248
Early Life History Traits of Japanese Anchovy in the Northeastern
Waters of Taiwan, with Reference to Larval Transport
Chih-Shin Chen and Tai-Sheng Chiu*
Department of Zoology, National Taiwan University, Taipei, Taiwan 10764, R.O.C. (Accepted Fedruary 11, 2003)
Chih-Shin Chen and Tai-Sheng Chiu (2003) Early life history traits of Japanese anchovy in the northeastern waters of Taiwan, with reference to larvae transport. Zoological Studies 42(2): 248-257. There is controversy concerning the location of spawning sites and recruiting routes of Japanese anchovy in the northeastern Taiwan. Early in the larval fish fishery season, we conducted a survey along a presumptive coastal current to Ilan Bay with the aim of obtaining anchovy larvae for demographic analysis. The obtained samples of the Japanese anchovy, Engraulis japonicus, were analyzed to elucidate the species,directional tendency, which was from offshore to inshore, and which reached coastal Ilan Bay along the western boundary of the Kuroshio Current during late April. Demographic analysis indicated that offshore anchovy larvae had the smallest body size, and that the size increased from offshore toward the coastal. Based on the sagittal microstructural analy-sis, detailed early life history traits, such as increment size, standard length and body weight at daily ages were back-calculated, then various relationships were estimated, and significant differences were shown to have an offshore-coastal tendency. Estimated developmental stability exhibited highest variation in transitional inshore area and lowest variation in coastal area. The mean daily growth rate in standard length was 0.28 mm d-1for larvae in the offshore area and 0.54 mm d-1for those in the coastal area. However, the inshore larval popula-tion exhibited a non-steady growth pattern, which was 0.27 mm d-1corresponding to an offshore environment during a period of 7-24 April, but had a subsequent higher growth rate of 0.59 mm d-1, whose value was similar to that of the coastal population. These findings combined with local flow pattern and sea surface temperature images support the hypothesis that anchovy larvae are transported from offshore at Pangaiyu to the coastal area of the Ilan Bay, along with a previously identified coastal current in northeastern Taiwan waters. Coastal area of Ilan Bay serves as an important nursery ground for late anchovy larvae.
http://www.sinica.edu.tw/zool/zoolstud/42.2/248.pdf
Key words: Otolith microstructure, Daily growth, Development instability.
J
apanese anchovy, Engraulis japonicus Schlegel, 1846, is a major component of the catches in the northeastern Taiwan coastal fishery, usually being found in spring and late summer (Chiu et al. 1997). It ranges from Japan to most of the Taiwan Strait (Hayasi 1967, Young et al. 1994). Fishery practices have revealed a series of rela-tively discrete areas in which it is distributed along the coast of Taiwan (Yu and Chiu 1994). Spawning of Japanese anchovy is concentrated temporally, usually from late Mar. to May and late July to Aug. (Chiu and Chen 2001). Spawning adults found concentrated somewhere in the off-shore areas of Ilan Bay, Taiwan were 11-12 cm instandard length with more than 50% of the group being mature (Young and Chiu 1994). The eggs are pelagic and hatch in about 4 d (Tsuji and Aoyama 1984). Late larvae are found concentrat-ed in coastal areas, and become a target of coastal trawl fisheries, but the abundance fluctuate among years, possibly due to annual differences in environmental conditions (Lee et al. 1990). The coastal phase of the larval anchovy lasts 6-8 wk, during which time most individuals become juve-niles; they subsequently leave coastal area at about 60 d of age when they are 40-50 mm in SL (Chiu and Chen 2001).
A monthly geographic distribution analysis
248
*To whom correspondence and reprint requests should be addressed. Tel: 886-2-23630231 ext. 2128. Fax: 886-2-23634014. E-mail: [email protected]
indicated that the larval stock first appears in offshore of northeastern Taiwan, and subsequently locates in a more extended area off northeastern Taiwan, being especially concentrated in Ilan Bay (Chiu et al. 1997). This result indicates a shoreward trans-portation of the larvae. However, the result also contradicts the findings of Young and Chiu (1994), who inferred that larvae were born in Ilan Bay. The Japanese anchovy has a protracted spawning time, which results in multiple recruitment cohorts with temperature-dependent growth rates (Wang and Tzeng 1999). By sampling at very short time interval of 3 d to elimination bias caused by asyn-chronous growth factors and other factors (Crecco and Savoy 1985), we compared spatial differences of the early life history traits of anchovies in an attempt to elucidate the offshore-coastal relation-ship.
