FimK regulation on the expression of type 1 fimbriae
in Klebsiella pneumoniae CG43S3
Zhe-Chong Wang,
13 Ching-Jou Huang,
23 Ying-Jung Huang,
1Chien-Chen Wu
1and Hwei-Ling Peng
1,2Correspondence Hwei-Ling Peng [email protected]
Received 15 March 2013 Accepted 15 May 2013
1Department of Biological Science and Technology, National Chiao Tung University, Hsin Chu, Taiwan, Republic of China
2Institute of Molecular Medicine and Bioengineering, School of Biological Science and Technology, National Chiao Tung University, Hsin Chu, Taiwan, Republic of China
Klebsiella pneumoniae CG43, a heavy encapsulated liver abscess isolate, mainly expresses type 3 fimbriae. Type 1 fimbriae expression was only apparent in CG43S3DmrkA (the type 3 fimbriae-deficient strain). The expression of type 1 fimbriae in CG43S3DmrkA was reduced by deleting the fimK gene, but was unaffected by removing the 39 end of fimK encoding the C-terminal EIL domain (EILfimK). Quantitative RT-PCR and promoter activity analysis showed that the putative DNA-binding region at the N terminus, but not the C-terminal EIL domain, of FimK positively affects transcription of the type 1 fimbrial major subunit, fimA. An electrophoretic mobility shift assay demonstrated that the recombinant FimK could specifically bind to fimS, which is located upstream of fimA and contains a vegetative promoter for the fim operon, also reflecting possible transcriptional regulation. EILfimKwas shown to encode a functional phosphodiesterase (PDE) via enhancing motility in Escherichia coli JM109 and in vitro using PDE activity assays. Moreover, EILfimKexhibited higher PDE activity than FimK, implying that the N-terminal DNA-binding domain may negatively affect the PDE activity of FimK. FimA expression was detected in CG43S3 expressing EILfimKor AILfimK, suggesting that FimA expression is not directly influenced by the c-di-GMP level. In summary, FimK influences type 1 fimbriation by binding to fimS at the N-terminal domain, and thereafter, the altered protein structure may activate C-terminal PDE activity to reduce the intracellular c-di-GMP level.
INTRODUCTION
Klebsiella pneumoniae is an important nosocomial
patho-gen that causes suppurative lesions, septicaemia, and
urinary and respiratory tract infections in
immunocom-promised patients (Han, 1995; Schelenz et al., 2007). A
steady increase in the incidence of Klebsiella liver abscesses
(KLAs) in patients with diabetes, malignancy, renal disease
or pneumonia has been observed in Taiwan (Fung et al.,
2002). Reports of KLAs have also increased in Western and
other Asian countries (Pope et al., 2011). Although KLA
pathogenic mechanisms remain obscure, several virulence
traits, including the vast amount of K1 capsular
polysac-charide surrounding the bacteria surface (Fung et al.,
2002), magA (Chuang et al., 2006), iron acquisition loci on
pLVPK (Tang et al., 2010) and type 1 fimbriae (Struve
et al., 2008, 2009; Stahlhut et al., 2012), have been reported.
Fimbriae (also called pili) allow bacteria to attach to host
cells to establish infection. Type 1 fimbriae expressed by
most members of the family Enterobacteriaceae are
com-monly associated with urinary tract infections and bind to
the mannose-containing structure present on host cells or in
the extracellular matrix (Ishikawa, 1991; Jones et al., 1995).
The expression of type 1 fimbriae is phase-variable and is
mediated by an invertible 314 bp cis element, fimS, located
upstream of the type 1 fimbrial major subunit gene, fimA.
The fimS switching, which alternates Escherichia coli between
type 1 fimbriated and non-fimbriated states, is controlled by
site-specific recombinases FimB and FimE. In addition,
DNA-binding proteins IHF and Lrp can specifically bend
fimS DNA, enabling proper positioning of the inverted
repeat sequences to facilitate recombination (Schwan, 2011).
Besides fimB, fimE and the fimAICDFGH gene cluster, a
unique gene, fimK, was found immediately downstream of
the fimH gene in the K. pneumoniae genome. The fimK
gene had also been shown to be transcribed as part of the
3These authors contributed equally to this paper.
Abbreviations: bis-pNPP, bis(p-nitrophenol) phosphate; c-di-GMP, cyclic di-GMP; EMSA, electrophoretic mobility shift assay; KLA, Klebsiella liver abscess; PDE, phosphodiesterase.
Two supplementary figures are available with the online version of this paper.
single fimAICDFGH operon (Rosen et al., 2008). The FimK
protein of a K. pneumoniae urinary tract infection isolate
negatively affects the expression of type 1 fimbriae. This may
be caused by the involvement of cyclic di-GMP (c-di-GMP)
phosphodiesterase (PDE) activity (Rosen et al., 2008). This
possibility is supported by the multiple sequence alignment
data of Fig. S1, available in Microbiology online, which
reveals a conserved EAL domain at the C terminus of FimK.
In addition to the EAL domain, a DNA-binding domain has
been predicted in FimK (Struve et al., 2008). Amino acid
sequence analysis on the basis of Pfam classification also
showed a putative helix–turn–helix DNA-binding motif
from the HTH_23 family (http://pfam.sanger.ac.uk/) at the
N-terminal region of FimK.
Analysis of the genome sequence of K. pneumoniae CG43
(unpublished data), a liver abscess isolate that belongs to
the K2 serotype (Chang et al., 1996), showed that the type 1
fimbriae gene cluster is physically linked and divergently
transcribed to the type 3 fimbriae operon. This study
reports FimK-mediated regulation of type 1 fimbriae
expression at the transcriptional level in K. pneumoniae
CG43S3, which may be achieved by binding with the HTH
domain of FimK to the fimS DNA. The changed protein
Table 1. Bacterial strains and plasmids used in this study
Strain or plasmid Properties* Reference or source
Strain K. pneumoniae
CG43S3 Smr, CG43 derived strain Lai et al. (2003)
CG43S3DmrkA CG43S3 with deletion of mrkA gene This study
CG43S3DfimK CG43S3 with deletion of fimK gene This study
CG43S3DEILfimK CG43S3 with deletion of EILfimKcoding region This study
CG43S3DmrkADfimK CG43S3 with deletion of mrkA and fimK gene This study
CG43S3DmrkADGTGfimK CG43S3 with deletion of mrkA and HTHfimKcoding region This study
CG43S3DmrkADEILfimK CG43S3 with deletion of mrkA and EILfimKcoding region This study
CG43S3DmrkADlacZ CG43S3DmrkA with deletion of lacZ gene This study
CG43S3DmrkADfimKDlacZ CG43S3DmrkADfimK with deletion of lacZ gene This study
CG43S3DmrkADEILfimKDlacZ CG43S3DmrkADEILfimKwith deletion of lacZ gene This study
E. coli
JM109 Cloning host Laboratory stock
NovaBlue BL21(DE3) Recombinant protein overexpression host Laboratory stock
Plasmid
pETQ31 Kmr; expression vector Wu et al. (2010)
pETQ33 Kmr; expression vector Wu et al. (2010)
pKAS46 Kmrand Apr; suicide vector Skorupski & Taylor (1996)
pRK415 Tcr; broad-host-range vector Keen et al. (1988)
pBAD33 Cmr; expression vector Guzman et al. (1995)
pET30a-mrkJ Kmr; mrkJ coding region cloned into pET30a This study
pQE31-fimK Apr; fimK coding region cloned into pQE31 This study
pETQ31-EILfimK Kmr; fimK-EIL domain region cloned into pETQ31 This study
pETQ31-AILfimK Kmr; fimK-AIL domain region cloned into pETQ31 This study
pETQ33-fimB Kmr; fimB coding region cloned into pETQ33 This study
pETQ33-ydeH Kmr; ydeH coding region cloned into pETQ33 This study
pETQ33-EILfimK3 Kmr; fimK-EIL domain region cloned into pETQ3 This study
pETQ33-AILfimK Kmr; fimK-AIL domain region cloned into pETQ33 This study
pETQ33-mrkJ Kmr; mrkJ coding region cloned into pETQ33 This study
pRK415-fimK Tcr; fimK gene cloned into pRK415 This study
pRK415-fimKE245A Tcr; fimK gene cloned into pRK415 This study
pRK415-HTHfimK Tcr; fimK N-terminal HTH domain cloned into pRK415 This study
pRK415-EILfimK Tcr; EILfimKcoding region cloned into pRK415 This study
