Plant Growth Regulation 27: 189–192, 1999.
© 1999 Kluwer Academic Publishers. Printed in the Netherlands. 189
Importance of ornithine-δ-aminotransferase to proline accumulation
caused by water stress in detached rice leaves
Chao-Wei Yang & Ching Huei Kao
∗Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China (∗author for corres-pondence: fax: 886-2-23620879)
Received 29 October 1998; accepted in revised form 28 January 1999
Key words: ornithine-δ-aminotransferase, proline accumulation, Oryza sativa, water stress
Abstract
Proline is synthesized either from glutamate or from ornithine in plants. Relatively little is known about the contribution of the pathway from ornithine to proline biosynthesis. In this paper we investigated the contribution of ornithine-δ-aminotransterase (OAT), an enzyme responsible for ornithine pathway, to proline accumulation in water-stressed detached rice leaves. Although OAT activity increased with the increase of water stress duration, a pattern similar to that obtained for proline accumulation, the ornithine pathway in rice leaves seems to contribute little, if any, to proline accumulation under water stress condition. This conclusion was based on the observations that (a) gabaculine (50 µM), an inhibitor of OAT, inhibited about 75% OAT activity caused by water stress but reduced only 20% of proline content and (b) cycloheximide, a protein synthesis inhibitor, had no effect on OAT activity induced by water stress but significantly reduced proline accumulation.
Abbreviations: CHI – cycloheximide; GAB – gabaculine; OAT – ornithine-δ-aminotransferase; WS – water stress
1. Introduction
Proline has been shown to accumulate in plant tissues under various types of stress such as high salinity and water stress [5, 11, 12, 13, 16]. The actual role of that proline, however, remains unclear [15], but has been speculated to have involvement in osmoregula-tion, maintenance of protein solubility, stablization of membrane phospholipids and provision of a store of carbon, nitrogen and energy.
It has been suggested that, in plants, proline is synthesized from glutamate [18]. The biosynthetic pathway from glutamate is thought to involve the conversion of glutamate to proline via the interme-diates γ -glutamyl phosphate, glutamate-γ -semialde-hyde and pyrroline-5-carboxylate, as is the case first established in bacteria. It has been shown from labeling experiments that ornithine can also serve as a precursor to proline in higher plants [3, 4, 6]. In animal systems, OAT has been shown to participate in proline synthesis by producing pyrroline-5-carboxylate from
ornithine and α-ketoglutarate [1]. The isolation of cDNAs encoding OAT in higher plants [7, 14] suggests that OAT participates in proline biosynthesis via the ornithine pathway in higher plants. The role of the glutamate pathway in proline accumulation under stress conditions is well established [18]. However, the importance of the relative contribution of the ornithine pathway in proline accumulation during stress is still a matter of discussion [9, 14]. In this paper, we shall examine the contribution of the ornithine pathway to proline accumulation in detached rice leaves induced by water stress.
2. Materials and methods
Rice (Oryza sativa cv. Taichung Native 1) seedlings were cultured as previously described [10]. The apical 3 cm segments excised from the third leaves of 12-d-old seedlings were used.
Water stress was applied by a previously described method [11]. Groups of twenty-leaf segments were
190
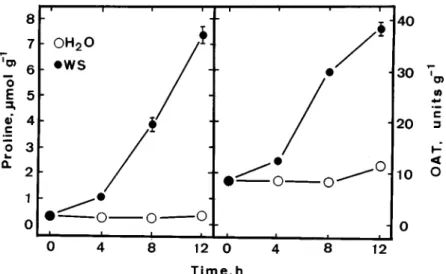
Figure 1. Time course of the effect of water stress (WS) on proline content and OAT activity in detached rice leaves. Vertical bars represent
standard errors (n = 4).
exposed to vapor above a solution of 0.5 M NaCl. Similar segments of leaves were floated on 20 mL of distilled water in a Petri dish to serve as turgid controls. All samples were kept at 27◦C under light (40 µmol m−2s−1) provided by fluorescent lamps.
Proline was extracted and its concentration deter-mined following the method of Bates et al. [2]. Leaf segments were homogenized in 3% (w/v) salfosali-cylic acid and centrifuged. The supernatant fluid was treated with acetic acid and acid-ninhydrin, boiled for 1 h and absorbance was determined at 520 nm. Proline content was expressed as nmol g−1initial fresh weight.
