252
* To whom correspondence and reprint requests should be addressed. Tel: 886-7-6051355. Fax: 886-7-6051365.
E-mail: [email protected]
S
patial translocations of exotic species have produced solid economic and social benefits globally (Pimentel et al. 2001). However, the inva-sion of exotic organisms due to species introduc-tion also affects native species through predaintroduc-tion, hybridization, parasite or disease transmission, and competition (Vos et al. 1990, Williamson 1996, Marsh and Douglas 1997). Fish invasions have commonly created negative ecological impacts internationally, such as the Nile perch, Latesnuloticus (Linnaeus, 1758), in Lake Victoria (Balirwa et al. 2003) and the brown trout, Salmo trutta Linnaeus, 1758, in New Zealand (Townsend 1996).
Sailfin catfish, Liposarcus multiradiatus (Hancock, 1828), grouped with the armored catfish species and previously named Pterygoplichthys multiradiatus, was originally distributed in the Orinoco River system in South America (FishBase 2004). A nocturnal species, L. multiradiatus
Size Structure, Reproductive Phenology, and Sex Ratio of an Exotic
Armored Catfish (Liposarcus multiradiatus) in the Kaoping River of
Southern Taiwan
Shih-Hsiung Liang1,*, Hsiao-Ping Wu1, and Bao-Sen Shieh2
1Graduate Institute of Life Science, National Kaohsiung Normal University, Kaohsiung, Taiwan 802, R.O.C.
2Department of Biomedical Science and Environmental Biology, Kaohsiung Medical University, Kaohsiung, Taiwan 807, R.O.C.
(Accepted January 22, 2005)
Shih-Hsiung Liang, Hsiao-Ping Wu, and Bao-Sen Shieh (2005) Size structure, reproductive phenology, and sex ratio of an exotic armored catfish (Liposarcus multiradiatus) in the Kaoping River of southern Taiwan.
Zoological Studies 44(2): 252-259. The sailfin catfish, Liposarcus multiradiatus (Hancock, 1828), was originally
introduced into Taiwan commercially for aquarium cleaning, and has now established a wild population in Taiwan. This study investigates the size structure, reproductive phenology, and sex ratio of this exotic species in the Kaoping River of southwestern Taiwan. In total, 537 individuals were respectively collected using gill nets and cast nets in the mainstream and tributaries of the Kaoping River from Dec. 1999 to Nov. 2000. The stan-dard lengths of collected individuals ranged from 12.5 to 43.7 cm, with an average of 28.64 cm. Females were longer than males. Variations in morphological differences between the sexes could not clearly be distin-guished by the 13 body measurements. The female L. multiradiatus is sexually mature at a standard length of 25 cm, and has a reproductive season extending from Mar. to Sept. in Taiwan. Some individuals, however, are able to reproduce from Oct. to Feb. The sex ratio was female-biased (female: male, 2.4: 1), and an even greater proportion of females was found during the reproductive season. Results show that L. multiradiatus exhibits detritus/algae and omnivorous feeding habits, a long reproductive period, a female-biased sex ratio, and possible male parental care. Considering that the sailfin catfish originally inhabits clear, shallow waters, L.
multiradiatus in Taiwan has not only expanded its population size in downstream regions but may also gradually
move to clearer waters upstream or into smaller channels. This species may create serious negative impacts on those native aquatic species such as periphyton feeders and bottom spawners. It is suggested that educat-ing people to stop releaseducat-ing sailfin catfish and provideducat-ing incentives for catcheducat-ing it in reproductive season are two possible ways to control the further spread of this exotic species in Taiwan.
http://www.sinica.edu.tw/zool/zoolstud/44.2/252.pdf
inhabits streams, lakes, and weedy, mud-bot-tomed channels (Page and Burr 1991). Bottom detritus and benthic algae are this species, major food sources, but it also feeds on worms, insect larvae, and various bottom-dwelling aquatic ani-mals (Yamamoto and Tagawa 2000).
