行政院國家科學委員會專題研究計畫 成果報告
胚胎幹細胞之建立與其生殖細胞系統之分化
計畫類別: 個別型計畫 計畫編號: NSC91-2314-B-002-374- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 國立臺灣大學醫學院婦產科 計畫主持人: 陳信孚 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 92 年 11 月 6 日
I 中文摘要 胚胎幹細胞是多能性細胞,最近有許多 證據顯示這些細胞在發育生物學上,藥物及 毒物之研究、以及移植醫學等方面都非常有 用,具有提供人類器官移植所需之細胞來源 的潛能。根據此研究計畫之原始目標;首先 本年度我們已建立了4株小鼠胚胎幹細胞株 (其中之一是C57BL/6種,另外3株是ICR 種),建立之效率是3.4%,其中二株已成長 30代以上,且注入免疫不全小鼠中,可以長 成三種胚層的細胞。目前我們正致力於提高 整體的建立效率以及對這些胚胎幹細胞之 特性作檢驗。我們已於2月底獲得Monash 大學所建立的人類胚胎幹細胞株(HES2), 並於近期開始進行相關之培養。但是目前的 培養環境仍未臻理想,HES2仍無法維持完 全未分化;不過在HES2分化實驗中倒也觀 察到一些特殊細胞型態,可能於生殖細胞有 關,因此我們正進行檢驗中。此外我們已經 在著手建立人類胚胎幹細胞株,但是由於人 類胚胎比較稀有,因此還未有具體成果,但 是目前包括建立人類胚胎幹細胞株所需之 免 疫 手 術 與 細 胞 培 養 之 各 個 步 驟 皆 已 完 備,且有一個內細胞團發展成類似胚胎幹細 胞之群落,並存活了三代。 因此關於本計畫之原定目標,大部分已 有成果,進度應屬可以接受。我們期待計畫 可以延續,以達成建立有效的人類胚胎幹細 胞培養與維持的能力與步驟,同時設定一個 初步的實驗模式,以進一步研究幹細胞分化 至生殖細胞之過程。 英文摘要
Embryonis stem (ES) cells are pluripotent cells which can differentiate into most lineages in the body. Recently a number of studies indicate that ES cell may be useful in the study of developmental biology and toxicology. These cells may also provide an unlimited source of cells or tissues or organs
for transplantation therapy. According to the original goals, we have derived in total 4 mouse ES cell lines (one is C57BL6 and 3 are ICR strain mice). The effieciency of establishment is about 3.4% and two of the lines (one is C57BL6 and one is ICR strain mice) have been passaged for more than 30 generations. After injection of these ES-like cells into the SCID mice, termatoma formed which contained cells originated from all three germ layers. Recently we are also characterizing these cells by examining the stem cell markers (SSEA-1, SSEA-3, SSEA-4 and alkaline phosphatase) of these ES cell lines. We also are working on establishing human ES cell line from spare human embryo obtained during IVF. However as yet we can only obtain ES-like colony and grow to 3rd passage. In addition, we are also mainiaining the human ES cell line (HES2) obtained from Monash University in Australia, the condition of these cells are also fair and in many circumstances the cell will differentiate. Therefore further refinement of the culture condition is needed. However during the differentiation of the HES2 cells, we observe some morphological changes of the cells which suggested the potential of germ cell differentiation. Therefore further study will hopefully identify the potential of these cells.
II
報告內容
前言Stem cells are unique cells with the ability of self-renewal and are capable of developing into one or sometimes many specialized cell types. Stem cells can roughly be divided into mono- (or multi-) potent and pluripotent ones. As yet, three pluripotent stem cell lines have been isolated – embryonic carcinoma (EC) cells, embryonic stem (ES) cells, and embryonic germ (EG) cells. Among them, ES cell is on the earliest up-stream cell type that theoretically is able to form all the cell lineages in the body. Recent studies suggest that under certain culture condition, ES cell can differentiate into trophoblast cell and even oocytes. These indicate that ES cell is very likely a totipotent cell type.
Human ES cell lines were first isolated by Thomson et al. in 1998 and subsequently be by Reubinoff in 2000. Many other informal reports also claimed the establishment of more than 70 human ES cell lines. These cell lines retain the ability to differentiate into all three primary embryonic germ layers. The study of human ES cells is scientifically very important due at least to the following reasons. First through the observation of ES cell differentiation into a number of cell lineages, the key factors and environmental influences that direct earliest decisions toward specific cell fate may be delineated. Secondly, from clinical and pharmaceutical point of view, the use of ES cells also provides an extremely efficient in vitro model for studying a number of chemicals and toxins. Last, the differentiated product of ES cells will likely provide an unlimited source of tissues or
organs for human transplantation therapy.
