薑黃素和鼠尾草酸抑制細胞激素誘發之人類大主動脈平滑肌細胞的遷移和基質金屬蛋白酶的活化; Curcumin and Carnosic acid inhibit MMP-9 activity and migration of cytokine-induced human aortic smooth muscle cells
107
0
0
全文
(2) 謝. 誌. 感謝指導教授余雅美博士兩年來於學業、研究等方面的悉心指導 及待人處事上的諄諄教誨,使本論文得以順利完成。師恩浩瀚,永銘 於心,謹誌卷首,以表誠摯之謝忱。 文稿初成,承蒙台灣大學沈立言教授與中國醫藥大學江素瑛助理 教授於百忙之中撥冗審閱,詳加斧正,並提供寶貴意見使論文內容得 以更加完善,謹此致上萬分謝意。 在學期間,多蒙學長姐的指導與同窗好友芩卉、珮玲、馨儀、美 瑜、美存、珊瑩、昇輝及同期研究所同學於實驗與生活上的幫助與支 持;實驗室之學妹瑜妏、于婷、郁婷、珮琪、榆茜的協助與幫忙;再 者,要感謝好友們佩文、映廷、子綺、Yurika、怡真、美玲,於艱辛 研究過程中給予諸多的協助與鼓勵,在此致上最誠摯的謝意。 最後由衷感謝男友育成及其家人伯父、伯母及俊宏,還有我最親 愛的家人們村村爸、珍珍媽、阿憲弟弟以及綉妃的關懷與支持,使我 能安心無慮的完成學業。謹此將本論文獻給所有關心及曾經幫助我的 人,謝謝你們。 林杏純 謹致於 中國醫藥大學 營養學系 中華民國 九十六年六月.
(3) Contents 摘 要 ……………………………………………………………………………… 1 Abstract………………………………………………………………………… 2 Chapter 1 Literature review…………………………………………………... 4 1.1. Morphology of artery........................................................................................ 4. 1.2. Oxidative stress and atherosclerosis ................................................................. 4. 1.3. Matrix metalloproteinases (MMPs) and atherosclerosis .................................. 6. 1.4. Nuclear factor-κB (NF-κB) and inflammation ................................................. 8. 1.5. Curcumin.………………………………… …………………………………..9. 1.6. Carnosic acid (CA)……………………………………………………………10. 1.7. The objective of this research…………………………………………………10. Chapter 2 Experimentation I…..……………………………………………... 20 2.1 Introduction …………………………………………………………………..21 2.2. Materials..………………………………………………………. .....................23. 2.3. Methods………………………………………………………………………..27. 2.4. Results………………………………………………………………………... 34. 2.5 Discussion……………………………………………………………………. 37 2.6. Conclusion…………………………………………………………………..... 41. Chapter 3 Experimentation II.………………………………………………... 51 3.1 Introduction …………………………………………………………………..52 3.2. Materials……………..………………………………………………………..54. 3.3. Methods……………………………………………………………………….58. 3.4. Results………………………………………………………………………...67. 3.5 Discussion…………………………………………………………………….70 3.6. Conclusion………………………………………………………………….... 74. I.
(4) Chapter 4 Summary………….………………………………………………... 85 Reference………….………….………………………………………………... 87 Appendix………….………….…………………………………………………..99. II.
(5) Table of contents Table 1. MMPs and their substrates………………………………………………..16. Table 3-1 Antioxidative capacities of CA in vitro…………………………………75. III.
(6) Figure of contents Fig. 1. The structure of the normal blood wall……………………………………..13. Fig. 2. The function of VSMCs during different stages of atherosclerosis………...14. Fig. 3. The regulation of MMPs…………………………………………………....15. Fig. 4. The signal transduction pathway of NF-κB activation……………………..17. Fig. 5. The structure of curcumin…………………………………………………..18. Fig. 6. The structure of carnosic acid………………………………………………19. Fig. 2-1. Cytotoxic effect of curcumin on HASMCs with MTT test. ……………...42. Fig. 2-2 Effect of curcumin on the MMP-9 activity of TNF-α-induced HASMCs..43 Fig. 2-3. Curcumin inhibits the protein expression of MMP-9 in HASMCs……….44. Fig. 2-4. Effect of curcumin on TNF-α-induced activation of NF-κB p50 in HASMCs. ···················································································································· 45. Fig. 2-5. Effect of curcumin on TNF-α-induced activation of NF-κB p65 in HASMCs. ···················································································································· 46. Fig. 2-6 Effect of curcumin on migrarion of HASMCs induced by TNF-α treatment. ···················································································································· 47 Fig. 2-7. Effect of curcumin on TNF-α-induced ROS production in HASMCs……49. Fig. 3-1. Cytotoxic effect of CA on HASMCs with MTT test……………………...76. Fig. 3-2. Effect of CA on the MMP-9 activity of TNF-α-induced HASMCs………77. Fig. 3-3. CA inhibits the protein expression of MMP-9 in HASMCs………………78. Fig. 3-4 Effect of CA on TNF-α-induced activation of NF-κB p50 in HASMCs….79 Fig. 3-5 Effect of CA on TNF-α-induced activation of NF-κB p65 in HASMCs….80 Fig. 3-6. Effect of CA on migrarion of HASMCs induced by TNF-α treatment……81. Fig. 3-7 Effect of CA on TNF-α-induced ROS production in HASMCs…………...83. IV.
(7) Fig. 4-1. The inhibitory effect of curcumin and carnosic acid on proanthersclerotic. mechanism in vitro…………………………………………………………………...86. V.
(8) Appendix Table 1.. Antioxidative capacities of curcumin in vitro……………………………99. VI.
(9) 摘要. 動脈粥狀硬化在美國及其他西方國家中是造成死亡的主要疾病。血管平滑肌 細胞的遷移和基質金屬蛋白酶的活化,在動脈粥狀硬化的發展中扮演一個重要的 角色。薑黃素是衍生自薑黃的多酚,目前已有數百萬人會每天攝取。一般而言, 薑黃素與數種生物和細胞活性有關,包含抗氧化、抗發炎、抗癌,以及降低血中 膽固醇的特性。鼠尾草酸是存在鼠尾草與迷迭香中的主要酚類化合物,在過去的 體外試驗中已被證實具抗氧化的能力。本研究之目的主要探討薑黃素和鼠尾草酸 抑制 TNF-α 誘發人類大主動脈的平滑肌細胞的遷移與基質金屬蛋白酶-9 (MMP-9) 的相關性。在遷移試驗中顯示薑黃素和鼠尾草酸可有效抑制 TNF-α 誘發之平滑肌 細胞的遷移。由 Gelatin zymography 試驗顯示薑黃素和鼠尾草酸會降低 MMP-9 的分泌,且西方墨點法也顯示薑黃素和鼠尾草酸會降低 MMP-9 的表現。經 NF-κB 酵素免疫分析試驗證實,薑黃素和鼠尾草酸能減少核轉錄因子 NF-κB p50 與 p65 的活化,以及活性氧物種的產生。總而言之,薑黃素和鼠尾草酸降低平滑肌細胞 的遷移是經由抑制 TNF-α 誘發 p50 和 p65 的核轉位,而抑制 MMP-9 的分泌及蛋 白質表現。因此,薑黃素和鼠尾草酸在預防動脈粥狀硬化中,具有抗發炎的特性 且可能扮演一個重要的角色。. 關鍵字:動脈粥狀硬化、平滑肌細胞遷移、基質金屬蛋白酶、薑黃素、鼠尾草酸、 核轉錄因子-κB、活性氧成分、腫瘤壞死因子-α。. 1.
(10) Abstract. In the United States and other Western countries, atherosclerosis is the leading cause of illness and death. The migration and matrix metalloproteinases (MMPs) activation of vascular smooth muscle cells (VSMCs) may play key roles in the development of atherosclerosis. Curcumin, which is consumed daily by millions of people, is a polyphenol derived from the plant Curcuma longa. In general, curcumin has been associated with a large number of biological and cellular activities, including antioxidative,. anti-inflammatory,. anticarcinogenic,. and. hypocholesterolemic. properties. Carnosic acid (CA) is the primary phenolic compound in rosemary and salvia. Previous study indicated that CA possesses antioxidant activity in vitro. In this study, we investigated the inhibitory effect of curcumin and CA on tumor necrosis factor-α (TNF-α)-induced the migration in human aortic smooth muscle cells (HASMCs) and MMP-9 activity. The migration assay showed that curcumin and CA effectively inhibited the TNF-α-induced migration of HASMCs as compared with the control group. Curcumin and CA lowered the secretion and protein expression of MMP-9 by gelatin zymography and western blot assays. They also decreased nuclear translocation of nuclear factor-κB (NF-κB) p50, p65, and ROS production. In conclusion, curcumin and CA inhibit TNF-α-induced nuclear translocation of p50 and p65, thereby suppressing the secretion and protein expression of MMP-9, resulting in decreased HASMCs migration. Thus, curcumin and CA have anti-inflammatory properties and may play important roles in the prevention of atherosclerosis.. 2.
(11) Key words: Atherosclerosis, vascular smooth muscle cells (VSMCs) migration, matrix metalloproteinases (MMPs), Curcumin, Carnosic acid (CA), NF-κB, ROS, TNF-α. 3.
