Vacuolar Invertases in Sweet Potato: Molecular Cloning,
Characterization, and Analysis of Gene Expression
†L
I-T
INGW
ANG,
‡A
I-Y
UW
ANG,
‡,§C
HANG-W
ENH
SIEH,
‡C
HIH-Y
UC
HEN,
‡ANDH
SIEN-Y
IS
UNG*
,‡,§Institute of Microbiology and Biochemistry and Department of Biochemical Science and Technology, National Taiwan University, Taipei 106, Taiwan
Two cDNAs (Ibβfruct2 and Ibβfruct3) encoding vacuolar invertases were cloned from sweet potato leaves, expressed inPichia pastoris, and the recombinant proteins were purified by ammonium sulfate fractionation and chromatography on Ni-NTA agarose. The deduced amino acid sequences encoded by the cDNAs contained characteristic conserved elements of vacuolar invertases, including the sequence R[G/A/P]xxxGVS[E/D/M]K[S/T/A/R], located in the prepeptide region, Wxxx[M/I/V]LxWQ, located around the starting site of the mature protein, and an intactβ-fructosidase motif. The pH optimum, the substrate specificity, and the apparentKmvalues for sucrose exhibited by the recombinant proteins were similar to those of vacuolar invertases purified from sweet potato leaves and cell suspensions, thus confirming that the proteins encoded by Ibβfruct2 and Ibβfruct3 are vacuolar invertases. Moreover, northern analysis revealed that the expression of the two genes was differentially regulated. With the exception of mature leaves and sprouting storage roots,Ibβfruct2 mRNA is widely expressed among the tissues of the sweet potato and is more abundant in young sink tissues. By contrast,Ibβfruct3 mRNA was only detected in shoots and in young and mature leaves. It appears, therefore, that these two vacuolar invertases play different physiological roles during the development of the sweet potato plant.
KEYWORDS: Vacuolar invertase; cDNA; sweet potato (Ipomoea batatas); recombinant invertase;Pichia pastoris; gene expression
INTRODUCTION
The sweet potato Ipomoea batatas L. is a dicotyledonous plant that belongs to the family Convolvulaceae. The major part of the plant used as human food is its storage root (tuberous roots), which is rich in starch and dietary fiber and is a good source of vitamin A; indeed, 100 g of cooked sweet potatoes provides about 11.5 mg of β-carotene or about four times the U.S. recommended daily allowance (1). The plant is also used as animal feed and for starch extraction. In addition, the leaves of sweet potato are eaten as a green vegetable in parts of Asia (2,
3).
Sweet potato production is affected by several internal and external factors, including the net photosynthetic rate, photo-synthate partition, storage root sink potential, temperature, moisture level, and pathogens (3, 4). It has been proposed that in many plants the sucrose-cleaving enzymes invertase and sucrose synthase play key roles in the control of the photosyn-thate partition and in the determination of the sink strength (5-8
and references therein). Invertase hydrolyzes sucrose into fructose and glucose, whereas sucrose synthase converts sucrose and UDP into fructose and UDP-glucose. Both enzymes have been purified from sweet potato, and their in situ activities have been well-studied (9-12).
As a group, the invertases areβ-fructofuranosidases that have been categorized as cytoplasmic, vacuolar, and cell wall-associated forms based on their solubility properties, subcellular localizations, and pH optima (5). The cytoplasmic form (neutral/ alkaline invertase) exhibits optimal activity within a pH range of 7.0-8.0, whereas both the vacuolar (soluble acid invertase) and the cell wall-associated (extracellular acid invertase) forms exhibit optimal activity at acidic pH values. Functionally, the cytoplasmic enzyme is probably involved in channeling sucrose into catabolism, while the proposed functions of the cell wall-associated enzyme include sucrose partitioning between source and sink organs and regulation of cell differentiation, plant development, and the responses to signals from various biotic and abiotic stresses. Vacuolar invertases have been proposed to play important roles in osmoregulation and cell enlargement, in the regulation of the sugar composition in fruits and storage organs, and in the response to cold (6-8 and references therein). In the previous studies, we have characterized several invertase isoforms and have cloned a cDNA encoding vacuolar †The nucleotide sequences reported in this paper have been submitted
to GenBank under the accession numbers AY037937 and AY037938. * To whom correspondence should be addressed. Tel/Fax: +886-2-23634729. E-mail: [email protected].