MATERIALS AND METHODS Study area and data collection
Ilan Bay is an estuarine, which receives fresh-water from the Lanyang River, the largest river on the eastern coast of Taiwan (Fig. 1). The water in Ilan Bay, which receives little freshwater runoff was defined as an inshore area for this study. The off-shore area was located in the vicinity of the Pangaiyu Island, where local upwelling is frequent-ly found (Liu et al. 1992). The coastal area was identified as waters influenced by fresh water runoff, where is primarily located in shallow water less than 20 m deep very closed to the shoal of the bay, and where larval anchovies has traditionally been harvested by regular trawling practices. Inshore and offshore transects were designed along a presumptive coastal current located at the western boundary of the Kuroshio Current in between either landmass or shelf edge (Fig. 1; see Chuang et al. 1993, Tang et al. 1999), and sam-ples were collected using an ichthyoplankton net on 30 Apr. - 1 May 2000, on board R/V Ocean
Research II (350 t). Since the research vessel
was too large to access the shallower areas, coastal samples were collected the next day (2 May 2000) on board a commercial fishing boat using the same ichthyoplankton net. At each sam-pling station, temperature and salinity were mea-sured at 1-m depth intervals to as closed to the bottom as possible using a CTD profiler (Sea-Bird Electronic, Seattle, USA), with a mounted fluores-cence sensor to measure relative primary
produc-tivity. The ichthyoplankton sampler consisted of a ring net of 1.3-m diameter, with 4-m length, and 1.0 mm mesh. At each station, the net was towed in an oblique haul to sample the water column to about 5 m above the bottom. The net was deployed and retrieved at wire speed of 0.5 m s-1
while speed of ship was estimated 2 mi s-1 to
maintain minimum headway. All eggs and fish were preserved in 95% ethanol. Elliptical eggs with 1.0-1.5 mm long and 0.5-0.7 mm width were identified as Engraulis japonicus, because hardly any other fish spawning in the early spring has egg with elliptic shape in the study area (Chang et al. 1985). Larvae of the Japanese anchovy were identified and measured to 0.01 mm standard length and 0.1 mg. The sagittae from both sides of the head were picked out and placed on a micro-scope slide under a binocular micromicro-scope. The otoliths were mounted in Permount and covered with a glass slip.
Data analysis
Both otoliths of each larva were observed under a compound microscope at 200x, and a clear image of the sagitta was obtained using a hard copier connected to the image analysis sys-tem. All distinctive increments presumably identi-fied as daily rings were marked with a pen. The coordinates of sagittal perimeter and increment position were obtained using a digitizer (MM1201 Data tablets, Summagraphics Corp., Seymour, USA), and stored in a computer file. Because the position of increments was first examined and marked on the hard copier output, and then digi-tized, the number of increments was exactly deter-mined, and the increment sizes were precisely cal-culated. Little difference was found in the shape outline of the left and right otolith of the same specimen, hence only the right sagitta was used in growth analysis. The number of the increments plus 4 was assumed to be the actual daily age of the fish according to the findings of Tsuji and Aoyama (1984). Procrustes distance (Goodall 1990), which measures the fluctuation asymmetry between the right and left otoliths, was used to measure the degree of fitness of the larva in adapting to their environments; i.e., a shorter dis-tance indicates a better environment for larval development, while a higher one indicates more unstable conditions which may imply a transitional stage (Somarakis et al. 1997). A back-calculation method was used to estimate growth differences based on the identified daily increments
(Moksness and Wespestad 1989, Jones 2000).
RESULTS
Hydrograph, fluorescence index, and larval densities
Hydrographic conditions were typical for waters of northeastern Taiwan during the early spring. Three water types could be identified from the temperature-salinity (T-S) diagrams: offshore (stns. C-F), inshore (stns. 1-3) and coast (stns. 0, A and B) (Fig. 2). The T-S line pattern of stns. E and F,s T-S line is identified as being within the Kuroshio Current proper, according to a previous study (Chiu and Hsyu 1994). The water mass from stn. C was very similar to that of inshore but with higher temperature and salinity is categorized as the shoreward margin of the offshore, because the location is removed from the estuary of the Ilan Bay. Upwelling and mixing waters is observed at stn. D, where the surface was relatively cold (ca. 23
°
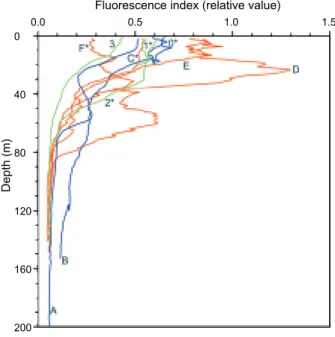
C) and the T-S line situated between stns. C and E. Coastal water is evidenced by a horizontal T-S line, where low salinities were observed.The depth profile of the fluorescence index is shown in figure 3. The highest value was located at the depths of 30-40 at stn. D, where TS dia-grams indicated upwelling and mixing water. The lowest value was located in the surface water of
stn. F, but a higher value was also found in the subsurface at a depth of 50-70 m depth. The inshore and coastal stations had median fluores-cence values.
The abundance of fish eggs, larvae, and
zoo-Fig. 1. Location of sampling stations in the northeastern
waters of Taiwan. Stations C-F are treated as offshore, stns. 1-3 as inshore and stns. 0, A, B as coastal areas. Japanese anchovy larvae were found at stations shown by filled circles. The schematic coastal current is a composite based on Wang and Chen (1990) and Chuang et al. (1993).
Fig. 2. Temperature-salinity (T-S) diagrams of the water mass
from northeastern Taiwan. T-S lines from offshore water are located to the right, an area which is identified as the origin of Kuroshio Current. The line of coastal waters is horizontal, which indicates freshwater intrusion. Anchovy larvae were found at starred stations.