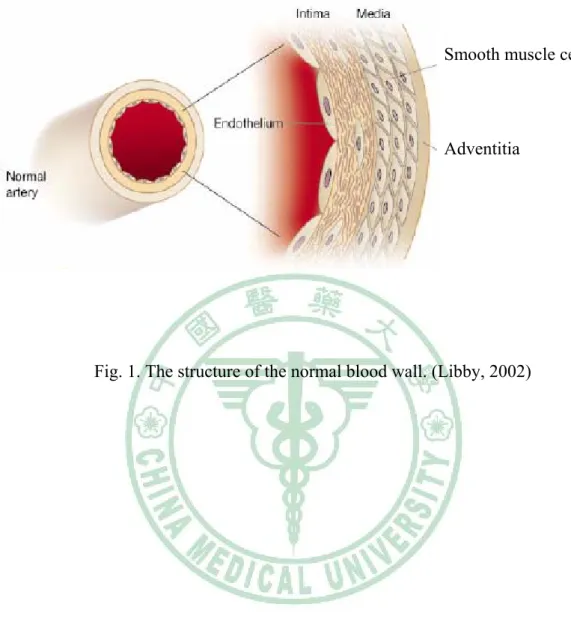
(12) Chapter 1 Literature review. 1.1 Morphology of artery. The structure of the normal blood wall comprises three layers: the innermost layer is called the intima, the middle layer is called the media, and the outermost layer is known the adventitia (Fig. 1). The intima is an inner monolayer of endothelial cells lining the lumen and is bound on the outside by internal elastic lamina, a fenestrated sheet of elastin fibers. The thin subendothelial space in between contains thin elastin and collagen fibers along with a few smooth muscle cells (SMCs). The extracellular matrix (ECM) of the vascular wall, largely elastin and collagen, subserves many functions essential for vessel homeostasis. ECM damage or remodeling in conditions such as rheumatoid arthritis (Weber and De Bandt, 2000), malignant tumors (Ohtani, 1998), aortic aneurysm (MacSweenety, 1994), and atherosclerosis (Libby, 1995; 2000) contributes to the formation, progression, and clinical expressions of these disorders.. 1.2 Oxidative stress and atherosclerosis. Several lines of evidence indicate that oxidative stress may play an important role in various pathological conditions, including cancer, neurodegeneration, atherosclerosis, diabetes, rheumatoid arthritis, postischemic reoxygenation injury, and aging. Oxidative stress may lead to many cellular events, such as inactivation of NO,. 4.
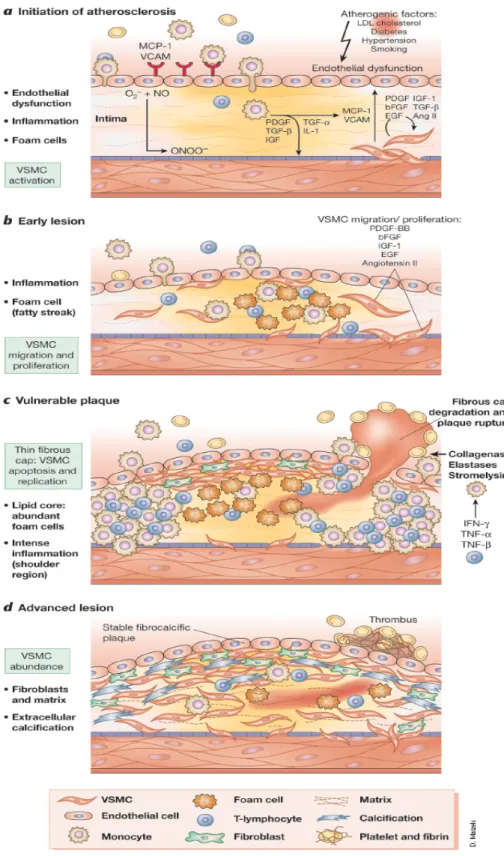
(13) oxidative modifications of DNA and proteins, lipid oxidation, enhanced mitogenicity and apoptosis of vascular cells, and increased expression and activation of redox-sensitive genes, such as the receptor for oxidized LDL, adhesion molecules, chemotaxis factors, NF-κB (Muller et al, 1997), proinflammatory cytokines, regulators of cell cycle progression, and matrix metalloproteinases (Wassmann et al, 2004). Vascular lesions form during pathological processes, which involve the accumulation of inflammatory cells and release of cytokines. Production of cytokines and growth factors within lesions may induce migration of SMCs. Proinflammatory cytokines, such as tumor necrosis factor (TNF) and interleukin-1 (IL-1), play important roles in the pathogenesis of atherosclerosis (Zhu et al, 1999; Rahman at al, 1998). TNF-α, an inflammatory cytokine released in greater quantities by obese humans and patients with insulin resistance, not only initiates but also propagates atherosclerotic lesion formation (Lau et al, 2005). Cardiovascular disease (CVD) due to atherosclerosis is the leading cause of death in the Western world and the second most common cause worldwide (Braunwald, 1997). Atherosclerosis comes from the Greek words athero (meaning gruel or paste) and sclerosis (hardness). Almost 200 years ago, London surgeon J. Hodgson observed the inflammatory characteristics of atherosclerotic lesions (Kaperonis et al, 2006). A causative role of inflammation and infection in the pathogenesis of atherosclerosis was first proposed in 1908 by Sir William Osler (Kaperonis et al, 2006). Atherosclerosis is a chronic inflammatory disease driven by risk factors that cause oxidative and inflammatory mechanisms. The major risk factors are gender, age (Lusis et al, 2004), obesity (Yudkin et al, 1999), cigarette smoking (Nordskog et al, 2003), exercise, hypercholesterolemia (Khoo et al, 1988) ,. 5.
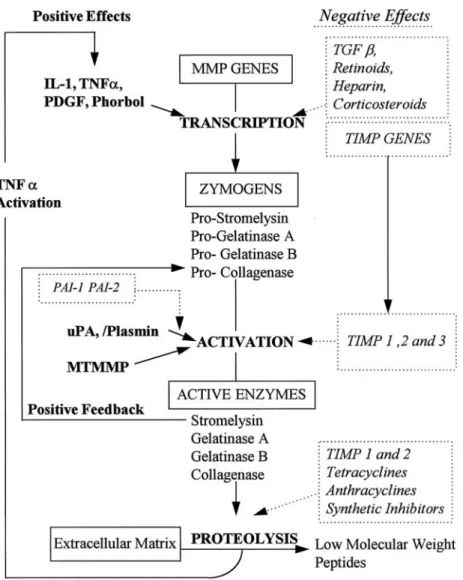
(14) hypertension (Griendling et al, 1997), hyperglycemia (Schmidt et al, 1999), hyperhomocysteinemia (Majors et al, 1997), and bacterial and viral infection (Jackson et al, 1997). The pathobiology of atherosclerosis comprises endothelial cells dysfunction, SMCs proliferation and migration, fibrous cap formation, plaque rupture and thrombosis. SMCs are important actors in the pathogenesis of atherosclerosis and restenosis after angioplasty or stent application (Hao et al, 2003). SMCs can synthesize and secrete biologically active mediators that regulate contraction and relaxation, inflammation, proliferation, apoptosis and matrix alterations (Dzau et al, 2002) (Fig. 2). The migration of SMCs from the tunica media to the subendothelial region is a key event in the development and progression of many vascular diseases including atherosclerosis and post-angioplasty restenosis (Maeda et al, 2002).. 1.3 Matrix metalloproteinases (MMPs) and atherosclerosis. MMPs were discovered in 1962, in an effort to establish how the metamorphosing tadpole of a frog lost its tail (Jones et al, 2003). MMPs are a family of Zn2+-dependent endopeptidases capable of cleaving components of ECM (Falk, 1999; Creemers et al, 2001). MMPs activity is regulated at three levels: gene transcription, activation of the latent proenzymes, and inhibition of proteolytic activity (Dollery et al, 1995; Birkedal-Hansen et al, 1993) (Fig. 3). MMPs can be categorized into the following groups: interstitial collagenases, gelatinases, stromelysins, membrane-type matrix metalloproteinases (MT-MMPs), and others. The collagenases include MMP-1, MMP-8, and MMP-13. Gelatinases are composed of MMP-2 and. 6.
(15) MMP-9. The stromelysin group is composed of MMP-3, MMP-10, and MMP-11. Six different membrane type (MT)-MMPs are MT-MMP1, MT-MMP2, MT-MMP3, MT-MMP4, MT-MMP5, and MT-MMP6. And others are MMP-7, MMP-12, MMP-19, MMP-20, MMP-23, and MMP-26 (Creemers et al, 2001) (Table 1). On the other hand, the activity of MMPs is controlled by specific inhibitors known as tissue inhibitors of MMPs (TIMPs). Three members of TIMPs have been identified to date: TIMP-1, TIMP-2, and TIMP-3. The balance between production of MMPs and TIMPs represents a critical point to maintain the homeostasis of the ECM (Birkedal-Hansen et al, 1993). MMPs activity may contribute to the pathogenesis of atherosclerosis by facilitating migration of VSMCs (Jones et al, 2003). MMPs (MMP-9 and MMP-2) production and SMCs migration may play key roles in the pathogenesis of neointima formation and atherosclerosis. Especially inducible MMP-9 expression was directly involved in the cancer cell invasion and SMCs migration through vascular wall (Cho and Reidy, 2002; Galis et al, 2002). Although MMP-2 and MMP-9 have similar substrate specificities, there are differences in the regulation of their expression. MMP-2 is constitutively expressed by several cell types, including SMCs, and its expression is not induced by cytokines or growth factors. In contrast, based on reports from several different laboratories, it has generally been concluded that the basal levels of MMP-9 are usually low, and its expression can be induced by treatment of cells with TNF-α, but not platelet-derived growth factor (PDGF) or thrombin (Cho et al, 2000; Galis et al, 1994).. 7.
