國立臺中教育大學環境教育及管理研究所
碩士論文
以批次詴驗及兩段式串聯反應槽提升
花生殼厭氧產氫之可行性研究
指導教授〆林明瑞 教授
研 究 生〆顏冠忠 撰
中 華 民 國 一 ○ ○ 年 一 月
謝 誌
在研究所修業期間,首先要感謝恩師林明瑞教授對於學生在專業知識及待人 處事上殷切的指導,使學生受益匪淺,謹致上最誠摯的敬意及謝意。論文口詴期 間,承蒙鄭幸雄、林秋裕、賴俊卲教授悉心指導,並提供學生許多寶貴的意見, 使本論文內容更臻完善,在此獻上最誠摯的感謝。 其中要感謝生命中的貴人─黃富昌老師,老師如同伯樂,引領學生進入豐富 的知識殿堂,並帶給我不一樣的人生觀,讓我得以發揮長才在合宜之處,總不忘 老師的提拔。另外感謝蔡瓊萩老師,因為老師的教導讓學生獲得更多專業知識, 為往後研究道路奠定良好基礎,使學生得以順利畢業。 回想起過去兩年多來艱辛做實驗的日子,幸虧有學長孟宏、一傑願意傳授實 驗之經驗,讓我在實驗上更加順利完成。同時要感謝班上同學宜曄、芝嬿、敬凱、 亮榮、廷甄、堯暉、瑞芳、鴻諭、玉如、佳青、正昌、嘉偉、天琦、素貞、春憲 以及敏如,陪伴著我一起同歡樂,論文也順利完成。也感謝劉惠元老師、林素華 老師、劉思岑老師及品詵學姐於課業及生活上的指導與鼓勵々感謝雅鳳學姐、瑋 育助理,在論文及實驗方面給予協助。最後感謝小賴學長、淳圓學姐、承恩學長、 晏權、嘉鴻、志豪,因為有你們給予的幫忙,我才可以很順利的完成。 最後以此論文獻給我最親愛的家人多年來的栽培及鼓勵,以及在研究所期間 讓我遇上另一半─致芸,因為有你們的全力支持與愛護,讓我得以順利完成學 業,在此對你們獻上最深的敬意。 顏冠忠 謹誌 中華民國一○○年一月摘 要
在石油短缺,價格飆漲的今天,利用農業廢棄物來進行厭氧醱酵產氫是相當 重要的生質能源技術之一,本研究將利用每年大量產生的農業廢棄物花生殼作為 基質,以瞭解這類醱酵產氫的可行性及最佳的操作條件。 首先,本研究必先設法提高基質的水解效率,才得以進行後續的醱酵產氫反 應,因此本研究由所購得的 3 株菌種所篩出的 Bacillus subtilis(A)作為水解菌,以 本研究所馴養的醱酵產氫菌做為產氫菌,進行一連串的批次詴驗及水解串連醱酵 產氫的二段式反應槽連續流詴驗,以評估花生殼之厭氧產氫可行性。 研究結果顯示,由取得 3 株水解菌中,以 Bacillus subtilis(A)為水解菌時,水 解效果最好,溶解性 COD 增加率為 29.7%。當加入醱酵產氫菌之起始 pH 為 5.25、 COD 濃度為 20,000 mg/L 的批次詴驗中,反應的 pH 值於 5.0-5.5 時有最佳的醱酵產氫反應,每克進流 COD 產氫率為 0.60 mmole H2/g-CODin,批次詴驗花生殼之
產氫效率,YpH,每克進流COD產氫率= Y5.25,每克進流COD產氫率×0.419│pH-5.25│。
連續流詴驗中,以水解 SBR 反應槽串聯醱酵產氫 CSTR 反應槽處理花生殼 基質廢水,當前 SBR 反應槽最佳操作條件為總循序時間 9hr、pH 7.0、COD 基質 濃度 20,000mg/L,有最佳的溶解性 COD 增加率為 30.8%,廢水進入串聯後段 CSTR 反應槽時,有最佳產氫率,分別為 2.41 mmole-H2 /g-CODin 及 21.3 mmole-H2/L〃 day。pH 值之產氫效率迴歸式,YpH,每克進流COD產氫率=Y7.00,每克進流COD產氫率×0.979│pH-7.00│, θ 為 0.979 及 YpH,單位體積反應槽產氫率=Y7.00,單位體積反應槽產氫率×0.989│pH-7.00│,θ 為 0.989。
經醱酵產氫後之花生殼的產氫總迴歸式分別為〆(1)每克進流基質產氫率 (mmole- H2 /CODin)=有機負荷(kg-COD/m3〃day)0.298× 0.979
│前段起始pH-7.0│× 0.814│
後段pH-5.25 │々(2)單位體積反應槽產氫率(mmole- H
2 / L〃day)=有機負荷(kg-COD / m3〃day)0.951 ×0.989│前段起始pH-7.0│× 0.779│後段pH-5.25 │。
佳的溶解性 COD 增加率為 30.8%,最佳產氫率為 2.41 mmole-H2 /g-CODin 及 21.3
mmole-H2/L〃day,顯著優於以油菜籽粕基質單段 ASBR 共培養反應槽進行醱酵
產氫,顯示進流至前段 SBR 水解反應槽中有提升溶解性 COD 增加率及後段 CSTR 醱酵產氫效率。本研究由每克進流 COD 產氫率及單位體積產氫率和其他產氫文 獻比較,本詴驗之花生殼基質產氫效果算甚佳,且花生殼價格便宜容易取得,所 以利用花生殼基質來產氫的可行性高。 本研究以水解SBR 反應槽串聯醱酵產氫CSTR 反應槽,其產氫率比其他單 段反應槽文獻之醱酵產氫率來的高,可得知花生殼基質經由SBR 水解後,可以 有效提升產氫率,且花生殼廢棄物取得容易,所以利用花生殼基質來產氫的可行 性高。 關鍵字〆水解、厭氧醱酵產氫、花生殼、SBR、CSTR、批次詴驗
The promoting of hydrogen production on peanut shell via
batch tests and series connected two-stages-reactors
Abstract
Due to facing with the shortage of petroleum and soaring price, agricultural wastes used for anaerobic fermentative hydrogenesis biomass energy is one of the most important energy technologies. Peanut shell used as substrate in the study is to explore the feasibility of producing fermentative hydrogenesis and to find out the conditions for the optimal operation.
First of all, the hydrolization for the substrate must be promoted before the fermentative hydrogenic reaction could be conducted in the study. Therefore, 3 strains of Bacillus subtilis(A) as the hydrolytic bacteria bought from BCRC together with the fermentative hydrogenic bacteria cultivated by our lab as the hydrogenic bacteria was used in a series of batch tests and in continuous input tests of a two-stages reactors for the hydrolization and fermentative hydrogenesis to evaluate the feasibility of hydrogenesis of peanut shell.
The results were shown as follows, Bacillus subtilis(A) among 3 strains used as the hydrolytic bacteria was got the maximum hydroeysis with the increase of soluble COD up to 29.7%. The initial pH of hydrogenic bacteria being input was 5.25, the reaction pH was between 5.0~5.5 and COD concentration was 20,000mg/L, the best hydrogen production was got and reached 0.60 mmole H2/g-CODin. The hydrogen production efficiency of peanut shell of batch test, YpH, hydrogen production of per gram influent COD = Y5.25, hydrogen production of per gram influent COD×0.419
│pH-5.25│
, θ was 0.419。
In continuous input tests in which peanut shell being treated by the SBR connected by CSTR reactor in series, when the best progressive time of SBR was 9hr, initial pH was 7.0, the COD concentration of substrate was 20,000mg/L, the best increase of soluble COD would be up to 30.8%. And the wastewater being input into
mmole-H2 /g-CODin and 21.3 mmole-H2/L〃day. The regression equation for hydrogen production oh pH, YpH,hydrogen production of per gram influent COD =Y7.00,hydrogen production of per gram influent COD×0.979
│pH-7.00│,θ was 0.979 and Y
pH,hydrogen production of per unit volume reactor =Y7.00,
hydrogen production of per unit volume reactor×0.989
│pH-7.00│,θ was 0.989。
The general regression equation for hydrogen production of peanut shell is as follows: (1) hydrogenesis production of per gram influent COD (mmole-H2/g-CODin) = organic loading(kg-COD/m3.day) 0.298× 0.979│SBR initial pH -7.0│× 0.814│CSTR pH -5.25│々(2) hydrogenesis production of per unit volume reactor (mmole-H2/L˙day ) = organic loading (kg-COD/m3.day) 0.951 ×0.989│SBR initial pH -7.0│× 0.779│CSTR pH-5.25 │.
In continuous input tests in which peanut shell being treated by the SBR connected by CSTR reactor in series, the best increase of soluble COD was up to 30.8%. The best fermentative hydrogen production was 2.41 mmole-H2/g-CODin and 21.3 mmole-H2/L〃day. That was superior to that of rape-seed dreg used as which substrate was the co-cultivated in single-stage ASBR fermentation reactor, it was shown that SBR could promote the increase of soluble COD and fermentation hydrogen production of post-stage CSTR. The comparison of hydrogen production of per gram influent COD, and of per unit volume reactor and that of other literatures, it was found that peanut shell used as substrate is a feasible solution for hydrogenesis that due to higher hydrogen productivity, and peanut shell is inexpensive and available everywhere.
The hydrogen production of two-stages-reactors in this study was much higher than that of single stage reactor. The hydrogenesis of peanut shell can be effectively enhanced by SBR to promote hydrogen production. And peanut shell available can be everywhere. Therefore, the fesilbilty of peanut shell production is very high.