Fig. 3. Depth profiles of fluorescence index from the waters of
northeastern Taiwan. Anchovy larvae were found at starred stations. Salinity (ppt) Latitude (
°
N) Depth (m) T emperature (°
C)Fluorescence index (relative value)
28 26 24 22 20 18 16 14 12 0 40 80 120 160 200 32 0.0 1.5 33 34 35 Longitude (
°
E) 26 25 24 121 122 123 124 Latitude (°
N) 28 26 24 22 20 Longitude (°
E) 118 120 122 124 126 0.5 1.0plankton are shown in table 1. No significant differ-ence (at p = 0.05) were found in comparisons across stations between fish egg and zooplankton densities. However, a significant difference (p = 0.0068) was found in larval fish density. Three homogeneous station groups were identified by multiple-range test, i.e., stns. 2, 3, and A-F; 0, 2, 3, and B-E; and 0-2, and B, however none of them can be discriminated without overlapping.
Size and age of larvae
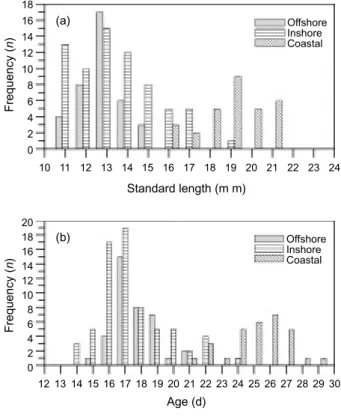
Larvae captured offshore ranged from 11.01 to 15.11 mm in length and ranged from 15 to 21 d in age; they were similar to inshore larvae at 10.80 to 19.13 mm and 14 to 24 d, and contrasted with the significantly bigger and older larvae at 16.48 to 21.44 mm and 21 to 29 d found in coastal area (Fig. 4). It is apparent that small-sized, young lar-vae were found in offshore and inshore areas, but not in the coastal area. There was no difference in the average standard length of anchovy larvae of the offshore and inshore (12.91 ± 0.41 mm and 13.52 ± 0.41 mm, respectively), while that of larvae in the coast was significant larger (19.04 ± 0.55 mm, p < 0.001). Similarly, the ages of anchovy in offshore and inshore waters were almost the same
(17.70 ± 0.52 d and 17.46 ± 0.48 d, respectively), while that of coastal larvae was significantly older (21.13 ± 0.69 d, p < 0.001) (Fig. 4b).
Relationship among length, increments, and otolith radius
The otolith radius (OR) exhibited a linear rela-tionship with the number of increments (NI) count-ed along the measurement axis supportcount-ed by a minimal probability level of 1%, OR = 6.31 * NI -13.48 (Fig. 5), however group differences could also be detected statistically. Analysis of each increment group and testing for lack of fit also showed a significant difference (lack of fit F14,121 = 16.955), in which the increment count at 11-14 had group averages of less than expected values, but they were compensated for to reach the expected value of the regression line after a count number of 15.
The standard length-otolith radius (SL-OR) relationships were different between the offshore and inshore groups (Fig. 6). The resulted formulas were SL = 10.78 + 0.045 OR and SL = 7.20 + 0.139 OR, for the offshore and inshore populations respectively. However, the coastal larvae exhibit-ed a relationship of SL = 8.46 + 0.110 OR. Despite the ranges of otolith radius having a small overlap between the inshore and coastal groups, we made all possible comparisons of the SL-OR relationships between the 3 groups. Finally, the same relationship was concluded to be statistically similar between the inshore and coastal groups (p < 0.001), and the combined relationship was SL = 8.30 + 0.113 OR (n = 98, r = 0.961).
Increment size
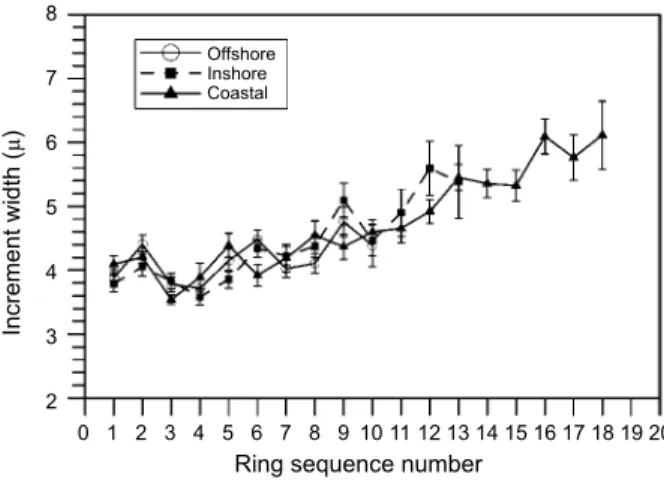
The pattern of otolith accretion history was similar between the offshore and inshore popula-tions (Fig. 7). All increment widths were compara-ble when ring number was less than 10 (ca. age of 14 d), while they are higher in coastal group when they were aged greater than 14 d (at an increment number 10). Only increment counts, which were duplicated more than 10 were combined in the analysis, because variations could be large as width counts were small. Daily increment widths ranged from 3.71 to 5.31 µ d-1 and from 3.57 to
5.75 µ d-1, for offshore and inshore populations,
respectively. However, the increment width of the coastal larvae ranged 3.69 to 6.09 µ when the lar-vae were aged less than 19 d. Variations in the increment width were found in 3 groups, but no
Fig. 4. Length and age frequency distributions of the Japanese
anchovy larvae captured in the waters of northeastern Taiwan.
