行政院國家科學委員會專題研究計畫 期中進度報告
在胞內菌感染中宿主細胞凋亡及樹突細胞作交叉抗原呈獻
及 CD8 T 細胞活化的機制(2/3)
期中進度報告(精簡版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2320-B-002-027- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學醫學院免疫學研究所 計 畫 主 持 人 : 伍安怡 處 理 方 式 : 期中報告不提供公開查詢中 華 民 國 96 年 05 月 31 日
行政院國家科學委員會補助專題研究計畫
□ 成 果 報 告
■期中進度報告
在胞內菌感染中宿主細胞凋亡及樹突細胞作交叉抗原呈獻及
CD8 T 細胞活化的機制(2/3)
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 95-2320-B-002-027
執行期間:2006 年 08 月 01 日至 2007 年 07 月 31 日
計畫主持人:伍安怡
共同主持人:
計畫參與人員:林志萱、林洋鼎、謝式弘、謝清智
成果報告類型(依經費核定清單規定繳交):■精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:台大醫學院免疫所
中 華 民 國 96 年 5 月 31 日
中文摘要
關鍵字:CD8 T 細胞,巨噬細胞細胞凋亡,抗原交叉呈獻,胞內病源菌,組織胞漿菌 CD4 T 細胞在對抗巨噬細胞內之非病毒性病原菌扮演重要角色。組織胞漿菌為主要居住在 巨噬細胞中之雙型態真菌。我們研究發現CD4 以及 CD8 T 細胞在組織胞漿菌感染後均會 被活化具有功能。我們証明巨噬細胞在吞噬此病菌後會引發細胞凋亡;樹突狀細胞吞噬被 感染之凋亡巨噬細胞可在體外實驗中活化CD8 T 細胞。因此組織胞漿菌特異性 CD8 T 細 胞是被表現外源性組織胞漿菌抗原之樹突狀細胞所活化的,此過程稱為”抗原交叉呈獻”。 為了更進一步探討抗原交叉呈獻在活體內的重要性,我們利用皮下路徑注射組織胞漿菌來 免疫老鼠。我們發現皮下注射活的組織胞漿菌會引起CD8 及 CD4 T 細胞產生干擾素γ。 巨噬細胞與嗜中性球會聚集到注射處並死亡。利用吞噬組織胞漿菌之凋亡巨噬細胞來皮下 免疫老鼠,可引起淋巴結中CD8 T 細胞的活化。我們目前正在深入探討皮下接種含抗原之 凋亡巨噬細胞是否會產生具保護性免疫反應來對抗致死劑量感染。此結果將會在接下來的 二個月寫成論文準備投稿。Abstract
Key words: CD8 T cell, macrophage apoptosis, cross-presentation, intracellular pathogen, Histoplasma capsulatum
It is widely accepted that cellular immune response via the activation of CD4 T cells plays an important role in the protection against endosomal intracellular non-viral pathogens of the macrophages. Histoplasma capsulatum is a dimorphic fungus which takes residence primarily within the macrophage. We found that not only CD4 but also CD8 T cells are activated and functional in Histoplasma infection. We demonstrated that uptake of live Histoplasma induces macrophage apoptosis and dendritic cells engulfing Histoplasma-infected apoptotic macrophages stimulate sensitized CD8 T cells in vitro. Therefore, Histoplasma-specific CD8 T cells are activated by dendritic cells that present exogenous Histoplasma antigens, a process known as ‘cross-presentation’. To further examine the in vivo significance of cross-presentation, we immunized mice with Histoplasma yeasts by the subcutaneous route. We found that injection of viable Histoplasma yeasts induces CD8 as well as CD4 T cell IFNγ production. Macrophages and neutrophils are recruited to the injection site and undergo apoptosis. Subcutaneous immunization of Histoplasma- pulsed apoptotic macrophages induces CD8 T cell activation in the draining lymph nodes. The study on whether immunization by antigen-pulsed apoptotic cells generates protective immunity against lethal challenge is ongoing. The results will be written up to publication in the next two months.