‡Institute of Microbiology and Biochemistry. §Department of Biochemical Science and Technology. 3672 J. Agric. Food Chem. 2005, 53, 3672−3678
10.1021/jf0480851 CCC: $30.25 © 2005 American Chemical Society Published on Web 04/09/2005
invertase from sweet potato (10, 13, 14). In this paper, we report the cloning of two other vacuolar invertase cDNAs and the functional expression of their encoded proteins in Pichia
pastoris. To gain insight into the functions of the different
vacuolar invertases in the sweet potato, the expression of the two invertase genes was analyzed by northern hybridization. The results revealed the differentially regulated expression of the two invertase genes and suggested different physiological roles of the different isozymes.
MATERIALS AND METHODS
Plant Materials. Sweet potato (I. batatas L. cv. Tainong 57) plants
were grown in a field in natural light. Leaves and storage roots were collected at various developmental stages, immediately frozen in liquid nitrogen, and stored at -80°C until required.
RNA Isolation. The frozen sweet potato samples were ground into
a fine powder in liquid nitrogen, after which total RNA was isolated using Trizol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. Poly(A)+RNA was purified from the total RNA using biotinylated-oligo(dT) with a PolyATract mRNA Isolation System (Promega, Madison, WI).
Cloning of Invertase cDNA. A cDNA library was constructed from
poly(A)+-RNA isolated from leaves at different developmental stages using a SuperScript Choice System (Invitrogen) and a Lamba ZAP II RI Library Construction Kit (Stratagene, La Jolla, CA). A 1.4 kb sweet potato invertase cDNA fragment (13) was first32P-labeled using the random primer method with a Rediprime DNA Labeling System (Amersham Biosciences, Buckinghamshire, United Kingdom) and then used as a probe to screen the cDNA library. The positive plaques were isolated and subjected to in vivo excision of the pBluescript SK-phagemids from the lambda ZAP II vector.
DNA Sequencing and Sequence Analysis. Both DNA strands were
sequenced using an ABI PRISM BigDye Terminator Cycle Sequencing Ready Reaction Kit with an ABI 3730 XL DNA Analyzer (Applied Biosystems, Foster City, CA). The nucleotide and protein sequence analyses were using the programs of Wisconsin Package Version 10.1 (Accelrys Inc., San Diego, CA). The cleavage site for the leader sequence was predicted by the SPScan program of Wisconsin Package and comparison with other vacuolar invertases.
Preparation of Gene Specific Probes. The DNA probe specific
for Ibβfruct2 was synthesized by polymerase chain reaction (PCR) using the cDNA as a template and gene specific primers selected from the 3′-untranslated region (3′-UTR) [forward, 5′ -CTTGGATGAGTTG-TAGTTCCA-3′(nucleotides 2001-2021); reverse, 5′ -GGGGATACT-TGTAAACTCATC-3′(nucleotides 2180-2196; the underlined nucle-otides were added to adjust the GC content)]. The identity of the amplified DNA fragment was confirmed by sequencing. For Ibβfruct3, the entire Ibβfruct3-1 cDNA was used as a probe. The DNA probe for 18S rRNA was a 1.6 kb DNA fragment containing part of the rice 18S rRNA gene, which was amplified from rice genomic DNA (15). For use in the northern hybridization, the Ibβfruct2 specific probe was labeled with [R-32P]dATP (NEN, Boston, MA) by PCR, while the Ibβfruct3 specific and 18S rRNA specific probes were labeled with [R-32P]dCTP (NEN) using the random primer method.
Northern Analysis. Twenty micrograms of total RNA from each
sample was separated on 1.2% (w/v) agarose gels containing formal-dehyde (16) and then transferred onto positively charged nylon membranes (Immobilon-Ny+, Millipore, Bedford, MA). Following hybridization, the blots were washed twice for 10 min at room temperature with 2× SSPE and 0.1% sodium dodecyl sulfate (SDS), once for 15 min at 42°C with 1× SSPE and 0.1% SDS, and then once for 15 min at 62°C with 0.1× SSPE and 0.1% SDS for the Ibβfruct2 and Ibβfruct3 probes or at 65°C for the 18S rRNA probe. The blots were then exposed to phosphorimaging plates, and the captured imagines were analyzed using a Bio Imaging Analyzer (Fujix BAS1000, Fuji Photo Film, Tokyo, Japan).