(16) 1.4 Nuclear factor-κB (NF-κB) and inflammation. Nuclear factor-κB (NF-κB) was first described in 1986 as a nuclear factor necessary for immunoglobulin kappa light chain transcription in B cells (hence the name, NF-κB) (Ghosh et al, 1998). Transcription factors are proteins responsible for the coordinated expression of genes through specific binding to gene promoter and enhancer sites. Transcription factor NF-κB and its target genes are involved in the pathogenesis of atherosclerosis (Kutuk and Basaga, 2003). The NF-κB family members include RelA (p65), RelB, c-Rel, p50 (NF-κB1), and p52 (NF-κB2), the latter two being synthesized from the inactive precursor molecules P105 and P100, respectively, as well as their inhibitory subunits IκBα, IκBβ, and IκBε (Martin et al, 2000). Each family member contains an N-terminal 300 amino acid conserved region known as the rel homology domain (RHD) (Ghosh et al, 1998). NF-κB subunits form homo- and heterodimers, the most prominent one is p50/p65 heterodimers (Fig. 4). The dimmer is retained in the cytoplasm in an inactive state through interaction with IκB. NF-κB is rapidly activated in response to variety of inflammatory and other stimuli that lead to degradation of IκB (Martin et al., 2000). Active DNA-binding complexes of the NF-κB/Rel family consist of different homo- and heterodimers that bind to a common κB site (5’-GGGPuNNPyPyCC-3’), where Pu is purine, Py is pyrimidine and N is any nucleotide (Ghosh et al., 1998). The nuclear translocation of NF-κB is induced by many agents, including inflammatory cytokines (e.g. TNF-α and IL-1), mitogens, bacterial products, protein synthesis inhibitors, oxidative stress (H2O2), ultraviolet light, and phorbol esters (Baeuerle, 1991; Grilli et al, 1993). Upon activation of NF-κB, a large number of genes are induced including various. 8.
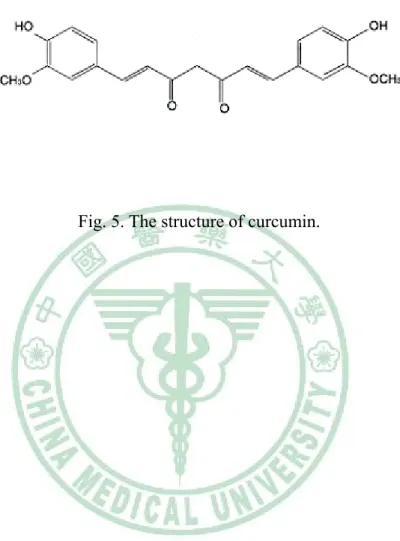
(17) inflammatory cytokines, adhesion molecules, and MMPs (Baeuerle, 1991; Grilli et al, 1993; Martin et al, 2000). The NF-κB is known regulator of inflammation, which represent key features of the response to vascular complications.. 1.5 Curcumin. The characteristic yellow color of turmeric is due to the curcuminoids, first isolated by Vogel in 1842. Curcumin is an orange-yellow crystalline powder practically insoluble in water. The structure of curcumin (C21H20O6) (Fig. 5) was first described in 1910 by Lampe and Milobedeska and shown to be diferuloylmethane (Aggarwal et al, 2003). Curcumin (diferuloylmethane) is the active yellow pigment in turmeric (the ground rhizome of Curcuma longa Linn), a popular plant-derived coloring spice and ingredient of many cosmetics and pharmaceuticals (Gafner et al, 2004; Shishodia et al, 2005; Aharma et al, 2005; Aggarwal and Shishodia, 2006; Maheshwari et al, 2006). Previous study also indicated that supplementation with Curcuma longa reduced oxidative stress and attenuates the development of fatty streaks in rabbits fed a high cholesterol diet (Quiles et al, 2002). In general, curcumin has been associated with a large number of biological and cellular activities, including antioxidative,. anti-inflammatory,. anticarcinogenic,. and. hypocholesterolemic. properties (Gafner et al, 2004; Shishodia et al, 2005; Aharma et al, 2005; Aggarwal and Shishodia, 2006; Maheshwari et al, 2006). Curcumin mediates its effects by modulating several important molecular targets, including transcription factors, enzymes, cell cycle proteins, cytokines, receptors, and cell surface adhesion molecules (Shishodia et al, 2005; Aharma et al, 2005; Aggarwal and Shishodia, 2006).. 9.
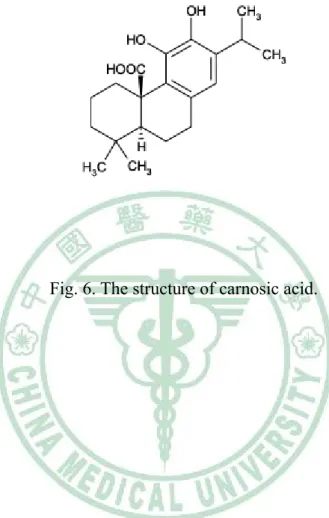
(18) 1.6. Carnosic acid (CA). Sage (Salvia officinalis L.) and rosemary (Rosmarinus officinalis L.) are popular Labiatae herbs used as spices and folk medicines around the world (Masuda et al, 2002). In the 1950s, it was reported that an extract of rosemary leaves contained high antioxidant activity (Huang et al, 2005). Extracts of Labiatae herbs can have both flavoring and antioxidant properties, and there has been particular interest in the use of rosemary extract over recent years. CA (Fig. 6) is a phenolic diterpene compound present in rosemary and sage leaves. CA has a typical O-diphenol structure and most diphenol compounds show potent chain-breaking antioxidant activity in food systems (Shahidi et al, 1992). This molecule has antimicrobial activity (Oluwatuyi et al, 2004; Moreno et al, 2006), is able to inhibit lipid absorption in humans (Ninomiya et al, 2004) and is a free radical scavenger, due to its phenolic skeleton (Masuda et al, 2001, 2002; del Bano et al, 2003). CA is a lipophilic antioxidant that scavenges singlet oxygen, hydroxyl radicals, and lipid peroxyl radicals, thus preventing lipid peroxidation and disruption of biological membranes (Aruoma et al, 1992; Haraguchi et al, 1995). Its radical scavenging activity follows a mechanism analogous to that of other antioxidants such as α-tocopherol and is caused by the presence of two O-phenolic hydroxyl groups found at C11 and C12 of the molecule (Richheimer et al, 1999).. 1.7 The objective of this research. Atherosclerosis, a progressive pathological disorder leading to cardiovascular. 10.
(19) and cerebrovascular diseases, is still the leading cause of mortality and morbidity in industrialized countries, in spite of improved pharmacological and lifestyle approaches (Ross, 1993). It is a chronic inflammatory disease driven by risk factors that cause oxidative and inflammatory mechanisms. Oxidative stress may lead to many cellular events, such as inactivation of NO, oxidative modifications of DNA and proteins, lipid oxidation, enhanced mitogenicity and apoptosis of vascular cells, and increased expression and activation of redox-sensitive genes, such as the receptor for oxidized LDL, adhesion molecules, chemotaxis factors, proinflammatory cytokines, regulators of cell cycle progression, and matrix metalloproteinases (Wassmann et al, 2004). Previous studies also indicated that proinflammatory cytokines, such as tumor necrosis factor (TNF) and interleukin-1 (IL-1), play an important role in the pathogenesis of atherosclerosis (Zhu et al, 1999; Rahman at al, 1998). The migration of SMCs from the tunica media to the subendothelial region is a key event in the development and progression of many vascular diseases including atherosclerosis and post-angioplasty restenosis (Maeda et al, 2002). MMPs activity may contribute to the pathogenesis of atherosclerosis by facilitating migration of vascular smooth muscle cells (VSMCs) (Jones et al, 2003). MMPs (MMP-9 and MMP-2) production and SMCs migration may play key roles in the pathogenesis of neointima formation and atherosclerosis. The activity of the 92 kDa (MMP-9) but not the 72 kDa (MMP-2) gelatinase is induced by IL-1α, TNF-α and phorbol esters, in a variety of cell types (Birkedal-Hansen et al, 1993; Fabunmi et al, 1996). Transcription factor NF-κB and its target genes are involved in the pathogenesis of atherosclerosis (Kutuk and Basaga, 2003). NF-κB subunits form homo- and heterodimers, the most prominent one is p50/p65 heterodimers. The dimmer is. 11.
(20) retained in the cytoplasm in an inactive state through interaction with IκB. NF-κB is rapidly activated in response to a variety of inflammatory and other stimuli that lead to degradation of IκB (Martin et al, 2000). Upon activation of NF-κB, a large number of genes are induced including various inflammatory cytokines, adhesion molecules, and MMPs (Baeuerle, 1991; Grilli et al, 1993; Martin et al, 2000). Curcumin, which is consumed daily by millions of people, is a polyphenol derived from the plant Curcuma longa. In general, curcumin has been associated with a large number of biological and cellular activities, including antioxidative, anti-inflammatory, anticarcinogenic, and hypocholesterolemic properties (Gafner et al, 2004; Shishodia et al, 2005; Aharma et al, 2005; Aggarwal and Shishodia, 2006; Maheshwari et al, 2006). In this study, we investigated the inhibitory effect of curcumin on TNF-α-induced human aortic smooth muscle cells (HASMCs) migration and MMP-9 activity. Carnosic acid (CA) is the primary phenolic compound in rosemary and salvia. Previous study indicated that CA has a typical O-diphenol structure and most diphenol compounds show potent chain-breaking antioxidant activity in food systems (Shahidi et al, 1992). This molecule has antimicrobial activity (Oluwatuyi et al, 2004; Moreno et al, 2006), is able to inhibit lipid absorption in humans (Ninomiya et al, 2004) and is a free radical scavenger, due to its phenolic skeleton (Masuda et al, 2001, 2002; del Bano et al, 2003). In this study, we investigated the inhibitory effect of CA on TNF-α-induced HASMCs migration and MMP-9 activity. In the present study, two polyphenolic compounds, Curcumin and CA, were examined for their effects on TNF-α-induced cell migration in HASMCs and also elucidate its possible mechanism.. 12.