Although there have been approximately 79 cell lines registered in NIH, there is as yet no domestic Taiwanese cell line. From the practical point of view, we in Taiwan need to have our own cell lines in order to make more researchers assessable to the cells. In addition, each ES cell line is not the same. It is therefore important to establish a practically workable protocol for researchers in Taiwan to isolate our own ES cell lines. Using these cell lines as a starting point, further study on the in vivo and in vitro differentiation of ES cells can be performed to expand and fulfill the potentiality of these pluripotent stem cells. However isolation of ES cell lines is not easy. Therefore, the original design of this project was first to establish murine ES cell line as a starting point. Using the experience from the animal works and also by learning ES cell culture from foreign lab (e.g. Dr. Pera’s lab in Monash University), we hoped to establish Taiwanese human ES cell line. During the period, we also tried to contact with Monash University about obtaining the established human ES cell line, so that culture and differentiation study can be performed in our lab as soon as possible even before establishment of our own cell line.
研究目的
[1] Modify the protocol of mouse ES cell isolation to increase the efficiency of cell line establishment
[2] Characterize the newly derived mouse ES cell lines
[3] To isolate and establish human ES cell line [4] Culture and maintain the human ES cell line
III
文獻探討
Isolation Of ES Cell Lines
The isolation of murine ES cells has been reported as early as 1981. Recently, successful derivation of primate ES cell has been reported in several groups. It has been found out that morphologically there are some subtle but important differences between primate and murine ES cells and there are also a number of differences in the cell surface markers between the two cell lines.
There have been a number of well established reports on the derivation of ES cell lines from human embryos (Thomson 1998; Reubinoff 2000). Spare embryos donated by patients participating in the in vitro fertilization (IVF) and embryos obtained by fertilization from donated gametes have both been used. According to those reports, the efficiency of isolation is not always satisfactory (a ES cell line production rate from 3/18 to 5/14 cell lines/inner cell mass have been achieved). It is thought that there is still room for improvement of isolation and establishment of human ES cell line.
Defining the pluripotency of a ES cell-like cell line
There are three independent methods [1;2] to test the differentiation potential of ES-like cells once it is established – including in vitro differentiation in a culture dish, injection of ES cell into an immune-compromised mice to produce a teratoma, and chimera formation after injection into a blastocyst. Only the first two methods are practically applicable in human ES cells and both have been used successfully.
Development and Differentiation of ES
cells
The differentiation of ES cells can be achieved by growing the cells in high density, on different feeder layers, by supplement of certain growth factors or introduction of transcription factors, or by culture on extracellular matrix substrates. The developmental and differential potential of murine ES cells have been tested in a big number of studies through trial and error and most denoted to the potential of ES cells to differentiate to many cell lineages when the culture environments are appropriately altered. Accumulating evidences also suggest that human ES cells can behave in a similar fashion.
(Methods of Differentiation Induction)
In vitro suspension culture leads to the formation of embryoid bodies that contains a number of differentiated cell types [1]. However these differentiations are generally undirected and uncontrollable. Addition of growth factors into the culture media is another effective way to induce the differentiation of ES cells [3]. However, the factors for the differentiation of a specific cell type have not been well established and no known single growth factor can induce ES cells to develop into a single cell lineage [3]. Finally coculture with a specific stroma cell lines may be proved to be an useful option since the feeder cells can provide the inherent dynamics of growth factor production that are suitable for the development of ES cells into specific lineage [4;5]. Different stroma types have been employed to promote hematopoietic commitment [4-6] and to direct production of dopaminergic neurons [7]. ES cells are induced to differentiate to hematopoietic cells by coculture with stroma
IV
cell line RP010 and cytokine IL-3 and IL-6 [4]. Nakano et al. showed that on a
macrophage colony-stimulating factor-deficient stroma cell line OP9, mouse
embryonic stem cells were shown to give rise to primitive and definite erythrocytes sequentially [6]. The group in the Kyoto University furthermore identified a stromal cell (PA6 stroma cell) derived inducing activity (SDIA) that promotes the neural differentiation of mouse ES cells [7]. The SDIA accumulates on the surface of the stroma cells and induce efficient differentiation of cocultured ES cells in a serum-free medium without addition of other growth factors. These observations indicate that specific stromal cell lines may provide the unknown but necessary growth factors to direct ES cell to develop to a certain cell lineage. Theoretically the stroma cells should arise from the environment where the target cell of ES cell differentiation is located. (Differentiation to germ cell)
Mouse ES cells have been repeatedly proved to be capable of differentiating to every cell lineage including germ line. Evidences from differentiation of human ES cells into teratoma also suggested that they are pluripotent, and specifically evidences are also accumulating that they can differentiate into germ cells in vivo [1;3;8;9]. However as far as our knowledge is concerned, in vitro development of human ES cells into germ cells or its related tissues have not been tested or, at least not reported previously. Irrespective of the potential ethical concerns that might arise, the study to examine this potential of ES cell has evident merits both in basic science and clinically.