Research on armored catfish has centered on their habitat quality and spatial distribution, feeding behavior, respiratory physiology, and life history, but a few researchers have focused on the genus Liposarcus Günther 1864 (Power 1984a 1984b 1990, Fernandes and Perna 1995, Fernandes et al. 1998). In recent years, L. multiradiatus has been introduced worldwide, and has established wild populations in tropical and subtropical regions such as Hawaii (Yamamoto 1992), Florida (Ludlow and Walsh 1991), and Puerto Rico (FAO 1997). In Taiwan, L. multiradiatus was imported commercial-ly for aquarium cleaning during the 1970s, and subsequently established populations in the wild after being released by amateur aquarium owners who had lost interest in ornamental fish (Jiang 1996). This exotic species has become widely distributed in streams, lakes, and reservoirs around the island, especially in the central and southern areas, and has become a serious threat to the freshwater fish diversity of Taiwan (Chen et al. 2003). To effectively manage this exotic species in the future, it is essential to study the population characteristics within the invaded habi-tats of Taiwan. The aim of this study was to inves-tigate the size structure, reproductive phenology, and sex ratio of L. multiradiatus in the Kaoping River, a major river of southwestern Taiwan.
MATERIALS AND METHODS Study site
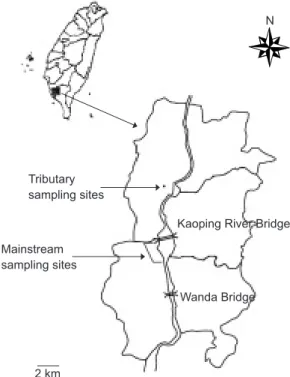
The Kaoping River has the largest drainage area and is the 2nd-longest river in Taiwan. This study was conducted in the downstream portions of the Kaoping River. The stream sections studied are seriously polluted because of the input of agri-cultural and industrial waste waters. Thirty-eight fish species, including 24 primary species and 14 secondary and peripheral species, have been recorded in the studied region (Huang et al. 1994).
Fish sampling
During Dec. 1999~Nov. 2000, fish samplings were conducted on both the mainstream (120
°
25' 31''E,22
°
37'31''N) and its tributaries, including branches and irrigation canals (Fig. 1). To collect fish speci-mens, a gill net (3 m high and 50 m long, with a mesh size of 5 cm) was placed in the water of the mainstream channel overnight for at least 12 h, because the channel is too wide and has strong currents. In the tributaries, a cast net (1 m long with a mesh size of 3.8 cm) was thrown 3 times at each sampling site. Because the minimum stan-dard lengths (LSs) of fish samples in both the mainstream (with a minimum LS of 20 cm) and tributaries (with a minimum LS of 12.2 cm) were much greater than the mesh size used in both stream sections, sources of misinterpretation, such as mesh bias resulting in capturing fish with different LSs, were eliminated. Three stream sec-tions were sampled at least once monthly for each channel type. Before sampling, we expected to catch at least 20 individuals monthly from the mainstream and tributary, respectively, for analysis.Data collection and analysis
Collected individuals were first sacrificed by placing them in a -20
°
C cooler for over 12 h. Males and females were distinguished based on 13 morphometric characteristics, including LS, total weight, body depth, caudal-peduncle depth, predorsal length, predorsal to caudal peduncle,Fig. 1. Sampling locations in the main stream and tributaries of the Kaoping River, Taiwan.
Mainstream sampling sites
Tributary sampling sites
Kaoping River Bridge
Wanda Bridge N
length of anal base, head length, gape width, mouth length, mouth width, eye diameter, and whisker length (Cailliet et al. 1986). After mea-sured these 13 morphometric characteristics, the sex was determined by dissecting the body and directly observing the gonads. Respective correla-tions between LS and body weight for males and females were then calculated. Analysis of covari-ance (ANCOVA) was used to compare the differ-ence of growth models between the sexes.
To morphologically distinguish male and female individuals, t-test was first performed to compare the 13 morphometric measurements between the sexes. Correlation analyses were then conducted for those morphometric variables which showed significant differences between the sexes. If several morphometric variables were sig-nificantly correlated, only one of them was select-ed for further analyses. After these 2 screening processes, a discriminant analysis was executed on those remaining morphometric variables to examine the correctness of the classification between the sexes.