pBAD33-mrkJ Cmr; mrkJ coding region cloned into pBAD33 This study
pBAD33-EILfimK Cmr; fimK-EIL domain region cloned into pBAD33 This study
pBAD33-AILfimK Cmr; fimK-AIL domain region cloned into pBAD33 This study
pBAD33-ydeH Cmr; ydeH coding region cloned into pBAD33 This study
Table 2. Oligonucleotide primers used in this study
Purpose Primer name Sequence (5§A3§)
Gene deletions mrkA SL0141 CGAGCTCAGCGTGATGTCTATCCAG SL0142 CGCGGATCCCGAATCAATGAGCACACT SL0143 CGCGGATCCACAATAATAAAGCGGCAAT SL0158 TGCTCTAGAGACTGCCGACAATAAAGC fimK K1 TCGCTTCCCGCTGCAGGCC K2 CTTCGCGGCGTTCAGCATC K3 AGATCTTCGTATTCGCGGGTG K4 CATGATCTGCGCGTCGAGG
EILfimK WCC32 AGGCTACTCGACGAGGCCTTGC
WCC33 GGATCCTTAAAACGCCGTCAGTGC WCC34 GGATCCGGCGGATCGTTGAGG WCC35 GGTACCCGGATGAAGTGGATGTCG HTHfimK WCC127 ACCTCTAGACAGGCGGTGATTAACGTCACCTATAC WCC128 ATTAAGCTTCGAGCAGGGCGAGAGGATATAAT WCC133 ATTAAGCTTAACGCACTGACGGCGTTTGAA WCC130 ATTGAGCTCAAGATTATCCCTCTCTGCCCG Gene expression
fimK/fimKE245A WCC71 GATGACCGATTATATCCTCTCGCCC
WCC74 GAATTCAACGATCCGCCGGATCG
WCC81 ACCTCTAGAAGGAGATCAATGATGACCGATTATATCCT
WCC84 ATTAAGCTTAACGATCCGCCGGATCGC
HTHfimK WCC72 GAATTCTCATTCAAACGCCGTCAGTGCGTT
WCC82 ACCAAGCTTATTCCTGTTCAAACGCCGT
EILfimK/AILfimK WCC73 GATGGCCCCCCGCTTTCC
WCC83 ATTTCTAGAAGGAGGTGGAATGCATCCGCAT ydeH WCC75 GATGATCAAGAAGACAACGGAAATTGATG WCC76 GAATTCTTAAACTCGGTTAATCACATTTTGTTCGTC WCC79 ACCTCTAGAGTGAAAAAGGAGTGGCAATG WCC80 ACCAAGCTTTGAATGTTAAACGGAGCTTA mrkJ WCC77 GATGAACACTAAAATATTCGAAGACAACATTTTATCTC WCC78 GAATTCTTACATGGCAATATCATCGGCGAC WCC85 ACCTCTAGAAGGAGGGATAATGAACACTAAAATATT WCC86 ACCAAGCTTACATGGCAATATCATCGG fimB B1 GGATCCGAAAACCAAAAATATGC B2 CCTTGGATCAGCGACGATCGC Site-directed mutation
fimKE245A M1 CTATCGTCGACAGTCGCTCACAGCTACAGGGGGTGGCGATCCTG
M2 CAGGATCGCCACCCCCTGTAGCTGTGAGCGACTGTCGACGATAG EMSA WCC89 GCCATATTTCCGCAAAAAAAAAT WCC90 GATCCGTCGCCAACGCC WCC91 CCACATAATACCAAGTTGACACAAAATA WCC93 biotin-TTGGGGCCAAACTGTTTATATCAT WCC94 TTGGGGCCAAACTGTTTATATCAT WCC95 biotin-TTGGGGCCATTTTGACTCG WCC96 TTGGGGCCATTTTGACTCG WCC98 GCCGGTGCAGCAGAGATATG WCC99 TTTTAAATTCTGCCAAATTTGGTTTT WCC121 biotin-AAGGAAAAGCGATGGCGTTGGCG WCC122 AAGGAAAAGCGATGGCGTTGGCG WCC123 AATGTTTTGACATATTTTGCAACTCACTGCGC qRT-PCR
fimA fimA-F GAACGATGTCGAAATAACGAACCGG
structure may induce C-terminal PDE activity to decrease
the intracellular c-di-GMP level.
METHODS
Bacterial strains, plasmid, primer and growth conditions.Table 1 lists the bacterial strains and plasmids used in this study, and Table 2 lists the primers. Bacteria were grown in Luria–Bertani (LB) broth with shaking at 37uC, unless otherwise indicated. Antibiotics used included ampicillin (100 mg ml21), kanamycin (25 mg ml21), strep-tomycin (500 mg ml21), chloramphenicol (35 mg ml21) and tetracy-cline (12.5 mg ml21).
Immunoelectron microscopy. K. pneumoniae CG43S3 was grown overnight in LB broth with shaking, and 20 ml of the bacterial suspension (108c.f.u. ml21) was added to collodion-coated 300-mesh copper grids.
The bacteria coated on the grids were then incubated with a 1 : 50 dilution of the previously prepared anti-MrkA polyclonal antibody (Huang et al., 2009), followed by incubation with a 1 : 65 dilution of 10 nm gold particles conjugated with protein A (Sigma- Aldrich P9660). After negatively staining with 2 % (w/v) phosphotungstic acid, pH 7.2, the grids were examined under a JEOL JEM 2000EXII transmission electron microscope at an operating voltage of 100 kV.
FimA antisera preparation. The fimA coding sequence was amplified by PCR from K. pneumoniae CG43S3 and ligated into expression vector pET30a. Recombinant plasmid pET30a-fimA was then transformed into E. coli NovaBlueBL21(DE3), and overexpres-sion of the recombinant protein His6-FimA was induced with 0.5 mM
IPTG for 5 h at 37uC. The insoluble fraction was denatured using 6 M urea and then the protein purified using a nickel column (Novagen) saturated with 6 M urea. Finally, 3 mg of the purified His6-FimA was used to immunize rabbits for the anti-FimA antibody. Constructing the gene deletion mutants. Specific gene deletion was introduced into the chromosome of K. pneumoniae CG43S3 using an allelic-exchange strategy (Lai et al., 2003). Briefly, DNA fragments of 1 kb flanking both ends of mrkA, fimK, EILfimKand
HTHfimK DNA were amplified by PCR with primer sets SL0141/
SL0142 and SL0143/SL0158, K1/K2 and K3/K4, WCC32/WCC33 and WCC34/WCC35, and WCC127/CC128 and WC133/WCC130, respectively. The amplified DNA fragments were individually cloned into suicide vector pKAS46 (Skorupski & Taylor, 1996). The resulting plasmid was transformed into E. coli S17-1lpir and then mobilized by conjugating to the streptomycin-resistant strain, K. pneumoniae CG43S3 (Lai et al., 2003). Several kanamycin-resistant transconju-gants, with the plasmid integrated into the chromosome by homologous recombination, were selected from the M9 agar plates supplemented with kanamycin and propagated in 2 ml LB broth
overnight. A small aliquot of the culture was plated on LB agar containing 500 mg streptomycin ml21. Colonies susceptible to
kanamycin were isolated, and the specific gene deletions were verified by PCR analysis.
Quantitative reverse-transcription PCR (qRT-PCR).Total RNA was isolated from bacteria using a High Pure RNA isolation kit (Roche), and residual DNA was eliminated with RNase-free DNase I according to the manufacturer’s instructions. The cDNAs used for PCR were synthesized from 1.5 mg RNA using a random hexamer primer form RevertAid H Minus First-strand cDNA synthesis kit (Fermentas). PCR was performed using an ABI Prism 7000 Detection system according to manufacturer’s instructions, and products were detected using SYBR Green PCR Master Mix (Roche). The RNA samples were normalized to the 23S rRNA level. Analysis was performed in triplicate in a reaction volume of 25 ml containing 12.5 ml SYBR Green PCR Master Mix, 300 nM primer pair, 9.5 ml distilled H2O and 1 ml cDNA. Samples were heated for 10 min at
95uC and amplified for 40 cycles of 15 s at 95 uC and 60 s at 60 uC. Quantification was performed using the 2–DDCTmethod (Tsai et al., 2009).
Western blot analysis of FimA and MrkA expression.Aliquots of total cellular lysates were resolved by SDS-PAGE, and proteins were electrophoretically transferred onto a PVDF membrane (Millipore). After incubation with 5 % skimmed milk at room temperature for 1 h, the membrane was washed three times in PBS with Tween 20 (PBST). The membrane was then incubated with anti-GAPDH (GeneTex), anti-FimA or anti-MrkA antiserum at room temperature for 2 h. After three washes with 16 PBST, the PVDF membrane was incubated with a 1 : 5000 dilution of the secondary antibody, alkaline phosphatase-conjugated anti-rabbit immunoglobulin G, at room temperature for 1 h. Finally, the blot was rewashed, and the secondary antibodies bound on the PVDF membrane were detected using chromogenic reagents 5-bromo-4-chloro-3-indolyl phosphate and nitro blue tetrazolium.
Yeast-cell agglutination. Agglutination of yeast Saccharomyces cerevisiae AH109 was conducted as described by Blumer et al. (2005). Bacteria (109c.f.u. ml21) were suspended in PBS with or without mannose and then mixed with yeast (10 mg ml21) on a glass slide.
The degree of clumping was assessed by observation.
Biofilm formation assay.As described previously (Lin et al., 2006; Wu et al., 2010), bacteria diluted 1 : 100 in LB broth supplemented with appropriate antibiotics were inoculated into each well of a 96-well microtitre dish (Orange Scientific) and statically incubated at 37uC for 20 h. After removing planktonic cells, the wells were washed once with distilled water to remove unattached cells. Crystal violet (0.1 % w/v; Sigma-Aldrich) was used to stain the attached cells for 30 min. Unattached dye was removed by washing three times with Table 2. cont.