(21) Smooth muscle cells. Adventitia. Fig. 1. The structure of the normal blood wall. (Libby, 2002). 13.
(22) Fig. 2. The function of VSMCs during different stages of atherosclerosis. (Dzau et al, 2002). 14.
(23) Fig. 3. The regulation of MMPs. (Dollery et al, 1995). 15.
(24) (Creemers et al, 2001). 16.
(25) Extracellular stimuli Bacteria. Cytokine. Viruses. Stress. IκB p50. p65. IκB Kinase. Cytoplasm P. P IκB p50. P. p65. P Ub. IκB. Ub p50. p65. IκB. p50. p65. Inflammation MMPs. Nucleus. Fig. 4. The signal transduction pathways of NF-κB activation. (Martin et al, 2000). 17.
(26) Fig. 5. The structure of curcumin.. 18.
(27) Fig. 6. The structure of carnosic acid.. 19.
(28) Chapter 2. 薑黃素抑制細胞激素誘發人類大主動脈平滑肌細胞的遷移和 基質金屬蛋白酶的活化. Curcumin inhibits MMP-9 activity and migration of TNF-α-induced human aortic smooth muscle cells. 20.
(29) 2.1 Introduction. Atherosclerosis, a progressive pathological disorder leading to cardiovascular and cerebrovascular diseases, is still the leading cause of mortality and morbidity in industrialized countries, in spite of improved pharmacological and lifestyle approaches (Ross, 1993). It is a chronic inflammatory disease driven by risk factors that cause oxidative and inflammatory mechanisms. Oxidative stress may lead to many cellular events, such as inactivation of NO, oxidative modifications of DNA and proteins, lipid oxidation, enhanced mitogenicity and apoptosis of vascular cells, and increased expression and activation of redox-sensitive genes, such as the receptor for oxidized LDL, adhesion molecules, chemotaxis factors, proinflammatory cytokines, regulators of cell cycle progression, and matrix metalloproteinases (Wassmann et al, 2004). Previous studies also indicated that proinflammatory cytokines, such as tumor necrosis factor (TNF) and interleukin-1 (IL-1), play an important role in the pathogenesis of atherosclerosis (Zhu et al, 1999; Rahman at al, 1998). The migration of SMCs from the tunica media to the subendothelial region is a key event in the development and progression of many vascular diseases including atherosclerosis and post-angioplasty restenosis (Maeda et al, 2002). MMPs activity may contribute to the pathogenesis of atherosclerosis by facilitating migration of vascular smooth muscle cells (VSMCs) (Jones et al, 2003). MMPs (MMP-9 and MMP-2) production and SMCs migration may play key roles in the pathogenesis of neointima formation and atherosclerosis. The activity of the 92 kDa (MMP-9) but not the 72 kDa (MMP-2) gelatinase is induced by IL-1α, TNF-α and phorbol esters, in a. 21.
(30) variety of cell types (Birkedal-Hansen et al, 1993; Fabunmi et al, 1996). Transcription factor NF-κB and its target genes are involved in the pathogenesis of atherosclerosis (Kutuk and Basaga, 2003). NF-κB subunits form homo- and heterodimers, the most prominent one is p50/p65 heterodimers. The dimmer is retained in the cytoplasm in an inactive state through interaction with IκB. NF-κB is rapidly activated in response to a variety of inflammatory and other stimuli that lead to degradation of IκB (Martin et al, 2000). Upon activation of NF-κB, a large number of genes are induced including various inflammatory cytokines, adhesion molecules, and MMPs (Baeuerle, 1991; Grilli et al, 1993; Martin et al, 2000). Curcumin, which is consumed daily by millions of people, is a polyphenol derived from the plant Curcuma longa. In general, curcumin has been associated with a large number of biological and cellular activities, including antioxidative, anti-inflammatory, anticarcinogenic, and hypocholesterolemic properties (Gafner et al, 2004; Shishodia et al, 2005; Aharma et al, 2005; Aggarwal and Shishodia, 2006; Maheshwari et al, 2006). In this study, we investigated the inhibitory effect of curcumin on TNF-α-induced human aortic smooth muscle cells (HASMCs) migration and MMP-9 activity.. 22.
(31) 2.2. Materials and Methods. 2.2.1 Materials. 2.2.1.1 Instruments. CO2 incubator. NUAIRE, MN, USA. Laminar flow. NUAIRE, MN, USA. Microscope. Nikon, Japan. pH meter. HANNA, RI, USA. Stirrer/Hotplate. Corning, Taiwan. Waterbath tank. TKS, Taiwan. Haemocytometer. Boeco, Germany. Eppendorf centrifugator. Hamburg, Germany. Pipetman. Gilson, France. Spectrophotometer. HITACHI, Japan. Spectrophotometer. Beckman Coulter, CA, USA. MicroPlate fluorescence reader. Bio-Tek, VT, USA. Shaking incubator. Orbital, VA, USA. ELISA plate reader. Bio-Tek, VT, USA. Electrophoresis tank. Bio-Rad, CA, USA. Transfer system. Bio-Rad, CA, USA. 23.
(32) Electrophoresis chamber. Bio-Rad, CA, USA. Power supply. Hoefer, CA, USA. 2.2.1.2 Chemicals. 40 % Acrylamide. Amresco, OH, USA. 2, 7-dichlorofluorescein diacetate (DCFH-DA). Molecular Probe, OR, USA. 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) Sigma, MO, USA Acetic acid. Amresco, OH, USA. Bovine serum albumin (BSA). Sigma, MO, USA. Bradford. Bio-Rad, CA, USA. Chloroform. BDH, Poole, England. Curcumin. Sigma, MO, USA. Diethyl pyrocarbonate (DEPC). Sigma, MO, USA. Dimethyl sulfoxide (DMSO). Amresco, OH, USA. Dodecylsulfate sodium salt (SDS). Amresco, OH, USA. Enhanced chemiluminescence (ECL). Upstate, CA, USA. Ethanol. 景明化工, Taichung, Taiwan. F12K. Sigma, MO, USA. Fetal bovine serum (FBS). Gibco, NY, USA. Gelatin. Amresco, OH, USA. Glycerol. Amresco, OH,USA. Glycine. Amresco, OH, USA. 24.
(33) Hepes. Gibco, NY, USA. Invasion assay kit. Chemicon, CA, USA. Isopropanol. Sigma, MO, USA. Methanol. Tedia, OH, USA. Nuclear extract kit. TransAM, CA, USA. NF-κB kit. TransAM, CA, USA. Penicillin-Streptomycin. Gibco, NY, USA. Recombinant human TNF-α. Cytolab, Rehovot, Israel. Sodium biocarbonate. Sigma, MO, USA. Sodium chloride. Sigma, MO, USA. Temed. Amresco, OH, USA. Tris base. Amresco, OH, USA. Tris-HCl. Amresco, OH, USA. Triton X-100. Amresco, OH, USA. Trizol. Invitrogen, CA, USA. Trypsin. Sigma, MO, USA. Trypan blue. Sigma, MO, USA. Tween-20. Amresco, OH, USA. 2.2.1.3. Antibodies. Primary antibodies Mouse anti-human β-actin. Abcam, Cambridge, UK. Rabbit anti-human matrix metallproteinases-9. Abcam, Cambridge, UK. 25.
(34) Secondary antibodies Sheep anti-mouse IgG antibody. Abcam, Cambridge, UK. Goat anti-rabbit IgG antibody. Abcam, Cambridge, UK. 26.
(35) 2.3. 2.3.1. Methods. Cell culture Human aortic smooth muscle cells (HASMCs) were purchased from Food. Industry Research and Development Institute, 新竹, Taiwan (CCRC 60293). They were maintained in Ham’s F12K containing 10 % fetal bovine serum, 2 mmol/l L-glutamine, 1.5 g/l sodium bicarbonate, 10 mmol/l HEPES, 10 mmol/l TES, 0.05 mg/ml ascorbic acid, 0.01 mg/ml transferrin, 0.01 mg/ml insulin, 10 ng/ml sodium selenite, 0.03 mg/ml ECGs. All experiments were performed with HASMCs from passages 21 to 31, which were grown to 80-90 % confluence and made quiescent by serum starvation (0.1 % FBS) for at least 24 h.. 2.3.2. Cell viability assay (MTT assay). The cytotoxic effect of curcumin on HASMCs was investigated using 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay (Chen et al, 2002). The principle of this assay is that mitochondria dehydrogenase in viable cells reduces MTT to a blue formazan. Briefly, the cells were grown in 96-well culture plates at a density of 1×104 cells per well in F-12K culture medium and incubated with various concentrations of curcumin for 24 hours. 10 μl MTT (5 mg/ml) were then added to each well and incubation continued at 37 °C for an additional 4 hours. The medium was then carefully removed, so as not to disturb the formazan crystals which had formed. Dimethyl sulphoxide (100 μl), which solubilizes formazan crystals, was. 27.