Gonads of sexually mature females are easier to identify and heavier than those of mature males because of enlarged eggs. Therefore, weighing the females, gonad to calculate a gonadosomatic index (GSI) was conducted for determining the reproductive phenology of Hypostomus and L. multiradiatus. (Mazzoni and Caramaschi 1995 1997). The GSI value is expected to increase as a female reaches its reproductive season. The fol-lowing formula was used to calculate the GSI
value:
GSI = [(gonad weight)/(body weight)] x 100. The minimum body length to reach sexual maturity of a female was decided by plotting the GSI value against LS. The reproductive season of females was determined by examining the monthly dynamics of the GSI value. Temporal variations in the sex ratio were also examined by Chi-square test to determine if differences existed between the reproductive and non-reproductive periods. To examine the sex ratio variations of different LS cat-egories, a frequency distribution of the LS was constructed at 3 cm intervals, and Chi-square test was performed for each body-length division.
RESULTS
In total, 537 individuals were collected, but more individuals were collected in tributaries (n = 437, 81.4%) than in the mainstream (n = 100, 18.6%) (Fig. 2). More than 20 individuals per month were collected from the tributaries, but it was not possible to accomplish the same require-ments in the mainstream except in Mar. and July.
More individuals with LSs of 30~35 cm were recorded in both stream sections; specimens with LSs of < 20 cm were collected only from the tribu-taries, while those with LSs of > 40 cm were found only in the mainstream.
Body length and weight
LSs of L. multiradiatus captured from the mainstream ranged from 20.0 to 43.7 ( 30.5 ± 4.8) cm, while LSs of L. multiradiatus from the tribu-taries ranged from 12.2 to 39 (28.3 ± 2.4) cm (Fig. 2). Those individuals collected from the main stream had significantly longer LSs than those from the tributaries (t535= 4.4, p < 0.05).
Body weight and LS were positively correlat-ed for both males and females. Similar growth patterns were found to exist between the sexes, with the exponential formulae between LS and body weight for males and females listed as fol-lows (Fig. 3): male, W = 0.14(LS)2.3 (r2= 0.83, n = 113, p < 0.01); and female, W = 0.19(LS)2.2 (r2= 0.79, n = 275, p < 0.01). Standard Length (cm) <20 20-25 25-30 30-35 35-40 40-45 Number 160 140 120 100 80 60 40 20 0 20 40 60
Fig. 2. Standard length (cm) distribution of collected
Liposarcus multiradiatus from the mainstream and tributaries of
the Kaoping River, Taiwan.
Tributaries Mainstream
No significant difference was found for the growth pattern between sexes based on the results of ANCOVA (p > 0.05). Thus, we pooled data of the two sexes together, and used an expo-nential model to evaluate the growth pattern of L. multiradiatus in the Kaoping River of southwestern Taiwan as follows:
W = 0.08(LS)2.5 (r2= 0.94, n = 388, p < 0.01).
Morphometric classification between sexes
The 388 sex-identified individuals (113 males and 275 females) were used for morphometric classification between the sexes. All of the 13 morphometric measurements except for mouth
width and whisker length of females exhibited sig-nificantly greater values than males (Table 1). In the following analysis, LS was selected to repre-sent body weight, predorsal length, predorsal to caudal peduncle, head width, and gape width, because a significant correlation was found among those variables (r > 0.8, p < 0.05). Six variables without significant correlations, including LS, body height, caudal-peduncle depth, length of anal base, mouth width, and eye diameter, were then selected for sexual classification. Results of the discriminant analysis showed that 61.7% (169/274) of the females and 66.1% (74/112) of the males could be correctly classified based on the selected variables.
Reproductive phenology
During the sampling period, 275 females were collected from both the mainstream and tributaries. The GSI values of females began to increase at a LS of 25 cm (Fig. 4).
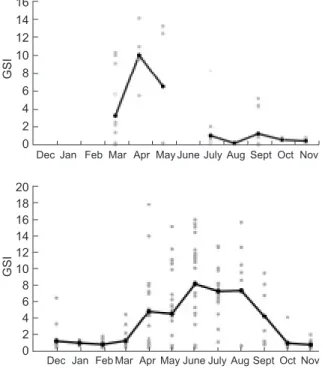
Only 62 females were caught from the main-stream (Fig. 5). No females were collected in June or during the period from Dec. to Feb. Mean GSI values of females from the mainstream showed an increase in Mar. and were close to a flat pattern from July to Nov.