Purpose Primer name Sequence (5§A3§)
23S rRNA 23S-F AGCGACTAAGCGTACACGGTGG
23S-R GATGTTTCAGTTTCAGTTCCCCCGGTTC
Promoter activity assay
PfimA-lacZ pfimA3 TTGGATCCATTTTGACTCGTTG
pfimA4 GGGGCCAAACTGTTTAGATCTT
PfimB-lacZ pfimB4 TGTCGGCGGGATTCCTCATGG
pfimB5 CAAGATCTTGAGCATACCACAGC
PfimE-lacZ pfimE1 ACGGATGGCGTTGTATCGCG
fimE fimE fimA fimA Bsa BI BsaBI BI 328 bp 322 bp 204 bp 446 bp IRL IRR IRL IRR Phase OFF (a) (b) (c) Phase ON P1 P2 P1 P2 1000 750 500 250 bp Marker CG43S3 Phase OFF Uncut PCR products CG43S3 MrkA FimA GAPDH CG43S 3ΔmrkA CG43S3 0 1 2 3 4 5
Relative transcription ratio
fimA transcript CG43S3 ΔmrkA P <0.0001 * 200 nm
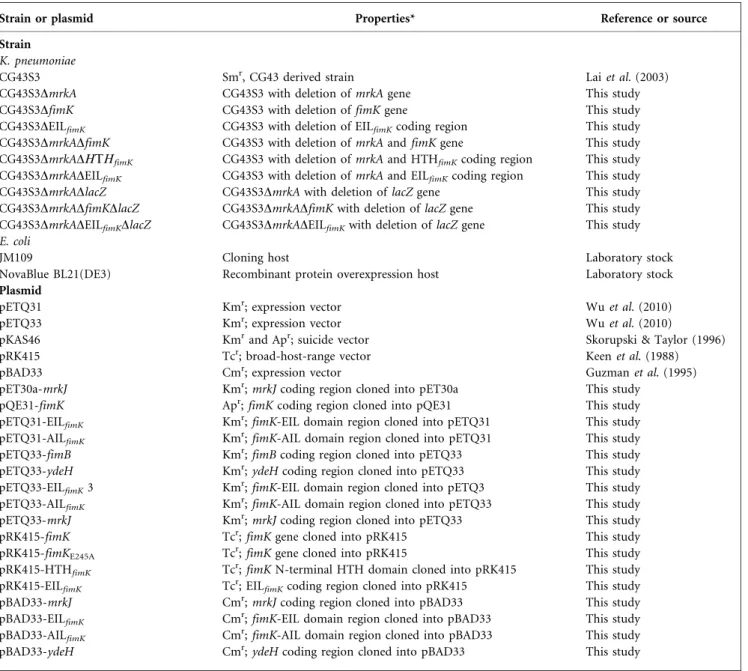
Fig. 1. Type 1 and type 3 fimbriae expression in K. pneumoniae CG43S3. (a) Transmission electron micrograph of K. pneumoniae CG43S3 labelled with anti-MrkA antibodies. (b) Diagrammatic representation of fimS promoter analysis (left). The primer pair P1 and P2 was used for amplifying the region containing fimS. The DNA pattern corresponding to incomplete digested amplicons or phase OFF of type 1 fimbriae is shown (right). (c) Analysis of FimA expression using qRT-PCR (left panel) and Western blot analysis (right panel). GAPDH was probed as a protein loading control. Values are means of three independent experiments. Error bars shown areSD, and asterisks denote statistically significant differences.
distilled water. The stained biomass was dissolved in 1 % (w/v) SDS, and the OD595was determined to assess biofilm-forming activity. Plasmid construction. The coding region of fimB, fimK, the N-terminal regions of the HTH domain (1–218 aa) or EIL domain (195–469 aa) of fimK, and mrkJ were PCR-amplified with primer pairs B1/B2, WCC71/WCC74, WCC71/WCC72, WCC73/WCC74 and WCC77/WCC78, respectively, from the CG43S3 genome. The amplified DNA was individually cloned into cloning vector yT&A (Yeastern Biotech). Site-directed mutation plasmid pyT-fimKE245A
was generated by substituting the glutamic acid at FimK position 245 with alanine using plasmid pyT-fimK as a template with overlapping primers M1 or M2 by PCR-based mutagenesis. Primer pair WCC73/ WCC74 was then used to amplify the AILfimKcoding region using
pyT-fimKE245Aas a template.
For complementation analysis or protein overexpression, the coding region from pyT-fimB, pyT-fimK, pyT-fimKE245A, pyT-HTHfimK,
pyT-EILfimK, pyT-AILfimKor pyT-mrkJ was subcloned into the
broad-host-range vector pRK415 (Keen et al., 1988) or expression vectors pETQ31, pETQ33 (Wu et al., 2010), pQE31 (Qiagen) or pET30a (Novagen). This resulted in low-copy-number complementation plasmids pRK415-fimK, pRK415-fimKE245A, pRK415-HTHfimK and
pRK415-EILfimKand overexpression clones pETQ33-fimB,
pETQ31-EILfimK, pETQ31-AILfimK, pQE31-fimK, pETQ33-EILfimK,
pETQ33-AILfimK, pETQ33-mrkJ and pET30a-mrkJ. The coding region of
gene ydeH (Jonas et al., 2008) was PCR-amplified with primer pair WCC75/WCC76 from the E. coli MG1655 genome and cloned into yT&A to generate plasmid pyT-ydeH. The ydeH coding DNA was then isolated from pyT-ydeH by restriction enzyme digestion and then cloned into pETQ33 to yield YdeH expression plasmid pETQ33-ydeH.
Promoter activity measurement.The putative promoter regions of fimA, fimB and fimE were PCR-amplified with primers pfimA3/ pfimA4, pfimB4/pfimB5 and pfimE1/pfimE2, respectively. The amplicons were then cloned into placZ15 (Lin et al., 2006) to generate PfimA-lacZ (locked on), PfimB-lacZ and PfimE-lacZ. The
promoter–reporter plasmids were individually mobilized into K. pneumoniae CG43S3 strains by conjugation from E. coli S17-1lpir. The bacteria were grown to the exponential phase in LB broth (OD600
of 0.7), and 100 ml of the culture was mixed with 900 ml of Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4,
50 mM b-mercaptoethanol), 17 ml of 0.1 % SDS and 35 ml chloro-form, followed by vigorous shaking. After incubation at 30uC for 5 min, 200 ml of a 4 mg ml21 concentration of ONPG
(Sigma-Aldrich) was added to the mixture to initiate the reaction. When the mixture became yellow, the reaction was stopped by adding 500 ml 1 M Na2CO3 and the absorbance at OD420 was recorded. The
promoter activity was expressed as Miller units. Each sample was assayed in triplicate and at least three independent experiments were conducted. The data were calculated from one representative experiment and are shown as the means andSDfrom three samples. Expression and purification of the recombinant proteins. Recombinant plasmids pETQ31-EILfimK and pETQ31-AILfimK were
transformed into E. coli JM109, and protein production was induced with 0.5 mM IPTG for 12 h at 22uC. The expression plasmid pET30a-mrkJ was introduced into E. coli NovaBlue BL21(DE3), and recombinant His6-MrkJ was induced with 0.5 mM IPTG for 5 h at
37uC. The recombinant protein His6-FimK was induced in E. coli
SG13009 [pREP4] with 0.01 mM IPTG for 24 h at 15 uC. All recombinant proteins were then purified from the soluble fraction of the cell lysate by affinity chromatography using His-Bind resin according to the QIAexpress expression system protocol (Qiagen). The purified proteins were dialysed against Tris-buffered saline (pH 7.4) containing 10 % (v/v) glycerol at 4uC overnight, followed
by condensation with PEG 20 000. Protein purity was determined by SDS-PAGE.