Introduction
It is widely accepted that cellular immune response via the activation of CD4 T cells plays an important role in the protection against endosomal intracellular non-viral pathogens of the macrophages. In our previous studies, we demonstrated that infection with Histoplasma capsulatum, an intracellular pathogen of the macrophage, activated not only CD4 but also CD8 T cells (1). CD8 T cells are important in limiting fungal replication in the absence of CD4 T cells to protect mice against histoplasmosis (2). The macrophage, being the primary host cell of Histoplasma, is induced to undergo apoptosis after engulfing the pathogen. The apoptotic macrophage can serve as antigen donor to dendritic cells. Histoplasma-specific CD8 T cells are stimulated by dendritic cells that present exogenous Histoplasma antigens, either through direct ingestion of yeasts or through uptake of apoptotic macrophage-associated fungal antigens, a process known as ‘cross-presentation’ (1). In addition to Histoplasma infection, it has been reported in Salmonella, Mycobacterium, and Shigella bacterial infections that intracellular non-viral pathogens induce macrophage apoptosis (3-8). DCs are capable of cross-presenting cell-associated antigens derived from apoptotic cells containing microbial antigens (6-8). Our previous work has focused on showing cross-presentation is an in vitro system; the in vivo significance of cross-presentation still awaits to be elucidated.
The importance of CD8 T cells and dispensability of CD4 T cells was reported in vaccine-induced immunity against Blastomyces and Histoplasma (9). Mice are resistant to lethal challenge after vaccination by high inocula of live yeasts at subcutaneous sites. Depleting CD4+ T cells does not reduce resistance to lethal challenge. The injection of live yeast cells at subcutaneous sites induces efficient CD8+ T cell-mediated protection. However, these experiments still did not address the question whether CD8 T cells are activated and whether cross-presentation of fungal antigens occurs in vivo. It was recently demonstrated that apoptotic vesicles from Mycobacteria-infected macrophages stimulate CD8 T cells in vivo (10). Vaccination with apoptotic vesicles from Mycobacteria-infected macrophages induces protection against tuberculosis compared to treatment with naïve vesicles. Therefore, the apoptotic bodies induce cross-presentation in vivo and mediate host defense against infection.
Based on these findings, we further our study to examine the in vivo significance of cross-presentation of Histoplasma fungal antigens. We immunized mice with Histoplasma yeasts by the subcutaneous route. We found that injection of live, but not heat-killed Histoplasma induced strong CD8 T cell response. Subcutaneous inoculation of viable Histoplasma induces
massive macrophage and neutrophil infiltration and the infiltrating cells become apoptotic. Furthermore, subcutaneous immunization by antigen-pulsed apoptotic cells also induced significant CD8 T cell response. Experiments are underway to study whether immunization by antigen-pulsed apoptotic cells generates protective immunity against lethal challenge.