Female Male Standard Length (cm) W eight (g) 1200 1000 800 600 400 200 0 46 43 40 37 34 31 28 25 22 19 16 13 10
Fig. 3. Body weight-standard length correlation of male and female Liposarcus multiradiatus from the Kaoping River, Taiwan. Standard Length (cm) GSI value 20 18 16 14 12 10 8 6 4 2 0 46 43 40 37 34 31 28 25 22 19 16 13 10
Fig. 4. Standard length plotted against the gonadosomatic index (GSI) for collected female Liposarcus multiradiatus from the Kaoping River, Taiwan.
Table 1. Means and standard error (SE) of 13
body measurements of male and female Liposarcus multiradiatus from the Kaoping River, Taiwan. Results of t-test between sexes are also shown (* p < 0.05; ns, non-significant)
Male (n = 113) Female (n = 275) Body measurements Mean (SE) Mean (SE) t386
Body weight (g) 449.6 (104.9) 470.6 (74.3) 4.6* Standard length (cm) 30.9 (2.6) 31.9 (2.6) 4.6* Body depth (cm) 6.0 (0.4) 6.2 (0.4) 2.8* Caudal-peduncle depth (cm) 3.9 (0.3) 4.0 (0.2) 4.3* Predorsal length (cm) 12.0 (1.0) 12.7 (0.8) 4.6* Predorsal to caudal 18.9 (1.5) 19.5 (1.4) 4.5* peduncle (cm)
Length of anal base (cm) 3.5 (0.3) 3.6 (0.2) 4.5* Head length (cm) 7.5 (0.5) 7.6 (0.4) 4.7* Gape width (cm) 8.0 (0.6) 8.1 (0.4) 4.3* Mouth length (cm) 3.6 (0.4) 3.6 (0.2) 1.6ns Mouth width (cm) 4.0 (0.3) 4.1 (0.2) 3.7* Eye diameter (cm) 0.9 (0.1) 0.9 (0.1) 4.2* Whisker length (cm) 2.5 (0.4) 2.5 (0.3) 4.5ns
Two hundred thirteen females (91 without enlarged eggs and 122 with enlarged eggs) from the tributaries were used to determine the repro-ductive phenology of L. multiradiatus in the Kaoping River (Fig. 5). Monthly means of GSI val-ues began to increase in Mar., reached a peak in June, and decreased to below 1 after Oct. Eggs of mature females were observed to gradually enlarge from 1.5 to 3~4 cm, on average, when entering the reproductive season. Most of the col-lected females were mature in Apr. (73.7%) and July (84.2%), while higher GSI values occasionally appeared from Oct. to Feb., suggesting that some mature females still existed.
Sex ratio
In total, 388 collected individuals, including 275 females and 113 males, were used to examine the sex ratio of L. multiradiatus in the Kaoping River. The annual sex ratio was female-biased (2.43 females: 1 male) and statistically different from 1: 1 (x2 = 25.7, df = 1, p < 0.05). Although
fewer males were collected in both the reproduc-tive and non-reproducreproduc-tive seasons, the proportion of females in the reproductive season (Mar.~Sept., 3.3 females: 1 male) was significantly greater than
that in the non-reproductive season (Oct.~Feb., 1.9 females: 1 male) (x2 = 5.63, df = 1, p < 0.05)
(Table 2).
The sex ratios of different LS divisions varied, and significantly more females were collected than males for those with a LS of 31~34 cm (3 females: 1 male; x2= 15.2, df = 1, p < 0.01), 34~37 cm (4.6
females: 1 male, x2= 18.5, df = 1, p < 0.01), and
37~40 cm (3.7 females: 1 male, x2= 6.3, df = 1, p
< 0.05) (Fig. 6). However, sex ratios of LSs of 25~28 cm and 28~31 cm did not significantly devi-ate from 1: 1 (Chi-square test, p > 0.05). Although the proportion of mature females slightly decreased for LSs > 34 cm, more than 50% of females for LSs of > 31 cm were sexually mature (Fig. 6).