For extraction of enzyme, leaf tissues were homo-genized with 100 mM potassium phosphate buffer (pH 7.4, containing 1 mM pyridoxal-5-phosphate, 1 mM EDTA, and 10 mM β-mercaptoethanol) in a chilled pestle and mortar. The homogenate was centrifuged at 12,000 g for 20 min and the resulting supernatant was used for determination of the OAT activity. The whole extraction procedure was carried out at 4 ◦C. OAT activity was assayed according to Vogel and Kopac [17]. In a final volume of 1 mL, the incubation medium contained: 50 mM ornithine, 20 mM α-ketoglutarate, 1 mM pyridoxal 5-phosphate and 100 mM potassium phosphate buffer, the final pH was 8.0. Reaction was initialed by adding α-ketoglutarate. The incubation was carried out at 37◦C for 30 min. The reaction was stopped by adding 0.5 mL trichloroacetic acid (10%) and color was developed by incubating the reaction mixture with 0.5 mL o-aminobenzaldehyde (0.5%) in ethanol (95%) for 1 h. After centrifugation at 12,000 g
for 10 min, the clear supernatant was taken to measure the optical density at 440 nm. One unit of OAT activity is defined as an increase of 1 A440per h. OAT activity
was expressed on the basis of g initial fresh weight.
3. Results and discussion
Proline has been shown to accumulate in various plant tissues in response to water stress [11, 12, 13, 16]. The time course of the effect of water stress on proline accumulation in detached rice leaves in the light is shown in Figure 1. In turgid control leaves, proline remained relatively unchanged during 12 h of incuba-tion. Water stress treatment resulted in a significant increase in proline content. The induction of proline accumulation by water stress in detached rice leaves is a rapid process. Proline accumulation is evident 4 h after water stress.
To determine the role of the ornithine pathway for proline accumulation caused by water stress, the effect of water stress on OAT activity was examined. The results are shown in Figure 2. It is clear that OAT activity increased with the increase of water stress duration similar to that observed for proline accumulation.
Gabaculine (GAB) at low concentration has been demonstrated to be an effective inhibitor of OAT in radish cotyledons [8]. Hervieu et al. [8, 9] considered that GAB is an useful tool to study the physiological consequence of OAT inhibition in plants. To charac-terize further the importance of the ornithine pathway
191
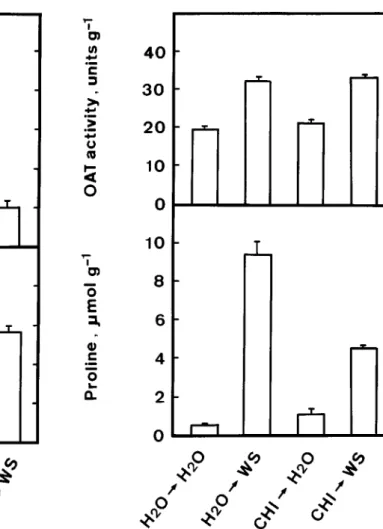
Figure 2. Effect of gabaculine on proline content and OAT activity
in detached rice leaves treated with water stress. Leaf segments were pretreated with either water or 50 µM gabaculine for 12 h and then treated with either water or water stress for 12 h in the light. Bars represent standard errors (n = 4).
for proline accumulation by water stress in detached rice leaves, the effect of GAB on OAT activity and proline accumulation was investigated. As expected, GAB was observed to be very effective in reducing OAT activity induced by water stress (Figure 2). GAB (50 µM) treatment resulted in about 75% decrease in OAT activity in detached rice leaves exposed to water stress (Figure 2). If the ornithine pathway plays an important role in regulating proline accumula-tion cause by water stress in detached rice leaves, GAB treatment is expected to decrease proline content markedly in water-stressed leaf tissues. However, only slight decrease (about 20%) of proline content in water-stressed rice leaves was obtained for GAB
treat-Figure 3. Effect of CHI on proline content and OAT activity in
detached rice leaves treated with water stress. Leaf segments were pretreated with either water or 10 µM CHI for 6 h and then treated with either water or water stress for 12 h in the light. Bars represent standard errors (n = 4).
ment, suggesting that the ornithine pathway is unlikely to be the major pathway for proline accumulation in detached rice leaves exposed to water stress.