(36) added to each well and absorbance of the solubilized blue formazan was measure the optical density at 590 nm using µQuant Microplate Spectrophotometer (Bio-Tek, VT, USA). All determinations were performed according to three individual experiments. The data were shown mean±SD as percentage of control.. 2.3.3. Gelatin zymography for MMP-9. MMP-9 activity in conditioned medium of cultured HASMCs was analyzed by substrate-gel electrophoresis (zymography) using SDS-PAGE (10 %) containing 0.1 % gelatin. Substrate gel zymography of the activity of MMP-9 was performed with a Mini-Protein II apparatus from Bio-Rad, according to a method described previously (Demeule et al, 2000). Cells were grown to sub-confluence and were rinsed with phosphate-buffered saline (PBS) and then incubated in serum-free medium for 24 h. Equal volumes of samples of conditioned cell culture medium were mixed with sample buffer containing 62.5 mmol/l Tris-HCl (pH 6.8), 10 % glycerol, 2 % SDS, and 0.00625 % (w/v) bromophenol blue, loaded onto the gel and separated by electrophoresis. Thereafter, gels were washed 3 times for 30 minutes at room temperature in buffer (50 mmol/l Tris-HCl, pH 8.0, 5 mmol/l CaCl2, 0.02 % NaN3, and 2.5 % Triton X-100) and incubated for 18 h at 37 °C with the same buffer except Triton X-100. Gels were stained with Coomasssie Brillant Blue R-2500 (0.1 %) and destained in 5 % methanol and 7 % acetic acid. Gelatinolytic activity appeared as a clear band on a blue background.. 2.3.4. Bradford protein assay. 28.
(37) The Bradford assay (Bradford, 1976), a colorimetric protein assay, is based on an absorbance shift in the dye Coomassie when bound to arginine and hydrophobic amino acid residues present in protein. The anionic (bound) form of the dye is blue and has an absorption spectrum maximum historically held to be at 595 nm. The cationic (unbound) forms are green and red. The increase of absorbance at 595 nm is proportional to the amount of bound dye, and thus to the amount (concentration) of protein present in the sample. Standard solutions contain a range of 0 to 25 micrograms protein (BSA) in 800 μl H2O, followed by adding 200 μl dye reagent and incubate 5 min. l μl of sample solution add into 799 μl H2O, followed by adding 200 μl dye reagent and incubated for 5 min. The absorbance was read at 595 nm. The results made a standard curve and the protein concentration of sample was determined by standard curve.. 2.3.5. Western blot analysis. HASMCs were treated with various concentrations of curcumin in the presence of 100 ng/ml TNF-α. Cellular lysates were prepared in a lysis buffer containing 10 mmol/l Tris/HCl (pH 8), 0.32 mol/l sucrose, 5 mmol/l Ethyienediamine Teraacetate Disodium Salt (EDTA), 1 % Triton X-100, 2 mmol/l 1, 4-Dithio-D,L-thereitol (DTT), 1 mmol/l PMSF. The cells were disrupted and extracted at 4 °C for 30 min. After centrifugation at 13,000 rpm for 15 min, the supernatant was obtained as the cell lysate. Protein concentrations were measured using the bradford assay. Total protein (20 μg) were subjected to SDS-PAGE (10 %) and blotted on PVDF membranes (Shishodia et al, 2003). Nonspecific binding was blocked by soaking the membrane in PBS-Tween. 29.
(38) 20 (PBST) buffer containing 50 g/l nonfat milk. The membrane was incubated with monoclonal mouse anti-human β-actin (1:1000) and polyclonal rabbit anti-human MMP-9 (1:1000). Subsequently, the membrane was incubated with sheep anti-mouse IgG antibody (1:5000) and goat anti-rabbit IgG antibody (1:5000). The protein levels were determined using the enhanced chemiluminescence detection reagents (Upstate, CA, USA) and high performance chemiluminescence film (Amersham, IL, USA). Incubation with mouse anti-human β-actin antibody was also performed as an internal control. Results were quantified with scanning densitometer using an image analysis system with software.. 2.3.6. Preparation of nuclear extract. Nuclear protein extracts of HASMCs were prepared using a nuclear extract kit (TransAM nuclear extract kit, CA, USA). Cells were lysed in hypotonic buffer and centrifuge suspension for 30 seconds at 14,000×g in a microcentrifuge pre-cooled at 4 °C (Dschietzig et al, 2001). Then resuspend nuclear pellet in 50 μl complete lysis buffer containing 10 mmol/l DTT, lysis buffer AM2, and protease inhibitor cocktail by pipetting up and down. The suspension was incubated for 30 min on ice, and centrifuged for 10 min at 14,000×g in a microcentrifuge pre-cooled at 4 °C. Transfer supernatant and stored at -80 °C. Protein concentrations were measured using the bradford protein assay.. 2.3.7. ELISA-Based Nuclear Factor-κB Assay. 30.
(39) Additionally to gel-shift assays, an ELISA-based kit was used for quantitative detection of NF-κB activity (TransAM NF-κB kit, CA, USA). For each sample, 20 μl of nuclear extracts (5 μg protein) were used according to the instructions of the manufacturer (Yu et al, 2007). Nuclear extracts were incubated with the oligonucleotide-coated wells for 60 min. Where indicated a competitor for NF-κB binding (NF-κB wild-type consensus oligonucleotide) was added in molar excess prior to the probe. The wells were then washed and incubated with the primary antibodies for p50 and p65 for 60 min. After incubation with a horseradish peroxidase-conjugated secondary antibody, a substrate was added to produce blue colour and then for quantitation by µQuant Microplate Spectrophotometer (Bio-tek, VT, USA). The absorbance was read at 590 nm and the blank was subtracted from all measurements.. 2.3.8. Cell migration assay. VSMCs invasion through the extracellular matrix was determined by using a commercial cell invasion assay kit (Chemicon, CA, USA). HASMCs (1.5×105 cells/300 μl) were resuspended in conditioned medium collected from pretreatment with curcumin and TNF-α-treated cells for 23 hours, and added to the upper components of migration chamber (Bedoui et al, 2005). Five hundred microliters of same conditioned medium were added to the lower compartment of migration chamber. Cells without TNF-α-treated conditioned medium served as control. The migration chambers were incubated at 37 °C for 24 hours in 5 % CO2. After incubation, the inserts were removed from the wells, and the cells on the upper side of the filter were removed using cotton swabs. The filters were fixed, and stained according to the. 31.
(40) manufacturer’s instructions. The cells that invaded and were located on the underside of the inserts. Then transfer 100 μl of the dye mixture to a 96-well plate, and measure the optical density at 560 nm.. 2.3.9 Measurement of intracellular ROS. HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 hour and induced by TNF-α (100 ng/ml) for 23 hours. Then were incubated with 10 μmol/l 2,7-dichlorofluorescein (DCF) diacetate (DCFH-DA) for 30 minutes, which is converted to DCF by intracellular esterase (Kim et al, 2006). The latter was then oxidized by ROS to the highly fluorescent DCF. The fluorescence of each dish was immediately analyzed at excitation wavelength of 485 nm and emission wavelength of 528 nm by FLx800 microplate fluorescence reader (Bio-tek, VT, USA). All measurements were at least triplicated.. 32.
(41) 2.3.10 Statistical analysis Results are shown as mean±SD. Statistical analyses of MTT were performed using One-way ANOVA followed by Dunnett’s test and others were performed using One-way ANOVA followed by Duncan’s Multiple Range Test. A value of p<0.05 was considered statistically significant.. 33.
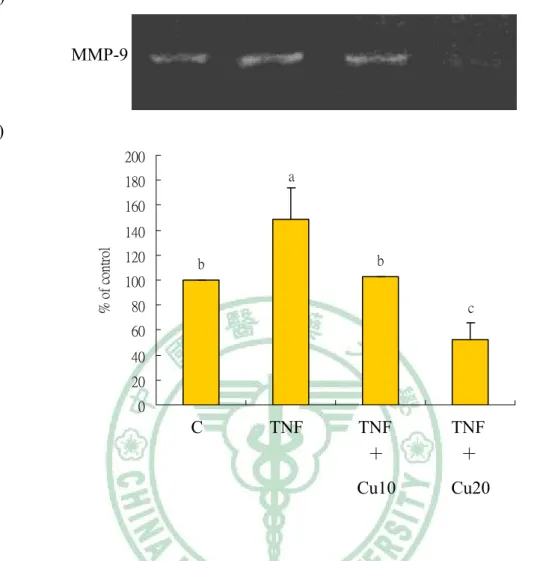
(42) 2.4 Results. 2.4.1. Cytotoxicity of curcumin on HASMCs.. The cytotoxity of curcumin on HASMCs were evaluated using MTT assay. The HASMCs (1×104 cells/well) were incubated for 24 h in cultures in 96-well with various concentrations of curcumin (0, 10, 20, 30, 50, and 75 μmol/l). Dose-dependent cytotoxic effect of curcumin against HASMCs was shown in Fig. 2-1 (100 %, 92±0.5 %, 91.2±0.8 %, 84.8±4.2 %, 81.8±6.5 %, and 67±13 %, respectively.). According to the MTT assay, we chose 10 and 20 μmol/l of curcumin to do all the following experiments.. 2.4.2. Curcumin prevents TNF-α-induced activation of MMP-9 in. HASMCs. The inhibitory effect of curcumin on TNF-α-induced MMP-9 activation were analysed by gelatin zymography. HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 h, and then induced by TNF-α (100 ng/ml) for additional 23 h. As shown in Fig. 2-2, MMP-9 secretion was markedly induced by TNF-α, and suppressed by curcumin. The 20 μmol/l curcumin treatment is more effective on activation of MMP-9 than 10 μmol/l curcumin.. 2.4.3. Curcumin suppresses TNF-α-induced MMP-9 expression in. HASMCs.. 34.