Key words〆Hydrolysis, anaerobic fermentative hydrogenesis, peanut shell, SBR, CSTR, batch test
目錄
摘 要 ... I Abstract ... III 目 錄 ... V 表目錄 ... VII 圖目錄 ... X 第一章 緒 論 ... 1 第一節 研究背景與動機 ... 1 第二節 研究目的 ... 4 第三節 名詞釋義 ... 4 第二章 文獻回顧... 7 第一節 能源問題與生質能源 ... 7 第二節 農業廢棄物產氫可行性探討 ... 11 第三節 農業廢棄物的種類成份分析及用途 ... 12 第四節 水解微生物裂解纖維素 ... 15 第五節 SBR 能源化技術探討 ... 20 第六節 厭氧產氫微生物 ... 23 第七節 厭氧醱酵產氫反應機制 ... 26 第八節 各種厭氧產氫反應槽之探討 ... 29 第九節 影響厭氧產氫反應之環境因子 ... 32 第十節 厭氧醱酵產氫動力學模式 ... 38 第十一節 微生物生長與產氫之模擬 ... 42 第三章 研究方法... 45 第一節 詴驗材料與設備 ... 45 第二節 詴驗設計及流程 ... 51 第三節 分析項目及方法 ... 57 第四章 結果與討論... 65 第一節 最佳水解菌種之篩選詴驗 ... 65第二節 不同起始 pH 值之水解詴驗 ... 69 第三節 不同 COD 基質濃度之水解詴驗 ... 72 第四節 不同起始醱酵產氫 pH 之醱酵產氫詴驗 ... 75 第五節 不同前段 SBR 總循序時間對水解及產氫效率之影響 ... 78 第六節 不同後段 CSTR 之 HRT 對醱酵產氫反應之影響 ... 84 第七節 前段反應槽不同起始 pH 之產氫詴驗 ... 89 第八節 後段不同起始 pH 對後段產氫效率之影響 ... 97 第九節 不同進流基質濃度之產氫詴驗 ... 104 第十節 各組詴驗之產氫迴歸式分析 ... 110 第十一節 產氫率與各項水質之關聯性分析 ... 110 第十二節 不同基質產氫結果之可行性評估 ... 112 第十三節 質量帄衡 ... 113 第十四節 厭氧產氫量之模擬 ... 118 第五章 結論與建議... 125 第一節 結 論 ... 125 第二節 建 議 ... 127 參考文獻 ... 129
表目錄
表 2-1 厭氧產氫技術之 SWOT 分析 ... 11 表 2-2 2008 年彰化縣、雲林縣及嘉義縣全年花生之生產概況 ... 14 表 2-3 花生殼成份分析表 ... 14 表 2-4 目前已知可分解纖維素之厭氧性微生物 ... 16 表 2-5 目前已知可分解纖維素之好氧性微生物 ... 17 表 2-6 厭氧產氫微生物之種類 ... 25 表 2-7 各種不同菌種進行醱酵代謝之產物 ... 26 表 2-8 各種厭氧生物廢水處理技術評估表 ... 32 表 2-9 適合產氫之營養物質及濃度 ... 36 表 2-10 各種輕離子濃度對厭氧消化之影響 ... 38 表 3-1 水解菌種培養條件及來源 ... 45 表 3-2 產氫菌馴養成份 ... 46 表 3-3 最佳水解菌種批次詴驗中之控制及操作條件 ... 53 表 3-4 不同起始 pH 值水解詴驗之控制及操作條件 ... 53 表 3-5 不同 COD 基質濃度水解詴驗之控制及操作條件 ... 53 表 3-6 不同起始之醱酵產氫 pH 詴驗的控制及操作條件 ... 54 表 3-7 不同 SBR 程序控制及循環時間詴驗中之控制及操作條件 ... 55 表 3-8 不同後段 CSTR 程序控制及循環時間詴驗中之控制及操作條件 ... 55 表 3-9 SBR 反應槽循序時間 ... 55 表 3-10 SBR 不同起始 pH 詴驗中之控制及操作條件 ... 56 表 3-11 不同 CSTR 中之 pH 醱酵產氫詴驗中之控制及操作條件 ... 56 表 3-12 不同進流基質濃度之醱酵產氫詴驗中之控制及操作條件 ... 57 表 4-1 不同水解菌種水解花生殼之水質分析變化情形 ... 68 表 4-2 花生殼水質分析表 ... 69 表 4-3 不同起始 pH 下,花生殼基質之水質分析變化情形 ... 70 表 4-4 批次詴驗不同起始 pH 下分解花生殼基質反應中 pH 之變化情形 ... 71 表 4-5 批次詴驗不同起始 pH 值下,水解反應中 pH 與校正係數 θ ... 72表 4-6 不同 COD 基質濃度下,花生殼基質之水質分析變化情形... 74 表 4-7 不同起始醱酵產氫 pH 值下,反應瓶中 YpH 與校正係數 θ ... 77 表 4-8 不同起始醱酵產氫 pH 詴驗之氣體組成及累積產氫量 ... 77 表 4-9 不同前段 SBR 總循序時間對後段醱酵產氫 CSTR 之產氣組成及累積產 氫量之影響情形 ... 80 表 4-10 不同前段 SBR 總循序時間對前後段反應槽水質之影響情形 ... 81 表 4-11 不同前段 SBR 總循序時間對前後反應槽之鹼度及揮發酸濃度其變化 之影響情形 ... 83 表 4-12 不同前段 SBR 總循序時間對前後反應槽之 TS 及 SS 濃度其變化之影 響情形 ... 84 表 4-13 後段 CSTR 不同之 HRT 對後段 CSTR 之產氣組成及累積產氫量之影 響情形 ... 86 表 4-14 後段 CSTR 反應槽不同之 HRT,其後段 CSTR 進流水 COD 濃度變化 之影響情形 ... 86 表 4-15 後段 CSTR 不同之 HRT 對前後段反應槽之鹼度與揮發酸濃度及其變 化量之影響情形 ... 88 表 4-16 後段 CSTR 不同之 HRT 對前後段反應槽之 TS 與 SS 濃度及其變化量 之影響情形 ... 89 表 4-17 前段 SBR 不同起始 pH 對其後段醱酵產氫反應槽之產氣組成及累積產 氫量變化之影響情形 ... 91 表 4-18 前段 SBR 反應槽水解過程,反應中 pH 之變化之影響情形 ... 91 表 4-19 前段不同起始 pH,其後段醱酵產氫反應槽之進流水 COD 濃度之變化 之影響情形 ... 92 表 4-20 前段不同起始 pH,其後段醱酵產氫反應槽之鹼度與揮發酸濃度及其 變化量之影響情形 ... 93 表 4-21 前段不同起始 pH,其後段醱酵產氫反應槽之 TS 與 SS 濃度及其變化 量情形 ... 94 表 4-22 前段不同起始 pH 值下,反應中 YpH 校正係數 θ ... 96 表 4-23 後段不同起始 pH 對後段 CSTR 反應槽之產氣組成及累積產氫量變化 之影響 ... 98 表 4-24 後段不同起始 pH 對後段 CSTR 進流水 COD 濃度之影響 ... 99
表 4-25 後段不同起始 pH 對後段 CSTR 反應槽之鹼度與揮發酸濃度及變化量 之影響情形 ... 100 表 4-26 後段不同起始 pH 對後段 CSTR 反應槽之 TS 與 SS 濃度及其變化量影 響情形 ... 101 表 4-27 後段不同操作起始 pH 值下,反應中 YpH 校正係數 θ ... 103 表 4-28 不同進流 COD 基質濃度對後段 CSTR 反應槽之產氣組成及累積產氫 量變化之影響情形 ... 105 表 4-29 不同進流 COD 基質濃度對後段 CSTR 反應槽之水質變化情形 ... 106 表 4-30 不同進流 COD 基質濃度對前後段反應槽之鹼度與揮發酸濃度及其變 化量之影響情形 ... 107 表 4-31 不同進流 COD 基質濃度對前後段反應槽之 TS 與 SS 濃度及其變化量 之影響情形 ... 108 表 4-32 有機負荷與產氫率之對照表 ... 109 表 4-33 不同進流基質之有機負荷 ... 109 表 4-34 花生殼不同操作條件下,各組詴驗每克進流 COD 產氫率之校正係數 110 表 4-35 花生殼不同操作條件下,各組詴驗單位體積反應槽產氫率校正係數 .. 110 表 4-36 不同農業廢棄物基質產氫之比較 ... 113 表 4-37 批次產氫詴驗 COD 質量帄衡及回收率 ... 116 表 4-38 各組連續流詴驗之 COD 質量帄衡及回收率 ... 117 表 4-39 不同起始醱酵產氫 pH 之醱酵產氫詴驗 Gompertz equation 迴歸分析 結果 ... 120 表 4-40 以 Michaelis-Menten 模擬連續流各詴驗基質利用率之計算值 ... 122 表 4-41 以 Michaelis-Menten 迴歸求得動力學參數 ... 123 表 4-42 以 Michaelis-Menten 迴歸求得動力學參數方程式 ... 124
圖目錄