In the search of the factors to direct ES
differentiation into germ cells, there are several candidates that need to be noted. The functional gametes first arise as a migratory population of cells, called the primordial germ cells. These cells migrate to and colonize on the genital ridges during early organogenesis period, and finally give rise to a completely differentiated gonad. During this migration they proliferate and the number of primordial germ cells in mouse embryo increase from less than 100 to approximately 4000. It has been shown that extraembryonic ectoderm-derived factors instruct the pluripotent epiblast cell (a part of inner cell mass of blastocyst) towards restricted primordial germ cell fate during murine gastrulation [10]. BMP4 and BMP8b proteins (which are expressed by extraembryonic ectoderm) produced by COS cells form heterodimer to direct murine primordial germ cell formation [10]. Subsequently, there were studies identifying that the proliferation and migration of primordial germ cells in culture were influenced by the soluble factors released from the genital ridge. It was shown that genital ridges exert long-range effects on murine primordial germ cells, including their cell number and direction of migration in culture [11]. Furthermore, the chemotropic effect is at least partly mediated by TGF- 1 [12]. It is therefore suggested that culture media conditioned by genital ridges might contain some factors that are closely related to this effect. Other feeder layers have been used for germ cells proliferation, including STO cell line, sertoli cells and granulosa cells [13]. Messenger RNA encoding primordial germ cell marker, vasa, could be present for more than 30 days when Zebrafish embryonic cells were co cultured with rainbow trout spleen
V
cell line RTS34st [14]. After this short-term culture, these embryonic cells were able to produce germ line chimera when injected into a host cell. Therefore, co-cultures with these cell lines may be able to direct ES cells toward germ line lineage. Since germ cells survive well on monolayer derived from the cells of embryonic gonads [15], the use of gonad as a feeder of culture is another alternative.
Growth differentiation factor 9 (GDF-9) is a cystine knot-containing hormone of the TGF- superfamily. In GDF-9 null mice, follicle development is arrested at the primary stage and GDF-9 treatment in vitro enhances preantral follicle growth. Presently however, there is no data to indicate the role of GDF-9 in stages during and before formation of primordial follicle.
材料及方法
Establishment of murine ES cell line
In this study, inbred mouse (C57BL/6 mice strain) and outbred ICR strain mouse embryos are used.
Culture medium and LIF
The culture medium for mouse ES cells isolation is DMEM supplemented with 0.1 mM 2-mercaptoethanol, 2 mM glutamine, and 15% FBS (Hyclone), 1% non-essential amino acid, 50-100 U/mL penicillin, and 50-100 g/mL streptomycin. LIF is also used at this stage.
Isolation of mouse ES cells from blastocyst
The steps for isolation follow the original report of Ledermann et al. and Wasserman et al. The blastocyst can be plated on to feeder layer directly and after 4 days of culture, the ICM colonies are collected by a Pasteur pipette and replated onto a new feeder.
After further 7-14 days, ES-like colony is collected and passaged. Blastocysts can also go through a procedure called immunosurgery as described below and the resulting ICM plated onto a feeder.
Establishment of human ES cell line
The protocols of isolation of human ES cell line is basically modified from some previous reports in primates [16](Suemori, Developmental Dynamics, 2001) and human [1;2;17;18].
Obtaining human embryos and in vitro culture to blastocyst
Fresh or frozen human embryos are donated from patients undergoing IVF. 4-8 cell stage embryos are first cultured using G1.2 and G2.2 sequential media until expanded blastocyst stage.