(43) The effect of MMP-9 expression by curcumin in HASMCs was assessed by Western blot. HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 h, and induced by TNF-α (100 ng/ml) for 23 h. MMP-9 expression was markedly induced by TNF-α, and suppressed by curcumin (Fig. 2-3). The 20 μmol/l curcumin treatment is more effective on protein expression of MMP-9 than 10 μmol/l curcumin.. 2.4.4. Curcumin suppresses nuclear translocation of NF-κB p50 and. p65 in TNF-α-induced HASMCs.. To determine whether the inhibitory effect of curcumin on the TNF-α-induced expression of MMP-9 is medicated via NF-κB, we measured the nuclear translocation of p50 and p65 of the NF-κB family. Treatment of TNF-α (100 ng/ml) for 23 h enhanced the nuclear translocation of p50 (Fig. 2-4) and p65 (Fig. 2-5). Pretreatment of HASMCs with 10 and 20 μmol/l curcumin prior to TNF-α stimulation did significantly prevent the nuclear translocation of p50 and p65. As shown in Fig. 2-4, the 20 μmol/l curcumin treatment is more effective on nuclear translocation of NF-κB p50 than 10 μmol/l curcumin. In Fig. 2-5, the 20 μmol/l curcumin treatment is more effective on decreased nuclear translocation of NF-κB p65 than 10 μmol/l curcumin.. 2.4.5. Curcumin suppresses TNF-α-induced HASMCs migration.. HASMCs (1.5×105 cells/300 μl) were pretreated with 10 and 20 μmol/l curcumin for 1 hour, and induced by TNF-α (100 ng/ml) for 23 h. As shown in Fig. 2-6, the migration of HASMCs was increased by TNF-α stimulation. The stimulatory. 35.
(44) effect of TNF-α was significantly reduced by curcumin pretreatment. The 20 μmol/l curcumin treatment is more effective on decreased HASMCs migration than 10 μmol/l curcumin.. 2.4.6. Curcumin suppresses TNF-α-induced ROS generation.. To characterize the events underlying TNF-α-induced migration, we examined the generation of ROS after TNF-α treatment in HASMCs. HASMCs were exposed to TNF-α (100 ng/ml) for 23 h, and DCF fluorescence produced was measured (Fig. 2-7). The production of ROS was induced by TNF-α and decreased by curcumin. The 20 μmol/l curcumin treatment is more effective on reduced ROS generation than 10 μmol/l curcumin.. 36.
(45) 2.5 Discussion. In this study, we investigated the effect of curcumin on HASMCs migration and MMP-9 activation induced by TNF-α. Curcumin lowered the secretion and protein expression of MMP-9 by gelatin zymography and Western blot assays. It also decreased nuclear translocation of nuclear factor-κB (NF-κB) P50 and P65. In addition, the migration assay showed that curcumin effectively inhibited the TNF-α-induced migration of HASMCs as compared with the control group. In our in vitro study, we also found that curcumin could scavenge DPPH (2, 2-diphenyl-1-picrylhydrazyl) radicals, alkoxyl radical (RO ), and peroxyl radical (ROO ). It is approximately 2-3-folds more potent than Trolox in antioxidative ability (Appendix 1). It also could suppress TNF-α-induced intracellular ROS production. Curcumin, which is consumed daily by millions of people, is a polyphenol derived from the plant Curcuma longa (Fang et al, 2005). It exhibits a variety of pharmacological effects including anti-tumor, anti-inflammtory, anti-infectious activities and is currently in clinical trials for AIDS patients (Mazumder et al,1995; Ruby et al, 1995; Surh, 2002). Commercial curcumin usually isolated from the rhizome of Curcuma longa Linn. which contain approximately 77 % of curcumin (Ahsan et al, 1994). Previous study indicated that the serum concentration was 1.77±1.87 μmol/l after 8 g curcumin intake in human (Cheng et al, 2001), therefore, curcumin was absorbable in digestive tract in human. In the present study, we found that 20 μmol/l of curcumin did not have any significant effect on the cytotoxity of HASMCs from MTT test. Therefore, we chose 10 and 20 μmol/l curcumin to do all. 37.
(46) the experiments (Fig. 2-1). The migration of SMCs from the tunica media to the subendothelial region is a key event in the development and progression of many vascular diseases including atherosclerosis and post-angioplasty restenosis (Maeda et al, 2002). MMPs (MMP-2 and MMP-9) activities may contribute to the pathogenesis of atherosclerosis by facilitating migration of VSMCs (Jones et al, 2003). Although MMP-2 and MMP-9 have similar substrate specificities, the regulation of their expression are different. MMP-2 is constitutively expressed by several cell types, including SMCs, and its expression is not induced by cytokines or growth factors. In contrast, MMP-9 can be induced by TNF-α in SMCs (Cho et al, 2000; Galis et al, 1994). Therefore, we have investigated the effect of curcumin on the migration of HASMCs and activation of MMP-9. The results indicated that the migration of HASMCs was significantly induced by TNF-α, and suppressed by curcumin (Fig. 2-6). This inhibition against TNF-α-induced migration of HASMCs is consistent with the inhibition of activation and expression of MMP-9 (Fig. 2-2 and Fig. 2-3). A similar result was seen when HASMCs were pretreated with other polyphenolic compound, such as tea flavonoid epigallocatechin-3-gallate (20 μmol/l), quercetin (40 μmol/l), and other flavonoids (Kim et al, 2005; Moon et al, 2003). The NF-κB family controls the expression of genes involved in the inflammation and immune response (Baeuerle, 1991). In the cytoplasm, inactive NF-κB exists as a heterodimeric complex of subunits p50 and p65 that binds to a cytoplasmic protein, IκB (Baeuerle and Henkel, 1994). Upon activation, IκB is rapidly degraded, and the p50/p65 heterodimer is translocated from the cytoplasm into the nucleus where the dimmer interacts with regulatory κB elements in promoters and. 38.
(47) enhancers, thereby controlling gene transcription (Baeuerle and Baltimore, 1988). NF-κB is activated by a multitude of stimuli, including various inflammatory cytokines and reactive oxygen intermediates (Baeuerle and Baltimore, 1998; Grilli et al, 1993; Muller et al, 1997), which are activated in atherosclerotic lesions (Brand et al, 1996). In the present study, curcumin reduced cytokine-induced nuclear translocation of p50 (Fig. 2-4) and p65 (Fig. 2-5) in HASMCs. Much further work, which is beyond the scope of the present study, is necessary to elucidate the mechanisms underlying the synergistic regulation of MMP secretion by cytokines. We focused instead on defining the role played by the NF-κB transcription factor in the regulation of MMPs in HASMCs. A functional NF-κB site occurs in the proximal stimulatory region of the MMP-9 promoter (Sato and Seiki, 1993; Fini et al, 1994) and deletion of this site reduces upregulation of reporter gene constructs in response to cytokines. Until now, however it has been unclear what contribution NF-κB plays in the upregulation of the endogenous MMP-9 gene. Previous study has demonstrated that transient overexpression of IκBα in vascular SMCs only partially impaired upregulation of MMP-9, suggesting that NF-κB simply plays a permissive role in the upregulation of MMP-9 (Bond et al, 2001). In the present study, curcumin reduced cytokine-induced expression of MMP-9 and prevented the nuclear translocation of p50 (Fig. 2-4) and p65 (Fig. 2-5) in HASMCs. We suggested that inhibitory mechanisms of curcumin might interrupt a signaling cascade involving MMP-9 transcription-mediated activation of NF-κB. Several studies have indicated that ROS are implicated in the activation of NF-κB (Muller et al, 1997). The current study shows that the ROS production stimulated by TNF-α was decreased by curcumin pretreatment in HASMCs (Fig. 2-7).. 39.
(48) Based on this result, we proposed that the inhibitory effect of curcumin on MMP-9 expression and NF-κB activation may be due to its antioxidant and anti-inflammatory properties. In our in vitro study, we also found that curcumin could scavenge DPPH (2, 2-diphenyl-1-picrylhydrazyl) radicals, alkoxyl radical (RO ), and peroxyl radical (ROO ) (Table 2-1). It is approximately 2-3-folds more potent than Trolox in antioxidative ability (Appendix 1). Since atherosclerosis is a chronic inflammatory disease associated with increased oxidative stress in the VSMCs, it would be conceivable that the anti-atherogenic effects of curcumin might due to its antioxidative and anti-inflammatory properties. The inhibition of cytokine-induced MMP-9 expression has been described for other substances with anti-inflammatory properties such as tea flavonoid epigallocatechin-3-gallate, quercetin, and other flavonoids (Kim et al, 2005; Moon et al, 2003).. 40.
(49) 2.6 Conclusion. In this study, curcumin effectively inhibited the TNF-α-induced migration of HASMCs as compared with the control group. The ROS production, MMP-9 secretion and expression, nuclear translocation of NF-κB p50 and p65 were reduced by curcumin pretreatment. These results led us to conclude that curcumin could restrict the migration of HASMCs by suppressing MMP-9 through down-regulation of NF-κB.. 41.
(50) 120. % of control. 100. *. *. *. 80 60 40 20 0 0. 10. 20. 30. 50. 75. Curcumin (μmol/l). Fig. 2-1. Cytotoxic effect of curcumin on human aortic smooth muscle cells (HASMCs) with MTT test. HASMCs were treated with increasing concentrations (10~75 μmol/l) of curcumin for 24 h in 10 % FBS-F12K. Statistical analyses of MTT were performed using One-way ANOVA followed by Dunnett’s test; n=3. *p<0.05 compared with control.. 42.