Results
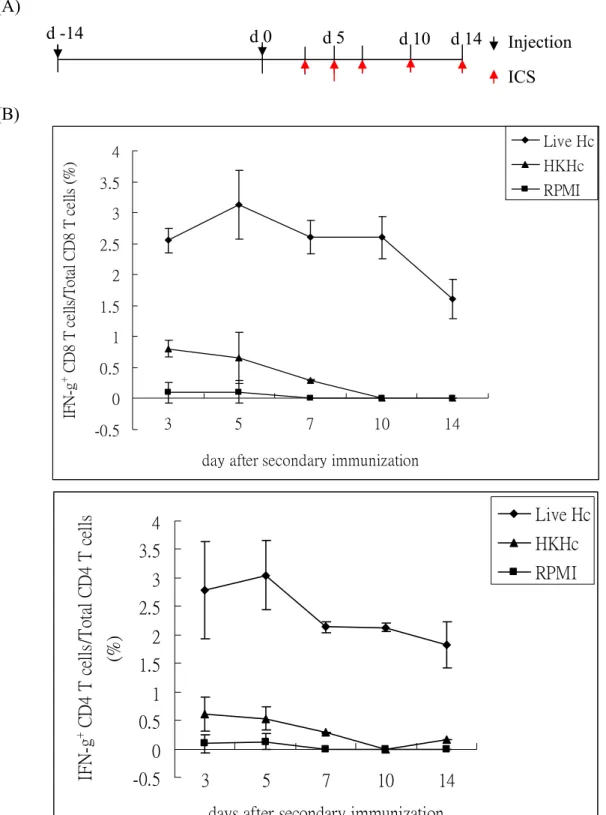
1. Subcutaneous injection of live Histoplasma induces CD8 as well as CD4 T cell activation. It was recently reported that injection of live yeast cells at subcutaneous sites induces efficient CD8 T cell-mediated immunity (9). Thus, we used subcutaneous injection of Histoplasma in this experiment and addressed whether CD8 T cells are activated. One million Histoplasma yeast cells were suspend in 50 μl RPMI 1640 solution and injected into the two sides of subcutaneous tissue of tail base (2 injection per mouse) at day-14 and day0. At 5 days after second injection, mice were sacrificed and the cells from draining lymph nodes were harvested. The cells were subjected to intracellular cytokine staining for IFNγ expression
(Figure 1A). As shown in Figure 1B, both CD8 and CD4 T cells in the draining lymph nodes
of mice subcutaneously injected with live Histoplasma yeasts (Live Hc) were activated and produced IFNγ after stimulation, while T cells from mice subcutaneously injected with heat-killed Histoplasma (HKHc) or medium only control (RPMI) were not activated. The kinetics of CD8 and CD4 T cell activation at different time points after injection was shown in Figure 2. The T cells were activated to produce IFNγ during the course of injection, and the percentage of IFNγ-producing T cell reached the maximal at day5 after second injection
(Figure 2B and 2C). We have shown that in mice with intravenously systemic infection of
Histoplasma, the percentage of IFNγ+ CD4 T cells is two to three times than that of IFNγ+
CD8 T cells (1). However, subcutaneous injection of live Histoplasma induced almost equal percentage and number of IFNγ+ CD8 and CD4 T cells (Figure 2B and 2C). These results suggest that subcutaneous injection is a potent route to stimulate CD8 T cell activation.
2. Subcutaneous injection of live Histoplasma recruits macrophage and neutrophil to the injection sites. Next, mice subcutaneously injected two times with live, heat-killed Histoplasma, or RPMI were sacrificed, and paraffin sections and cryosections of injection sites were prepared. In the H&E staining of cryosection sample, it can be observed that lots of cells were infiltrated into the injection site of mice inoculated with live Histoplasma (Figure
3A). In contrast, there was few or no cell infiltration in mice with HKHc or RPMI inoculation.
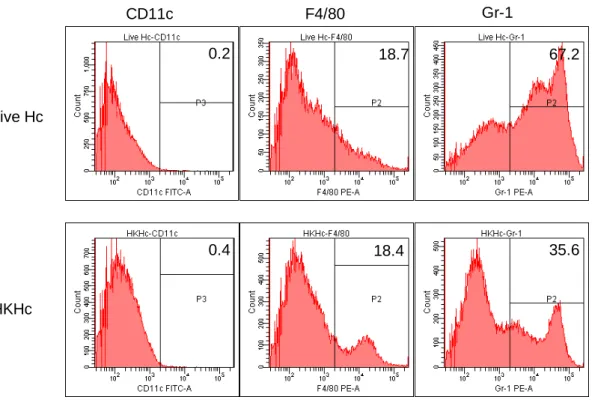
To further analyze what cell types were infiltrated, the cyrosection sample was stained with monoclonal antibodies against F4/80, Gr-1, CD3, or B220. Figure 3B shows that most infiltrating cells were F4/80+ and Gr-1+. These cells also expressed high level of CD11b (data not shown). There were few T or B cells in these cell populations. To quantify the constitution
of these cells, infiltrating cells in injection sites were isolated by centrifugation after tissue were treated with type 4 collagenase. Figure 3C shows that 67.2% and 18.7% of infiltrating cells in mice with live Hc injection were Gr-1+ and F4/80+, respectively. Although there were very few cells infiltrating into the injection site of mice injected with HKHc, 35.6% were Gr-1+ and 18.4% were F4/80+. Taken together, it indicates that macrophages and neutrophils are the major cell populations infiltrating into Histoplasma injection sites.