Production and Purification of Recombinant Invertases in Yeast.
The coding regions of Ibβfruct2 and Ibβfruct3 without the putative leader sequences were amplified by PCR. The sequences of the primers
were 5′-AAATCGATGTCCGGCGCCGGCAAT-3′(forward primer for Ibβfruct2, nucleotides 355-369), 5′ -GGTCTAGAAACAACTCATC-CAAGGG-3′(reverse primer for Ibβfruct2, nucleotides 1999-2014), 5′-AAGGAATTCATGTCCGGCGGTCGGATTT-3′(forward primer for Ibβfruct3, nucleotides 339-355), and 5′ -GGGGGTTCTAGAAA-CAATTGATTGATGA-3′ (reverse primer for Ibβfruct3, nucleotides 1969-1984). The underlined nucleotides shown in the forward primers were added to incorporate the translation initiation codon and restriction sites (ClaI and EcoRI for Ibβfruct2 and Ibβfruct3, respectively), and those in the reverse primers were added for incorporation of XbaI site. The amplified Ibβfruct2 DNA fragment was digested with ClaI and XbaI and ligated into ClaI-XbaI-digested pPICZRC (Invitrogen) to form pIT2-M. The amplified Ibβfruct3 DNA fragment was digested with EcoRI and XbaI and ligated into plasmid pPICZRA that had been digested with the same restriction enzymes, yielding pIT3-M. To express recombinant invertases in yeast, P. pastoris strain X-33 was transformed with pIT2-M or pIT3-M using a Pichia EasyComp Kit (Invitrogen). Growth of the transformed cells and induction of expression with methanol were carried out as described previously (13), except that the induction time was 24 h.
Purification of the recombinant enzymes was carried out at 0-4 °C. The methanol-induced cultures were centrifuged for 5 min at 6000g, after which solid ammonium sulfate was added to the supernatant. Proteins fractionated by 35-65% saturation with ammonium sulfate were pelleted by centrifugation, dissolved in PB 8.0 (50 mM sodium phosphate, pH 8.0), and dialyzed with the same buffer. The resultant enzyme solution was mixed with a suspension of Ni-NTA agarose [50% (v/v) in equilibrium buffer (50 mM sodium phosphate, pH 8.0, 150 mM NaCl, 5 mM imidazole, 0.1% Triton X-100, 20 mM β-mercap-toethanol)] and then incubated for 1 h at 4°C. The enzyme-Ni-NTA agarose mixture was then packed into a column, washed with buffer A (50 mM sodium phosphate, pH 8.0, 100 mM NaCl, 10 mM imidazole, 0.1% Triton X-100, and 20 mMβ-mercaptoethanol), and then eluted with buffer B (50 mM sodium phosphate, pH 8.0, 300 mM NaCl, 250 mM imidazole, 0.1% Triton X-100, and 20 mMβ-mercaptoethanol). Fractions showing invertase activity were collected.
Enzyme Assay and Protein Analytic Methods. The invertase
activity was assayed at pH 5.0 as described by Sung and Huang (17). The amount of reducing sugar produced was measured using the method of Somogyi-Nelson (18). The protein concentration was determined using the protein dye binding method (19), with bovine serum albumin serving as the standard protein. Enzymatic deglycosylation of the recombinant invertase rIT2 by N-glycosidase F (PNGase F, Roche Applied Science, Mannheim, Germany) was performed as described previously (13) except that the incubation time was 16 h. To complete deglycosylation of the recombinant invertase rIT3, the enzyme was heated at 100°C for 10 min in the presence of 1% SDS before PNGase F was added. SDS-polyacrylamide gel electrophoresis (PAGE) was carried out according to the method of Laemmli (20). The protein bands in gels were stained with Coomassie Blue R-250 or silver nitrate.