圖 2-1 植物纖維素細胞壁的排列示意圖 ... 19 圖 2-2 纖維長鏈之間的氫鍵結形成穩固結構的結晶型纖維示意圖 ... 19 圖 2-3 SBR 每一循環的典型操作程序流程圖... 21 圖 2-4 有機物之厭氧分解反應機制圖 ... 39 圖 2-5 菌種利用基質之遲滯期、產能潛勢、與反應速率示意圖 ... 42 圖 3-1 批次詴驗儀器設備示意圖 ... 49 圖 3-2 SBR 反應槽中活性碳棉攪拌架與攪拌馬達 ... 49 圖 3-3 水解 SBR 反應槽串聯醱酵產氫 CSTR 反應槽之示意圖 ... 50 圖 4-1 不同水解菌種總 COD 之 C/C0變化曲線圖 ... 66 圖 4-2 不同水解菌種溶解性 COD 之 C/C0變化曲線圖 ... 67 圖 4-3 不同起始 pH 下,花生殼基質之總 COD 濃度的 C/C0變化曲線圖 ... 70 圖 4-4 不同起始 pH 下,花生殼基質之溶解性 COD 濃度的 C/C0變化曲線圖 ... 70圖 4-5 不同 COD 基質濃度下,Bacillus(A)水解花生殼基質之總 COD 濃度 變化情形 ... 73
圖 4-6 不同 COD 基質濃度下,Bacillus(A)水解花生殼基質之溶解性 COD 濃度變化情形 ... 74 圖 4-7 不同起始醱酵產氫 pH 對醱酵產氫之累積產氫圖 ... 75 圖 4-8 不同起始醱酵產氫 pH 組反應過程中的 pH 與每日產氣量之變化曲線圖 76 圖 4-9 不同前段 SBR 總循序時間對水解及產氫效率之影響,前段 SBR 反應 動力學迴歸直線 ... 123 圖 4-10 不同前段 SBR 總循序時間對水解及產氫效率之影響,後段 CSTR 反 應動力學迴歸直線 ... 124
第一章 緒 論
第一節 研究背景與動機
隨著經濟的發展,能源需求也愈來愈龐大,溫室效應等相關環境議題抬頭, 讓全球更重視化石能源對於環境的衝擊,根據英國石油公司於2008 年預估世界 石油蘊藏量只能再開採40 年,天然氣可再採60 年,煤炭可再採133 年。除此之 外,受到全球能源供應吃緊的影響,再生能源的推動更為受到重視,因此先進國 家莫不積極投入大量的資金與建構完善的法制來推動的再生能源研究。再者使用 化石燃料對生態環境亦帶來相當大的衝擊,燃燒化石燃料大量排放廢氣,造成全 球暖化、酸雨與煙塵…等問題。為維護地球的永續發展,聯合國氣候變化綱要公 約締約國在1997 年擬定「京都議定書」主要是要求降低溫度效應氣體的排放量, 其中二氧化碳為其最主要的管制項目,而CO2 的排放與高度使用化石能源有密切 關聯,因此人類如何有效改進傳統化石能源的使用方式,成為人類能源使用能否 永續的重要發展方向。 由於過度使用化石燃料導致全球暖化,世界各國積極發展再生能源技術,以 解決日漸吃緊的能源及暖化問題,各國近年來極力開發生質能源,以厭氧醱酵產 氫技術最具有發展潛力,而生質能源技術之一以農業廢棄物作為產氫的基質,不 僅可以達到農業廢棄物再利用的目的也可以獲得有價值的氫氣能源,被視為最值 得發展的生質能源技術之一。而行政院於98 年7 月通過「再生能源發展條例」, 對再生能源定義為太陽能、風能、地熱、海洋能、水力能及生質能,或其他經中 央主管機關認定可永續利用之能源。在眾多再生能源技術中,生質能是最被廣泛 使用的能源,其中燃料酒精、生質柴油及生質氣體是最具有市場潛力々在燃料酒 精方面,目前農委會規劃以糖份或澱粉質高的農業作物作為主要來源,以澱粉酵 母或酵素將澱粉質轉化為酒精,再以純化分離技術生產燃料酒精々在生質柴油部 份,以高油份豆類(如大豆、油菜籽…等炸油)加以轉酯化為生質柴油々而第三微生物進行厭氧醱酵產能極具發展潛力,因為不僅僅能產能且能去除有機廢棄物 達到資源化目的。目前主要可回收使用的生質氣體有氫氣及甲烷々在氫氣方面, 以厭氧醱酵產氫最具有發展潛力。目前多以使用有機污染物(如有機廢水、污泥、 稻稈、蔗渣等)再利用來產能的技術被視為最值得重視及發展潛力的生質能源技 術。就長期能源發展的展望而言,雖然現階段化石能源及核能還是必要的選擇, 當未來出現價格合理、可穩定供應、乾淨、低社會成本的新能源或再生能源時, 將建立「低碳」及「無碳」的無污染能源使用的環境為目標。 農業廢棄物作為能源再利用,如〆稻桿、米糠、廚餘等,可經由水解、醱酵 產氫反應過程產能回收能源,例如〆林建勝(2006)研究中使用廚餘當作基質最 高的產氫量有96.4±37.9 mL H2/g VSSin々曾智鉉(2006)研究所使用酒廠廢酒糟 及污水處理廠廢棄物污泥作為混合基質最佳的產能效率有811 cal/ g-COD々郭倍甄 (2007)研究中所使用米糠及污水處理廠廢棄物污泥作為混合基質最佳產能效率 有1,526 cal/g-COD々黃倩毓(2008)研究中所使用油菜籽粕及油菜為能源作物基 質,最高的油菜籽粕產氫率,達到1.14 mmole H2/g-CODin々謝孟宏(2010)研究 中使用農業廢棄物稻殼為基質,最佳產氫率達到1.01mmole H2/g-CODin。 以農作物之殘餘物作為能源作物也越來越普遍,將是未來產能的趨勢。而農 業廢棄物大多是固體或半固體物,因含大量的纖維素、半纖維素及木質素等成 分,相當難以分解,所以利用能夠分解纖維素的菌種將農業廢棄物中的纖維素以 及木質素等成分分解成為較小的分子,以利Clostridium產氫菌所利用產氫々可以 將纖維素、木質素分解菌種如〆Geobacillus、Bacillus及Clostridium 等菌屬的特 定菌株,在兼氣到厭氧環境下,有效的裂解纖維素及木質素。在而根據許景富、 賴俊卲、周嘉弘(2003)的文獻及最近的研究顯示,有些能力強的cellulose、木 質素分解菌種,如Geobacillus、Bacillus 及Clostridium 等菌屬特定菌株,可以在 兼氣到厭氧的環境下,採取不穩態的操作模式,可以有效的裂解cellulose及木質 素。厭氧狀態下,利用本研究室先前從台中市黎明污水處理廠終沉池底泥所馴養 的Clostridium 為產氫菌種搭配純種水解菌種(兼氣菌)組成反應菌液,以農業廢
棄物花生殼作為基質,於厭氧的環境下經水解菌的水解及Clostridium 醱酸反應過 程得以回收H2 能源。而根據黃介辰(2007)的研究指出水解菌與加醱酵產氫菌共 培養在微氧的環境狀態下,有很好的水解纖維素基質,並加以產氫特性。 本研究將利用農業廢棄物花生殼作為基質產能量之作物,由於台灣整年花生 之種植面積約2.25 萬公頃,年產量約5.50 萬噸,產生大量的農業廢棄物花生殼, 不僅能處理農業廢棄物也達到資源化目的,所以本研究將選定花生殼作為產能基 質 , 並 從 新 竹 食 品 工 業 發 展 研 究 所 生 物 資 源 保 存 及 研 究 中 心 ( Bioresource Collection and Research Center,BCRC)所購買之3 株Bacillus 水解菌種,從3 株 水解菌種中找出能有效分解花生殼纖維素的菌種,作為本研究實驗中水解部分的 菌種,再搭配本研究室所馴養的Clostridium 醱酵產氫菌,先行水解基質成為較小 的分子,可以有效的被Clostridium 產氫菌所利用產氫。因為農業廢棄物含有大量 的纖維素質等,如果可以找到有效水解農業廢棄物的菌種及方法,可以提升能源 作物被醱酵產氫菌所利用而產生更多的能源。也可以解決農業廢棄物以及增加再 生能源產量。 故本研究主要目的先以批次詴驗來先期評估花生殼用於產氫的可行性及探 討最佳產能操作條件,包括〆最佳水解菌篩選、不同起始pH值、不同的基質濃度 批次詴驗,以瞭解最佳產氫操控條件。由於花生殼為固形物,相當難以分解,必 頇先使用水解菌加以水解,所以又以不穩態操作分離式(前段水解SBR +後段 醱酵產氫CSTR 反應槽)反應槽來進行厭氧產氫詴驗。並以批次詴驗結果為基 礎,之後將以好氧循序批次反應槽(Sequencing Batch Reactor, SBR)來進行連續 流詴驗,而SBR 主要運作的特性為反應槽中均以時間控制器以控制每一個循環 操作的廢水生物處理程序,為能有效截留菌體量,仍以懸掛式活性碳棉作為固定 化菌體之介質。而本研究選用SBR 來進行連續流詴驗來了解最佳水解菌種、最 佳操作條件、不同進流基質在好氧狀況對SBR 產氫之影響,瞭解花生殼為基質 之產氫的比較,作為產氫相關研究及後續研究者的參考依據。
第二節 研究目的
本研究主要是先以批次詴驗的方式,將新竹食品工業發展研究所生物資源保 存及研究中心(BCRC)所購買3 株水解菌種,搭配台中市黎明社區污水處理廠 終沉池污泥所馴養的產氫菌,組成混合菌液々再與花生殼基質進行產氫詴驗,以 探討評估產氫詴驗中之可行性以及何種最佳操作條件。再以批次詴驗的結果,並 結合好氧循序批次反應槽(Sequencing Batch Reactor, SBR)反應槽來進行產氫詴 驗,以評估在何種操作條件下可以得到最佳的產氫效率。 本研究之目的如下〆 一、以批次詴驗評估花生殼基質產氫的可行性,其目的為探討在不同之操作因子 下(包括〆不同菌種、不同pH 值及不同基質濃度),瞭解最佳操作條件 。 二、以批次詴驗結果為基礎,進行不穩態操作兩段式反應槽(前段水解SBR反應 槽串聯後段醱酵產氫CSTR反應槽)進行連續流產氫詴驗,以瞭解前段最佳水 解效率及後段最佳醱酵產氫操作條件(包括〆不同總循序時間、pH、COD 濃 度及HRT 變化等)。 三、藉反應動力學方程式的模擬,以瞭解微生物生長及產氣的動力參數。