DISCUSSION Body size and habitat selection
Data obtained from the main stream and trib-utaries indicated differences in habitat use between large and small L. multiradiatus. Smaller fish (LS < 20 cm) were collected only from the trib-utaries, whereas larger fish (LS > 40 cm) were col-lected only from the main stream. The uneven size distribution in different habitats suggests that the early development of this species occurs in smaller channels of streams. After L. multiradiatus has passed the 1st phase of development, it may
16 14 12 10 8 6 4 2 0
Dec Jan Feb Mar Apr May June July Aug Sept Oct Nov
Dec Jan Feb Mar Apr May June July Aug Sept Oct Nov
GSI 20 18 16 14 12 10 8 6 4 2 0
Fig. 5. Temporal variations of the gonadosomatic index (GSI) and its mean (•) of female Liposarcus multiradiatus from the main stream (top) and tributaries (bottom) of the Kaoping River, Taiwan. The mean values were connected to indicate the reproductive phenology. Standard Length (cm) Number 40 20 0 20 40 60 80 <25 25-28 28-31 31-34 34-3737-40 Female
Sexually Mature Female Male
Fig. 6. Collected male and female individuals of Liposarcus
multiradiatus in 6 standard size classes. Standard size classes
are divided into 3 cm intervals. Sexually mature females were determined by the presence of enlarged eggs in their ovaries.
then move from side channels to the mainstream, or stay in tributaries for a longer time. A similar behavior of habitat switching in different life stages has also been reported for other armored catfish species (Power 1984a, Mazzoni and Caramaschi 1995) as well as various other fish species (Schlosser 1982). A lower current velocity, fewer predators, and greater feeding opportunities in smaller streams have all been proposed as rea-sons for armored catfishes switching habitats dur-ing different life stages (Mazzoni and Caramaschi 1995). Because it has evolved external bony plates, and potential endemic predators have little or no experience with this species, predation pres-sure on young L. multiradiatus may be less intense in Taiwan than in its original distribution range. Thus, habitat adaptations, such as to water current velocity or food supply, may play more-important roles in shaping the uneven distribution of this species in different life stages after having accli-matized to Taiwan.
Morphometric classification and growth patterns
Distinguishing the two sexes of L. multiradia-tus based on 13 morphological measurements was unsatisfactory in this study because the classifica-tion correctness was only 61%~66%. In addiclassifica-tion, certain similar growth patterns were documented in both sexes. Therefore, it is difficult to separate individuals of L. multiradiatus between sexes based on morphological characteristics such as body length, body weight, or the 13 morphological characteristics we analyzed. According to our field experience, one way to quickly identify a female fish in the reproductive season is to press the fish,s belly and observe whether yellow eggs appear at the vent.
The exponential power parameters of the growth model for L. multiradiatus, for males only (2.3), females only (2.2), and the 2 sexes together (2.5), were all smaller than 3. This finding indi-cates that the body shape of L. multiradiatus becomes less rotund as the length increases, pos-sibly due to the external restriction of the bony plates (Nielsen and Johnson 1983).
Reproductive phenology
Monthly analysis of female GSI values sug-gested that the reproductive period for L. multira-diatus in Taiwan lasts for at least seven mo, from Mar. to Sept. In the lower section of the Paraiba
do Sul River in Brazil, the reproductive season, which was also based on seasonal variations of mean GSI values, extends for a 10-mo period from July/Aug. to Mar./Apr. for Hypostomus luetkeni (Steindachner 1878) and Hypostomus affinis (Steindachner 1877) (Mazzoni and Caramaschi 1995). The disparity in reproduction times between the armored fish in Brazil and Taiwan may be due to Brazil,s location in the Southern Hemisphere and Taiwan being situated in the Northern Hemisphere. Nevertheless, from a sea-sonal perspective, the reproductive periods of these three armored fish species all peak in mid-summer and reach their nadir during the winter season.
The fact that some females had elevated GSI values in Dec. indicates that the reproductive peri-od of L. multiradiatus in Taiwan might last all year. The ability of this species to reproduce all year has also been reported in Hawaii (Yamamoto and Tagawa 2000). Fractional spawning may be another possible explanation for the temporal asynchronous maturity found in females, as has been suggested for H. luetkeni (Mazzoni and Caramachi 1997). Further investigation is needed to clarify these two possible explanations.
Sex ratio
The overall sex ratio of L. multiradiatus in Taiwan was found to be significantly female-biased. Variations of the sex ratio in different body length categories were also documented. A 1: 1 sex ratio was found for fish with LSs of < 25 cm. For those with LSs of 31~40 cm, significantly more females were collected than males. The female-biased sex ratio found in Taiwan differs from the two Hypostomus Lacèpéde 1803 species in Brazil,s lotic waters. Equal sex ratios were observed for H. luetkeni and H. affinis with LSs of < 25.6 cm (Mazzoni and Caramachi 1995). Furthermore, for
Table 2. Collected individuals of male and female
Liposarcus multiradiatus in the sampling period, during the reproduction season (Mar.~Sept.), and the non-reproduction season (Oct.~Feb.)