Cycloheximide (CHI) treatment had no effect on OAT activity of water-stressed rice leaves (Figure 3), suggesting that an increase in OAT activity requires no protein synthesis. However, CHI treatment resulted in a significant reduction of proline level in detached rice leaves exposed to water stress. It is most likely that CHI inhibited the synthesis of enzymes responsible for glutamate pathway, which is turn caused the reduction of proline synthesis in water-stressed rice leaves.
Hervieu et al. [9] demonstrated that the ornithine pathway contributed, via an increase of OAT activity, to proline synthesis, as well as the glutamate pathway, in salt-treated radish cotyledons. Roosens et al. [14]
192
also concluded that the ornithine pathway could serve an important role in very young, but not adult,
Arabidopsis thaliana plantlets. Based on the data
presented here, we feel that the ornithine pathway in rice leaves contributes little if any to proline accumu-lation under water stress condition.
Acknowledgements
This work was funded by the National Science Council of the Republic of China.
References
1. Adams E and Frank (1980) Metabolism of proline and hydroxyproline. Annu Rev Biochem 49: 1005–1061 2. Bates LS, Waldren RP and Teare ID (1973) Rapid
determina-tion of the free proline for water stress studies. Plant Soil 39: 205–207
3. Brown DH and Fowden L (1966) Metabolism of δ-acetylor-nithine in two Legumious species. Phytochemistry 5: 887–892 4. Chiang H-H and Dandekar AM (1995) Regulation of proline accumulation in Arabidopsis thaliana (L.) Heynh during development and in response to desication. Plant Cell Environ 18: 1280–1290
5. Chou IT, Chen CT and Kao CH (1991) Characteristics of the induction of the accumulation of proline by abscisic acid and isobutyric acid in detached rice leaves. Plant Cell Physiol 32: 269–272
6. Coleman RG and Hegarty MP (1957) Metabolism of D, L-ornithine-2 14C in normal and potassium-deficient barley. Nature 179: 376–377
7. Delanney AJ, Hu CAA, Kavi Kishor PB and Verma DPS (1993) Cloning of ornithine δ-aminotransferase cDNA from
Vigna aconitifolia by trans-complementation in Escherichia coli and regulation of proline biosynthesis. J Biol Chem 268:
18673–18678
8. Hervieu F, Le Dily F, Le Saos J, Billard J-P and Huault C (1993) Inhibition of plant ornithine aminotransferase by gabaculine and 4-amino-5-hexynoic acid. Phytochemistry 34: 1231–1234
9. Hervieu F, Le Dily F, Huault C and Billard J-P (1995) Contri-bution of ornithine aminotransferase to proline accumulation in NaCl-treated radish cotyledons. Plant Cell Environ 18: 205–210
10. Kao CH (1980) Senescence of rice leaves IV. Influence of benzyladenine on chlorophyll degradation. Plant Cell Physiol 21: 1255–1262
11. Kao CH (1981) Senescence of rice leaves VI. Comparative study of the metabolic changes of senescing turgid and water-stressed excised leaves. Plant Cell Physiol 22: 683–688 12. Moftah AE and Michel BE (1987) The effect of sodium
chloride on solute potential and proline accumulation in soybean leaves. Plant Physiol 83: 238–240
13. Munns R, Rrady CJ and Barlow EW (1979) Solute accumula-tion in the apex and leaves of wheat during water stress. Aust J Plant Physiol 6: 379–389
14. Roosens NHCJ, Thu TT, Iskandar HM and Jacobs M (1998) Isolation of the ornithine-δ-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana. Plant physiol 117: 263–271
15. Stewart GR and Larher F (1980) Accumulation of amino acids and related compounds in relation to environmental stress. In: Miflin BJ (ed) Biochemistry of Plants, Vol 5. Amino Acids and Derivatives. New York: Academic Press, pp 609–635 16. Venekamp JH and Kost JTM (1988) The sources of free
proline and asparagine in field bean plants, Vicia faba L., during and after a short period of water withholding. J Plant Physiol 132: 102–109
17. Vogel RH and Kopac MJ (1960) Some properties of
ornithine-δ-transaminase from Neurosproa. Biochem Biophys Acta 37:
539–540
18. Yoshiba Y, Kiyosue T, Nakashima K, Yamaguchi-Shinozaki K and Shinozaki K (1997) Regulation of levels of proline as an osmolyte in plants under water stress. Plant Cell Physiol 38: 1095–1102