Frequency ( n ) Frequency ( n ) 18 16 14 12 10 8 6 4 2 0 20 18 16 14 12 10 8 6 4 2 0 Standard length (m m) Age (d) 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 (a) (b) Offshore Inshore Coastal Offshore Inshore Coastal
significant difference was found when larvae were aged less than 14 d (ring number ≤ 10).
Developmental stability
Larvae captured in inshore had the largest range of Procrustes distance (Pd) of from 8.29 to 36.21 µ, followed by those from the offshore areas at 8.29-32.44 µ (Fig. 8). Larvae found in the coastal area had a minimal Pd range of 8.76-25.69 µ. A decreasing trend for Pd as SL increased is found when the scatter plots of Pd on SL were elaborat-ed (Fig. 8). To eliminate the size effect on the Pd score, SL was taken as the covariate, and the group means of the Pd value per unit SL were esti-mated by Pd = 30.702 - 0.914 SL (n = 11, r = 0.8951, p < 0.001). Size adjusted Pd values indi-cated that the coefficients of variation of Pd value were 47.0%, 46.1%, and 19.2%, with variances of 6.22, 6.66, and 3.83 µ for the offshore, inshore, and coastal areas respectively (Fig. 9a-b).
Significant differences among groups were support-ed by analysis of variance (ANOVA) (p = 0.0002). Multiple range analysis yielded 2 homogeneous groups of offshore + inshore and coastal areas. Back-calculated growth history
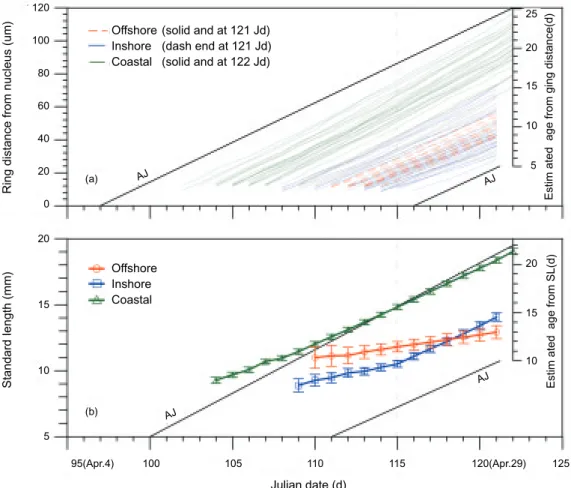
Individual growth histories were back-traced based on otolith microstructural analysis (Fig. 10). Larvae captured in the offshore areas had hatched as early as 14 Apr., and as late as 19 Apr., while those of inshore areas had hatched between 7 and 21 Apr. Larvae in the coastal area were older, having hatched from 7 to 15 Apr. The hatching period of larvae captured inshore covered both periods of the offshore and coastal areas,
indicat-Fig. 5. The relationship of otolith radius and number of
incre-ments.
Fig. 6. The relationships of standard length and otolith radius.
Fig. 7. Variation of increment width with the sequence of the
daily ring counted from the center of otolith. The vertical bar indicates 1 standard deviation.
Fig. 8. Scatter plots of relative developmental instability
esti-mated from Procrustes distance of asymmetrical property of each anchovy larva. Group means were estimated from each unit standard length. The regression line was fit to the group means.
Ring sequence number
Increment width ( µ ) Otolith radius ( µ ) Developmental instability Otolith radius (µ) Number of increments (n) Standard length (mm) Pd = 30.702-0.914 SI(n = 11, r = 0.8951, p < 0.001) Offshore Inshore Coastal Offshore SL=10.78+0.045 OR Inshore SL= 7.20+0.139 OR Coastal SL= 8.46+0.110 OR 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 2 40 10 30 50 70 90 100 130 30 20 10 0 3 4 5 6 7 8 Offshore Inshore Coastal Offshore Inshore Coastal 9 12 15 18 21 24 25 0 5 10 15 20 25 20 15 10 5 120 100 80 60 40 20 OR = 6.31*NI-13.48 N=137 R2=0.986 Standard length (mm)
ing a link (transition) between them. Reading the daily SL trajectory is quite complex, but individual growth can be back-estimated based on the ring distance read from otoliths, and compared to a theoretical age-Julian (AJ) dateline that shows an ideal growth pattern. For instance in figure 10a, retarded growth track is shown by the otolith ring-distance line which is flatter than the AJ line, such as that for offshore larvae; while normal growth is shown by being parallel to the AJ line as that for coastal larvae. This phenomenon indicates that the growth of otoliths accelerated in coastal larvae, but indicated arrested growth for larvae found in offshore areas.
The daily growth trajectory of SL measured at group means is shown in figure 10b. Almost uni-form growth was observed in offshore and coastal larvae with growth rate higher in coast than off-shore. However, an apparently discontinues growth pattern was found in inshore larvae which
showed an accelerated growth after Apr. 24 (115 Julian date) at estimated group age of about 10 d. Growth rate is lowest in offshore larvae, averaged 0.28 mm d-1 and highest in coast of 0.54 mm d-1.