Number (%)
Male Female Total
Sampling period 113 (29.1) 275 (70.9) 388 Reproductive season 44 (23.5) 143 (76.5) 187 Non-reproductive season 69 (34.3) 132 (65.7) 201
those H. affinis with LSs of > 25.6 cm, a male-biased sex ratio was found. Higher female mortali-ty due to a greater reproductive investment was suggested as a possible reason for the male-biased sex ratio in larger H. affinis. Compared to the two Hypostomus species, L. multiradiatus in Taiwan has a later onset of sexual maturity (LSs of 25 cm for L. multiradiatus, 14.4 cm for H. affini, and 16 cm for H. luetkeni) (Mazzoni and Caramaschi 1995). This indicates that female L. multiradiatus in Taiwan might invest less energy in reproduction than do H. affinis and H. luetkeni in Brazil. Given that L. multiradiatus was only recent-ly introduced into Taiwan, a female-biased sex ratio may result from less energy input of females towards reproduction and more energy for environ-mental adaptation, thereby lowering female mortal-ity. Another possible and simpler explanation for the female-biased sex ratio in Taiwan is that more females have been imported than males, and pro-portionally more females have been released.
In this study, fewer males than females were found during the reproductive period. Similar observations were documented for two Hypostomus species, in which fewer males than females were found. Males practicing parental care during the reproductive season have been proposed as a possible explanation (Mazzoni and Caramachi 1995). Recently, males of L. multira-diatus have been reported to exhibit parental care behavior in Hawaii (Yamamoto and Tagawa 2000). Thus, it is quite likely that males of L. multiradiatus in Taiwan may also be performing parental care during the reproductive season.
Environmental impact and management implications
The results of this study show that L. multira-diatus in Taiwan is equipped with a high tolerance to polluted environments from its possible air-breathing ability (Fernandes and Perna 1995), has at least a 7-mo, and possibly extended, reproduc-tive period, and may exhibit parental care. Power (1984a) reported that the armored catfish species (Ancistrus Bleeker, 1862) in Panama occupies clear water, and younger Ancistrus commonly sheltered on cobble substrate in riffles (5~40 cm in depth) until they grow to > 4 cm LS. Additionally, larger Ancistrus may enter shallow waters and rif-fles at night or during periods of high stream flow. Upstream invasion of L. multiradiatus has recently been reported in eastern Taiwan, where pristine streams still exist (Chen et al. 2003). Thus, it is
expected that L. multiradiatus in southern Taiwan will very likely move from polluted downstream regions to clear or least-polluted upstream reaches in the future.
Few aquatic species in Taiwan have been found to use the bottom detritus as their major food sources as does L. multiradiatus (Huang et al. 1994). Power (1990) observed that only 6 armored catfish (mainly Ancistrus)/m2 were
need-ed to deplete all algae and clear sneed-ediment for bedrock substrata in their original distribution range. If the population of L. multiradiatus contin-ues growing in the Taiwan,s lotic waters, L. multira-diatus will potentially have serious negative impacts on those endemic species with bottom-attached eggs or with benthic algae/detritus feed-ing habits.
Separating males and females L. multiradia-tus based on morphological measurements was ineffective. Additionally, the growth patterns of males and females are similar. It seems difficult, at least for the present, to establish or execute a management strategy or technique requiring sex identification in an effort to extirpate this invading species. A temporary constraining method, such as using incentives to encourage fishermen to catch this species during its reproductive season from Mar. to Sept., may have some merit. However, as formerly suggested, educating the general public not to release young or adult L. mul-tiradiatus indiscriminately into local waters may be the most-effective way to control its further expan-sion in Taiwan.
Acknowledgments: This study was supported by
the Council of Agriculture, Taiwan through a research grant (COA-89-1-5-Foresty-05(4)) award-ed to S.H. Liang and B.S. Shieh. We are really grateful to two anonymous reviewers for extremely helpful comments on the manuscript.
