In vivo protection of synaptosomes by ferulic acid ethyl ester (FAEE) from oxidative stress mediated by 2,2-azobis(2-amidino-propane)dihydrochloride (AAPH) or Fe(2+)/H(2)O(2): insight into mechanisms of neuroprotection and relevance to oxidative stress-related neurodegenerative disorders. Neurochem. Int. 48: 318-327, 2006.
Kook, S.Y., K.M. Lee, Y. Kim, M.Y. Cha, S. Kang, S.H. Baik, H. Lee, R. Park and I. Mook-Jung. High-dose of vitamin C supplementation reduces amyloid plaque burden and ameliorates pathological changes in the brain of 5XFAD mice. Cell Death Dis. 5: e1083, 2014.
Lee, J.S., H.G. Kim, J.M. Han, D.W. Kim, M.H. Yi, S.W. Son, Y.A. Kim, J.S. Lee, M.K. Choi and C.G. Son. Ethanol extract of Astragali Radix and Salviae Miltiorrhizae Radix, Myelophil, exerts anti-amnesic effect in a mouse model of scopolamine-induced memory deficits. J. Ethnopharmacol. 153: 782-792, 2014.
Lin, Z., J. Gu, J. Xiu, T. Mi, J. Dong and J.K. Tiwari. Traditional chinese medicine for senile dementia.
Evid-Based Compl. Alt.: eCAM 2012: 692621, 2012.
Mamiya, T., M. Kise and K. Morikawa. Ferulic acid attenuated cognitive deficits and increase in carbonyl proteins induced by buthionine-sulfoximine in mice. Neurosci. Lett. 430: 115-118, 2008. McCord, J.M. and I. Fridovich. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 244: 6049-6055, 1969.
McEntee, W.J. and T.H. Crook. Serotonin, memory, and the aging brain. Psychopharmacology. 103: 143-149, 1991.
McGeer, P.L. and E.G. McGeer. Inflammation of the brain in Alzheimer's disease: implications for therapy. J. leukocyte Biol. 65: 409-415, 1999.
Mori, T., N. Koyama, M.V. Guillot-Sestier, J. Tan and T. Town. Ferulic acid is a nutraceutical beta-secretase modulator that improves behavioral impairment and alzheimer-like pathology in transgenic mice. PloS one 8: e55774, 2013.
Nabeshima, T. and A. Nitta. Memory impairment and neuronal dysfunction induced by beta-amyloid protein in rats. Tohoku J. Exp. Med. 174: 241-249, 1994.
Nitta, A., A. Itoh, T. Hasegawa and T. Nabeshima. beta-Amyloid protein-induced Alzheimer's disease animal model. Neurosci. Lett. 170: 63-66, 1994.
Park, D., H.J. Lee, S.S. Joo, D.K. Bae, G. Yang, Y.H. Yang, I. Lim, A. Matsuo, I. Tooyama, Y.B. Kim and S.U. Kim. Human neural stem cells over-expressing choline acetyltransferase restore cognition in rat model of cognitive dysfunction. Exp. Neurol. 234: 521-526, 2012.
Peng, W.H., M.T. Hsieh and C.R. Wu. Effect of long-term administration of berberine on scopolamine-induced amnesia in rats. Jpn. J. Pharmacol. 74: 261-266, 1997.
Shinagawa, S. and M. Shigeta. [Acetylcholinesterase inhibitors for treatment of Alzheimer's disease].
Brain and nerve = Shinkei kenkyu no shinpo 66: 507-516, 2014.
Stepanichev, M., N. Lazareva, G. Tukhbatova, S. Salozhin and N. Gulyaeva. Transient disturbances in contextual fear memory induced by Abeta(25-35) in rats are accompanied by cholinergic dysfunction.
Behav. Brain Res. 259: 152-157, 2014.
Sultana, R., A. Ravagna, H. Mohmmad-Abdul, V. Calabrese and D.A. Butterfield. Ferulic acid ethyl ester protects neurons against amyloid beta- peptide(1-42)-induced oxidative stress and neurotoxicity: relationship to antioxidant activity. J. Neurochem. 92: 749-758, 2005.
Sutalangka, C., J. Wattanathorn, S. Muchimapura and W. Thukham-mee. Moringa oleifera mitigates memory impairment and neurodegeneration in animal model of age-related dementia. Oxid. Med. Cell.
Longev. 2013: 695936, 2013.
Sutalangka, C., J. Wattanathorn, S. Muchimapura, W. Thukham-Mee, P. Wannanon and T. Tong-un. Laser acupuncture improves memory impairment in an animal model of Alzheimer's disease. J.
Acupunct. Meridian Stud. 6: 247-251, 2013.
Terry, A.V., Jr. and J.J. Buccafusco. The cholinergic hypothesis of age and Alzheimer's disease-related cognitive deficits: recent challenges and their implications for novel drug development. J. Pharmacol.