Immunosurgery
Blastocysts are first subjected to proteinase to remove the zona pellucida. Then, the blastocysts are exposed to 1:10 dilution of rabbit antiserum for 30 min, and then exposed to heat-inactivated guinea pig complement for 30 min (1:5 dilution) to remove the trophectoderm.
Culture of ICM and subsequent propagation of ES cells
ICM is cultured on the mitomycin C-treated PMEF feeder using the ES medium. After 3-7 days, expanded ICM is mechanically removed from differentiated cell outgrowth, treated with 0.25% trypsin/1 mM EDTA, and replaced in a new PMEF feeder. After 7-14 days, the resulting ES-like colonies are propagated in clumps of cells (about 50-100 cells/clump) every 3-7 days, using type IV collagenase (1 mg/mL; Gibco/BRL).
Characterization of ES-like cell line
VI
characterized by a number of experiments. Four studies will be done – (1) examination of the ES cell marker expressions (2) examination of teratoma formation after injection into SCID mice (3) examination of length of telomere and the expression of telomerase (4) examination of the karyotype.
Maintenance and culture of HES2 cell line
After thawing of the HES2 cell line, the colony pieces are cultured using the protocol provided by ESI. Differentiation of these cell colonies are also observed and characterized.
結果 (Results)
Establishment of murine ES cell line
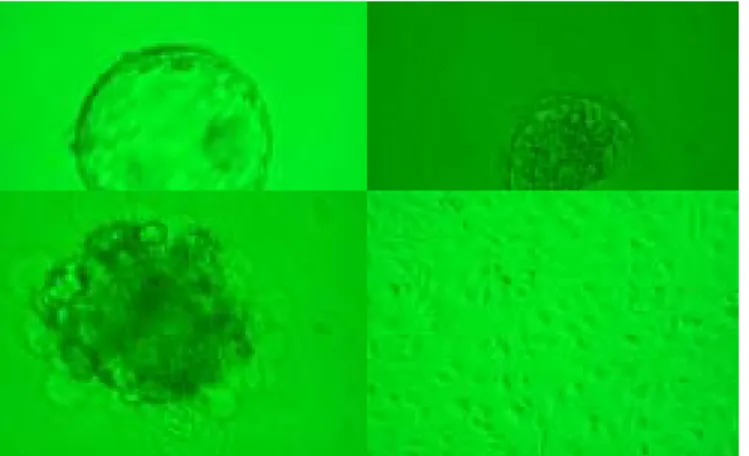
After serial experiments right from scratch, we finally establish 4 murine ES cell lines after many experiments (Table 1 and figure 1). One is from the inbred C57BL/6 mouse and the others from the outbred ICR strain mouses. The C57BL/6 line has been propagated to the 32th passage and was terminated due to contamination. At the 32th passage, the growth and proliferation of this cell line was still very vivid and the gross morphology was normal. Similarly one of the ICR cell line has been propagated to 31th passage and the passage was terminated due to the New Year. Again, the growth and proliferation speed remained perfect at that passage. Now we have ample stocks of these two cell lines in liquid nitrogen. Recently, we have used immuosurgery for mouse ICM harvet and the ICM morphology seems to be improved. At passage 10, both cell lines (one C57BL/6 mouse and one ICR strain) have been injected into SCID mice and both developed teratoma. After removal and stain, these tumors showed cell types from all three germ layers (including neural tissues, bone,
cartilage, GI tissues and renal tissues) (histological slides are available). We are now characterizing these cell lines (two of them) by examining SSEA-1,3, 4 and AP.
Obtain human ES cell line (HES2)
We have obtained a human ES cell line (HES2) from ESI (ES cell International (Figure 2). This cell line was shipped here in late February 2003 and we started to culture this line since June. At present the culture is not smooth yet and further refinement of the culture condition is needed to maintain its undifferentiated state. However in the observation of the in vitro differentiation of HES2, we find out some morphological change which may suggest the differentiation to germ cell lineage. Now we are using immunohistochemical staining and PCR to identify the potential germ cell marker (c-kit, vasa etc.), which hopeful may appear in these cells.