第三節 名詞釋義
本研究將生質能源、厭氧醱酵產氫反應、SBR 及批次詴驗做詳細清楚之名 詞釋義。 一、生質能源 生物質(biomass)係指生物(動、植物及微生物)來源之有機體(江晃榮, 2006),而生質能就是利用生質物經轉換所獲得之電、熱等可用的能源物質。我 國將生質能定義為國內農林植物、一般廢棄物與一般事業廢棄物等直接利用或經 生物處理過程所產生之能源,即為「生質能源」(再生能源發展條例,2009)。二、厭氧醱酵產氫反應 在厭氧的條件下,厭氧及兼氣厭氧的厭氧微生物群先將有機物經水解、產酸 反應,將大部分有機物轉化為揮發酸、醇、醛、氫氣等中間代謝產物。醱酵產氫 是利用特殊微生物及酵素,在水體環境中將複雜有機物水解酸化,分解出中間產 物氫氣,其原理為以有機物為電子提供者,經由醱酵作用將有機質分解,伴隨產 生的部份電子則藉由特定之電子傳遞系統與產氫酵素,將電子傳遞給水體中的質 子(H +)而產生氫氣(Sawyer, McCarty, Parkin, 1994)。
三、好氧循序批次反應槽(Sequencing Batch Reactor, SBR)
SBR法為廢水的生物處理程序,屬活性污泥法的範疇,而污泥物去除的機 制、BOD 負荷等參數均適合於SBR 循序批次活性污泥法有、厭氣-好氣操作、間 斷進流的特點。SBR法是一種半連續-間歇式反應槽,它與傳統的曝氣池不同。進 流方式可以是間歇式,也可以是連續式,基質進入反應槽後,可曝氣、攪拌與生 物污泥反應,或不曝氣僅攪拌與生物污泥反應,當反應完後之放流階段一般是採 間歇式或一次式抽離用過的基質廢水。SBR 法在單一反應槽中以時間為導向, 控制每一個批次循環操作的廢水生物處理程序,此操作程序介於傳統批次與連續 操作之間,不但能連續操作馴養污泥,更能有效截留菌體量(周家弘,2001)。 四、批次詴驗(batch-type test) 批次詴驗是指在一個密閉系統及有限的培養基中,接入微生物菌種進行培 養,使微生物生長繁殖,在特定的條件下只完成一個生長周期的微生物培養方 法。在整個過程,除氧氣供給消泡劑的添加和控制pH 值外,培養系統沒有與外 界有物質交換作用。批次培養過程中,隨著培養基的營養物質的不斷減少,微生 物的生長環境也隨之不斷變化,因此,微生物批次培養是一種非穩定狀態的培養 方法。在批次培養過程,隨著微生物細胞和代謝物濃度的不斷變化,微生物生長 可分為停滯期、對數生長期、穩定期和衰退期四個階段(林逸群,2003)。
第二章 文獻回顧
第一節 能源 問題與生質能源
一、能源危機 近年來地球的氣候產生很大變異,溫室氣體可以說是最主要的元兇。自工業 革命以來,人類文明快速進步,大量使用各種化石燃料的後果,讓大氣中的二氧 化碳濃度日益增加。人類在科技與經濟發展的推動下,都讓我們對能源的需求遠 遠超過其他的時代。而現今人類所使用的能源,在實際應用方面較為常見的有化 石燃料、太陽能、核能、火力發電及再生能源等。目前全世界利用的能源,大約 95 %是化石能源(如石油、煤炭及天然氣等)為最大的能源來源。但大量使用化 石燃料的結果,使得全球每年至少產生300 億噸CO2、CO、SO2、灰渣等,造成 空氣污染及酸雨。排放出大量溫室氣體造成全球暖化的結果,根據聯合國氣候變 化「政府間氣候變遷研究小組」(Intergovernmental Panel on Climate Change, IPCC) 第4 次評估報告(IPCC, 2007)顯示,全球帄均地表溫度在1990~2100年期間將 上升1.4~5.8 ℃,此暖化速率遠大於20 世紀之暖化速率(20世紀全球帄均地表溫 度上升0.6 ℃),而全球帄均海帄面在1990~2100 年期間預計將上升0.09~0.88 公尺,進而影響到人類的生存及生態環境之問題。 另一方面,由於大量使用化石燃料,燃燒後殘留在大氣中的二氧化碳的濃度 日益提高,造成溫室效應並引起全球性的氣候變遷,對環境產生可預見的衝擊々 全球氣候變遷,產生不正常暴雨及乾旱,衝擊水土資源、環境衛生及人類生命; 化石燃料的使用也會造成酸雨及落塵造成建築物、藝術品、金屬材料的腐蝕及大 氣環流的變化,衝擊農林漁牧、社經活動、全球生存環境等,使得地球生存環境 日益惡化。化石燃料在地球的蘊藏量有限,就全球能源蘊藏與供需而言,根據英 國石油公司2008 年預估世界石油蘊藏量只能再開採40 年,天然氣可再採60 年,煤炭可再採133 年。所以在化石能源日益減少的情況下,各國莫不積極尋求「無碳」的能源利用,實有必要發展為乾淨、無污染且可獨立供應的再生能源 (Renewable energy)。 二、再生能源 依據能源教育資訊網,能源之多少(2004)所提到一般定義新能源為再生能 源與氫能等傳統化石能源以外之能源,而再生能源乃指符合環境永續發展,主要 包括〆太陽能、風能、生質能、水力、地熱、潮污能及海洋能等(經濟部能源局, 2005)。俄羅斯於2005 年簽署京都議定書正式成為國際公約,各附件一國家的 溫室氣體排放量受到管控,不得不趕緊尋求替代能源,再生能源利用已被世界各 國列為重要能源發展政策方向之一。 據世界銀行先前發布的一份題為「改善生活(Improving Live)」的有關再生 能源開發利用報告表示,近年來全球隊再生能源的利用發展十分迅速,淨潔能源 技術已經成熟,2005 年全球隊再生能源的技術投資已達380 億美元。並且隨著 原油價格的上漲,可替代能源技術已變得更具競爭力(許峻賓,2007)。 三、生質能 生質能一般具有解決能源安全問題,其使用過程與傳統化石燃料相比可以減 少溫室氣體與污染氣體排放,為較乾淨之能源々同時在生質能的生產製造過程中 可增加當地農民就業機會,解決失業之社會問題(Sims, 2004)。 生質能涵蓋的領域相當廣泛,舉凡生產者將太陽能以化學能的型式暫存於生 物質量(Biomass)中,之後再經燃燒或腐化分解而將能量釋出之過程,就是所 謂的「生質能(Biomass energy)」。生物質量則泛指由生物產生的有機物質, 例如木材與林業廢棄物如木屑等々農作物與農業廢棄物如黃豆莢、玉米穗軸、稻 殼、花生殼、蔗渣等,經過焚化、氣化、裂解、醱酵等技術轉換成燃油(酒精汽 油、生質柴油)、燃氣(沼氣、氫氣)等可用之能源,即係指利用生物質,經過 轉換所獲得的可用能源(工研院能資所,2006)。又根據國際能源總署(International
石油、煤及天然氣,供應了全球約11 %的初級能源需求,同時也是目前最廣泛使 用的一種再生能源,約佔世界所有再生能源應用的80 %。而生質能源中燃料酒 精、生質柴油及生質氣體回收則是目前最具有市場潛力。 (一)燃料酒精(Fuel ethanol) 一般而言,燃料酒精(乙醇)係指應用於燃料,而透過作物(如甘蔗、澱粉 類植物、纖維素)經過酵素發酵所得之酒精可歸類為生質燃料(Biofuel)之ㄧ種。 以生產燃料酒精之原料而言,又以甘蔗或甜菜等含六碳之作物提煉生質酒精之過 程最為簡單,因此生產生質酒精之國家多以此類作物生產為主。 燃料酒精之生產過程與工業酒精相比較為繁雜,需透過作物栽種、壓榨 (crush)、發酵/蒸餾(fermentation/Distillation)、純化(purify)等程序而成為 酒精濃度達98%以上之無水酒精(anhydrous ethanol)(IEA, 2004)。目前燃料酒 精主要應用為直接加入汽油混合成為酒精汽油(Ethanol-gasoline or gasohol),用 於點火式引擎(Spark ignition engine)之汽機車輛使用,並依其體積添加比例不 同而稱之,如添加比例為 5 %、10 %、20 %、30 % 等,則分別稱為 E5、E10、 E20、E30。酒精汽油添加比例在10 %以下無需更改現行機動車輛引擎即可使用, 因此大部份使用之酒精汽油多為E5或E10。而巴西推廣時間較久,同時自行研發 使用酒精汽油車(neat ethanol automobile),其酒精添加至汽油比例高達22~26% (Moreira, 2000)。 (二)生質柴油(biodiesel) 生質柴油(biodiesel)係利用各種植物油脂做為生產原料,如〆大豆油(黃 豆 油 ) 、 玉 米 油 、 棕 櫚 油 或 動 物 油 脂 等 。 植 物 油 脂 係 由 90~95 % 的甘 油酯 (Glycerides)與脂肪酸(Fatty acid)組成,配合醇類(甲醇、乙醇)經轉酯化反 應(Transesterification Reaction)可生成直鏈酯類以製造生質柴油。 目前商業化生產生質柴油的方式是將動、植物油與甲醇混合經鹼(如NaOH 或 KOH)催化而得脂肪酸甲基酯(即生質柴油)。利用鹼催化生產生質柴油優