Exp. Ther. 306: 821-827, 2003.
Thukham-Mee, W. and J. Wattanathorn. Evaluation of Safety and Protective Effect of Combined Extract of Cissampelos pareira and Anethum graveolens (PM52) against Age-Related Cognitive Impairment. Evid. Based Complementary Altern. Med.: eCAM 2012: 674101, 2012.
Tran, M.H., K. Yamada and T. Nabeshima. Amyloid beta-peptide induces cholinergic dysfunction and cognitive deficits: a minireview. Peptides. 23: 1271-1283, 2002.
Tsai, F.S., H.Y. Cheng, M.T. Hsieh, C.R. Wu, Y.C. Lin and W.H. Peng. The ameliorating effects of luteolin on beta-amyloid-induced impairment of water maze performance and passive avoidance in rats.
Am. J. Chinese Med. 38: 279-291, 2010.
Tsai, F.S., W.H. Peng, W.H. Wang, C.R. Wu, C.C. Hsieh, Y.T. Lin, I.C. Feng and M.T. Hsieh. Effects of luteolin on learning acquisition in rats: involvement of the central cholinergic system. Life Sci. 80: 1692-1698, 2007.
Wattanathorn, J., S. Muchimapura, W. Thukham-Mee, K. Ingkaninan and S. Wittaya-Areekul. Mangifera indica fruit extract improves memory impairment, cholinergic dysfunction, and oxidative stress damage in animal model of mild cognitive impairment. Oxid. Med. Cell. Longev. 2014: 132097, 2014.
Wenk, G.L., K. McGann-Gramling, B. Hauss-Wegrzyniak, D. Ronchetti, R. Maucci, S. Rosi, L. Gasparini and E. Ongini. Attenuation of chronic neuroinflammation by a nitric oxide-releasing derivative of the antioxidant ferulic acid. J. Neurochem. 89: 484-493, 2004.
Wu, C.R., M.T. Hsieh, S.C. Huang, W.H. Peng, Y.S. Chang and C.F. Chen. Effects of Gastrodia elata and its active constituents on scopolamine-induced amnesia in rats. Planta Med. 62: 317-321, 1996. Yamada, K., T. Tanaka, T. Mamiya, T. Shiotani, T. Kameyama and T. Nabeshima. Improvement by nefiracetam of beta-amyloid-(1-42)-induced learning and memory impairments in rats. Brit. J.
Pharmacol. 126: 235-244, 1999.
Yan, J.J., J.Y. Cho, H.S. Kim, K.L. Kim, J.S. Jung, S.O. Huh, H.W. Suh, Y.H. Kim and D.K. Song. Protection against beta-amyloid peptide toxicity in vivo with long-term administration of ferulic acid.
Figure Legend
Figure 1. Effects of ferulic acid (FA) on the impairment of inhibitory avoidance performance induced by mecamylamine (MECA, 10 mg/kg) or scopolamine (SCOP, 0.3 mg/kg) plus MECA (3 mg/kg) in rats. Data are expressed as mean ± SEM (n = 8-14). ** p < 0.01, *** p < 0.001 compared with MECA/VEH group. ## p < 0.01 compared with SCOP plus MECA group.
Figure 2. Effects of scopolamine methylbromide (M-SCOP, 0.5 mg/kg) on ferulic acid (FA, 100 mg/kg)-induced reversal from the scopolamine (SCOP, 1 mg/kg)-induced impairment of inhibitory avoidance performance in rats. Data are expressed as mean ± SEM (n = 12-14). *** p < 0.001 compared with SCOP/VEH group.
Figure 3. Effects of ferulic acid (FA, 50 and 100 mg/kg) on the impairment of inhibitory avoidance performance induced by acetylcholine mustard hydrobromide (AF64A, 3 nmol/3 l/ brain) in rats. Data are expressed as mean ± SEM (n = 10-12). *** p < 0.001 compared with AF64A group.
Figure 4. Effects of ferulic acid (FA, 100 and 300 mg/kg) or vitamin C/E (Vit E/C, 150 mg/kg) on the impairment of inhibitory avoidance performance induced by intracisternal infusion with amyloid β peptide 1-40 (Aβ1-40) in rats. Data are expressed as mean ± SEM (n = 10-12). * p < 0.05, ** p < 0.01, p <
0.001 compared with Aβ1-40 group.
Figure 5. Effects of ferulic acid (FA, 100 and 300 mg/kg) or vitamin C/E (Vit E/C, 150 mg/kg) on the spatial performance impairment of water maze induced by intracisternal infusion with amyloid β peptide 1-40 (Aβ1-40) in rats. Data are expressed as mean ± SEM (n = 10-12). * p < 0.05, ** p < 0.01, *** p <
0.001 compared with Aβ1-40 group.