3. Subcutaneous injection of Histoplasma yeasts induces cell apoptosis in vivo. We have demonstrated that engulfing live Histoplasma yeasts induce macrophage apoptosis in vitro. The dendritic cells which engulfed Ag-pulsed apoptotic macrophages were efficient in stimulating CD8 T cell responses (2). To further examine whether subcutaneous injection of live Histoplasma yeasts induce apoptosis in vivo, we used TUNEL staining to detect apoptotic cells in paraffin section sample of injection sites. Subcutaneous injection of live or heat-killed Histoplasma but not RPMI induced cell apoptosis at day5 after primary injection (data not shown). By flow cytometric analysis of isolated infiltrating cells, 73.7% of cells from mice with live Hc injection were TUNEL+ apoptotic cells, while only 8.5% of cells from mice with HKHc injection were TUNEL+ (data not shown). Furthermore, Figure 4 shows that the apoptotic cells were Gr-1+ (Figure 4, A-D) or F4/80+ (Figure 4, E-H). Taken together, these results indicate that inoculation of viable Histoplasma into subcutaneous tissue induces massive macrophage and neutrophil infiltration and cell apoptosis.
4. Injection of HKHc-pulsed apoptotic cells induces functional CD8 T cell in vivo. The next question was whether immunization by apoptotic cells pulsed with Histoplasma antigens through the subcutaneous route induces CD8 T cell responses. Thioglycolate-elicited peritoneal macrophages were harvested and allowed to ingest heat-killed Histoplasma yeasts. After wash to get rid of undigested yeasts, the macrophages were treated with LPS and ATP to induce cell apoptosis (2). First, mice were injected with different doses of HKHc-pulsed apoptotic macrophages through the subcutaneous route, and CD8 T cell response was evaluate by intracytoplasmic cytokine staining. Mice injected with live Hc, HKHc or RPMI were also included. Figure 5 shows that CD8 T cells from mice injected with HKHc-pulsed apoptotic macrophages were activated to produce IFNγ. The number of IFNγ-producing CD8 T cells was dose-dependent on the number of apoptotic cells injected. However, injection of HKHc-pulsed apoptotic macrophages was not as efficient as injection of live Hc in inducing CD8 T cell IFN γ production (Figure 5). The result implies that
cross-presentation of antigens acquired from antigen-pulsed apoptotic macrophages occurs in vivo. It also suggests that apoptotic cell-induced cross-priming may not the only way to elicit CD8 T cell activation after live Hc injection.