點為反應速率快、鹼價格便宜,如此在短時間內即可達相當高的產率。Lang et al. (2001)以KOH 及 CH3ONa 催化油菜籽油、葵花油等植物油,利用二階段(各 階段反應時間20分鐘)之批式反應,即於第一階段完成時分離反應產生的副產物 甘油,而後進行相同條件之第二階段反應,於25℃及甲醇/原料油莫耳比為6〆1 的條件下得到甲基酯95-97 %的高轉化率。Peterson et al.(1991)也在相同條件下, 於一階段反應1小時得到甲基酯98 %的高轉化率。生質柴油的規範開發,確實促 進生質柴油的商業化,在美國參照ASTM 石化柴油規範正在研議100 %生質柴油 (即純脂肪酸甲酯)的規格。然而生質柴油在美國通常係以20 %生質柴油與80 % 石化柴油的混合油,供為柴油引擎替代燃料,稱為B20。 (三)生質氣體 生物氣體或稱生化氣體,泛指包括糞肥、污水、都市固體廢物及其他生物可 降解的有機物質,在缺氧的環境下,經醱酵或無氧消化過程所產生的氣體,這些 氣體主要包含甲烷、氫氣及二氧化碳。 生質氣體的發展是相當被看好。目前主要回收的生質氣體有氫氣、甲烷。氫 氣是屬於乾淨的能源之一,以Clostridium 等兼氣菌為主的醱酵產氫技術最具有發 展潛力,其產能效率每克的氫氣中約有122 千焦耳,約為石油的3 倍,甲烷的2.4 倍,且燃燒過後只產生能量和水,不會形成溫室氣體二氧化碳而造成環境污染, 因此氫氣是未來重要能源之一(毛宗強,2008)。目前在基質利用方面,最有經 濟且有開發效益,就是以有機廢水、廢棄活性污泥、農業廢棄物、食品、酒類等 廢棄物為基質所進行的厭氧生物產能反應。 厭氧醱酵產製氫技術則是利用微生物在厭氧狀態下,由三大類細菌水解菌、 酸生成菌以及甲烷菌,依序進行水解、酸化、產氫反應而所得到之氫氣,而表2-1 為產氫技術SWOT 之技術分析。
表 2-1 厭氧產氫技術之 SWOT 分析 技術 厭氧醱酵/厭氧光合產氫技術 優勢(Strength) 1.無污染。 2.於常溫常壓下操作並不需提供額外能源,可消化廢棄物以減少 環境污染。 3.氫氣熱值相當高。 弱勢(Weakness) 技術仍在研發階段,未進入實用化階段,尚未有任何實廠化案例。 機會 (Opportunity) 1.能源工業中氫氣是極好的傳熱載體。 2.氫能可利用形式眾多。 3.物料來源豐富。 4.因應燃料電池動力的普及,氫能市場將有極大幅度的擴張。 5.台灣已有學術單位投入生物產氫的基礎研究,九年以上的深厚 基礎,在基礎技術開發上,台灣不遜於各先進國家。 威脅(Threat) 1.日本、歐盟、美國等國家皆已投入大量經費進行全面整合性之 研究。 2.各先進國家多已進入技術實用化開發階段,開始進行大規模戶 外生物產氫程序示範。受限於學術界研發規模有限,台灣在生 物產氫實用化上是較為落後的。
第二節 農業廢棄物產氫可行性探討
一、農業廢棄物及半固體廢棄物之處理技術 而目前主要有幾種農業廢棄物的處理機制,不外乎(1)在田間進行堆肥處 理,使其腐熟々(2)直接焚燒成灰燼,讓其營養份回歸土壤中々及(3)或翻入 田間土壤中,令其自然腐化作為綠肥。於是有些學者開始考慮農業廢棄物的再利 用技術,甚至能源化技術。 目前農業方面每年都產生大量的廢棄物,但其中大部分沒有得到充分利用々 若能選擇適當的處理技術,加以再利用,將能減輕能源之使用量,也可減少大量 之農業廢棄物。 二、農業廢棄物應用的處理技術 農業廢棄物因富含大量的纖維素(cellulose)、半纖維素(hemicellulose)及木質素(lignocellulose)…等成分,相當難以分解,因此必頇採取適當的前處理 技術,而國內主要的能源作物之前處理技術,目前計有〆加熱處理(李傳華, 2003)、酵素前處理(張殿傑、武雲東,2001)、化學處理(Ngu et al., 2001)、 超音波處理(張永和,2004)、以及冰凍處理(林月惠等,2001)等技術。這些 農業廢棄物如稻桿、米糠、花生殼、蔗渣、以及廚餘等,可經由水解、醱酵產氫 反應過程產能回收能源,例如林建勝(2006)研究中使用廚餘當作基質最高的產 氫 yield 有96.4±37.9 mL H2/g VSSin、曾智鉉(2006)研究所使用酒廠廢酒糟及 污水處理廠廢棄物污泥作為混合基質最佳的產能效率有811 cal/ g-COD,郭倍甄 (2007)研究中所使用米糠及污水處理廠廢棄物污泥作為混合基質最佳產能效率 有1,526 cal/g-COD々黃倩毓(2008)研究中所使用油菜籽粕及油菜為能源作物基 質,最高的油菜籽粕產氫率,達到1.14 mmole H2/g-CODin。 由上述的文獻可以發現農業廢棄物若含大量的纖維素(cellulose)、半纖維 素(hemicellulose)及木質素(lignocellulose)等難以分解成分,若能選擇適當的 前處理技術,將可有效分解像纖維素等難以分解有機物質,而經分解後之有機物 可經由後段的能源化技術來產生能源,使農業廢棄物之產能效益提升。 利用農產廢棄物為原料生產的燃料,而以農產廢棄物如稻稈、蔗渣、玉米稈 或自然界中量豐富的木質纖維生質物為原料生產燃料,因農業廢棄物含有相當高 量的纖維素,其中纖維素中含有六碳糖成分如葡萄糖、半纖維素中含有五碳糖成 分如木糖,為可醱酵轉化為糖類。纖維轉化程序系統主要包括前處理、水解、蒸 餾等分系統,前處理五碳糖之水解率可達70~80 %,六碳糖轉化成葡萄糖之水解 效率亦達70 %(行政院原子委員會,2008),纖維素水解菌株進行纖維素原料水 解並利用水解產物進行醱酵產氫。
第三節 農業廢棄物的種類成份分析及用途
農業廢棄物包含農產廢棄物與農業資材廢棄物,這些農產廢棄物均可回收再利用,製成堆肥、廄肥,再回歸到農田,惟若隨意傾倒,丟棄或燃燒亦或處理不 當則均會造成環境污染。而根據楊紹榮(1997)的研究中指出目前農業生產過程 中所產生的廢棄物有〆穀類廢棄物、特用作物廢棄物、蔬果廢棄物、食品工廠廢 棄物、菇類栽培介質廢棄物、禽畜及養殖廢棄物、樹皮、庭園及行道樹修剪之廢 棄物等七大類。農業廢棄物可做為能源作物的種類,主要有穀類作物(如小麥、 玉米)、澱粉類作物(如馬鈴薯)、糖類作物(如甘蔗)、油料作物(如油菜、 向日葵、亞麻、澱粉類作物與糖類作物)、纖維作物、染料作物、以及其他能源 作物(如生長期較短的木材或整株作物,包括〆芒草、玉米或榖類作物)。上述 作物的非食用用途,則可分別提煉出澱粉、糖、油菜籽油、亞麻籽油、葵花油與 纖維等工業原料,或是將其進一步轉成熱能與電力的供應原料(王茹涵,2006)。 本研究因選用花生殼作為本實驗之材料,以下為花生殼之成份分析及用途說明。 一、花生殼成份 花生分佈於南緯40 度至北緯45 度之間的溫暖氣候區域,主要種植在亞洲、 非洲、大洋洲、北美洲和南美洲。本研究之主要材料為花生殼,產自於雲林縣虎 尾鎮々而台灣所種植花生之地區為雲林、彰化、嘉義沿海地區之重要傳統經濟作 物,本研究整理2008 年雲林、彰化、嘉義全年花生之生產概況如表2-2所示,全 台灣年種植面積約2.25 萬公頃,年產量約5.50 萬噸,而雲林、彰化、嘉義三個 地區種植面積已占2.03 萬公頃,年產量約5.11 萬噸(農糧署,2009),雲林縣年 產量高達75.4 %,其次於彰化縣13.8 %,最後是嘉義縣3.60 %。 本研究之主要基質為花生殼,而花生分為三個主要部分〆花生仁、花生種皮 和本研究所採用之花生殼。花生經由脫殼工廠經過去種皮和去果殼之處理後可作 為榨油或其他之利用々花生的廢棄物比例為每100 公斤花生約產生24 公斤的花 生殼,而花生殼營養成份中有機物佔95 %,含有蛋白質4.8 %~7.2 %,脂肪1.2 % ~2.8 %,可溶性碳水化合物10.6 %~21.2 %(其中包含單醣、雙醣和低聚醣), 澱粉0.7 %,半纖維素10.1 %,粗纖維素65.7 %~79.3 %,灰分1.9 %~4.6 %(如
%,熱水溶出物16.5 %,苯醇溶出物2.7 %(黃金城,1984)。 表 2-2 2008 年彰化縣、雲林縣及嘉義縣全年花生之生產概況 縣市 種植面積(公頃) 每公頃產量(公斤) 產量(公噸) 百分比(%) 彰化縣 3,462 2,200 7,618 13.8 雲林縣 16,079 2,583 41,528 75.4 嘉義縣 858 2,332 2,000 3.6 小計 20,399 7,115 51,146 92.8 全國合計 22,537 38,830 55,076 100 資料來源〆農委會,2009 表 2-3 花生殼成份分析表 成份 百分比(%) 蛋白質 4.8 脂肪 1.2 碳水化合物 10.6 澱粉 0.7 半纖維素 10.1 粗纖維素 70.7 灰分 1.9 合計 100 二、花生殼之用途 目前花生殼大多用於與農業廢棄物混合醱酵製成堆肥,再回歸到農田以及當 燃料燃燒,較少用於當能源作物來研究。而花生殼中含有高量的纖維素約80 %(其 中包含木質素28 %、半纖維素10.1 %),適合用於水解醱酵以產氫々所以本研究 打算以花生殼作為本研究農業廢棄物批次詴驗及SBR 產氫的基質來源。儘管有 大量農業廢棄物原料可重新使用,但這些材料在技術上與循環使用上仍存在著一 定的障礙,這正是本研究需努力的地方。
第四節 水解微生物裂解纖維素