RESULTS AND DISCUSSION
Isolation and Characterization of Invertase cDNA Clones from Sweet Potato Leaves. We previously cloned a cDNA (Ibβfruct1) encoding vacuolar invertase from the sink leaves of the sweet potato (13). Northern analysis using the 3′-UTR of Ibβfruct1 as a gene specific probe indicated that Ibβfruct1 mRNA was more abundant in sink leaves than in either source leaves or storage roots. Nevertheless, when the northern blots were hybridized with a probe containing a sequence conserved among plant vacuolar invertases, similar levels of invertase mRNA in sink leaves, source leaves, and storage roots were observed (21). This suggested that at least two isoforms of vacuolar invertase are expressed in sweet potato. To isolate other cDNA clones encoding invertase, a cDNA library was con-structed using mRNA collected from leaves at various stages in their growth. By screening the cDNA library with a sweet potato invertase probe containing the conserved region of plant
vacuolar invertase, 21 potential clones were obtained. Four clones whose insert length was close to that predicted for the full-length vacuolar invertase cDNA were selected for charac-terization. One of the four cDNAs, Ibβfruct2, was 2207 bp in length and included a 42-bp 5′-UTR, a 1974 bp open reading frame, and a 191 bp 3′-UTR. The open reading frame encoded a 71.8 kDa polypeptide with a pI of 4.88. The other three cDNAs, Ibβfruct3-1, Ibβfruct3-2, and Ibβfruct3-3, had identical sequences, except that at their 3′-ends they showed differences in the numbers of TA repeats and in the polyadenylation sites within their 3′-UTRs (Figure 1). All three Ibβfruct3 clones lacked a 5′-UTR sequence. Ibβfruct3-1, which was 2202 bp in length, had the largest open reading frame; its sequence began with an ATG codon and encoded 661 amino acid residues. The calculated molecular mass and pI of the encoded protein were 72.47 kDa and 4.93, respectively. The deduced amino acid sequences encoded by Ibβfruct2 and Ibβfruct3 showed 74% identity to one another and, respectively, 86 and 80% identity to Ibβfruct1. Figure 2 shows the alignment of the deduced amino acid sequences of the three Ibβfruct cDNAs and other dicot vacuolar invertases. With the exception of the N- and C-terminal regions, the sequences are highly homologous. Unexpectedly, the first 146 amino acid residues of Ibβfruct1 and Ibβfruct3 are identical (Figure 2). This might reflect an error that occurred during cloning of the 5′-part of Ibβfruct1 cDNA using 5′-RACE, since the nucleotide sequence of the primer used happened to be the same as the corresponding region in Ibβfruct3.
Plant vacuolar invertases are synthesized as preproproteins with a leader sequence containing a signal peptide and an N-terminal extension that are trimmed off during transport and protein maturation (5). Signal peptides of 63 and 68 amino acid residues were predicted from the deduced amino acid sequences of Ibβfruct2 and Ibβfruct3, respectively. The cleavage site of the N-terminal extension was predicted to be between Gly104 and Ser105 for Ibβfruct2 and between Val113 and Ser114 for
Ibβfruct3. The calculated molecular masses of the mature proteins encoded by Ibβfruct2 and Ibβfruct3 were 61.03 and 60.89 kDa, respectively, and both had a pI of 4.72. The deduced amino acid sequence of Ibβfruct2 contained five potential N-glycosylation sites, while that of Ibβfruct3 contained ten.
The amino acid sequences of plant acid invertases and fructan-metabolizing fructosyltransferases are reported to be highly homologous, sharing conserved regions that include the sucrose-binding motif, RDP motif, and EC motif (22, 23). However, vacuolar type acid invertases are distinguishable from cell wall-associated acid invertases and from fructan-metabolizing fruc-tosyltransferases by the presence of several characteristic amino acid sequence elements, which include the sequence R[G/A/P]-xxxGVS[E/D/M]K[S/T/A/R], located in the prepeptide region, Wxxx[M/I/V]LxWQ, located around the starting site of mature protein, and an intact β-fructosidase motif (sucrose-binding motif) NWMNDPNG (24). The deduced amino acid sequence of Ibβfruct2 contained all three of these conserved sequences [RGVSQGVSEKA (amino acids 90-100), WSNLMLSWQ (amino acids 115-123), and NWMNDPNG (amino acids 134-141)] and did the Ibβfruct3-encoded protein. Notably, the last
Figure 1. Alignment of the 3′-UTR of the three Ibβfruct3 cDNA clones. The TA repeat regions are boxed. The putative polyadenylation signal is underlined.