(51) (A) MMP-9. (B) 200 a. 180 160 % of control. 140 120. b. b. 100 80. c. 60 40 20 0. C. TNF. TNF +. TNF +. Cu10. Cu20. Fig. 2-2. Effect of curcumin on the MMP-9 activity of TNF-α-induced human aortic smooth muscle cells (HASMCs). HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 h, and induced by TNF-α (100 ng/ml) for additional 23 h. The activation of MMP-9 was assessed by gelatin zymography (A). Densitometric analysis was conducted with image analysis system software to quantify gelatin zymography data (B). Values are mean±SD, n=3.. a-c. Means with different letters are significantly. different compared at p<0.05. Abbreviation: C (control without TNF-α or curcumin), TNF (TNF-α), Cu10 (curcumin 10 μmol/l), Cu20 (curcumin 20 μmol/l).. 43.
(52) (A) MMP-9. β-actin (B) 180 a. 160. % of control. 140 120. b b. c. 100 80 60 40 20 0. C. TNF. TNF +. TNF +. Cu10. Cu20. Fig. 2-3. Curcumin inhibits the protein expression of MMP-9 in human aortic smooth muscle cells (HASMCs). HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 h, and induced by TNF-α (100 ng/ml) for additional 23 h. The expression of MMP-9 was assessed by western blot analysis. Representative western blot showing MMP-9 protein levels in cell lysates (top) and β-actin (bottom) (A). Densitometric analysis was conducted with image analysis system software to quantify western blot data (B). Values are mean±SD, n=3.. a-c. Means with different letters are significantly. different compared at p<0.05. Abbreviation: C (control without TNF-α or curcumin), TNF (TNF-α), Cu10 (curcumin 10 μmol/l), Cu20 (curcumin 20 μmol/l).. 44.
(53) 450. a. 400. % of control. 350 300. b. 250 200 150. c. c. 100 50 0. C. TNF. TNF +. TNF +. Cu10. Cu20. Fig. 2-4. Effect of curcumin on TNF-α-induced activation of NF-κB p50 in human aortic smooth muscle cells (HASMCs). HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 h and induced by TNF-α (100 ng/ml) for 23 h. Nuclear extracts were prepared and analyzed for activation of NF-κB family. Five micrograms of nuclear protein was used in each experiment. Values are mean±SD, n=3. a-dMeans with different letters are significantly different compared at p<0.05. Abbreviation: C (control without TNF-α or curcumin), TNF (TNF-α), Cu10 (curcumin 10 μmol/l), Cu20 (curcumin 20 μmol/l).. 45.
(54) a. 500. % of control. 400 b. 300. c. 200 d 100 0. C. TNF. TNF +. TNF +. Cu10. Cu20. Fig. 2-5. Effect of curcumin on TNF-α-induced activation of NF-κB p65 in human aortic smooth muscle cells (HASMCs). HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 h and induced by TNF-α (100 ng/ml) for 23 h. Nuclear extracts were prepared and analyzed for activation of NF-κB family. Five micrograms of nuclear protein was used in each experiment. Values are mean±SD, n=3. a-dMeans with different letters are significantly different compared at p<0.05. Abbreviation: C (control without TNF-α or curcumin), TNF (TNF-α), Cu10 (curcumin 10 μmol/l), Cu20 (curcumin 20 μmol/l).. 46.
(55) (A) (a). (b). (c). (d). 47.
(56) (B) 250 Migrated cells (% of control). a 200 150 b. b. c. 100 50 0. C. TNF. TNF +. TNF +. Cu10. Cu20. Fig. 2-6. Effect of curcumin on migrarion of human aortic smooth muscle cells (HASMCs) induced by TNF-α treatment. (A) Microphotographs of migrated cells without TNF-α or curcumin (a), with TNF-α (100 ng/ml) (b), with TNF-α (100 ng/ml) and 10 μmol/l of curcumin (c), and with TNF-α (100 ng/ml) and 20 μmol/l of curcumin (d) were captured. (B) HASMCs (1.5×105 cells/300 μl) were resuspended in conditioned medium collected from TNF-α treated cells for 23 h, and added to the upper components of migration chamber in the presence of 10 and 20 μmol/l curcumin. Values are mean±SD, n=3.. a-c. Means with different letters are significantly different. compared at p<0.05. Abbreviation: C (control without TNF-α or curcumin), TNF (TNF-α), Cu10 (curcumin 10 μmol/l), Cu20 (curcumin 20 μmol/l).. 48.
(57) (A) (a). (b). (c). (d). 49.
(58) (B) 160 a. 140. % of control. 120. b c. c. 100 80 60 40 20 0. C. TNF. TNF +. TNF +. Cu10. Cu20. Fig. 2-7. Effect of curcumin on TNF-α-induced ROS production in human aortic smooth muscle cells (HASMCs). (A) Microphotographs of ROS production in HASMCs without TNF-α or curcumin (a), with TNF-α (100 ng/ml) (b), with TNF-α (100 ng/ml) and 10 μmol/l of curcumin (c), and with TNF-α (100 ng/ml) and 20 μmol/l of curcumin (d) were captured. (B) HASMCs were pretreated with 10 and 20 μmol/l curcumin for 1 h and induced by TNF-α (100 ng/ml) for 23 h. Values are mean±SD, n=3.. a-c. Means with different letters are significantly different compared at p<0.05.. Abbreviation: C (control without TNF-α or curcumin), TNF (TNF-α), Cu10 (curcumin 10 μmol/l), Cu20 (curcumin 20 μmol/l).. 50.
(59) Chapter 3. 鼠尾草酸抑制細胞激素誘發人類大主動脈平滑肌細胞的遷移 和基質金屬蛋白酶的活化. Carnosic acid inhibits MMP-9 activity and migration of TNF-α-induced human aortic smooth muscle cells. 51.
(60) 3.1. Introduction. Atherosclerosis, a progressive pathological disorder leading to cardiovascular and cerebrovascular diseases, is still the leading cause of mortality and morbidity in industrialized countries, in spite of improved pharmacological and lifestyle approaches (Ross, 1993). It is a chronic inflammatory disease driven by risk factors that cause oxidative and inflammatory mechanisms. Oxidative stress may lead to many cellular events, such as inactivation of NO, oxidative modifications of DNA and proteins, lipid oxidation, enhanced mitogenicity and apoptosis of vascular cells, and increased expression and activation of redox-sensitive genes, such as the receptor for oxidized LDL, adhesion molecules, chemotaxis factors, proinflammatory cytokines, regulators of cell cycle progression, and matrix metalloproteinases (Wassmann et al, 2004). Previous studies also indicated that proinflammatory cytokines, such as tumor necrosis factor (TNF) and interleukin-1 (IL-1), play an important role in the pathogenesis of atherosclerosis (Zhu et al, 1999; Rahman at al, 1998). The migration of SMCs from the tunica media to the subendothelial region is a key event in the development and progression of many vascular diseases including atherosclerosis and post-angioplasty restenosis (Maeda et al, 2002). MMPs activity may contribute to the pathogenesis of atherosclerosis by facilitating migration of VSMCs (Jones et al, 2003). MMPs (MMP-9 and MMP-2) production and SMCs migration may play key roles in the pathogenesis of neointima formation and atherosclerosis. The activity of the 92 kDa (MMP-9) but not the 72 kDa (MMP-2) gelatinase is induced by IL-1α, TNF-α and phorbol esters, in a variety of cell types. 52.
(61) (Birkedal-Hansen et al, 1993; Fabunmi et al, 1996). Transcription factor NF-κB and its target genes are involved in the pathogenesis of atherosclerosis (Kutuk and Basaga, 2003). NF-κB subunits form homo- and heterodimers, the most prominent one is p50/p65 heterodimers. The dimmer is retained in the cytoplasm in an inactive state through interaction with IκB. NF-κB is rapidly activated in response to variety of inflammatory and other stimuli that lead to degradation of IκB (Martin et al, 2000). Upon activation of NF-κB, a large number of genes are induced including various inflammatory cytokines, adhesion molecules, and MMPs (Baeuerle, 1991; Grilli et al, 1993; Martin et al, 2000). CA is the primary phenolic compound in rosemary and salvia. Previous study indicated that CA has a typical O-diphenol structure and most diphenol compounds show potent chain-breaking antioxidant activity in food systems (Shahidi et al, 1992). This molecule has antimicrobial activity (Oluwatuyi et al, 2004; Moreno et al, 2006), is able to inhibit lipid absorption in humans (Ninomiya et al, 2004) and is a free radical scavenger, due to its phenolic skeleton (Masuda et al, 2001, 2002; del Bano et al, 2003). In this study, we investigated the inhibitory effect of CA on TNF-α-induced HASMCs migration and MMP-9 activity.. 53.
(62) 3.2. Materials and Methods. 3.2.1 Materials. 3.2.1.1 Instruments. CO2 incubator. NUAIRE, MN, USA. Laminar flow. NUAIRE, MN, USA. Microscope. Nikon, Japan. pH meter. HANNA, RI, USA. Stirrer/Hotplate. Corning, Taiwan. Waterbath tank. TKS, Taiwan. Haemocytometer. Boeco, Germany. Eppendorf centrifugator. Hamburg, Germany. Pipetman. Gilson, France. Spectrophotometer. HITACHI, Japan. Spectrophotometer. Beckman Coulter, CA, USA. MicroPlate fluorescence reader. Bio-Tek, VT, USA. Shaking incubator. Orbital, VA, USA. ELISA plate reader. Bio-Tek, VT, USA. Electrophoresis tank. Bio-Rad, CA, USA. Transfer system. Bio-Rad, CA, USA. 54.