REFERENCES
Balirwa JS, CA Chapman, LJ Chapman, IG Cowx, K Geheb, L Kaufman, RH Lowe-McConnell, O Seehausen, JH Wanink, RJ Welcomme, F Witte. 2003. Biodiversity and fishery sustainability in the Lake Victoria Basin: an unex-pected marriage? BioScience 53: 703-715.
Cailliet GM, MS Love, AW Ebeling. 1986. Fishes: a field and laboratory manual on their structure, identification, and natural history. Belmont, CA: Wadsworth Publication. Chen RT, PH Ho, HH Lee. 2003. Distribution of exotic
fresh-water fishes and shrimps in Taiwan. Endem. Sp. Res. 5: 33-46. (in Chinese)
Rome. Italy: Food and Agriculture Organization.
Fernandes MN, SA Perna. 1995. Internal morphology of the gill of a loricariid fish, Hypostomus plecostomus: arterio-arterial vasculature and muscle organization. Can. J. Zool. 73: 2259-2265.
Fernandes MN, SA Perna, SE Moron. 1998. Chloride cell api-cal surface changes in gill epithelia of the armoured cat-fish Hypostomus plecostomus during exposure to distilled water. J. Fish Biol. 52: 844-849.
FishBase. Apr. 2004. Available at http://www.fishbase.org/ home.htm
Huang LS, IS Chan, CC Hung. 1994. Fishes in Kaoping River. Kaohsiung, Taiwan: Office of Kaohsiung County. (in Chinese)
Jiang GH. 1996. New dominator in the streams. Taipei, Taiwan: Freedom Times, 12 June ed. (in Chinese) Ludlow ME, SJ Walsh. 1991. Occurrence of a South American
armored catfish in the Hillsborough River, Florida. Fla. Sci. 54: 41-48.
Marsh PC, ME Douglas. 1997. Predation by introduced fish on endangered humpback chub and other native species in the Little Colorado River, Arizona. Trans. Am. Fish. Soc. 126: 343-346.
Mazzoni R, EP Caramaschi. 1995. Size structure, sex ratio, and onset of sexual maturity of two species of
Hypostomus. J. Fish Biol. 47: 841 - 849.
Mazzoni R, EP Caramaschi. 1997. Observations on the repro-ductive biology of female Hypostomus luetkeni Lacèpéde 1803. Ecol. Freshw. Fish 6: 53-56.
Nielsen LA, DL Johnson. 1983. Fisheries techniques. Bethesda, MD: American Fisheries Society.
Page LM, BM Burr. 1991. A field guide to freshwater fishes of
North America north of Mexico. Boston, MA: Houghton Mifflin.
Pimentel D, S McNair, J Janecka, J Wightman, C Simmonds, C O,Connell, E Wang, L Russel, J Zern, T Aquino, T Tsomondo. 2001. Economic and environmental threats of alien plants, animals, and microbe invasions. Agric. Ecosyst. Environ. 84: 1-20.
Power ME. 1984a. Habitat quality and the distribution of algae-grazing catfish in a Panamanian stream. J. Anim. Ecol. 53: 357-374.
Power ME. 1984b. The importance of sediment in the grazing ecology and size class interactions of an armored catfish,
Ancistrus spinosus. Environ. Biol. Fish. 10: 173-181.
Power ME. 1990. Resource enhancement by indirect effects of grazers: armored catfish, algae, and sediment. Ecology 71: 897-904.
Schlosser IJ. 1982. Trophic structure, reproductive success, and growth rate of fishes in a natural and modified head-water stream. Can. J. Fish. Aquat. Sci. 39: 968-978. Townsend CR. 1996. Invasion biology and ecological impacts
of brown trout in New Zealand. Biol. Conserv. 78: 13-22. Vos LD, J Snoeks, DT Van Den Audenaerde. 1990. The
effects of tilapia introductions in Lake Luhondo, Rwanda. Environ. Biol. Fish. 27: 303-308.
Williamson M. 1996. Biological invasion. New York, NY: Chapman and Hall Press.
Yamamoto MN. 1992. Occurrence, distribution and abundance of accidentally introduced freshwater aquatic organisms in Hawaii. Honolulu, HI: Freshwater Fisheries Research and Surveys.
Yamamoto MN, AW Tagawa. 2000. Hawaii,s native and exotic freshwater animals. Honolulu, HI: Mutual Publishing.