在都市、森林與農業廢棄物中,纖維質廢棄物約佔有40-70 %(Burrell et al., 2004々Van Wyk, 2001)纖維素含量甚高。纖維素在自然界中是一種很豐富的多醣 體,而對環境來說更是種友善的生質材料,亦可再經生物轉換成其它再生能源。 近年有許多學者發現以纖維素或纖維質廢棄物進行生物分解產能之研究比直接 焚化更具有能源再生的潛能(Cooney et al., 1987々Van Wyk, 2002々Cowan, 1992; Yang et al., 2004々Thanakoses et al., 2003)。而本研究所選用之基質為農業廢棄物 花生殼,花生殼含有高量的纖維素,適合用於水解醱酵以產氫。 一、具纖維素分解能力的生物 具纖維素分解能力的微物,包括、真菌或放線菌等,對此類微生物而言,纖 維素分解酵素可以幫助其分解纖維素,作為碳源利用,部分微生物產生纖維素分 解酵素以幫助孢子萌發,或破壞宿主細胞的細胞壁,例會與豆科植物形成共生的 根瘤菌。而真核生物的植物,會生產纖維素水解酵素,造成細胞的生長,又如植 物種子含有纖維素分解酵素,可幫助種子萌發。 具纖維素分解能力的生物多數為細菌、真菌和放線菌,就目前研究所知,真 菌類可產生各種纖維素水解酵素,包含內切型、外切型纖維素水解酵素和β-葡萄 糖苷酵素,構成一完整纖維素水解系統,其中木黴菌(Trichoderma reesei)是研 究應用上最多也最常見的一種。相對於真菌類完整的纖維素水解系統,只有少部 分細菌可生產外切型纖維素水解酵素,例如部分細菌並不生產 cellobiase,而是以 cellobiose phosphorylases 和 cellodextrin phosphorylases 來 分 解 纖 維 雙 醣 (cellobiose)和纖維糊精(cellodextrins)。目前已發現能產生纖維素分解酵素的 細 菌 包 括 〆 Clostridium, Cellulomonas, Bacillus, Thermomonospora, uminococcus,Bacteroides, Erwinia, Acetivibrio, Streptomyces, Cellvibrio, Pseudomonas,Rhizobium 和 Microbispora 等。在許多細菌中也具有分解纖維素的
其纖維素被水解的反應式如下〆 (C5H8O4)n+nH2O → nC5H10O5 ………(2-1) (C6H10O5)n+nH2O → mC6H12O6 ………(2-2) C6H12O6+6O2 → 6CO2+6H2O ………(2-3) C6H12O6 → 3CH4+3CO 2 ………(2-4) 表 2-4 目前已知可分解纖維素之厭氧性微生物 Anaerobes (厭氧性微生物)/Facultative aerobes
Acetivibrio cellulolyticus
Bacteroides (Fibrobacter) succinogenes Bacteroides cellulosolvens Bacteroides xylanolyticus Butyrivibrio fibrisolvens Caldocellum saccharolyticum Clostridium acetobutylicum Clostridium cellobioparum Clostridium cellulolyticum Clostridium cellulovorans Clostridium josui Clostridium papyrosolvens Clostridium stercorarium Clostridium thermocellum Clostridium thermocopriae Erwinia carotovora Erwinia chrysanthemi
Eubacterium (cillobacterium) cellulosolvens Micromonospora propionici a Micromonospora ruminantium a Pseudonocardia thermophila Ruminococcus albus Ruminococcus flavefaciens Thermoanaerobacter cellulolyticus
a No longer recognized as true members of genus, or by this name or by inclusion in Bergey’s Manual (1984-1989). 資料來源〆Bergey, David, Krieg, Moeir & John(2001)
表 2-5 目前已知可分解纖維素之好氧性微生物 Aerobes(好氧性微生物)/Facultative anaerobes
Acidothermus cellulolyticus Cytophaga johnsonae
Actinoplanes brasiliensis Cytophaga krzemieniewskae a Actinoplanes phillipinensis Cytophaga pectinovora Actinopolyspora halophila Cytophaga rubra a
Archangium spp. Cytophaga tenuissima a
Arthrobacter spp. Cytophaga winogradskii a
Bacillus brevis Frankia sp
Bacillus cereus Herpetosiphon geysericolus
Bacillus coagulans Microbispora bispora
Bacillus firmus Micromonospora aurantiaca a
Bacillus licheniformis Micromonospora chalcae
Bacillus pasteurii Micromonospora inositola
Bacillus polymyxa Micromonospora melanosporea
Bacillus pumilus Micromonospora olivasterospora
Bacillus stearothermophilus Polyangium spp.
Bacillus subtilis Pseudomonas fluorescens var. cellulosa Cellulomonas biazotea Pseudomonas solanacearum
Cellulomonas cellulans (formerly C. cartae) Serratia marcescens
Cellulomonas cellasea Sorangium spp.
Cellulomonas fermentans a Sporocytophaga myxococcoides Cellulomonas fimi Staphylococcus saprophyticus Cellulomonas flavigena Streptomyces albogriseolus Cellulomonas gelida Streptomyces celluloflavus Cellulomonas turbata a Streptomyces cellulosae
Cellulomonas uda Streptomyces flavogriseus
Cellvibrio flavescens a Streptomyces lividans Cellvibrio fulvus a Streptomyces nitrosporeus Cellvibrio gilvus a Streptomyces olivochromogenes Cellvibrio mixtus a Streptomyces thermodiastaticus Cellvibrio ochraeus a Streptomyces thermovulgaris Cellvibrio viridus a Streptomyces viridosporus Cellvibrio vulgaris a Thermoactinomyces cellulosae Corynebacterium michiganense Thermomonospora curvata
Cytophaga aprica Thermomonospora fusca
Cytophaga aquatilis Xanthomonas albilineans Cytophaga aurantiaca Xanthomonas campestris Cytophaga hutchinsonii
a No longer recognized as true members of genus, or by this name or by inclusion in Bergey’s Manual(1984-1989).