Figure 2. Alignment of the deduced amino acid sequences derived fromIbβfruct cDNAs and vacuolar invertase cDNAs from other dicot plants. The sequences compared are from carrot (Daucus carota, CAA53097.1), pie cherry (P. cerasus, AAL05427.2), Arabidopsis thaliana (AtβFRUCT3, NP_567498.1; AtβFRUCT4, NP_567901.1), sugar beet (Beta vulgaris, CAD19321.1), potato (Solanum tuberosum, CAA49831.1), and tobacco (Nicotiana tabaccum, CAC83577.2). The characteristic conserved elements of vacuolar invertases are indicated by lines above the sequences.
two residues in theβ-fructosidase motif of the latter were DG instead of NG, which has also been noted in vacuolar invertases from pie cherry (Prunus cerasus, accession number AAL05427.2) and tulip (Tulipa gesneriana, accession number CAA64953). The presence of the three characteristic elements confirmed that
Ibβfruct2 and Ibβfruct3 encode vacuolar type acid invertases and not cell wall-associated acid invertases or fructan-metabolizing fructosyltransferases.
Expression and Characterization of the Ibβfruct2- and
Ibβfruct3-Encoded Proteins in Yeast. To express the
Ibβfruct2-and Ibβfruct3-encoded proteins, P. pastoris cells were, respec-tively, transformed with the expression plasmids pIT2-M and pIT3-M. The putative mature proteins, which contained a c-myc epitope and a His tag fused at their C terminus, were expected to be expressed under the control of the alcohol oxidase (AOX1) promoter and to be secreted into the culture medium via a yeast R-factor secretion signal. Consistent with that expectation, when the transformed cells were grown in the presence of methanol, the acid invertase activity accumulated in the growth medium (Figure 3), such that the activity in the intracellular fraction was only about 10% of that in the growth medium. Table 1 shows the results of the purification using ammonium sulfate fractionation and chromatography on Ni-NTA agarose of the recombinant Ibβfruct2- and Ibβfruct3-encoded proteins (des-ignated rIT2 and rIT3, respectively) from the conditioned media of transformed P. pastoris cells. This two step protocol was sufficient to remove most of the impurities from the samples (Figure 4).
Both rIT2 and rIT3 exhibited a pH optimum of 5.0, with activities declining sharply at higher pH values (data not shown). In addition to sucrose, the two recombinant enzymes also hydrolyzed raffinose and stachyose, although the activities were substantially lower than with sucrose (Table 2) and neither enzyme displayed any activity with cellobiose, maltose, lactose, or inulin. The pH optimum and substrate specificity exhibited by the recombinant proteins are consistent with vacuolar invertases purified from sweet potato leaves (10) and cell suspensions (14). Moreover, the apparent Kmvalues obtained for rIT2 and rIT3 with sucrose (4.97 and 10.1 mM, respectively)
Figure 3. Expression of recombinant invertases inP. pastoris. Cells transformed with pIT2-M (b), pIT3-M (2), pPICZRC (O), or pPICZRA
(4) were grown in media without methanol at 30°C untilA600values
reached 2.0, after which expression of recombinant proteins was induced by adding methanol to the cultures (13). Samples collected at various times after induction were centrifuged at 3000g for 5 min. Proteins secreted into the culture medium (A) and in the intracellular fraction (B) were assayed for invertase activity at pH 5.0.
Table 1. Purification of the Recombinant Invertases rIT2 and rIT3 from TransformedP. pastorisa purification step enzyme total activity (unitsb) total protein (mg) specific activity (units/mg) yield (%) purification (fold) centrifugal supernatant of the culture rIT2 578 7.9 73 100 1.0 rIT3 140 9.7 14 100 1.0 ammonium sulfate fractionation rIT2 377 6.9 55 65 0.8 rIT3 101 8.3 12 72 1.9 Ni-NTA agarose rIT2 166 0.3 553 29 7.6 rIT3 7 0.1 70 5 5.0
aThe data were obtained from the centrifugal supernatant of a 500 mL culture of methanol-inducedP. pastoristransformed with pIT2-M or pIT3-M.bOne unit of invertase was defined as the amount of enzyme that catalyzed the formation of 1 µmol of reducing sugar from sucrose per minute at 37°C and pH 5.0.