DNA electrophoretic mobility shift assay (EMSA).The variant truncated putative promoter of fimA was PCR-amplified using biotin-labelled primer pairs WCC93/WCC96, WCC93/WCC99, WCC93/ WCC98, WCC95/WCC91, WCC95/WCC90, WCC95/WCC89 and WCC121/WCC123 or non-labelled primer pairs WCC94/WCC96 and WCC122/WCC123. The DNA-binding reaction was performed in 20 ml binding buffer [100 nM MnCl2, 1 mM MgCl2, 0.5 mM DTT, CG43S3 [pETQ33]CG43S3 [pETQ33-fimB ] CG43S3 ΔfimK [pETQ33-fimB ] CG43S3 ΔEILfimK [pETQ33-fimB ] MrkA FimA GAPDH GAPDH FimA CG43S3 ΔmrkA CG43S3 ΔmrkA ΔfimK CG43S3 ΔmrkA ΔEILfimK CG43S3 ΔmrkA ΔHTH fimK FimA GAPDH CG43S3 ΔmrkA [pRK415] CG43S3 ΔmrkA ΔfimK [pRK4 15-fim K] CG43S3 ΔmrkA ΔfimK [pRK415-HTH fimK ] CG43S3 ΔmrkA ΔfimK [pRK415] (a) (b) (c)
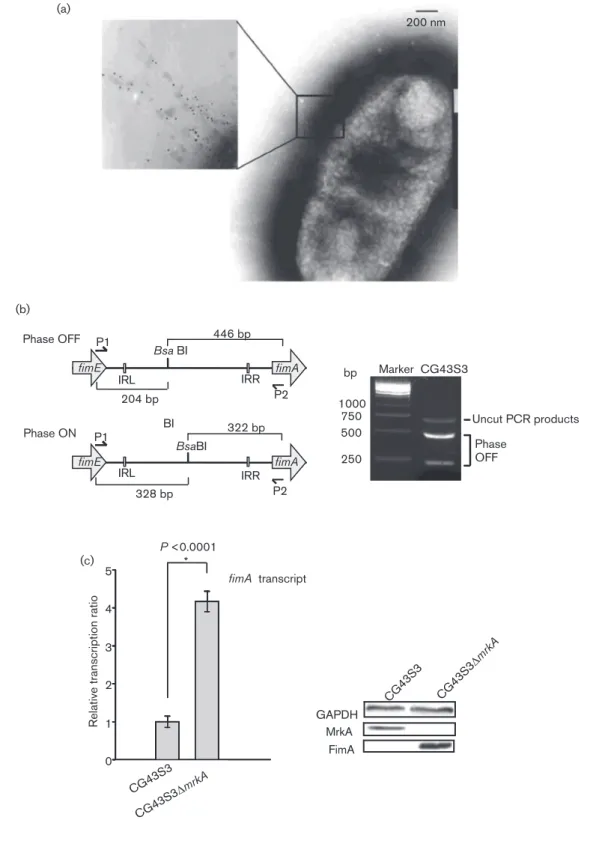
Fig. 2. Western blot analysis of fimK deletion effects on type 1 fimbriae expression. Expression of the type 1 fimbriae major pilin FimA was determined in (a) FimB recombinase overexpression bacteria CG43S3[pETQ33-fimB], and the derived strains CG43S3DfimK[pETQ33-fimB] and CG43S3DEILfimK
[pETQ33-fimB] and (b) CG43S3DmrkA, CG43S3DmrkADfimK, CG43S3 DmrkADEILfimK and CG43S3DmrkADHTHfimK. (c) Analysis of
FimK complementation by comparing FimA production of CG-43S3DmrkA[pRK415], CG43S3DmrkADfimK[pRK415], CG43S3 DmrkADfimK[pRK415-fimK] and CG43S3DmrkADfimK[pRK415-HTHfimK]. The bacteria carrying pETQ33 or pETQ33-fimB plasmid
were grown at 37 6C for 2 h, and subsequently grown in the presence of 0.5 mM IPTG for 4 h. The bacteria carrying pRK415 or the derivative plasmids were grown in LB broth supplemented with 0.01 mM IPTG for 20 h at 37 6C.
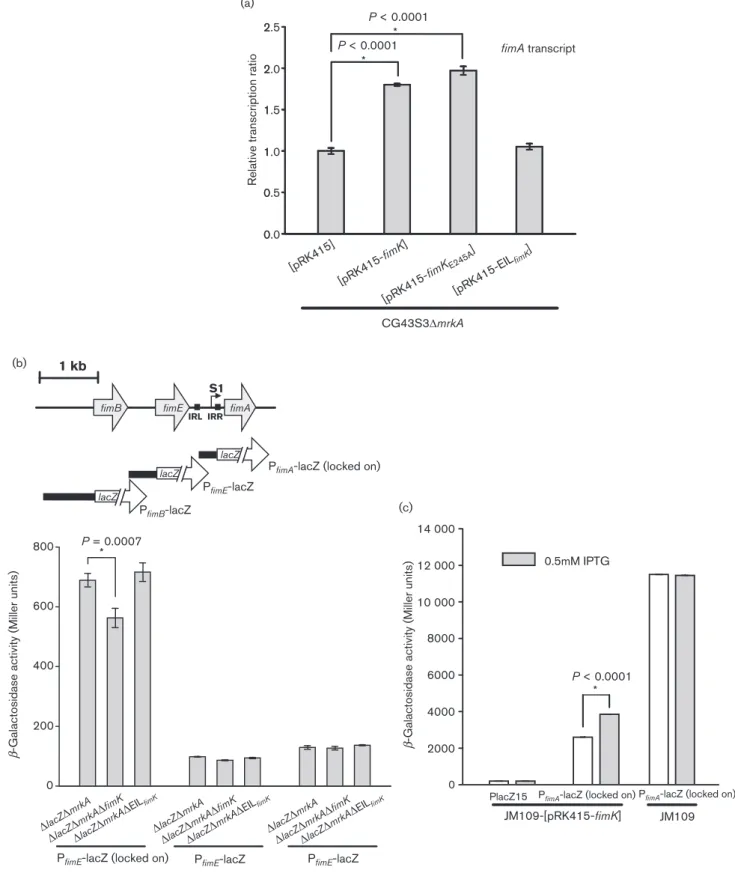
0 0 1 1 2 2 0.0 0.5 1.0 1.5 2.0 2.5 (a) (b) (c) fimA transcript [pRK415] [pRK415-fimK ] [pRK415-fimKE245A ] [pRK415-EIL fimK ]
Relative transcription ratio
CG43S3ΔmrkA P < 0.0001 * P < 0.0001 * 1 kb IRL lacZ lacZ
PfimA-lacZ (locked on)
PfimB-lacZ IRR
S1
fimB fimE fimA
PfimE-lacZ
PfimE-lacZ PfimE-lacZ
PfimE-lacZ (locked on)
lacZ
β
-Galactosidase activity (Miller units)
0 200 400 600 800 P = 0.0007 * ΔlacZΔ mrkA ΔlacZΔ mrkAΔ fimK ΔlacZΔ mrkAΔ EILfimK ΔlacZΔ mrkA ΔlacZΔ mrkAΔ fimK ΔlacZΔ mrkAΔ EILfimK ΔlacZΔ mrkA ΔlacZΔ mrkAΔ fimK ΔlacZΔ mrkAΔ EILfimK JM109-[pRK415-fimK] 0 2000 4000 6000 8000 10 000 12 000 14 000
PlacZ15 PfimA-lacZ (locked on)PfimA-lacZ (locked on) JM109 0.5mM IPTG
P < 0.0001 *
β
-Galactosidase activity (Miller units)
Fig. 3. FimK positively affects type 1 fimbriae transcript levels. (a) qRT-PCR analysis of the type 1 fimbriae (fimA) transcription. K. pneumoniae CG43S3DmrkA carrying pRK415 or the derivative plasmids was grown in LB broth supplemented with 0.01 mM IPTG at 37 6C for 20 h with agitation. RNA extraction and qRT-PCR detection were performed as described in Methods. qRT-PCR assays were conducted in triplicate. The error bars indicate the variations of three replicates from the mean. (b) The upper panel depicts the promoterless lacZ fusion to the putative promoter of fimA, fimB and fimE. IRR, IRL and S1 are,
50 mM KCl, 10 mM Tris/HCl (pH 7.5), 0.05 mg BSA ml21and 4 %
(v/v) glycerol]. In the reaction, sheared salmon sperm DNA was added at 0.05 mg ml21 to prevent non-specific DNA binding. After
transferring to a Biodyne B nylon membrane, biotin-labelled DNA was detected using a LightShift chemiluminescent EMSA kit (Pierce). PDE activity measurement. An in vitro PDE activity assay was performed as described by Johnson & Clegg (2010) with 300 nM of the purified recombinant proteins or a non-phosphodiesterase control, BSA (Sigma-Aldrich), in assay buffer [50 mM Tris/HCl, 1 mM MnCl2(pH 8.5)] supplemented with 5 mM bis(p-nitrophenol)
phosphate (bis-pNPP). Reaction mixtures were incubated at 37uC for 3 h, and the release of p-nitrophenol was quantified at 405 nm. Swimming activity analysis.The coding regions of EILfimK, AILfimK,
mrkJ and ydeH were amplified from pyT-fimK, pyT-fimKE245A,
pyT-mrkJ and pyT-ydeH, respectively, with primer pairs WCC83/WCC84, WCC85/WCC86 and WCC79/WCC80 (Table 2). The amplified DNA products were individually cloned into expression vector pBAD33 (Guzman et al., 1995), and the resulting plasmids were transformed into E. coli JM109. Swimming activity analysis for the recombinant bacteria was then conducted as described by Wood et al. (2006). Four microlitres of the bacteria grown overnight carrying pBAD33-EILfimK,
pBAD33-AILfimK, pBAD33-mrkJ or pBAD33-ydeH was spotted onto a
tryptone swimming plate (0.25 % Bacto agar, 0.25 % NaCl, 1 % tryptone, 35 mg chloramphenicol ml21and 0.1 % arabinose), and the plate was incubated for 12 h at 37uC.