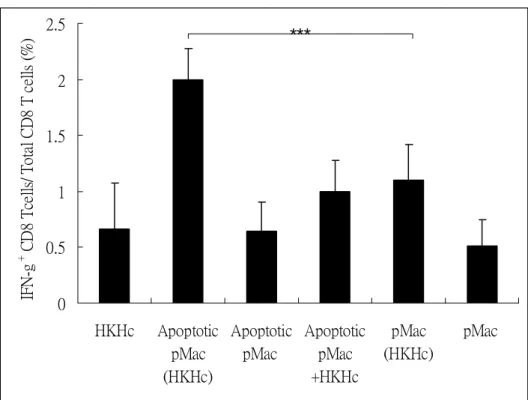
5. Cell apoptosis is important for CD8 T cell cross-priming in vivo. To further investigate the role of cell apoptosis in CD8 T cell activation, macrophages were cocultured with or without HKHc and then induce apoptosis or not. These cells were injected into subcutaneous sites of mice, and IFNγ-producing CD8 T cells were detected. The results in Figure 5 show that injection of HKHc-pulsed apoptotic macrophages [Apoptotic pMac (HKHc)] efficiently induced CD8 T cell IFNγ production. In contrast, HKHc-pulsed non-apoptotic macrophages [pMac (HKHc)] did not further trigger T cell response as compared to that of HKHc injection [HKHc]. Neither macrophages [pMac] nor apoptotic macrophages [Apoptotic pMac] without antigen pulse stimulated IFNγ-producing CD8 T cells. These results suggest that cell apoptosis and antigen-containing are important for CD8 T cell activation in vivo. Moreover, it has been published that apoptotic cell can serve as adjuvant rather than antigen processing cell to activate TLR2 signaling (10). Thus, we injected apoptotic macrophages mixed but not cocultuered with HKHc [Apoptotic pMac+HKHc] to study whether Histoplasma antigens required for CD8 T cell cross-priming need to be processed in apoptotic cell. Results showed that injection of apoptotic macrophages mixed with HKHc was not any more efficient than that injection of HKHc in stimulating IFNγ+ CD8 T cells (Figure 5). This confirms cell process of antigen is important for cross-presentation. Hence, we proposed that Histoplasma antigens which are processed in apoptotic cells are the major sources for cross-presentation and cross-priming.
References
1. Lin, J. S., and B. A. Wu-Hsieh. 2004. Functional T cells in primary immune response to histoplasmosis. Int Immunol 16:1663-1673.
2. Lin, J. S., C. W. Yang, D. W. Wang, and B. A. Wu-Hsieh. 2005. Dendritic cells
cross-present exogenous fungal antigens to stimulate a protective CD8 T cell response in infection by Histoplasma capsulatum. J Immunol 174:6282-6291.
3. Rojas, M., L. F. Barrera, G. Puzo, and L. F. Garcia. 1997. Differential induction of apoptosis by virulent Mycobacterium tuberculosis in resistant and susceptible murine macrophages: role of nitric oxide and mycobacterial products. J Immunol 159:1352-1361. 4. Klingler, K., K. M. Tchou-Wong, O. Brandli, C. Aston, R. Kim, C. Chi, and W. N. Rom.
1997. Effects of mycobacteria on regulation of apoptosis in mononuclear phagocytes. Infect Immun 65:5272-5278.
5. Navarre, W. W., and A. Zychlinsky. 2000. Pathogen-induced apoptosis of macrophages: a common end for different pathogenic strategies. Cell Microbiol 2:265-273.
6. Yrlid, U., and M. J. Wick. 2000. Salmonella-induced apoptosis of infected macrophages results in presentation of a bacteria-encoded antigen after uptake by bystander dendritic cells. J Exp Med 191:613-624.
7. Yrlid, U., M. Svensson, A. Kirby, and M. J. Wick. 2001. Antigen-presenting cells and anti-Salmonella immunity. Microbes Infect 3:1239-1248.
8. Schaible, U. E., F. Winau, P. A. Sieling, K. Fischer, H. L. Collins, K. Hagens, R. L. Modlin, V. Brinkmann, and S. H. Kaufmann. 2003. Apoptosis facilitates antigen presentation to T lymphocytes through MHC-I and CD1 in tuberculosis. Nat Med 9:1039-1046.
9. Wuthrich, M., H. I. Filutowicz, T. Warner, G. S. Deepe, Jr., and B. S. Klein. 2003. Vaccine immunity to pathogenic fungi overcomes the requirement for CD4 help in exogenous antigen presentation to CD8+ T cells: implications for vaccine development in immune-deficient hosts. J Exp Med 197:1405-1416.