目前至少有 85 種 Clostridium (梭狀芽孢桿菌屬)被發現(Cato et al., 1986), 雖然大部分的 Clostridium 無法分解纖維素,但在能夠分解纖維素的微生物中,
Clostridium 仍佔了相當量的族群。Clostridium 屬於中溫菌,而依過去許多的研
究中便可發現,在許多有機物中可分離出 Clostridium(Duong et al., 1983)。例 如,在動物糞肥中分離出(Viljeon et al., 1926; McBee, 1950, 1954)々在污泥中分 離出 Clostridium thermocellum LQRI(Lamed & Zeikus, 1980)々以及在棉絮中分 離出 Clostridium thermocellum JW20(Ljungdahl et al., 1981)。
國內生物資源保存及研究中心生物資料庫發現,在所有分解纖維素的細菌 中,以 Clostridium thermocellum 及 Clostridium cellulolyticum 最有分解纖維素潛 力,其生長環境,Clostridium thermocellum JW20 可生長在纖維素、木質素中々而 Clostridium thermocellum 生長在纖維素中的一個特徵是會產生一種黃色色素物質 (YAS)(Ljungdahl et al., 1983),除此之外,此株菌亦可生長在葡萄糖、果糖、 木糖中,在最適生長環境方面,pH 約介於 6.1-7.5 之間,溫度為 28-69 ℃間(最 理想之溫度為 60℃),而當以 cellulose 及 cellobiose 為基質時,其生長的適應時間 分別為 6.5 hrs 及 2.5 hrs。而 Clostridium cellulolyticum 在 pH 7.5、溫度 35℃時, 有最佳的生長狀況。 二、纖維素水解 纖維素是植物結構的主要物質(如圖 2-1 所示),其主要以 D-glucose 為基本 單體,單體之間的鍵結藉由〆α-1, 4-glucosidic bonds 聚合成直鏈狀巨分子,與相 鄰的葡萄醣分子重複形成直鏈狀巨分子,而鏈的長度約為 100~14,000 個葡萄醣 分子,鏈與鏈之間,利用氫鍵彼此鍵結,以帄行排列並折疊成緊密且結晶度高的 多醣體,纖維長鏈之間的氫鍵結形成穩固結構的結晶型(crystalline domain)纖 維(如圖 2-2 所示),具有極穩定不溶於水且不易分解的特性,需要以化學或生物 方式水解成簡單的醣類,這些分子較小的醣類即可提供其它生物利用,水解方式 包括化學與酵素或生物方式,化學方式包含酸水解、超臨界二氧化碳與濕氧化等 方式,化學方式多以強酸或高溫高壓進行水解,且處理反應劑量與時間控制不
易,容易過度分解成其它副產物或反應不完全(Kim and Hong,2001々Fox and Noike, 2004)。生物處理可以免除化學處理的缺點,利用添加纖維分解酵素(cellulases) 或真菌、纖維素分解菌群將纖維水解成可溶解醣類(Shoham et al.,1999),操作簡 易更可降低操作管理的成本,且纖維素分解菌群本身具有纖維分解酵素,與直接 添加酵素的成效相當,皆具有纖維素分解為醣類並同時醱酵的功能,由纖維素分 解後產生的醣類或酸類如葡萄醣、纖維雙醣、醋酸等,可加以運用成為生質能材 料來源,對於環境是種友善兼具產能的方式(Zeikus, 1979々Van Wyk, 2001)。
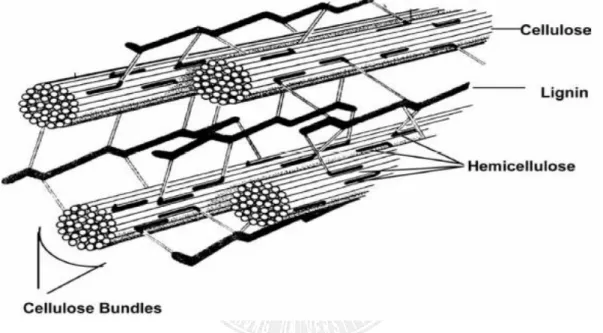
圖 2-1 植物纖維素細胞壁的排列示意圖(Murphy and McCarthy, 2005 a)
第五節 SBR 能源化技術探討
一、基本原理
廢水按批次操作(operated in batch mode)的概念最早於1914 年提出(Torrijos, 1994),而在半個多世紀以後,才由美國諾羅丹大學的Irvine提出了循序批次反 應槽SBR(Sequencing Batch Reactor)的名稱,並嘗詴在實際廢水工程上加以應 用。SBR 法為廢水的生物處理程序,屬活性污泥法的範疇,而污泥物去除的機 制、BOD 負荷等參數均適合於SBR循序批次活性污泥法有、厭氣-好氣操作、間 斷進流的特點。實際上,SBR 法是一種半連續-間歇式反應槽,它與傳統的曝氣 池不同。進流方式可以是間歇式,也可以是連續式,而放流階段一般是採間歇式。 SBR 法作為廢水的生化處理方法,屬活性污泥法的範疇,而污泥物去除的 機制、BOD 負荷等參數均適合於SBR,程序批次活性污泥法兼有推流、厭氣-好 氣操作、間斷進流的特點。實際上,SBR 法是一種半連續-間歇式裝置,它與傳 統的充放式曝氣池不同。進流方式可以是間歇式,也可以是連續式,而放流階段 一般是採間歇式々從曝氣方式而言,可以採用進流時不曝氣的限制曝氣方式,進 流時曝氣的非限制曝氣方式或進流後曝氣的半限制曝氣方式(李軍,1994)。 SBR 法的運作以間歇式操作為主要特徵,所謂序列間歇有兩種含義〆一是 運行操作在空間上是按程序排列、間歇進行,由於污水是連續按序列進入反應 器,它們運作時的相對關係是序列、間歇的々二是每個SBR 的運行操作在時間 上也是按序排列、間歇進行,一般反應槽操作分為下列四個不同的階段〆(1) 進流(fill)々(2)攪拌(react)々(3)沉降(settle)々以及(4)抽離(draw), 這四個階段組成一個週期,如圖2-3。這樣SBR 就把單一反應槽在時間上進行分 割,各部分獨立完成連續穩定的處理過程(任立斌,1998)。
1.進流 2.攪拌 3.沈降 4.抽離 進流基質 上澄液 污 泥 有 效 體 積 圖 2-3 SBR 每一循環的典型操作程序流程圖 (一)進流階段 進流階段為一循環的開始,緊接上一週期的放流狀態,反應槽內留有活性污 泥,且槽內以達最低水位,進流後基質開始流入,隨著進流時間的增加,水位不 斷上升。通常在進流階段可以有兩種不同操作情形〆(1)進流即同時開始攪拌 階段々(2)進流後才開始攪拌階段。至於採用何種方法,乃依廢水的性質和處 理目標而定。 (二)攪拌階段 在此階段反應槽內保持最大水量,並藉著攪拌讓微生物與基質完全混合,達 到處理的目的。 (三)沉降階段 反應槽停止攪拌,反應停止後,活性污泥絮狀體藉著重力沉澱,達到固液分 離的目的々此時污泥是在完全靜置狀況下沉降,固沉降效果極佳,能有效截流菌 體濃度(VSS)。 (四)抽離階段 反應槽內水位達到設計的最高水位並經沉澱後,開始排放上澄液,當槽內水 位恢復到處理週期開始的最低水位(大於污泥高度)停止放流。在放流後,因尚
有一部份上澄液殘留在反應槽內,故在下一循環進流階段時,此殘留的上澄液有 稀釋的作用,可避免突增負荷(Shock Loadind)的影響。 二、SBR 之優點 SBR 法在單一反應槽中以時間為導向,控制每一個批次循環操作的廢水生 物處理程序,此操作程序介於傳統批次與連續操作之間,不但能連續操作馴養污 泥,更能有效截留菌體量。SBR 法除了具有設備及操作成本低廉、節省佔地面 積、節省能源、易於控制等優點外,尚有下列優點〆 (一)耐衝擊負荷強 SBR 兼具有耐衝擊負荷強及反應推力大的優點,並且SBR 法的沉降階段為 靜止沉澱,沉降性能好及不需要污泥迴流,使反應器中維持較高的MLSS 濃度。 故SBR 法更具耐衝擊負荷能力。 (二)能充分防止污泥膨脹 污泥膨脹多為絲狀膨脹,在活性污泥法中,間歇式最不易發生膨脹,完全混 合式最容易引起膨脹。按照發生膨脹難易程度的順序為〆間歇式、傳統推流式、 階段曝氣式和完全混合式,同時其降解有機物速率也是遵循此順序。SBR 法能 夠有效地控制絲狀菌的過量繁殖(Heinrich, 1995),這一特性是由缺氧好氧狀態 並存、反應中底物濃度較大、泥齡短、比增長速率大決定的。 (三)沉澱效果佳 SBR 反應槽固液分離在近乎理想的靜止狀態下進行,且沉降時不會有短流 (short circuiting)現象發生,故污泥沉降性較連續式佳,且不易有鬆化現象。 本研究選定 SBR 的原因是上述(一)~(三)之原因,不穩太操作有利於水 解,才選用 SBR。故本研究選定具有分解 cellulose 之 Bacillus 菌株與進行大量 培養之醱酵產氫菌,進行不同之配比組合批次研究,選出分解力較佳的分解菌, 作為 SBR 反應槽中主要菌種。又因 SBR 其操作過程可以適當調整其溶氧含量經
過一段時間的反應後,可以達到完全的厭氧,因此可以適合 Bacillus 及 Clostridium 的分離培養反應,而達到有效分解 cellulose、木質素,以產氫為目的。因此本研 究將第一段連續流反應槽將以 SBR 來水解農業廢棄物花生殼中的纖維素、木質 素等成分。
第六節 厭氧產氫微生物
產 氫 微 生 物 可 分 為 二 大 類 , 一 為 光 合 產 氫 微 生 物 ( photosynthetic microorganisms ),另一類為醱酵產氫微生物(fermentative hydrogen-producing microorganisms),後者具有較佳的生長速率,進行生物產氫的反應速率較前者 為快,且不需要光能,因此較具優勢(Das & Veziroglu, 2001)。生物產氫的程序 可區分為〆(一)藻類及藍綠細菌以光合作用分解水,作為電子供給,進行產氫。 (二)光合細菌以無氧光合作用分解有機物作為電子供給者,進行產氫。 (三)發酵細菌以有機物進行發酵產氫。