The inshore larvae has a median group growth rate of 0.27 mm d-1 estimated from the period of
Apr. 7-24 at average of ca. 10 d, but has a higher
Fig. 9. Residual plots of the Procrustes distance along
stan-dard length (a), and coefficient of variation relative to SL (b).
Fig. 10. Back-traced accretion of sagittal otolith for each larva captured from 3 hydrographic areas of northeastern Taiwan (a). The
group means and standard errors of the mean of standard length were also estimated at each Julian date (b). Theoretical daily age scales converted from ring distance and standard length were also plotted at the date of capture. The age-Julian (AJ) datelines were plotted to show relatively accelerated or retarded growth.
Julian date (d)
Ring distance from nucleus (um)
Standard Iength (mm) Estim ated age from SL(d)
Estim ated age from ging distance(d)
Offshore Inshore Coastal Residual of Pd (um) Ooef ficient of variation 20 10 0 -10 60 40 20 0 10 120 100 80 60 40 20 0 20 15 10 5
12 14 16 18 20 22 Offshore Inshore Coastal
(a) (b) 95(Apr.4) 100 105 110 115 120(Apr.29) 125 20 25 20 15 10 5 15 10 (a) (b) Offshore Inshore Coastal
Offshore (solid and at 121 Jd) Inshore (dash end at 121 Jd) Coastal (solid and at 122 Jd)
AJ AJ
AJ
growth rate of 0.59 mm d-1, which is similar to that
of coastal population.
DISCUSSION Distribution of anchovy eggs
Larvae of Japanese anchovy (13 - 30 mm SL) are abundant in the coastal waters of northeastern Taiwan (Chiu and Chen 1998). Young larvae (< 13 mm) of Japanese anchovy are relatively rare in the estuaries of the Ilan coast (Young et al. 1992). In the waters of northeastern Taiwan, the Japanese anchovy exhibited averaged increasing gradient of egg abundance (Table 1) from coastal to offshore areas, and inversed gradient shown by larval size and age (Fig. 4) from offshore to coastal areas during the 3-d study across the 3 inshore-offshore areas, however a significant difference among off-shore-coastal categories was not confirmed statis-tically. The offshore occurrence of abundant fish egg patches was not uncommon event in ichthyo-plankton surveys (Boehlert and Mundy 1988, Tzeng and Wang 1993, Chiu et al. 1997). This phenomenon may cause by offshore spawning (Nelson et al. 1997) or offshore entrainment (Nakata et al. 2000). In our data, coastal stn. B showed a high density of anchovy eggs (8790 eggs 1000 m-3, Table 1), which may imply
wide-spread spawning by mature adults. The ubiquity anchovy egg occurrence may have resulted from offshore eddies via advection (Bartsch et al. 1989, Townsend 1992). On the other hand, late anchovy
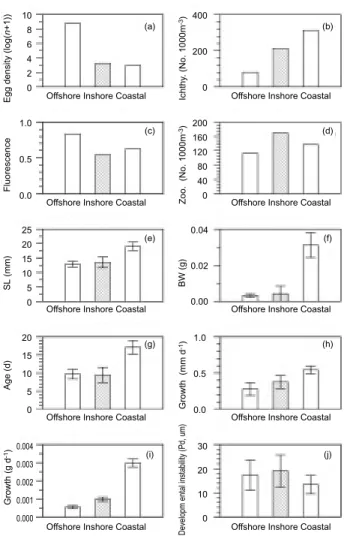
Fig. 11. Summary of demographic traits categorized into 3
environments of offshore, inshore, and coastal area off north-eastern Taiwan. Bars in the histograms indicate 1 standard deviation. Ichthyoplankton (No. 1000m-3) Egg (No. 1000m-3) Larvae obtained (No.) Depth (m) Table 1. Abundance of zooplankton, ichthyoplankton, and Japanese anchovy egg and larvae in the waters of northeastern Taiwan during studying period of 30 April and 2 May 2000 Station Zooplankton (g 1000m-3) Offshore C 41.5 128.9 29.4 35 151 D 21.1 71.1 16662.3 - 220 E 254.4 77.6 5612.1 - 130 F 130.5 45.0 3666.3 3 100 Inshore 1 125.2 349.2 37.7 54 40 2 309.9 159.7 18.1 17 93 3 74.5 129.6 19.1 - 180 Coastal 0 136.0 313.0 18.9 30 20 A 25.0 22.8 21.2 - 190 B 40.4 78.2 8797.4a - 150
The stn B (a), a strong mixing between the coastal and offshore waters is indicated. Please see the
geographic relationships in figure 1, and water mass properties in figure 2.