10. Winau, F., S. Weber, S. Sad, J. de Diego, S. L. Hoops, B. Breiden, K. Sandhoff, V. Brinkmann, S. H. Kaufmann, and U. E. Schaible. 2006. Apoptotic vesicles crossprime CD8 T cells and protect against tuberculosis. Immunity 24:105-117.
Figures and legends
Figure 1. Subcutaneous injection of Histoplasma yeast cells induces CD8 and CD4 T cell activation. (A) Wild-type B6 mice were subcutaneous injected with live Histoplasma yeasts
(Live Hc), heat-killed Histoplasma (HKHc) or medium only control (RPMI) at day-14 and day0 (black arrows). At 5 days after second injection (red arrow), lymph node cells from draining lymph nodes (DLN) were isolated. The cells were stimulated with (Stimulated) or without (Unstimulated) HK Hc for 24 h and stained for surface CD4, CD8 and intracytoplasmic IFNγ. (B) The data were analyzed by flow cytometry FACS Canto. The numbers in the figure indicate the percentage of CD4 or CD8 T cells that produce IFNγ. One representative result was shown.
Figure 1. d -14 d 0 ICS Injection d 5 (A) CD8 IF N -g 0 0 0.63 3.04 0 0.42 Stimulated Unstimulated RPMI HKHc Live Hc (B) CD4 IF N -g 0.56 0 0 3.11 0.3 0.37 Stimulated Unstimulated RPMI HKHc Live Hc
Figure 2. Histoplasma injection induces CD8 and CD4 T cell activation during the course after secondary immunization. Mice were subcutaneous injected two times with live Hc, HKHc,
or RPMI as described in Figure 1. (A) The mice were sacrificed and the DLN cells were collected at day3, 5, 7, 10 and 14 after secondary injection (red arrow). The cells were stimulated and stained for IFNγ, CD4 and CD8. (B) The percentage of IFNγ-producing CD8 or CD4 cells in total CD8 or CD4 T cells. (C) Total number of IFNγ-producing CD8 or CD4 T cells in the DLN.
Figure 2. d 14 d -14 d 0 d 5 d 10 Injection ICS (A) (B) -0.5 0 0.5 1 1.5 2 2.5 3 3.5 4 3 5 7 10 14
day after secondary immunization
IF N -g + C D8 T c el ls /T ot al C D8 T c ells (% ) Live Hc HKHc RPMI -0.5 0 0.5 1 1.5 2 2.5 3 3.5 4 3 5 7 10 14
days after secondary immunization
IFN -g + C D 4 T c ells /T ot al C D 4 T c ells (% ) Live Hc HKHc RPMI
Figure 2. (C) -100 0 100 200 300 400 500 3 5 7 10 14
days after secondary immunization
IF N -g + CD 8 T c el l n um b er ( x1 0 3 ) Live Hc HKHc RPMI -100 0 100 200 300 400 500 600 3 5 7 10 14
days after secondary immunization
IF N -g + C D 4 T c el l numbe r (x10 3 ) Live Hc HKHc RPMI
Figure 3. Macrophages and neutrophils are the major populations of infiltrating cells. (A)
Subcutaneously injection of live Hc induces massive cell infiltration. Mice were injected with live Hc, HKHc or RPMI for 7 days. The cyrosection of injection sites were stained with H&E. (right column, magnification, ×40). The arrow indicated infiltrating cells. (B) Most of the infiltrating cells are F4/80+ and Gr-1+. Mice were injected subcutaneously with live Hc. At day 5 after primary infection, the cyrosection of injection sites were prepared for staining. PE-conjugated monoclonal antibodies against F4/80, Gr-1, CD3 and B220 were used. The slides were counterstained with Hoechst and analyzed by confocal microscope (magnification, ×630). The white line indicates the 40 μm scale bar. (C) Infiltrating cells are isolated and analyzed by FACS. At day 5 after primary injection of live Hc or HKHc, the subcutaneous injection sites were collected and treated with 5 mg/ml type 4 collagenase for 1 h at 37℃. The debris was removed by filter and the cells were collected by centrifugation. The cells were stained with anti-CD3, B220, CD11c, CD11b, F4/80, and Gr-1 antibodies to analyze the cell population.