Measuring c-di-GMP level in K. pneumoniae CG43S3. Expression plasmids pETQ33, pETQ33-EILFimK, pETQ33-AILFimK
or pETQ33-ydeH were individually transformed into K. pneumoniae CG43S3, and overexpression of the recombinant proteins was induced with 0.5 mM IPTG for 4 h at 37uC. To measure the c-di-GMP content, cellular extracts were prepared as described by Simm et al. (2004). The IPTG-induced bacteria were collected and treated with formaldehyde (0.19 % final concentration), and then pelleted by centrifugation. The pellet was suspended in de-ionized water and heated to 95uC for 10 min before nucleotides were extracted using 65 % ethanol. The lyophilized samples were resuspended in de-ionized water, and the suspension was used to detect c-di-GMP with the cyclic diguanylate ELISA kit (Wuhan EIAab Science).
RESULTS
Inverse expression of type 1 and 3 fimbriae in
K. pneumoniae CG43S3
Transmission electron microscopy detection with an
immunogold-labelled antibody against the major pilin,
MrkA, was used to demonstrate the synthesis of type 3
fimbriae on the surface of K. pneumonaie CG43S3 (Fig. 1a).
Expression of the fim operon could be assessed by the
restriction pattern of the fimS DNA amplified from the
bacterial culture. Fig. 1(b) shows some incomplete digested
amplicons and the BsaBI restriction fragments of
approxi-mately 450 and 200 bp, corresponding to the phase ‘OFF’
fimS in CG43S3. This restriction pattern reflects no
detectable expression of the type 1 fimbriae. The expression
of type 1 fimbriae pilin FimA was only evident in the mrkA
deletion mutant, as shown by qRT-PCR and Western blot
analysis (Fig. 1c). The results show that deleting the
predominant type 3 fimbriae pilin mrkA gene increased
type 1 fimbriae expression. The inverse expression pattern
between the two types of fimbriae was further observed by
introducing pETQ33-fimB, which overexpresses the FimB
recombinase, into K. pneumoniae CG43S3. Fig. 2(a) shows
that overexpression of the FimB recombinase turns on
expression of FimA but depletes MrkA production.
FimK plays a positive regulatory role in the
expression of type 1 fimbriae
As shown in Fig. 2(a), deleting fimK from
CG43S3[pETQ33-fimB] slightly decreased the expression of FimA but greatly
increased the expression of MrkA as judged by immunoblot
analysis targeting MrkA and FimA. However, deleting only
the DNA coding for the FimK EIL domain (195–469 aa) in
an in-frame fashion from CG43S3[pETQ33-fimB] had no
apparent effect on FimA production. To further confirm the
regulatory role of FimK on the expression of type 1 fimbriae,
the FimK-coding DNA and the DNA coding for the HTH
domain were individually deleted in an in-frame fashion
from CG43S3DmrkA. Compared with CG43S3DmrkA,
CG-43S3DmrkADfimK and CG43S3DmrkADHTH
fimKreduced
FimA production slightly while CG43S3DmrkADEIL
fimKhad
no apparent effect on FimA expression (Fig. 2b). The results
suggest that the FimK regulation of type 1 fimbriae is
dependent on the N-terminal DNA-binding domain. As
shown in Fig. 2(c), introducing plasmid pRK415-fimK or
pRK415-HTH
fimK, which expressed an intact FimK or the
FimK DNA-binding domain, into CG43S3DmrkADfimK
increased FimA expression levels. The FimK-dependent type
1 fimbriae activities were also observed using
mannose-sensitive yeast agglutination assay. As shown in Fig. S2(a, b),
deleting fimK from CG43S3[pETQ33-fimB] or CG43S3-DmrkA
had no dramatic change on the mannose-sensitive
agglutina-tion activity. However, the complementaagglutina-tion analysis clearly
revealed an FimK- or HTH domain-dependent
mannose-sensitive agglutination activity (Fig. S2c). These results suggest
that FimK positively affects the type 1 fimbriae expression
possibly through its N-terminal DNA-binding domain.
respectively, the inverted repeat right and left of fimS and the transcription start site. Lower panel: the promoter activity of fimA, fimB and fimE as, respectively, assessed by expression of the b-galactosidase reporter plasmid PfimA-lacZ (locked on), PfimB-lacZ and PfimE-lacZ, in K. pneumoniae CG43S3DlacZDmrkA and its isogenic derivatives DlacZDmrkADfimK and
DlacZDmrkADEILfimKwere determined. (c) Two-plasmid heterologous expression analysis in E. coli JM109 for the regulation
of FimK on the fimA promoter activity. Expression of fimA assessed via b-galactosidase activity of the strains JM109[PfimA
-lacZ][pRK415-fimK] and JM109[PlacZ15][pRK415-fimK], and also the control strain JM109[PfimA-lacZ] with or without IPTG
addition into the cultures. The bacteria were grown in LB broth at 37 6C and FimK expression was induced with 0.5 mM IPTG for 5 h. Data represent an average of three independent experiments.
The N-terminal domain of FimK positively
influences P
fimAactivity
To investigate whether the fimK gene affects type 1 fimbriae
biosynthesis at a transcriptional level, the fimA transcript
level was determined using qPCR. As shown in Fig. 3(a),
introducing either plasmid fimK or
pRK415-fimK
E245Ainto CG43S3DmrkA increased the fimA transcripts
to approximately twice the value of CG43S3DmrkA[pRK415]
or CG43S3DmrkA[pRK415-EIL
fimK]. This suggests that the
fimA transcript-level change might not be directly influenced
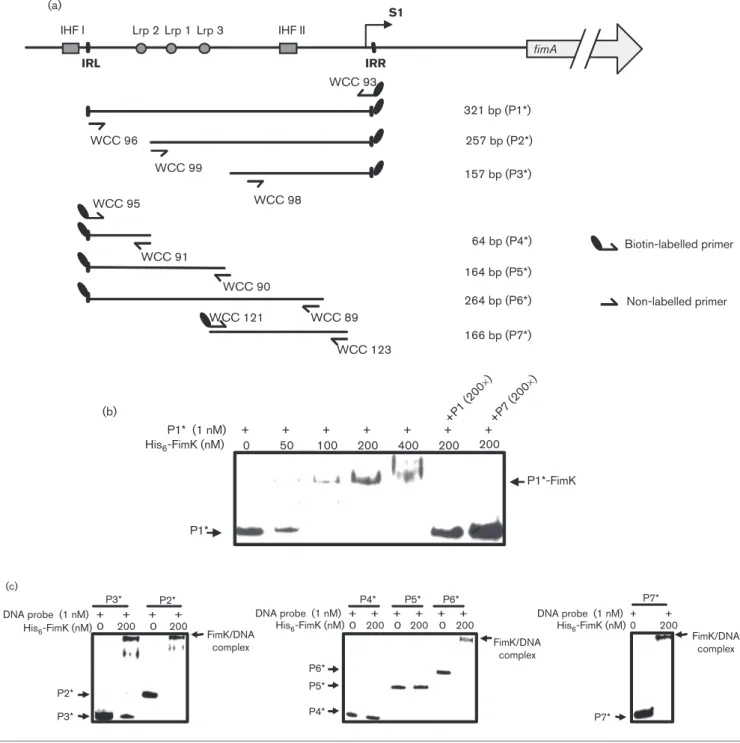
S1 IRR IRL WCC 91 WCC 90 WCC 89 WCC 98 WCC 99 WCC 93 WCC 96 WCC 95 257 bp (P2*) 321 bp (P1*) 157 bp (P3*) 64 bp (P4*) 164 bp (P5*) 264 bp (P6*) Biotin-labelled primer Non-labelled primer IHF I (a) (b) (c) IHF II Lrp 2 Lrp 1 Lrp 3 fimA WCC 121 WCC 123 166 bp (P7*) + + + + + + 200 400 200 100 50 0 200 + P1* (1 nM) His6-FimK (nM) P1*-FimK +P1 (200 ×) +P7 (200 ×) P1* FimK/DNA complex FimK/DNA complex FimK/DNA complex DNA probe (1 nM) His6-FimK (nM) DNA probe (1 nM) His6-FimK (nM) DNA probe (1 nM) His6-FimK (nM) P2* P3* + + + + 0 P2* P3* P4* P5* P5* P4* P6* P6* 0 200 P7* + + P7* 200 0 200 + + + + + + 0 200 0 200 0 200
Fig. 4. EMSA of the interaction between His6-FimK and fimS. (a) Schematic diagram of the DNA probes for the analysis. The
relative positions of the primer sets used in PCR amplification of the DNA probes are indicated. Names and sizes of the biotin-labelled DNA probes are shown on the right. S1, the transcription start site; IHF I and IHF II, the binding sites for integration host factor IHF; Lrp 1, Lrp 2 and Lrp 3, the leucine-responsive regulatory protein (Lrp) binding sites. (b) Interaction of the His6-FimK
and the putative promoter DNA. The biotin-labelled PfimA(P1*) was incubated with increasing amount of recombinant FimK
protein. Binding specificity was investigated by adding 200-fold unlabelled specific competitor DNA fragments (P1 or P7). (c) The biotin-labelled DNA probes P2*, P3*, P4*, P5*, P6* and P7* were applied for the specific binding region.
by the C-terminal EIL domain. To investigate whether the
N-terminal DNA-binding domain of FimK enhanced type 1
fimbriae expression by regulating fimB or fimE promoter
activity or directly affected fimA promoter activity, a LacZ
reporter assay was performed. Fig. 1(b) shows that the fimS is
mostly OFF-phased in CG43S3, and a locked ON fimA
promoter was therefore used. These promoter activities
in CG43S3DfimK appeared to be at similar levels to those
in CG43S3 (data not shown). We reason that fimK is
transcribed as part of the fimAICDFGH operon (Rosen
et al., 2008) and expression of type 1 fimbriae in CG43S3 is
not detected in the culture condition. Therefore, the
promoter activities of fimA, fimB and fimE were measured
in CG43S3DmrkA to examine the fimK gene deletion effect.