(63) Electrophoresis chamber. Bio-Rad, CA, USA. Power supply. Hoefer, CA, USA. 3.2.1.2 Chemicals. 40 % Acrylamide. Amresco, OH, USA. 2, 7-dichlorofluorescein diacetate (DCFH-DA). Molecular Probe, OR, USA. 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) Sigma, MO, USA Acetic acid. Amresco, OH, USA. Bovine serum albumin (BSA). Sigma, MO, USA. Bradford. Bio-Rad, CA, USA. CA. Sigma, MO, USA. Chloroform. BDH, Poole, England. Diethyl pyrocarbonate (DEPC). Sigma, MO, USA. Dimethyl sulfoxide (DMSO). Amresco, OH, USA. Dodecylsulfate sodium salt (SDS). Amresco, OH, USA. Enhanced chemiluminescence (ECL). Upstate, CA, USA. Ethanol. 景明化工, Taichung, Taiwan. F12K. Sigma, MO, USA. Fetal bovine serum (FBS). Gibco, NY, USA. Gelatin. Amresco, OH, USA. Glycerol. Amresco, OH,USA. Glycine. Amresco, OH, USA. 55.
(64) Hepes. Gibco, NY, USA. Invasion assay kit. Chemicon, CA, USA. Isopropanol. Sigma, MO, USA. Methanol. Tedia, OH, USA. Nuclear extract kit. TransAM, CA, USA. NF-κB kit. TransAM, CA, USA. Penicillin-Streptomycin. Gibco, NY, USA. Recombinant human TNF-α. Cytolab, Rehovot, Israel. Sodium biocarbonate. Sigma, MO, USA. Sodium chloride. Sigma, MO, USA. Temed. Amresco, OH, USA. Tris base. Amresco, OH, USA. Tris-HCl. Amresco, OH, USA. Triton X-100. Amresco, OH, USA. Trizol. Invitrogen, CA, USA. Trypsin. Sigma, MO, USA. Trypan blue. Sigma, MO, USA. Tween-20. Amresco, OH, USA. 3.2.1.3. Antibodies. Primary antibodies Mouse anti-human β-actin. Abcam, Cambridge, UK. Rabbit anti-human matrix metallproteinases-9. Abcam, Cambridge, UK. 56.
(65) Secondary antibodies Sheep anti-mouse IgG antibody. Abcam, Cambridge, UK. Goat anti-rabbit IgG antibody. Abcam, Cambridge, UK. 57.
(66) 3.3. 3.3.1. Methods. DPPH scavenging assay. Free radical scavenging effect was determined using the free radical generator DPPH (2, 2-diphenyl-1-picrylhydrazyl) by a similar method to Yamaguchi et al, 1998. Briefly, the reaction mixture contained 500 μl of CA concentrations (0-200 μmol/l) and 500 μl of DPPH (0.5 mmol/l in methanolic solution). The DPPH radical scavenging activity was evaluated by measuring the decrease of DPPH radical detected at 517 nm and by determining the difference in the peak area between control and reaction mixture. Inhibition % was calculated via Eq. Inhibition % = (B1-B0/B0) × 100 (where B0: is the absorbance of control. B1: is the absorbance of reaction mixture.) The decoloration was plotted against the sample extract concentration in order to calculate the IC50 values (inhibitory concentration 50 μmol/l), which is the amount of sample necessary to decrease the absorbance of DPPH by 50 %.. 3.3.2 Trolox equivalent antioxidant capacity (TEAC) assay. The assay was carried out using a spectrophotometer by the improved ABTS + method as described by Re et al, 1999 with slight modification. Briefly, ABTS + radical cation. 58.
(67) was generated by a reaction of 7 mmol/l 2,2’-azinobis(3-ethyl-benzothiazoline-6sulfonicacid-diammoniumsalt) (ABTS) and 2.45 mmol/l potassium persulfate. The reaction mixture was allowed to stand in the dark for 16 h at room temperature and used within 2 days. The ABTS + solution was diluted with ethanol to an absorbance of 0.700±0.050 at 734 nm. All samples were diluted appropriately to provide 20-80 % inhibition of control absorbance. Fifty microliters of the diluted sample were mixed with 1.9 ml of diluted ABTS + solution. The assay with the mixture was carried out in triplicate, the mixture was allowed to stand for 6 min at room temperature and the absorbance was immediately recorded at 734 nm (Li et al, 2007). Trolox solution (final concentration 0~15 μmol/l) was used as a reference standard. The results were expressed as μmol/l Trolox of CA.. 3.3.3 Isolation of low density lipoproteins. Blood was collected from healthy donors after a 12 h overnight fasting using EDTA as anticoagulant. Plasma was obtained after low-speed centrifugation of the blood, adjusted to a density of 1.21 g/ml with KBr and a discontinuous density gradient was made by overlaying the plasma solution with a 10 mmol/l PBS and 1 mmol/l EDTA pH 7.4. LDL were isolated after ultracentrifugation as described previously (Vieira et al, 1996). The LDL fraction was exhaustively dialyzed against PBS buffer pH 7.4 without EDTA in the dark, filtered through a 0.45 μm filter, stored at 4 °C under nitrogen and used in 24-72 h.. 3.3.4 Inhibition of LDL Oxidation 59.
(68) LDL were oxidised using the classical copper-induced LDL auto-oxidation. Incubations were carried out at 37 °C for measuring the diene conjugated formation (Vieira et al, 1996). Increasing concentrations of CA (0~10 μmol/l) dissolved in DMSO were present in the incubation media. Briefly, 0.9 mg/ml of LDL total cholesterol was incubated in PBS in the presence of CuSO4 (50 μmol/l). After incubation, 150 µL EDTA (2 mmol/l) was added. A 100 µL portion of the mixture was then transferred to vials containin 0.9 ml of 2-propanol. The precipitates were removed by centrifugation. The concentration of conjugated diene in the supernatant was determined by absorption at 234 nm.. 3.3.5. Cell culture HASMCs were purchased from Food Industry Research and Development. Institute, 新竹, Taiwan. (CCRC 60293). They were maintained in Ham’s F12K containing 10 % fetal bovine serum, 2 mmol/l L-glutamine, 1.5 g/l sodium bicarbonate, 10 mmol/l HEPES, 10 mmol/l TES, 0.05 mg/ml ascorbic acid, 0.01 mg/ml transferrin, 0.01 mg/ml insulin, 10 ng/ml sodium selenite, 0.03 mg/ml ECGs. All experiments were performed with HASMCs from passages 21 to 31, which were grown to 80-90 % confluence and made quiescent by serum starvation (0.1 % FBS) for at least 24 hours.. 3.3.6. Cell viability assay (MTT assay). The cytotoxic effect of CA on HASMCs was investigated using 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay (Chen et al,. 60.
(69) 2002). The principle of this assay is that mitochondria dehydrogenase in viable cells reduces MTT to a blue formazan. Briefly, the cells were grown in 96-well culture plates at a density of 1×104 cells per well in F-12K culture medium and incubated with various concentrations of CA for 24 hours. 10 μl MTT (5 mg/ml) were then added to each well and incubation continued at 37 °C for an additional 4 hours. The medium was then carefully removed, so as not to disturb the formazan crystals which had formed. Dimethyl sulphoxide (100 μl), which solubilizes formazan crystals, was added to each well and absorbance of the solubilized blue formazan was measure the optical density at 590 nm using µQuant Microplate Spectrophotometer (Bio-Tek, VT, USA). All determinations were performed according to three individual experiments. The data were shown mean±SD as percentage of control.. 3.3.7. Gelatin zymography for MMP-9. MMP-9 activity in conditioned medium of cultured HASMCs was analyzed by substrate-gel electrophoresis (zymography) using SDS-PAGE (10 %) containing 0.1 % gelatin. Substrate gel zymography of the activity of MMP-9 was performed with a Mini-Protein II apparatus from Bio-Rad, according to a method described previously (Demeule et al, 2000). Cells were grown to sub-confluence and were rinsed with phosphate-buffered saline (PBS) and then incubated in serum-free medium for 24 h. Equal volumes of samples of conditioned cell culture medium were mixed with sample buffer containing 62.5 mmol/l Tris-HCl (pH 6.8), 10 % glycerol, 2 % SDS, and 0.00625 % (w/v) bromophenol blue, loaded onto the gel and separated by electrophoresis. Thereafter, gels were washed 3 times for 30 minutes at room. 61.
數據


+7


相關文件
• 由於細胞代謝、紫外線的電離輻射、不良的生 活習慣 ( 煙、酒、肥胖 ) 、各種感染物所引起 的中性粒細胞、巨噬細胞的激活, 會催化分子 氧發生單價還原產生的 "
SF12144A 張基晟 比較 Nanoplatin 併用 Pemetrexed 和 Cisplatin 併用 Pemetrexed 作為第一線治 療用於非鱗狀細胞性之非小細胞肺癌
the lymphocyte function-associated antigen 1, or LFA-1, was so named because antibodies recognizing this structure interfere with lymphocyte cell adhesion events and
一般飲食中,都會攝取些微的葉黃素,由於葉黃體
• 是細胞不正常增生,且這些增生的細胞可
基因編輯技術以人工核酸酶辨識特定 DNA 位置,並於此處切斷雙股 DNA。DNA 斷
肥大细胞内 Ca 2+ 增加可导致过敏介质释放, Ca 2+ 进入 胞浆也可导致支气管平滑肌收缩. 钙通道阻滞剂 ——抑制Ca
2.滴加蒸餾水或亞甲藍液製成水埋標本 3.觀察 石細胞 (厚壁細胞)及