Figure 3.
Figure 3.
(B)
Hoechst Antibody staining
Isotype control
F4/80
Gr-1
CD3
Figure 3. HKHc Live Hc CD11c F4/80 Gr-1 35.6 18.4 0.4 67.2 18.7 0.2 (C)
Figure 4. Subcutaneous injection of live Histoplasma induces macrophages and neutrophils apoptosis. Mice were injected with live Hc for 5 days, and paraffin sections of injection sites
were prepared and subjected to TUNEL staining (B, F) following the manufacturer’s instruction. The slides were also stained for surface Gr-1 (A-D) or F4/80 (E-H) by PE-conjugated mAbs and counterstained with Hoechst (A, E). The slides were analyzed by confocal microscope (magnification, ×630). The images of TUNEL and surface Gr-1 or F4/80 staining were merged (D, H). The white line indicates the 40 μm scale bar. The white arrows indicate the apoptotic Gr-1+ or F4/80+ cells.
Figure 4.
D B
A C
E F G H
Hoechst TUNEL Surface Merge
F4/80
Figure 5. Apoptotic HKHc-containing peritoneal macrophage induce CD8 T cell IFNγ production in a dose-dependent manner. Thioglycolate-elicited peritoneal macrophages were
harvested and allowed to ingest heat-killed Histoplasma yeasts. After wash, the macrophages were treated with LPS and ATP to induce cell apoptosis. Mice were injected with 1, 5, 10, or 20× 105 apoptotic macrophages twice with two week interval, and lymph node cells were harvested at day5 after secondary injection. Mice injected with live Hc, HKHc, or RPMI was used as control. The cells were stimulated and analyzed by flow cytometry as described in Figure 1.
Figure 5. 0 100 200 300 400 500 600 RPMI 1x10^5 5x10^5 1x10^6 2x10^6 Live Hc HKHc IF N -g + CD 8 T c el l n um be r ( x1 0 3 )
Figure 6. Antigen-engulfing peritoneal macrophage apoptosis is important to induce CD8 T
cell IFNγ production in vivo. Thioglycolate-elicited peritoneal macrophages (2×106) were
allowed to ingest heat-killed Histoplasma yeasts [(HKHc)] or left untreated. After wash, the macrophages were treated with LPS and ATP to induce apoptosis or not. Mice were injected with apoptotic [Apoptotic pMac (HKHc)] or non-apoptotic [pMac (HKHc)] HKHc-pulsed peritoneal macrophages, or apoptotic [Apoptotic pMac] or non-apoptotic [pMac] macrophages without antigen pulse. Apoptotic macrophages mixed with HKHc [Apoptotic pMac+HKHc] were injected into mice. IFNγ+ CD8 T cells were analyzed at day5 after secondary injection. *** p<0.005, n=3~4.
Figure 6. 0 0.5 1 1.5 2 2.5 HKHc Apoptotic pMac (HKHc) Apoptotic pMac Apoptotic pMac +HKHc pMac (HKHc) pMac IF N -g + C D 8 T ce lls/ T ota l CD8 T c ells ( % ) *** *** (A) 0 50 100 150 200 250 300 HKHc Apoptotic pMac (HKHc) Apoptotic pMac Apoptotic pMac +HKHc pMac (HKHc) pMac IF N -g + C D8 T c ell n um ber ( x1 0 3 ) *** *** (B)