As shown in Fig. 3(b), the promoter activity of fimA, but not
of fimB or fimE, was reduced by deleting fimK. This suggests
that FimK affects type 1 fimbriae expression by directly
influencing the fimA promoter. The negative effect was not
observed for P
fimAin CG43S3DmrkADEIL
fimk, implying that
the N-terminal region of FimK plays a regulatory role. As
shown in Fig. 3(c), P
fimAactivity in E. coli JM109 was not
changed with IPTG addition, while heterologous expression
of FimK by IPTG induction increased the expression levels of
fimA, further supporting a direct regulation of FimK on
fimA.
Recombinant FimK exhibits a specific
DNA-binding activity with P
fimAThe DNA fragments encompassing the full-length fimS (P1)
and the truncated forms P2*, P3*, P4*, P5*, P6* and P7* as
depicted in Fig. 4(a) were isolated and biotin-labelled for the
analysis. As shown in Fig. 4(b), formation of the P1*/FimK
complex could be observed as the amount of His
6-FimK
increased, and the binding specificity was demonstrated as
the complex diminished in the presence of excess
non-labelled P1 or P7 acting as specific competitor DNA. The
sheared salmon sperm DNA was added as non-specific
competitor reagent. Fig. 4(c) shows that the purified
recombinant His
6-FimK protein was able to bind to the
DNA probes P2, P3, P6 and P7, but not to P4 or P5. The
results support the conclusion that the recombinant FimK
protein could specifically interact with fimS DNA and also
suggest that the FimK-binding site is located within P7.
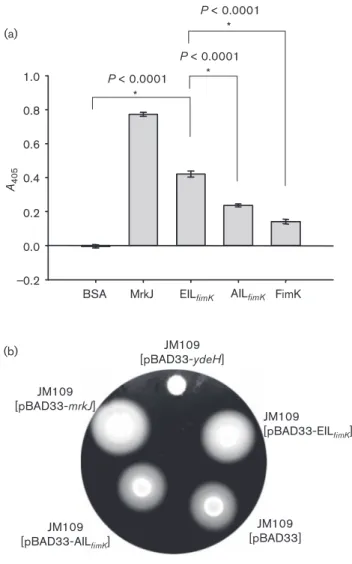
The recombinant FimK protein exerts PDE activity
Some PDE domain proteins do not exhibit PDE-specific
enzyme activity (Ro¨mling, 2009), so we studied whether
FimK possesses PDE activity. As shown in Fig. 5(a), the
purified recombinant FimK exhibited considerably less
bis-PNPP catalytic activity than the recombinant MrkJ, for
which PDE activity has been reported (Johnson et al.,
2011). Recombinant clones expressing EIL
fimKand AIL
fimKcontaining the C-terminal EIL domain of FimK and the
domain with critical E245 residue replaced with alanine,
respectively, were also generated and the corresponding
overexpressed proteins were purified. The recombinant
EIL
fimKexhibited a higher level of PDE activity than the
recombinant FimK and AIL
fimK. The FimK protein
exhibited the lowest PDE activity of all the tested proteins,
except the negative control BSA.
The second-messenger c-di-GMP levels determine whether
E. coli and many other bacteria are in a motile or sessile
state (Jonas et al., 2009). As assessed using swimming
activity analysis shown in Fig. 5(b), the heterologous
expression in E. coli JM109 of the EIL domain of FimK or
the intact MrkJ increased the swimming zone compared
with that of JM109[pBAD33] or JM109[pBAD33-ydeH],
_0.2 0.0 0.2 0.4 0.6 0.8 1.0 (a) (b)
BSA MrkJ EILfimK AILfimK FimK A405 P < 0.0001 * P < 0.0001 * P < 0.0001 * JM109 [pBAD33-EILfimK] JM109 [pBAD33-AILfimK] JM109 [pBAD33-mrkJ] JM109 [pBAD33-ydeH] JM109 [pBAD33]
Fig. 5. The recombinant FimK protein exerts PDE activity. (a) The recombinant MrkJ, the EIL domain and AIL domain of FimK, and FimK were analysed for PDE activity. BSA was used as a negative control for activity determination. Approximately 300 nM of each of the purified proteins was incubated with the substrate bis-pNPP at 37 6C for 3 h. Release of p-nitrophenol was determined at 405 nm. Results shown are representative of three independent experiments. Error bars represent SD. (b) Motility assay for the recombinant E. coli JM109[pBAD33], JM109[pBAD33-mrkJ], JM109[pBAD33-EILfimK], JM109[pBAD33-AILfimK] and JM109
which expresses diguanylate cyclase activity (Jonas et al.,
2008). The result supports that the FimK EIL domain
exerts PDE activity, which is able to lower the intracellular
c-di-GMP content, thereby increasing motility of the
recombinant E. coli.
FimA production is not directly dependent on
c-di-GMP levels
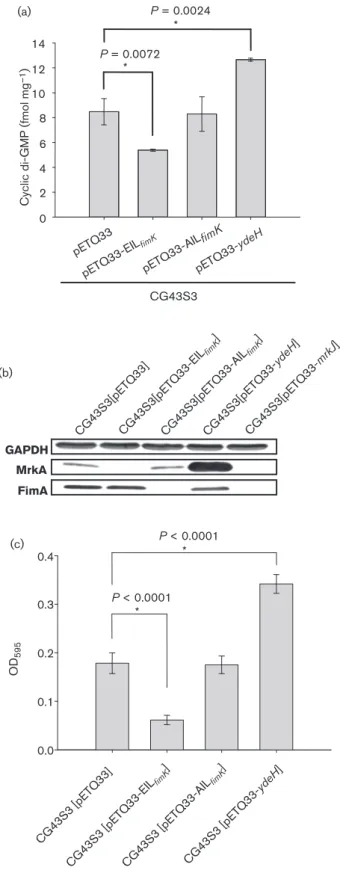
To further confirm EIL
fimKPDE activity, the c-di-GMP levels
in K. pneumoniae CG43S3 transformed with pETQ33,
pETQ33-ydeH, pETQ33-EIL
fimKand pETQ33-AIL
fimKwere
measured. As shown in Fig. 6(a), the c-di-GMP concentration
was approximately 12.7 fmol mg
21in
CG43S3[pETQ33-ydeH], which was higher than that in CG43S3[pETQ33] or
CG43S3[pETQ33-AIL
fimK]. The CG43S3[pETQ33-EIL
fimK]
contained the lowest level at 5.3 fmol mg
21, also confirming
c-di-GMP PDE activity of EIL
fimK.
Comparative analysis of the bacteria which carry different
c-di-GMP levels was conducted to determine if the
second-messenger levels regulate the expression of type 1 fimbriae.
As shown in Fig. 6(b), FimA production was observed
in CG43S3[pETQ33-EIL
fimK], CG43S3[pETQ33-AIL
fimK]
and CG43S3[pETQ33-mrkJ] but not in CG43S3[pETQ33]
or CG43S3[pETQ33-ydeH], reflecting MrkA expression.
Approximately the same level of c-di-GMP was detected in
CG43S3[pETQ33-AIL
fimK] and CG43S3[pETQ33],
indic-ating that FimA production is not directly affected by
c-di-GMP level. Despite this, the c-di-GMP level change
altered MrkA production and biofilm formation (Fig. 6c).
DISCUSSION
Research shows that cross-talk regulation may occur
between different fimbriae (Snyder et al., 2005; Sjo¨stro¨m
et al., 2009). In E. coli, during coordinate regulation for
type 1 and type P fimbriae, PapB, a regulator for type P
fimbriae expression, inhibits FimB-promoted
recombina-tion, thereby affecting the expression of type 1 fimbriae
(Xia et al., 2000). Analysing K. pneumoniae CG43 genome
sequences revealed at least ten fimbriae-coding gene
clusters (unpublished data). It is possible that the
expression of individual adhesins must be cooperatively
regulated in the bacteria. Fimbriated and non-fimbriated
planktonic cells display different outer-membrane protein
patterns (Otto et al., 2001). Decreased expression of type 1
fimbriae was reported for the E. coli K1 ompA deletion
mutant (Teng et al., 2006). Misfolding of the P fimbriae
subunit triggered the 2CS Cpx and s
Eregulatory pathways
(Jones et al., 1997). These studies suggest that deleting
mrkA may change the outer-membrane protein pattern or
trigger an envelope stress system, leading to the expression
of type 1 fimbriae in K. pneumoniae CG43S3.