Human ES cell line establishment
Due to the lack of donated human embryos, we have as yet tested on 20 human embryos to establish the ES cell line. However, most of them are poor embryos (1P or 3P embryos, i.e. very likely chromosomally abnormal embryos). The results of immunosurgery on these embryos are basically favorable (Table 2, Figure 3) and one of the tested embryo resulted in a ES cell–like colony which survived and growed for three passages. However as yet no ES line has been established.
討論
The efficiency of establishment of mouse ES cell line at this early stage was approximately 3.3~3.4%. This is comparatively lower than those reported in
VII
literature (which is approximately at the level of 5-20%). Therefore there is still room for improvement. However since the result was obtained at an early stage when we have just learned to establish cell line and we did not use immunosurgery for ICM collection. Therefore, future modification of the protocols and the use of immunosurgery will probably improve the efficiency. Actually we have done immunosurgery on mouse embyos and it was found that the resulting ICM has a better morphology than those without immunosurgery. In addition, the formation of all three germ layer cells after injection into SCID mice indicates that these cell lines have the potential to develop to many cell types in the body. This result (accompanied with prolonged ability to proliferate up to 32 passages) basically fulfills the criteria of embryonic stem cells. However we still need to test other stem cell parameters to confirm the characteristics.
We have tested the viability and proliferation efficiency of HES2 in our lab. Preliminary experience indicate that it is quite difficult to maintain its long-term undifferentiated state. Similar experience actually have been met in 工研院. Therefore we need more time and experience to obtain a smooth culture of this cell line. However the observation of the differentiated cell morphology, which reminds us of the germ cell lineage, suggested that our initial goal to direct human ES cell differentiation to germ cell lineage may be feasible.
The process of human ES cell line establishment is delayed recently basically due to the lack of donated human embryos. However since the feeder layer maintenance and the immunosurgery is basically smooth
recently, we will more actively contact with IVF patients for the possibility of embryo donation. We have come to the idea that the success of ES establishment will depend mostly on the availability of good embryos.
計畫成果自評
The goal of this study was: [1] to isolate and establish mouse ES cell line; [2] to establish Taiwanese human ES cell line; [3] to culture and maintain HES2 cell line; [4] to direct the differentiation of human ES cell to germ cell lineage. The experiments going on in this year are basically agreeable with the original design. Up to now, we have established the mouse ES cell lines, have obtained human ES cell line from Monash and culture, and also have record some preliminary observation of in vitro HES2 cell differentiation. However, the establishment of our own human ES cell line is still going on and not yet successful (which in the original design, we actually did not think it definitely possible to establish in a year). The culture of the HES2 cell cannot be regarded as smooth, neither.
In conclusion, from a practical point of view, the present study basically follows the pre-designed way and the results can be rated as O.K. In addition, the results available up to now are not yet sufficient for publication and one more year of study will be needed for a more thorough analysis of the data. However it is likely that these data would provide some or enormous contribution to the future academic and clinical science.
參考文獻
1. Reubinoff BE, Pera MF, Fong CY, Trounson A, Bongso A. Embryonic stem
VIII
cell lines from human blastocysts: somatic differentiation in vitro. [see comments]. [erratum appears in Nat Biotechnol 2000 May;18(5):559]. Nature Biotechnology 2000; 18: 399-404.
2. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. [see comments]. [erratum appears in Science 1998 Dec 4;282(5395):1827]. Science 1998; 282: 1145-1147.
3. Schuldiner M, Yanuka O, Itskovitz-Eldor J, Melton DA, Benvenisty N. From the cover: effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proceedings of the National Academy of Sciences of the United States of America 2000; 97: 11307-11312.
4. Palacios R, Golunski E, Samaridis J. In vitro generation of hematopoietic stem cells from an embryonic stem cell line. Proceedings of the National Academy of Sciences of the United States of America 1995; 92: 7530-7534.
5. Nakano T, Kodama H, Honjo T. Generation of lymphohematopoietic cells from embryonic stem cells in culture. Science 1994; 265: 1098-1101.
6. Nakano T. In vitro development of hematopoietic system from mouse embryonic stem cells: a new approach for embryonic hematopoiesis. International Journal of Hematology 1996; 65: 1-8.
7. Kawasaki H, Mizuseki K, Nishikawa S, Kaneko S, Kuwana Y, Nakanishi S, Nishikawa SI, Sasai Y. Induction of midbrain dopaminergic neurons from ES
cells by stromal cell-derived inducing activity. Neuron 2000; 28: 31-40.