Figure 4. Purification of the recombinant invertases rIT2 and rIT3. Protein samples collected at various stages of the purification were separated on 10% SDS-polyacrylamide gels, which were then stained with Coomassie blue R-250. Lane 1, molecular mass markers; lane 2, proteins precipitated by ammonium sulfate (35−65%); and lane 3, proteins purified by Ni-NTA agarose chromatography.
Table 2. Substrate Specificity of the Recombinant Invertases rIT2 and rIT3
relative activity (%)
substrate rIT3 rIT3
sucrose (0.1 M) 100 100 raffinose (0.1 M) 37±1 25±3 stachyose (0.1 M) 13±3 9±2 cellobiose (0.1 M) 0 0 maltose (0.1 M) 0 0 lactose (0.1 M) 0 0 inulin (2%) 0 0
were situated between those for the enzymes purified from leaves (3.05 mM) and from cell suspensions (17.6 mM). Thus, the identity of rIT2 and rIT3 as vacuolar invertases was further confirmed by their functionality.
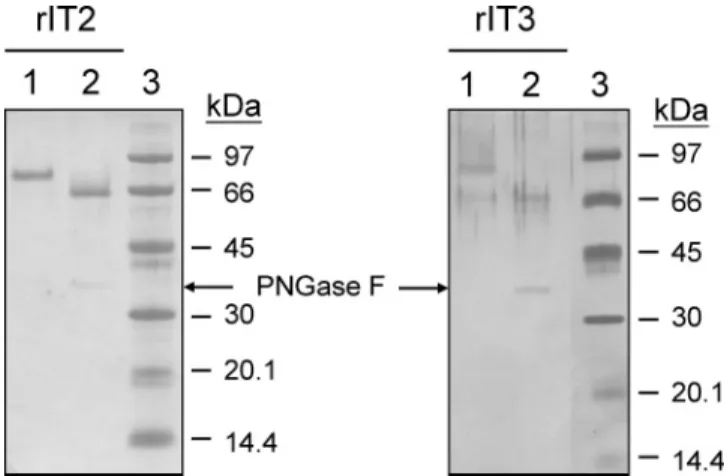
The N-terminal sequences of rIT2 and rIT3 determined by Edman degradation were SGAGNGVS (amino acid residues 105-112 of the cDNA-derived sequence) and SGGRRISY (114-121), respectively. The results were consistent with the expected sequences for the predicted mature proteins. The molecular masses of the purified rIT2 and rIT3 estimated by SDS-PAGE were approximately 82 and 87 kDa, respectively. These values are higher than the calculated molecular masses for the two recombinant enzymes and also higher than those of the enzymes purified from leaves (61 kDa, 10) and cell suspensions (63 kDa, 14). To examine whether the differences in molecular mass were due to glycosylation of the recombinant enzymes, the purified rIT2 and rIT3 were subjected to degly-cosylation with PNGase F (Figure 5). The observed molecular masses of the two deglycosylated enzymes were both about 66 kDa, which are very close to the calculated molecular masses for the c-myc epitope and His tag fused recombinant enzymes (64.03 and 63.29 kDa, respectively). The result indicated that the extent of glycosylation is different in P. pastoris and in sweet potato.
Expression of Ibβfruct2 and Ibβfruct3 Genes in Sweet
Potato. The expression of the two Ibβfruct genes in sweet potato was examined by northern analysis. The hybridization probe specific for Ibβfruct2 was a DNA fragment containing the 3′ -UTR of the gene. For Ibβfruct3, on the other hand, we used the entire Ibβfruct3-1 cDNA as the hybridization probe because the lengths and sequences of the 3′-UTRs differed among the three Ibβfruct3 clones. Although the specificity of the Ibβfruct3 probe was verified by cross-Southern hybridization with full-length Ibβfruct2 and Ibβfruct3 cDNAs (data not shown), the presence of other vacuolar invertase genes that are highly homologous to Ibβfruct3 cannot be ruled out. Consequently, the result obtained using the Ibβfruct3-1 cDNA probe may not reflect exclusively the gene expression of Ibβfruct3.