As shown in Fig. 2(b), deleting fimK from the mrkA
deletion mutant reduced FimA production. These
reduc-tion effects were not observed in CG43S3DmrkADEIL
fimK,
suggesting that FimK regulates fim expression through
the N-terminal region. Complementation analysis
con-firms that the HTH
fimKdomain, but not the C-terminal
EIL domain, positively regulates type 1 fimbriae
expres-sion via increasing fimA transcription (Figs 2c and 3a).
Our LacZ reporter assay and EMSA data indicated that
FimK may bind to an fimS region upstream of the fimA
transcriptional start site and then activate fimA promoter
activity. In E. coli, IHF and Lrp could specifically bind on
and bent fimS DNA, enabling proper positioning of the
inverted repeat sequences to facilitate recombination
(Schwan, 2011). EMSA data indicate that FimK binds
to P7* DNA. Regional sequence analysis identified a
putative
IHF
binding
sequence,
59-TNYAANNNR-TTGAT-39, where Y is pyrimidine and R is purine
(Eisenstein et al., 1987). In Salmonella enterica serovar
Typhimurium, the fimA promoter is activated when FimZ
forms a complex with FimY protein but repressed by the
FimZ–FimW complex (McFarland et al., 2008). Hence,
FimK may regulate fim expression by cooperating with
IHF or Lrp or an unidentified protein such as FimZ in S.
Typhimurium. Nevertheless, FimK-mediated regulation
may be also achieved by changing the structure of
on-phase DNA, thereby improving transcription, or by
facilitating fimS switch from off to on through altering
the interaction between DNA and FimB or FimE
recombinase.
As shown in Fig. S1, the FimK EIL domain includes the
critical residues involved in Mg
2+substrate binding and
Loop 6 required for signal transduction (Rao et al., 2008,
2009; Ro¨mling, 2009). This suggests PDE activity for
FimK. Several studies have shown that PDE activity was
inactivated when critical residue E from the EAL domain
was replaced with alanine (Kuchma et al., 2007; Bassis &
Visick, 2010). However, recombinant proteins EIL
fimKand
AIL
fimKexpressed PDE activity, and only AIL
fimKexhibited
lower enzyme activity levels. When responding to different
internal and external signals, the sensory modules, such as
PAS, GAF, HAMP, REC and HTH domains, commonly
present with the GGDEF, EAL and HD-GYP domains,
activate diguanylate cyclase or PDE activity (Ho et al.,
2000; Galperin et al., 2001; Christen et al., 2005; Tamayo
et al., 2007; Cruz et al., 2012). FimK exerted higher PDE
activity when the N-terminal DNA-binding domain was
removed, suggesting that PDE activity may be activated
after the N-terminal domain is stimulated by a specific
signal.
This study provides novel insight into the function and
mechanism of FimK regulating type 1 fimbriation in K.
pneumoniae. The data demonstrate that FimK positively
affects the expression of type 1 fimbriae at the
transcrip-tional level by modulating P
fimApromoter activity after
binding to fimS. When interaction between the N-terminal
domain and DNA occurs, FimK PDE activity may be
activated to reduce intracellular c-di-GMP levels, thus
negatively affecting the expression of other surface
structures, such as type 3 fimbriae (Wilksch et al., 2011).
ACKNOWLEDGEMENTS
The work was supported by grants from the National Science Council (NSC97-2320-B-009-001-MY3 and NSC100-2320-B-009-003-MY3) and ‘Aim for the Top University Plan’ of the National Chiao Tung University and Ministry of Education (100W962), Taiwan, ROC.
REFERENCES
Bassis, C. M. & Visick, K. L. (2010).The cyclic-di-GMP phospho-diesterase BinA negatively regulates cellulose-containing biofilms in Vibrio fischeri. J Bacteriol 192, 1269–1278.
Blumer, C., Kleefeld, A., Lehnen, D., Heintz, M., Dobrindt, U., Nagy, G., Michaelis, K., Emo¨dy, L., Polen, T. & other authors (2005). Regulation of type 1 fimbriae synthesis and biofilm formation by the transcriptional regulator LrhA of Escherichia coli. Microbiology 151, 3287–3298.
Chang, H. Y., Lee, J. H., Deng, W. L., Fu, T. F. & Peng, H. L. (1996). Virulence and outer membrane properties of a galU mutant of Klebsiella pneumoniae CG43. Microb Pathog 20, 255–261.
Christen, M., Christen, B., Folcher, M., Schauerte, A. & Jenal, U. (2005).Identification and characterization of a cyclic di-GMP-specific phosphodiesterase and its allosteric control by GTP. J Biol Chem 280, 30829–30837.
Chuang, Y. P., Fang, C. T., Lai, S. Y., Chang, S. C. & Wang, J. T. (2006). Genetic determinants of capsular serotype K1 of Klebsiella pneumo-niae causing primary pyogenic liver abscess. J Infect Dis 193, 645– 654.
Cruz, D. P., Huertas, M. G., Lozano, M., Za´rate, L. & Zambrano, M. M. (2012). Comparative analysis of diguanylate cyclase and phospho-diesterase genes in Klebsiella pneumoniae. BMC Microbiol 12, 139. Eisenstein, B. I., Sweet, D. S., Vaughn, V. & Friedman, D. I. (1987). Integration host factor is required for the DNA inversion that controls phase variation in Escherichia coli. Proc Natl Acad Sci U S A 84, 6506–6510.
Fung, C. P., Chang, F. Y., Lee, S. C., Hu, B. S., Kuo, B. I. T., Liu, C. Y., Ho, M. & Siu, L. K. (2002). A global emerging disease of Klebsiella pneumoniae liver abscess: is serotype K1 an important factor for complicated endophthalmitis? Gut 50, 420–424.
Galperin, M. Y., Nikolskaya, A. N. & Koonin, E. V. (2001). Novel domains of the prokaryotic two-component signal transduction systems. FEMS Microbiol Lett 203, 11–21.
Guzman, L. M., Belin, D., Carson, M. J. & Beckwith, J. (1995).Tight regulation, modulation, and high-level expression by vectors contain-ing the arabinose PBAD promoter. J Bacteriol 177, 4121–4130.
Cyclic di-G MP (fmol mg –1 ) 0 2 4 6 8 10 12 14 (a) (b) (c) pETQ33 pETQ 33-E ILfimK pETQ33-AIL fimK pETQ33-ydeH P = 0.0072 * P = 0.0024 * CG43S3 MrkA FimA GAPDH CG43S3[pETQ33]CG43S3[pETQ33-EIL fimK ] CG43S3[pETQ33-AIL fimK ] CG43S3[pETQ33-ydeH ] CG43S3[pETQ33-mrkJ ] OD 595 0.0 0.1 0.2 0.3 0.4 CG43S3 [pETQ33] CG43S3 [pETQ33-EIL fimK ] CG43S3 [pETQ33-AIL fimK ] CG43S3 [pETQ33-ydeH ] P < 0.0001 * P < 0.0001 *
Fig. 6. c-di-GMP effects on the expression of type 1 fimbriae. (a) Quantification of c-di-GMP using ELISA according to the manual (Wuhan EIAab Science). Measurement of c-di-GMP was per-formed in three independent experiments. Error bars shown areSD, and asterisks indicate statistically significant differences. (b) Western blot analysis of FimA and MrkA production in K.
pneumoniae CG43S3[pETQ33], CG43S3[pETQ33-EILfimK],
CG-43S3[pETQ33-AILfimK], CG43S3[pETQ33-ydeH] and CG43S3
[pETQ33-mrkJ]. Bacteria carrying pETQ33 and other derivative plasmids were grown to mid-exponential phase and then supplemented with 0.5 mM IPTG, and the culture continued for 4 h at 37 6C. Thereafter, total proteins were isolated and resolved by SDS-PAGE. The proteins were transferred to PVDF membrane and hybridized with anti-FimA, anti-MrkA or anti-GAPDH. (c) Biofilm formation activity. Bacteria were inoculated into each well of a 96-well microtitre dish and statically incubated at 37 6C for 20 h, and then the biofilm-forming activity measurement was performed as described in Methods.
Han, S. H. B. (1995). Review of hepatic abscess from Klebsiella pneumoniae. An association with diabetes mellitus and septic endophthalmitis. West J Med 162, 220–224.
Ho, Y. S. J., Burden, L. M. & Hurley, J. H. (2000).Structure of the GAF domain, a ubiquitous signaling motif and a new class of cyclic GMP receptor. EMBO J 19, 5288–5299.
Huang, Y. J., Liao, H. W., Wu, C. C. & Peng, H. L. (2009).MrkF is a component of type 3 fimbriae in Klebsiella pneumoniae. Res Microbiol 160, 71–79.
Ishikawa, S. (1991).[The role of fimbriae of Escherichia coli in urinary tract infections]. Hinyokika Kiyo 37, 953–956 [in Japanese].
Johnson, J. G. & Clegg, S. (2010).Role of MrkJ, a phosphodiesterase, in type 3 fimbrial expression and biofilm formation in Klebsiella pneumoniae. J Bacteriol 192, 3944–3950.