Figure 6. Effects of ferulic acid (FA, 100 and 300 mg/kg) or vitamin C/E (Vit E/C, 150 mg/kg) on the deficit of reference memory induced by intracisternal infusion with amyloid β peptide 1-40 (Aβ1-40) in
rats. Data are expressed as mean ± SEM (n = 10-12). ** p < 0.01 compared with Aβ1-40 group.
Figure 7. Effect of ferulic acid (FA, 100 and 300 mg/kg) or vitamin E/C (Vit E/C. 150 mg/kg) on the non-spatial performance (A) and velocity (B) in Aβ 1-40 (300 pmol/day)-infused rats. Data are
expressed as mean ± SEM (n = 10-16). p < 0.05 compared with Sham group. # p < 0.05, compared with Aβ 1-40 group.
Figure 9. Effect of ferulic acid (FA, 100 and 300 mg/kg) or vitamin E/C (Vit E/C, 150 mg/kg) on the levels of cortical and hippocampal GSH altered by intracisternal infusion with Aβ 1-40 infusion (300 pmol/day) in rats. Data are expressed as mean ± SEM (n = 10-16). ** p < 0.01 compared with VEH group. # p < 0.05, ## p < 0.01 compared with Aβ 1-40 group.
Figure 10. Effect of ferulic acid (FA, 100 and 300 mg/kg) or vitamin E/C (Vit E/C, 150 mg/kg) on the activities of cortical and hippocampal AChE altered by intracisternal infusion with Aβ 1-40 infusion (300 pmol/day) in rats. Data are expressed as mean ± SEM (n = 10-16). * p < 0.05, *** p < 0.001 compared with VEH group. # p < 0.05, ### p < 0.001 compared with Aβ 1-40 group.
Figure 1.
S
te
p-th
ro
ug
h
la
te
nc
y
(s
ec
)
0 50 100 150 200 250 300 ##VEH VEH 50 100 VEH 100
FA (mg/kg)
MECA 10 mg/kg
*** **FA
SCOP + MECA
***Figure 2.
S
te
p-th
ro
ug
h
la
te
nc
y
(s
ec
)
0
50
100
150
200
250
300
***
***
VEH M-SCOP VEH FA FA + M-SCOP
SCOP 1 mg/kg
Fig. 3.
S
te
p-th
ro
ug
h
la
te
nc
y
(s
ec
)
0 50 100 150 200 250 300 *** ***VEH VEH 50 100
FA (mg/kg)
AF64A 3 nmol/l/brain
***Fig. 4.
S
te
p
th
ro
ug
h
la
te
nc
y
(s
ec
)
0
50
100
150
200
250
300
**
*
Sham VEH 100 300
FA (mg/kg)
A
1-40Vit E/C
***
Fig. 5.
Days
0 1 2 3 4T
im
e
(s
ec
)
0 20 40 60 80 100 120 VEH A1-4 0 A1-4 0+FA 100 mg/kg A1-4 0+FA 300 mg/kgA1-4 0+Vit E/C 150 mg/kg
*
*** ** ***
Fig. 6.
T
im
e
sp
en
t i
n
th
e
ta
rg
et
a
re
a
(s
ec
)
0
3
6
9
12
15
18
Sham VEH 100 300
FA (mg/kg)
A
1-40Vit E/C
**
**
**
Fig. 7.
(A) non-spatial performance
T im e (s ec ) 0 20 40 60 # # Sham VEH 100 300 FA (mg/kg) A1-40 Vit E/C * (B) velocity V el oc ity ( cm /s ec ) 0 10 20 30 40 50 Sham VEH 100 300 FA (mg/kg) A1-40 Vit E/C
Fig. 8.
U
/m
g
pr
ot
ei
n
0.00 0.02 0.04 0.06 0.08 0.10VEH
A 1-40
A 1-40 + FA 100
A 1-40 + FA 300
A 1-40 + Vit E/C
Cortex
Hippocampus
Cortex
Hippocampus
Mn-SOD
Cu,Zn-SOD
** * ## # ### ## # # # # * # #Fig. 9
T
he
c
on
ce
nt
ra
tio
n
of
G
S
H
(
ng
/m
g
pr
ot
ei
n)
0 1 2 3VEH
A 1-40
A 1-40 + FA 100
A 1-40 + FA 300
A 1-40 + Vit E/C
** # ## # #
Cortex
Hippocampus
** #GSH (ng/mg protein)
Fig. 10 T he c on ce nt ra tio n of A ch E ( U /m g pr ot ei n) 0.0 0.2 0.4 0.6 0.8 1.0 1.2 VEH A 1-40 A 1-40 + FA 100 A 1-40 + FA 300 A 1-40 + Vit E/C
* ** * # ## # # #