The observed differences in the hybridization patterns of the two probes revealed that the expression of the two genes is differentially regulated (Figure 6). Levels of Ibβfruct2 mRNA were higher in sprouting shoots, immature leaves, stems, and storage roots than in fibrous roots but were barely detected in
mature leaves and sprouting storage roots. Vacuolar invertases are thought to play an important role during sink initiation and the initial expansion growth of many sinks (8 and references therein). The presence of high levels of Ibβfruct2 mRNA in young sink tissues is consistent with such a function. In contrast to the wide distribution of Ibβfruct2 mRNA, the transcripts detected using the Ibβfruct3 probe were restricted to the shoots and leaves. Furthermore, the changes in the levels of the two gene transcripts during the maturation of the leaves differed. Levels of Ibβfruct2 mRNA were highest in shoots and then gradually declined until they were undetectable in large (mature) leaves. By contrast, levels of Ibβfruct3 mRNA were low in shoots and at a maximum in small leaves, although a significant amount of Ibβfruct3 mRNA was still present in mature leaves. Expression of the two genes thus appears to be developmentally regulated during maturation of the leaves. Notably, the temporal and spatial patterns of the Ibβfruct3-encoded enzyme suggest that, in addition to functioning during the cell expansion of young leaves, it may play a role in the storage of photosynthate in the vacuoles of mature leaves.
In summary, our results reveal that the vacuolar invertases in sweet potato are encoded by at least three genes. Expression of Ibβfruct2 and Ibβfruct3 in sweet potato is both spatially and temporally regulated. The differential expression of the two genes suggests that different vacuolar invertases play different physiological roles during the development of the sweet potato plant. The Ibβfruct2- and Ibβfruct3-encoded proteins were successfully overexpressed and purified from P. pastoris. The recombinant proteins exhibited similar biochemical properties to the vacuolar invertases purified from sweet potato. The results not only confirm the identity of Ibβfruct2 and Ibβfruct3 as vacuolar invertase genes but also indicate that the heterologous expression system together with the two-step easy purification procedure is a highly feasible system to study the structure-function relationships, regulatory and mechanistic properties, and industrial applications of the enzyme.
LITERATURE CITED
(1) U.S. Department of Agriculture, Agricultural Research Service. USDA Nutrient database for standard reference, release 17. Nutrient Data Laboratory Home Page, http://www.nal.usda.gov/ fnic/foodcomp, 2004.
Figure 5. Analysis of the deglyosylated recombinant invertases rIT2 and rIT3. Purified rIT2 and rIT3 were incubated with PNGase F at 37°C for 16 h and then separated on 12.5% SDS−PAGE. The resolved proteins were stained with Coomassie blue R-250 (left panel) or silver nitrate (right panel). Lane 1, purified recombinant invertase; lane 2, purified recombinant invertase treated with PNGase F; and lane 3, molecular mass markers.
Figure 6. Expression ofIbβfruct2 and Ibβfruct3 in sweet potato. Total RNA was isolated from sprouting shoots (Sh), small leaves (S, 2 cm in length), middle-sized leaves (M, 5 cm in length), large leaves (L, 9 cm in length), stems (St), fibrous roots (FR), small storage roots (S, 6 cm in length), large (mature) storage roots (L, 16 cm in length), and sprouting storage roots (Sp). An equal amount of RNA (20µg) was loaded into each lane of a formaldehyde agarose gel and analyzed by northern blot hybridization. Identical blots were hybridized with Ibβfruct2 specific, Ibβfruct3 specific, and 18S rRNA specific probes.
(2) Woolfe, J. A. Sweet potatospast and present. In Sweet Potato: An Untapped Food Resource; Cambridge University Press: Cambridge, 1992; pp 15-40.
(3) Jansson, R. K.; Raman, K. V. Sweet potato pest management: A global overview. In Sweet Potato Pest Management: A Global PerspectiVe; Jansson, R. K., Raman, K. V., Eds; Westview Press: Boulder, Colorado, 1991; pp 1-12.