Johnson, J. G., Murphy, C. N., Sippy, J., Johnson, T. J. & Clegg, S. (2011).Type 3 fimbriae and biofilm formation are regulated by the transcriptional regulators MrkHI in Klebsiella pneumoniae. J Bacteriol 193, 3453–3460.
Jonas, K., Edwards, A. N., Simm, R., Romeo, T., Ro¨mling, U. & Melefors, O. (2008).The RNA binding protein CsrA controls cyclic di-GMP metabolism by directly regulating the expression of GGDEF proteins. Mol Microbiol 70, 236–257.
Jonas, K., Melefors, O. & Ro¨mling, U. (2009). Regulation of c-di-GMP metabolism in biofilms. Future Microbiol 4, 341–358. Jones, C. H., Pinkner, J. S., Roth, R., Heuser, J., Nicholes, A. V., Abraham, S. N. & Hultgren, S. J. (1995).FimH adhesin of type 1 pili is assembled into a fibrillar tip structure in the Enterobacteriaceae. Proc Natl Acad Sci U S A 92, 2081–2085.
Jones, C. H., Danese, P. N., Pinkner, J. S., Silhavy, T. J. & Hultgren, S. J. (1997).The chaperone-assisted membrane release and folding pathway is sensed by two signal transduction systems. EMBO J 16, 6394–6406.
Keen, N. T., Tamaki, S., Kobayashi, D. & Trollinger, D. (1988). Improved broad-host-range plasmids for DNA cloning in gram-negative bacteria. Gene 70, 191–197.
Kuchma, S. L., Brothers, K. M., Merritt, J. H., Liberati, N. T., Ausubel, F. M. & O’Toole, G. A. (2007). BifA, a cyclic-di-GMP phosphodiesterase, inversely regulates biofilm formation and swarming motility by Pseudomonas aeruginosa PA14. J Bacteriol 189, 8165–8178.
Lai, Y. C., Peng, H. L. & Chang, H. Y. (2003).RmpA2, an activator of capsule biosynthesis in Klebsiella pneumoniae CG43, regulates K2 cps gene expression at the transcriptional level. J Bacteriol 185, 788– 800.
Lin, C. T., Huang, Y. J., Chu, P. H., Hsu, J. L., Huang, C. H. & Peng, H. L. (2006). Identification of an HptB-mediated multi-step phosphorelay in Pseudomonas aeruginosa PAO1. Res Microbiol 157, 169–175.
McFarland, K. A., Lucchini, S., Hinton, J. C. D. & Dorman, C. J. (2008). The leucine-responsive regulatory protein, Lrp, activates transcription of the fim operon in Salmonella enterica serovar typhimurium via the fimZ regulatory gene. J Bacteriol 190, 602–612.
Otto, K., Norbeck, J., Larsson, T., Karlsson, K. A. & Hermansson, M. (2001). Adhesion of type 1-fimbriated Escherichia coli to abiotic surfaces leads to altered composition of outer membrane proteins. J Bacteriol 183, 2445–2453.
Pope, J. V., Teich, D. L., Clardy, P. & McGillicuddy, D. C. (2011). Klebsiella pneumoniae liver abscess: an emerging problem in North America. J Emerg Med 41, e103–e105.
Rao, F., Yang, Y., Qi, Y. N. & Liang, Z. X. (2008).Catalytic mechanism of cyclic di-GMP-specific phosphodiesterase: a study of the EAL
domain-containing RocR from Pseudomonas aeruginosa. J Bacteriol 190, 3622–3631.
Rao, F., Qi, Y. N., Chong, H. S., Kotaka, M., Li, B., Li, J. M., Lescar, J., Tang, K. & Liang, Z. X. (2009).The functional role of a conserved loop in EAL domain-based cyclic di-GMP-specific phosphodiesterase. J Bacteriol 191, 4722–4731.
Ro¨mling, U. (2009).Rationalizing the evolution of EAL domain-based cyclic di-GMP-specific phosphodiesterases. J Bacteriol 191, 4697– 4700.
Rosen, D. A., Pinkner, J. S., Jones, J. M., Walker, J. N., Clegg, S. & Hultgren, S. J. (2008). Utilization of an intracellular bacterial community pathway in Klebsiella pneumoniae urinary tract infection and the effects of FimK on type 1 pilus expression. Infect Immun 76, 3337–3345.
Schelenz, S., Bramham, K. & Goldsmith, D. (2007).Septic arthritis due to extended spectrum beta lactamase producing Klebsiella pneumoniae. Joint Bone Spine 74, 275–278.
Schwan, W. R. (2011).Regulation of fim genes in uropathogenic Escherichia coli. World J Clin Infect Dis 1, 17–25.
Simm, R., Morr, M., Kader, A., Nimtz, M. & Ro¨mling, U. (2004). GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol Microbiol 53, 1123– 1134.
Sjo¨stro¨m, A. E., Balsalobre, C., Emo¨dy, L., Westerlund-Wikstro¨m, B., Hacker, J. & Uhlin, B. E. (2009).The SfaXII protein from newborn meningitis E. coli is involved in regulation of motility and type 1 fimbriae expression. Microb Pathog 46, 243–252.
Skorupski, K. & Taylor, R. K. (1996).Positive selection vectors for allelic exchange. Gene 169, 47–52.
Snyder, J. A., Haugen, B. J., Lockatell, C. V., Maroncle, N., Hagan, E. C., Johnson, D. E., Welch, R. A. & Mobley, H. L. T. (2005). Coordinate expression of fimbriae in uropathogenic Escherichia coli. Infect Immun 73, 7588–7596.
Stahlhut, S. G., Struve, C., Krogfelt, K. A. & Reisner, A. (2012). Biofilm formation of Klebsiella pneumoniae on urethral catheters requires either type 1 or type 3 fimbriae. FEMS Immunol Med Microbiol 65, 350–359.
Struve, C., Bojer, M. & Krogfelt, K. A. (2008).Characterization of Klebsiella pneumoniae type 1 fimbriae by detection of phase variation during colonization and infection and impact on virulence. Infect Immun 76, 4055–4065.
Struve, C., Bojer, M. & Krogfelt, K. A. (2009). Identification of a conserved chromosomal region encoding Klebsiella pneumoniae type 1 and type 3 fimbriae and assessment of the role of fimbriae in pathogenicity. Infect Immun 77, 5016–5024.
Tamayo, R., Pratt, J. T. & Camilli, A. (2007).Roles of cyclic diguanylate in the regulation of bacterial pathogenesis. Annu Rev Microbiol 61, 131–148.
Tang, H. L., Chiang, M. K., Liou, W. J., Chen, Y. T., Peng, H. L., Chiou, C. S., Liu, K. S., Lu, M. C., Tung, K. C. & Lai, Y. C. (2010).Correlation between Klebsiella pneumoniae carrying pLVPK-derived loci and abscess formation. Eur J Clin Microbiol Infect Dis 29, 689–698. Teng, C. H., Xie, Y., Shin, S., Di Cello, F., Paul-Satyaseela, M., Cai, M. & Kim, K. S. (2006).Effects of ompA deletion on expression of type 1 fimbriae in Escherichia coli K1 strain RS218 and on the association of E. coli with human brain microvascular endothelial cells. Infect Immun 74, 5609–5616.
Tsai, K. W., Lai, H. T., Tsai, T. C., Wu, Y. C., Yang, Y. T., Chen, K. Y., Chen, C. M., Li, Y. S. J. & Chen, C. N. (2009). Difference in the regulation of IL-8 expression induced by uropathogenic E. coli between two kinds of urinary tract epithelial cells. J Biomed Sci 16, 91.
Wilksch, J. J., Yang, J., Clements, A., Gabbe, J. L., Short, K. R., Cao, H. W., Cavaliere, R., James, C. E., Whitchurch, C. B. & other authors (2011).MrkH, a novel c-di-GMP-dependent transcriptional activator, controls Klebsiella pneumoniae biofilm formation by regulating type 3 fimbriae expression. PLoS Pathog 7, e1002204.
Wood, T. K., Gonza´lez Barrios, A. F., Herzberg, M. & Lee, J. (2006). Motility influences biofilm architecture in Escherichia coli. Appl Microbiol Biotechnol 72, 361–367.
Wu, C. C., Huang, Y. J., Fung, C. P. & Peng, H. L. (2010).Regulation of the Klebsiella pneumoniae Kpc fimbriae by the site-specific recombi-nase KpcI. Microbiology 156, 1983–1992.
Xia, Y., Gally, D., Forsman-Semb, K. & Uhlin, B. E. (2000).Regulatory cross-talk between adhesin operons in Escherichia coli: inhibition of type 1 fimbriae expression by the PapB protein. EMBO J 19, 1450–1457. Edited by: D. Demuth

![Fig. 2. Western blot analysis of fimK deletion effects on type 1 fimbriae expression. Expression of the type 1 fimbriae major pilin FimA was determined in (a) FimB recombinase overexpression bacteria CG43S3[pETQ33-fimB], and the derived strains CG43S3DfimK](https://thumb-ap.123doks.com/thumbv2/9libinfo/7629462.134566/6.892.471.824.271.841/analysis-deletion-expression-expression-determined-recombinase-overexpression-bacteria.webp)