(四)結合光合及發酵細菌進行產氫(Debarata & Nejat Veziroglu,2001)。而光 合細菌及發酵產氫均要在厭氧條件下才能進行。 一、厭氧產氫微生物 至 目 前 為 止 , 許 多 研 究 發 現 最 適 合 產 氫 的 菌 屬 包 括 梭 狀 芽 孢 桿 菌 屬 (Clostridium)及腸桿菌屬(Enterobacter)。梭狀芽孢桿菌屬於目前產氫能力較 佳之菌種(Gottschalk, 1986),適合生長在厭氧、中溫及中性pH 值的環境(Minton and Clarke, 1989),但在不適合生長的環境中會產生內生孢子而進入休眠狀態, 這是具孢子微生物的特性,可藉此抵抗惡劣的生存環境。一般而言,各種產孢菌 在不同的生長環境條件會形成內生孢子,這些生長環境條件,包括〆pH 值、溫 度、營養成分與供氧狀況等。因此,由過去許多的研究中發現,有很多微生物均 具有產氫能力,如表2-6(Das & Veziro lu, 2001)。但因各產氫菌生理特性的不
同,其產氫能力也不同,對於環境的要求亦有所差異(林秋裕、林明正、陳晉照, 1999)。
產 氫 菌 主 要 分 為 厭 氧 醱 酵 產 氫 菌 ( Fermentative hydrogen producing microorganisms)及光合產氫菌(Photosynthetic microorganisms)二大類(Yokoi et
al., 1995)。光合作用微生物部份包括〆紫色非硫菌、藻類等。醱酵產氫微生
物又依其對氧需求性分為兼性厭氧菌(如Bacillus、Enterobacter aerogenes等), 及絕對厭氧菌(如Clostridium 等)。各方資料(Yokoi, 1997々Odom, 1983)均顯 示〆目前研究以紫色非硫菌產氫效率甚佳,但需要照光及靜置的環境中生長々而 醱酵產氫微生物中以Clostridium 菌屬為主,此菌屬大部分都可以產氫,其中以
Clostridium butyricum 菌 種 產 氫 更 是 可 觀 , 醱 酵 產 物 為 butyrate 、 butanol 及
acetone。表2-7為各種不同產氫菌進行厭氧醱酵之產物(Gottschalk, 1986)。 目前文獻中有關厭氧產氫純菌的研究是以Enterobacter 及Clostridium 兩屬 微生物為主要的研究菌群。Entrobacter 為革蘭氏陰性(gram negative)、兼性厭 氧菌(facultative anaerobe),主要醱酵產物為氫氣、二氧化碳及揮發酸。而
Clostridium 為革蘭氏陽式(gram positive)、絕對厭氧菌(obligately anaerobe),
且會形成內生孢子(endo spore),主要發酵生成物為氫氣、二氧化碳、揮發酸及 醇類。
Clostridium 菌屬的產氫能力近年來已逐漸受到重視,主要是因為針對混合菌
統產氫程序的菌種鑑定結果發現主要的菌群是以Clostridium 為主(Ueno et al., 2001; Fang et al., 2002a)。Yokoi 等人(1998)指出Clostridium 屬菌單位基質的 產氫率是Enterobacter 的二倍(2 mol H2/mol-hexose),因此在植種菌中加入 clostridia 是有利於產氫的,但缺點為極易受氧氣的抑制,且孢子萌發時有特殊的 環境及營養需求。
表 2-6 厭氧產氫微生物之種類 分類 微生物名稱 Green algae (綠藻) Scenedesmus obliquus Chlamydomonas reinhardii Chlamydomonas movewusii Cynobacteria Heterocystous (藍綠菌) Anabaena azollae Anabaena CA Anabaena variabilis Anabaena cylindrical Nostoc muscorum Nostoc spongiaeforme Westiellopsis prolifica
Nonheterocystous Plectonema boryanum Oscillotoria Miami BG7 Oscillotoria limnetica Synechococcus sp. Alphanothece halophytico Mastidocladus Phormidium valderianum Photosynthetic bacteria (厭氧光合細菌) Rhodobater sphaeroides Rhodobater capsulatus Rhodobater sulidophilus Rhodopseudomonas sphaeroides Rhodopseudomonas palustris Rhodopseudomonas capsulata Rhodospirillum rebnum Chromatium sp. Miami PSB 1071 Chlorobium limicola Chloroflexu aurantiacus Thiocapsa roseopersicina Halobacterium halobium Fermentative bacteria (厭氧醱酵菌) Enterobacter aerogenes Enterobacter cloacae Clostridium butyricum Clostridium pasteurianum Desulfovibrio vulgaris Magashaera elsdenii Citrobacter intermedius Escherichia coli
表 2-7 各種不同菌種進行醱酵代謝之產物 醱酵種類 菌種 產氫量及產物 乙醇 Zymomonas anaerobica 少量氫氣 Sarcina ventriculi 醋酸 Erwinia amylovora 乳酸 丁酸(酯) Clostrium 大量的氫氣 丁醇 Clostridium acetobutylicum 丁酸(酯) 丙酮 Clostridium beijerinckii 己酸 Clostridium tetanomorphum Clostridium aurantibutylicum Clostridium kluyveri Butyrivibrio Eubacterium Fusobacterium 混酸 Escherichia coli 中量的氫氣 丁二醇 Enterbacter aerogenes 資料來源〆Gottschalk,(1986) Clostridum 除具有不錯的產氫能力外,還有一項有利於菌種篩選的功能,就 是會形成內生孢子,因此,在厭氧甲烷化系統中,提供一個較為惡劣的環境,如 加入強酸、強鹼、抑制劑或加熱煮沸等方式以殺死甲烷菌並保留已經形成孢子的 Clostridum ,藉此達到菌種篩選之目的,以利於進行之後的厭氧生物產氫程序。
第七節 厭氧醱酵產氫反應機制
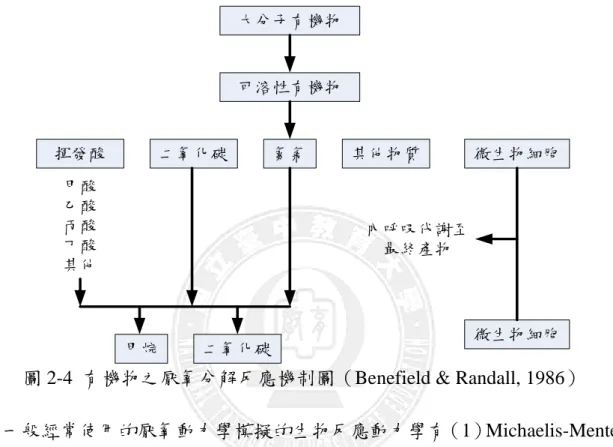
一、厭氧醱酵產氫反應 厭氧消化乃是指在厭氧條件下,藉著兼氧性及厭氧性微生物生化代謝作用, 將複雜之高分子有機物(如蛋白質、碳水化合物、脂質等)經過在傳統厭氧反應 分解有機物的過程中,厭氧及兼氣性微生物群先將有機物經水解產酸反應,將大 部分有機物轉化為揮發酸、醇、醛、氫氣等中間代謝產物,若生長條件適當,這 些中間代謝產物又會被甲烷生成菌分解或轉換成甲烷及二氧化碳。而厭氧醱酵就產氫反應,則是傳統厭氧反應進行至醱酵階段中止反應之繼續進行,而達到醱酵 產氫之目的,這則需要一些特殊操控技術及能利用特殊微生來達成。
由Das and Vezirolu(2001)的研究中指出厭氧醱酵產氫反應有非常高的氫氣 轉換率、穩定的氫氣產生量及良好的微生物生長率等優點,因此在微生物共培養 狀況良好的系統中,電子、氫與能量在水解、酸化與甲烷化反應之間傳遞著,氫 氣很難被釋放至氣相之中,但是反應槽中微生物共共培養系統一旦失去帄衡,氫 氣未被甲烷化反應快速消耗,則將會被釋放至氣相中。在厭氧醱酵產氫機制方 面,由於能量是微生物存活所必頇的原動力,而微生物細胞內能量的來源,主要 來自可釋放能量的化學反應,故此能量為一種化學能。此種化學能貯存在許多的 化合物內,當分子分解時,可將貯存的能量釋出(楊美桂,2003)。
Girbal, Crox, Vasconcelos and Soucaille ( 1995 ) 的 研 究 提 出 Clostridium
acetobutylicum 分解葡萄糖的厭氧醱酵路徑。不同路徑所生成的代謝產物也不一
樣,包括lactate、acetone、ethanol、butanol、butyrate、acetate等。而最佳的厭氧 產氫反應為產乙酸的反應途徑。醣類轉換到pyruvate 的過程可產生NADH,而 NADH 在hydrogenase 及ferredoxin 的作用下轉回NADH 並釋出H2。在電子傳遞 的 過 程 中 可 發 現 , 在 產 acetate 的 路 徑 上 NADH 不 會 被 消 耗 , 幾 乎 全 部 由 hydrogenase 及ferredoxin 的作用而釋放出H2。由電子的轉移、能量的轉換探討, 微生物分解葡萄糖是傾向產生氫氣與乙酸的途徑。所以葡萄糖及蔗糖是最常用來 進行產氫研究的基質。理論上每莫耳的葡萄糖可產生4莫耳的氫氣,此時乙酸是 醱酵的副產物々當醱酵的副產物為丁酸時,產氫量僅為產乙酸時的一半而已 (Girbal et al., 1995)。 一般傳統厭氧消化反應可分為三階段,即水解、酸化、及甲烷化三階段。而 厭氧醱酵產氫反應則發生在水解及酸化階段,以下就這二階段進行討論。 (一)水解階段 此階段是利用細胞外水解(extracellular enzymes)將複雜的有機物(如蛋白 質、脂肪、多醣類等)分解成簡單的化合物(如胺基酸、甘油、單醣等),而水