(4) Kays, S. J.; Magnuson, C. E.; Fares, Y. Assimilation patterns of carbon in developing sweet potatoes using11C and14C. In Sweet Potato. Proceedings of the First International Symposium; Villareal, R. L., Griggs, T. D., Eds; Asian Vegetable Research and Development Center: Tainan, Taiwan, 1982; pp 95-118. (5) Tymowska-Lalanne, Z.; Kreis, M. The plant invertases: Physiol-ogy, biochemistry and molecular biology. In AdVances in Botanical Research; Callow, J. A., Ed.; Academic Press: New York, 1998; Vol. 28, pp 71-117.
(6) Sturm, A.; Tang, G.-Q. The sucrose-cleaving enzymes of plants are crucial for development, growth and carbon partitioning. Trends Plant Sci. 1999, 4, 401-407.
(7) Sturm, A. Invertase. Primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiol. 1999, 121, 1-8.
(8) Koch, K. Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235-246.
(9) Murata, T. Regulatory properties of sucrose synthetase in sweet potato roots. Agri. Biol. Chem. 1971, 35, 297-299.
(10) Wang, Y.-L. Studies on soluble acid invertase from sweet potato leaves. Master thesis, Graduate Institute of Agricultural Chem-istry, National Taiwan University, Taipei, Taiwan, 1995. (11) Takahata, Y.; Noda, T.; Sato, T. Relationship between acid
invertase activity and hexose content in sweet potato storage roots. J. Agric. Food. Chem. 1996, 44, 2063-2066.
(12) Yatomi, M.; Kubota, F.; Saito, K.; Agata, W. Evaluation of root sink ability of sweet potato (Ipomoea batatas Lam.) cultivars on the basis of enzymatic activity in the starch synthesis pathway. J. Agron. Crop. Sci. 1996, 177, 17-23.
(13) Huang, W.-C.; Wang, A.-Y.; Wang, L.-T.; Sung, H.-Y. Expres-sion and characterization of sweet potato invertase in Pichia pastoris. J. Agric. Food. Chem. 2003, 51, 1494-1499.
(14) Wu, L.-J.; Huang, W.-C.; Sung, H.-Y. Partial purification and characterization of soluble acid invertases from sweet potato suspension cells. Food Sci. Agric. Chem. 2000, 2, 49-54. (15) Liao, Y.-C.; Wang, A.-Y. Sugar-modulated gene expression of
sucrose synthase in suspension-cultured cells of rice. Physiol. Plant. 2003, 118, 319-327.
(16) Lehrach, H.; Diamond, D.; Wozney, J. M.; Boedtker, H. RNA molecular weight determinations by gel electrophoresis under denaturing conditions, a critical reexamination. Biochemistry
1977, 16, 4743-4751.
(17) Sung, H.-Y.; Huang, W.-C. Purification and characterization of cell-wall-bound invertase from rice (Oryza satiVa) grains. Biotechnol. Appl. Biochem. 1994, 19, 75-83.
(18) Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375-380.
(19) Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248-254. (20) Laemmli, U. K. Cleavage of structural proteins during assembly
of the head of bacteriophage T4. Nature 1970, 227, 680-685. (21) Huang, W.-C. Cloning and characterization of a cDNA encoding vacuolar invertase from sweet potato leaves, and expression of the cDNA in Pichia pastoris. Ph.D. Thesis, Graduate Institute of Agricultural Chemistry, National Taiwan University, Taipei, Taiwan, 2000.
(22) Vijn, I.; Smeekens, S. Fructan: More than a reserve carbohy-drate? Plant Physiol. 1999, 120, 351-360.
(23) Ritsema, T.; Smeekens, S. C. M. Engineering fructan metabolism in plants. J. Plant Physiol. 2003, 160, 811-820.
(24) Van den Ende, W.; Michiels, A.; Roy, K. L.; Van Laere, A. Cloning of a vacuolar invertase from Belgian endive leaves (Cichorium intybus). Physiol. Plant. 2002, 115, 504-512.
Received for review November 16, 2004. Revised manuscript received March 8, 2005. Accepted March 15, 2005. This work was supported by grants from the National Science Council, the Republic of China (Taiwan).