8. Amit M, Carpenter MK, Inokuma MS, Chiu CP, Harris CP, Waknitz MA, Itskovitz-Eldor J, Thomson JA. Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Developmental Biology 2000; 227: 271-278.
9. Itskovitz-Eldor J, Schuldiner M, Karsenti, D, Eden A, Yanuka O, Amit M, Soreq H, Benvenisty N. Differentiation of human embryonic stem cells into embryoid bodies compromising the three embryonic germ layers. Molecular Medicine 2000; 6: 88-95.
10. Ying Y, Qi X, Zhao GQ. Induction of primordial germ cells from murine epiblasts by synergistic action of BMP4 and BMP8B signaling pathways. Proceedings of the National Academy of Sciences of the United States of America 2001; 98: 7858-7862.
11. Godin I, Wylie C, Heasman J. Genital ridges exert long-range effects on mouse primordial germ cell numbers and direction of migration in culture. Development 1990; 108: 357-363.
12. Godin I, Wylie CC. TGF beta 1 inhibits proliferation and has a chemotropic effect on mouse primordial germ cells in culture. Development 1991; 113: 1451-1457.
13. De Felici M, Siracusa G. Adhesiveness of mouse primordial germ cells to follicular and Sertoli cell monolayers. Journal of Embryology & Experimental Morphology 1985; 87: 87-97.
IX
Collodi P. Production of zebrafish germ-line chimeras from embryo cell cultures. Proceedings of the National Academy of Sciences of the United States of America 2001; 98: 2461-2466. 15. Bachvarova R, Baran MM, Tejblum A.
Development of naked growing mouse oocytes in vitro. Journal of Experimental Zoology 1980; 211: 159-169.
16. Thomson JA, Kalishman J, Golos TG, Durning M, Harris CP, Becker RA, Hearn JP. Isolation of a primate embryonic stem cell line. Proceedings of the National Academy of Sciences of the United States of America 1995; 92: 844-7848.
17. Thomson JA, Odorico JS. Human embryonic stem cell and embryonic germ cell lines. Ophthalmic Genetics 2000; 18: 53-57.
18. Lanzendorf SE, Boyd CA, Wright DL, Muasher S, Oehninger S, Hodgen GD. Use of human gametes obtained from anonymous donors for the production of human embryonic stem cell lines. Fertility & Sterility 2001; 76: 132-137. 19. Ledermann B, Burki K. Establishment of
a germ-line competent C57BL/6 embryonic stem cell line. Experimental Cell Research 1991; 197: 254-258.
20. Thomson JA, Kalishman J, Golos TG, Durning M, Harris CP, Becker RA, Hearn JP. Isolation of a primate embryonic stem cell line. Proceedings of the National Academy of Sciences of the United States of America 1995; 92: 7844-7848.
21. Reubinoff BE, Pera MF, Fong CY, Trounson A, Bongso A. Embryonic stem cell lines from human blastocysts:
somatic differentiation in vitro. [see comments]. [erratum appears in Nat Biotechnol 2000 May;18(5):559]. Nature Biotechnology 2000; 18: 399-404.
22. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. [see comments]. [erratum appears in Science 1998 Dec 4;282(5395):1827]. Science 1998; 282: 1145-1147.
23. Lanzendorf SE, Boyd CA, Wright DL, Muasher S, Oehninger S, Hodgen GD. Use of human gametes obtained from anonymous donors for the production uman embryonic stem cell lines. Fertility & Sterility 2001; 76: 132-137.
24. Solter D, Knowles BB. Immunosurgery of mouse blastocyst. Proceedings of the National Academy of Sciences of the United States of America 1975; 72: 5099-5102.
10
Table 1: Successfully established mouse ES cell lines Mouse strain Number of 3.5 d.p.c. embryo (blastocyst) plated Number of ICM colonies formation Number of ES cell line established ICR 132 116 4 (4/116; 3.3%) C57BL/6 40 30 1 (1/30; 3.4%)
Table 2: Result of ES cell Establishment on Human Embryos Embryo Initial condition Successful
trophectoderm destruction by immunosurgery Successful implantation of ICM on feeder ES cell line formation Fresh embryos 12 (mostly 1P or 3P) 8 6 0 Frozen embryo 3 (morphologically good in 1) 3 3 0
11
Figure 1: mouse ES cell line (ICR strain, left; C57BL6, right)
Figure 2: human ES cell line (HES2)
Figure 3: Immunosurgery