Offshore Inshore Coastal Offshore Inshore Coastal
Offshore Inshore Coastal Offshore Inshore Coastal
Offshore Inshore Coastal Offshore Inshore Coastal
Offshore Inshore Coastal Offshore Inshore Coastal
Offshore Inshore Coastal Offshore Inshore Coastal
Egg density (log(
n +1)) Fluorescence SL (mm) Age (d) Growth (g d -1) Ichthy . (No. 1000m -3) Zoo. (No. 1000m -3) BW (g) Growth (mm d -1)
Developm ental instability (Pd, um)
10 8 6 4 2 0 400 200 0 25 20 15 10 5 0 0.04 0.02 0.00 20 15 10 5 0 1.0 0.5 0.0 0.004 0.003 0.002 0.001 0.000 30 20 10 0 1.0 0.5 0.0 200 160 120 80 40 0 (a) (b) (c) (d) (e) (f) (g) (h) (i) (j)
larvae were found to be abundant in the coastal area close to the shoal where larval fisheries oper-ate in Ilan Bay. There are several summarized conceptual models to explain offshore planktonic larval movements to estuarine nursery areas (Norcross and Shaw 1984, Boehlert and Mundy 1988). Despite anchovy populations supporting large coastal fisheries in Taiwan, the link between recruitment dynamics and mechanism of fishing ground formation remain unclear as compared to other systems such as those of the North Sea or the Northwest Atlantic (Bartsch et al. 1989, Lee et al. 1990, Townsend 1992).
Shoreward migration
The migration ability of the larval anchovy of less than 13 mm SL is limited; however, the off-shore-to-coastal gradient of early life history traits is obvious (Fig. 11). Several explanations for the shoreward accumulation of late larval anchovy or herring have been proposed, such as surface drift-ing by Ekman transport (Nelson et al. 1977), water column dwelling by internal wave or tides (Shanks 1998) or movement by a frontal jet of geostrophic flow (Lafuente et al. 2002). Comprehensive test-ing of relevant hypotheses of the recruitment of anchovy larvae to Ilan Bay requires data from extensive systematic survey, but the data are apparently still unavailable. Nonetheless, flow pat-terns in the waters of northeastern Taiwan have been shown to possibly be effective in transporting anchovy larvae shoreward to estuary (Tang et al. 2000). The most significant current in
northeast-ern Taiwan is the Kuroshio, which flows northward along 200-m isobaths, and in which no anchovy larvae were found in this study (Fig. 1). A part of the surface Kuroshio water branches onto the shelf (Hsueh et al. 1992), on the other hand, the subsur-face water from about 300 m also raise to the shelf through the shelf-edge upwelling (Chern et al. 1990, Wong et al. 1991). The environment down-stream of the upwelling may benefit the growth of early anchovy larvae, yet these hydrographic events seem irrelevant to the southward move-ment of the anchovy. Chuang et al. (1993) identi-fied a southwestward flow located between the inshore edge of the Kuroshio and the continental slope. The southwestward flow is quasi-steady in the vicinity of the Pangaiyu and along the shelf edge, and becomes the long shore current of northeastern Taiwan (Tang et al. 1999, Fig. 12a). This current is the major mechanism, which poten-tially explains the transportation of anchovy larvae to Ilan Bay estuaries. In the mean while, the sea surface temperature image of the month of Apr. 2000 (our sampled larvae lived from 7 Apr. to 1 May 2000, see text) also suggests a distinct water mass stretching from Pangaiyu to the Ilan Bay in which the dispersal of larvae might have been channeled (Fig. 12b).
In this study, surface mixing was recognized at stn. D, because the T-S line was located between stns. E and C (Fig. 2), and the water also upwelling from the subsurface since low tempera-ture were observed relative to those in the vicini-ties of stns. C, E, and F. This was also evidenced from the highest fluorescence indices at stns. D
Fig. 12. (a) Near surface (at 16 m) flow pattern of the northeast Taiwan during summer (adopted from Tang et al. 1999 with
permis-sion), and (b) Average sea surface temperature in the water north of Taiwan, April, 2000 (available from the web site of National Center for Ocean Research (NCOR), Taiwan, ROC; used with permission).
27
°
N 26°
N 25°
N 24°
N 28°
N 26°
N 24°
N 16 m →100 cm s-1 118°
E 110°
E 120°
E 122°
E 124°
E (a) (b)and E (Fig. 3). No anchovies were found at stns. D and E, but they were found at stns. C and F, which are downstream of the upwelling and close to the shelf edge where southeastward current occurred (Fig. 12a). On the northeastern tip of Taiwan, a frontal jet may exist that transports anchovies to Ilan Bay, because this oceanographic pattern is very similar to the Atlantic Ionian Stream (AIS) system, which generates a geostrophic flow in the Sicily Channel (Lafuente 2002). The AIS flowing through southwestern Sicily waters gener-ates a local upwelling facilitating primary produc-tion in the upstream, and a jet flow transporting
Engraulis encrasicolus to the downstream coastal
areas. Stations A and B were found to have prop-erties of coastal water since fresh water dilution was exhibited in the T-S diagrams. However, no anchovy were captured in the stns. A and B, because the dilution was too great, i.e., the resul-tant salinities were less than 33.5 ppt at surface. Hydrographic discontinuity in the waters of north-eastern Taiwan may become established during winter especially when the northeastern monsoon is strong, since a thermohaline front situated between the inshore and offshore areas from northeastern coast to Pangaiyu (Wang and Chern 1990). During spring season as the northeastern monsoon wanes, the front recedes and the com-munication between the offshore upwelling and the Ilan Bay estuaries is established (Fig. 12b).
Nursery ground
Little evidence has been shown for multiple stocks of the Japanese anchovy in the waters of northeastern Taiwan, yet different life parameters were found for the larvae entered different marine environments (Yu et al. 2002). We summarize findings in this study in figure 11, where anchovy larvae transport to coastal area is supported by distributions of egg density (a), ichthyoplankton density (b), standard length (e), body weight (f), daily age (g), growth rate by length (h) and by weight (i), and developmental instability (j); howev-er, the coastal area has neither the highest primary productivity (c) nor the most abundant in zooplank-ton density (d). It is worth noting that almost con-tinuous growth of larval SL was observed from inshore to coastal waters, but lower daily SL increases for larvae from offshore environments (Fig. 6, 10b); therefore the coastal environment is apparently important for larvae to gain weight (compare Fig. 11h-i).
Anchovy larvae greater than 15 mm SL at an
age of greater than 16 d (see Fig. 5) preferred to the coastal environment, as compare to those lar-vae with a SL below the estimated value at rings counted to 10-14. This is indirectly evidence by the fact that the growth rate was lowest for off-shore larvae (0.28 mm d-1), highest for coastal
lar-vae (0.54 mm d-1), and median for inshore larvae
(0.27 mm d-1before the age of 10 d, and 0.59 mm
d-1 after the age of 10 d). Stable larval
develop-ment was achieved as the larval reached the coastal area (Fig. 8). Possible causes of these apparent differences in growth rates and measures of developmental instability among environments are either migration (transport) or differential mor-tality. However, size segregation observed almost simultaneously in this study and spatial abundance differences in the Tanshui River estuary (Wang and Tzeng 1997) should favorite the migration (transport) hypothesis. The abundance of late anchovy larvae is possibly influenced by the fac-tors of calm water, food availability, predation pres-sure, and effects of temperature and salinity. Higher growth rates and low developmental insta-bility of the larvae in the coast environment may indicate that the estuary of Ilan Bay are a suitable nursery environment for late larvae and probably early juveniles, yet the factors affecting the estuar-ine distribution of the larvae still need further elab-oration.
Acknowledgments: The authors thank the crewmembers of the R/V Ocean Research II for their helps in the field. Thanks are also extended to Ms. K.Z. Chang and Ms. K.J. Chen for specimen identification and otolith preparation. The material used in the present paper was collected as part of the research project financially supported by the National Science Council of R.O.C. (NSC 89-2611-B-002-002). We are grateful to Dr. T.Y. Tang for making flow chart of figure 12a available and to National Center for Ocean Research for the sea surface temperature image of figure 12b. Comments from 2 anonymous reviewers are highly appreciated.
REFERENCES
Bartsch J, K Brander, M Heath, P Munk, K Richardson, E Svendsen. 1989. Modelling the advection of herring lar-vae in the North Sea. Nature 340: 632-636.
Boehlert GW, BC Mundy. 1988. Roles of behavior and physical factors in larval and juvenile fish recruitment to estuarine nursery areas. Am. Fish. Soc. Sympos. 3: 51-67. Chang ZZ, SF Lu, CC Chao, LF Chen, ZC Chang, IS Chiang.
1985. Fish egg and larvae in neritic waters of China. Shanghai: Shanghai Science and Technology Publisher, 206 pp. (in Chinese)
Chern CS, J Wang, DP Wang. 1990. The exchange of Kuroshio and East China Sea shelf water. J. Geophys. Res. 95: 16017-16023.
Chiu TS, CS Chen. 2001. Growth and temporal variation of two Japanese anchovy cohorts during their recruitment to the East China Sea. Fish. Res. 53: 1-15.
Chiu TS, CS Chen. 1998. A study on the status of ichthyo-plankton in the waters around Taiwan. J. Fish. Soc. Taiwan 25: 161-170.
Chiu TS, YH Hsyu. 1994. Interannual variation of ichthyoplank-ton density and species composition in the waters off northeastern Taiwan. Mar. Biol. 119: 441-448.
Chiu TS, SS Young, CS Chen. 1997. Monthly variation of larval anchovy fishery in I-lan Bay, NE Taiwan, with an evalua-tion for optimal fishing season. J. Fish. Soc. Taiwan 24: 273-282.
Chuang WS, HW Li, TY Tang, CK Wu. 1993. Observations of the countercurrent on the inshore side of the Kuroshio northeast of Taiwan. J. Oceanogr. 49: 581-592.
Crecco VA, TF Savoy. 1985. Effects of biotic and abiotic fac-tors on growth and relative survival of young American shad, Alosa sapidissima, in the Connecticut River. Can. J. Fish. Aquat. Sci. 42: 1640-1648.
Goodall C. 1990. Procrustes methods in the statistical analysis of shape. J. Roy. Stat. Soc. B. 53: 285-339.
Hayasi S. 1967. A note on the biology and fishery of the Japanese anchovy Engraulis japonica (Houttuyn). Rep. Cal. Coop. Oceanic Fish. Invest. 11: 44-57.
Hsueh Y, J Wang, CS Chern. 1992. The intrusion of the Kuroshio by the continental shelf northeast of Taiwan. J. Geophys. Res. 97: 14323-14330.
Jones CM. 2000. Fitting growth curves to retrospective size-at-age data. Fish. Bull. 46: 123-129.
Lafuente JG, A Gacia, S Mazzola, L Quintanilla, J Delgado, A Cuttita, B Patti. 2002. Hydrographic phenomena influenc-ing early life stages of Sicilian Channel anchovy. Fish. Oceanogr. 11: 31-44.
Lee MA, KT Lee, HC Ou. 1990. The larval anchovy fishing ground formation in relation to osmotic pressure changes of the coastal waters along southern Taiwan. J. Fish. Soc. Taiwan 17: 233-245.
Liu KK, GC Gong, S Lin, CY Yang, CL Wei, SC Pai, CK Wu. 1992. The year-round upwelling at the shelf break near the northern tip of Taiwan as evidenced by chemical hydrography. Terrestrial, Atmosphere and Oceanography
3: 243-276.
Moksness EM, V Wespestad. 1989. Ageing and back-calculat-ing growth rates of Pacific herrback-calculat-ing, Clupea pallasii, larvae by reading daily otolith increments. Fish. Bull. 87: 509-513.
Nakata H, S Kimura, Y Okazaki, A Kasai. 2000. Implications of meso-scale eddies caused by frontal disturbances of the Kuroshio current for anchovy recruitment. ICES J. Mar. Sci. 57: 143-152.
Nelson W, MC Ingham, WE Schaaf. 1977. Larval transport and year-class strength of Atlantic menhaden, Brevoortia
tyrannus. Fish. Bull. 75: 23-41.
Norcross BL, RF Shaw. 1984. Oceanic and estuarine transport of fish eggs and larvae: a review. Trans. Am. Fish. Soc.
113: 153-165.
Shanks AL. 1988. Further support for the hypothesis that inter-nal waves can cause shoreward transport of larval inver-tebrates and fish. Fish. Bull. 86: 703-714.
Somarakis S, I Kostikas, N Peristeraki, N Tsimenides. 1997. Fluctuating asymmetry in the otoliths of larval anchovy
Engraulis encrasicolus and the use of developmental
instability as an indicator of condition in larval fish. Mar. Ecol. Prog. Ser. 151: 191-203.
Tang TY, Y Hsueh, YJ Yang, JC Ma. 1999. Continental slope flow northeast of Taiwan. J. Phys. Oceanogr. 29: 1353-1362.
Tang TY, JH Tai, YJ Yang. 2000. The flow pattern north of Taiwan and the migration of the Kuroshio. Cont. Shelf Res. 20: 349-371.
Townsend DW. 1992. Ecology of larval herring in relation to the oceanography of the Gulf of Maine. J. Plankton Res. 14: 467-493.
Tsuji S, T Aoyama. 1984. Daily growth increments in otoliths of Japanese anchovy larvae Engraulis japonicus. Bull. Jpn. Fish. Soc. 50: 1105-1108.
Tzeng WN, YT Wang. 1993. Hydrography and distribution dynamics of larval and juvenile fishes in the coastal waters of the Tanshui River estuary, Taiwan, with refer-ence to esturine larval transport. Mar. Biol. 116: 205-217. Tzeng WN, YT Wang. 1997. Movement of fish larvae with tidal
flux in the Tanshui River estuary, northern Taiwan. Zool. Stud. 36: 178-185.
Wang J, CS Chern. 1990. A transect across the thermohaline front north of Taiwan in wintertime. Acta Oceanogr. Taiwanica 25: 31-54.
Wang YT, WN Tzeng. 1997. Temporal succession and spatial segregation of clupeoid larvae in the coastal waters off the Tanshui River estuary, northern Taiwan. Mar. Biol.
129: 23-32.
Wang YT, WN Tzeng. 1999. Differences in growth rates among cohorts of Encrasicholina puntifer and Engraulis japonicus larvae in the coastal waters off Tanshui River estuary, Taiwan, as indicated by otolith microstructure analysis. J. Fish Biol. 54: 1002-1016.
Wong GTF, SC Pai, KK Liu, CT Liu, CTA Chen. 1991. Variability of the chemical hydrography at the frontal region between the East China Sea and the Kuroshio northeast of Taiwan. Estuar. Coast. Shelf S. 33: 105-120. Young SS, CC Chen, TS Chiu. 1992. Resource characteristics
of young herring-like fish in the I-lan Bay area--Fishing season, major species and size variation. J. Fish. Soc. Taiwan 19: 273-281.
Young, SS, TS Chiu, SC Shen. 1994. A revision of the family Engraulidae (Pisces) from Taiwan. Zool. Stud. 33: 217-227.
Yu HT, YJ Lee, SW Huang, TS Chiu. 2002. Genetic analysis of the populations of Japanese anchovy (Engraulidae:
Engraulis japonicus) using microsatellite DNA. Mar.
Biotechnol. 4: 471-479.
Yu PT, TS Chiu. 1994. Fishery target species of larval anchovy fishery in the western central Taiwan. J. Fish. Soc. Taiwan 21: 227-239.