行政院國家科學委員會專題研究計畫 成果報告
P53 基因在眼翳中的突變型態及其和 p53 蛋白表現的關係
計畫類別: 個別型計畫 計畫編號: NSC94-2314-B-039-020- 執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 中國醫藥大學醫學系 計畫主持人: 蔡宜佑 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 95 年 10 月 11 日
中文摘要 目的:翳狀贅肉(pterygium)長久以來被認為是一種退化性疾病,但在發現翳狀贅 肉中的 p53 蛋白會有異常表現後,翳狀贅肉被認為可能是一種腫瘤。但是翳狀贅 肉中是否真有 p53 基因突變及 p53 基因發生突變的成因至今未明。本研究首先分 析翳狀贅肉組織中 p53 基因之突變率、突變形式及和 p53 蛋白表現程度間的關係。 方法:我們收集 51 個翳狀贅肉組織,在其上皮組織以 DNA 定序法來定序 p53 基 因的 exons 4 到 8 的突變率和突變形式。同時以免疫組織化學法 (immunohistochemistry)分析這 51 個翳狀贅肉組織的 p53 蛋白表現。我們同時選 擇 10 個正常結膜組織當控制組。 結果:在 8 (15.7%)個翳狀贅肉中發現有 p53 基因突變,且每一個組織只發現一 個突變。全部的突變形式均為點突變(point mutation),其中有 6 個為取代 (substitution)突變,2 個為缺失(deletion)突變。這 8 個突變有 3 個在 exon 6、2 個 在 exon 7、其餘 exon 4、5、8 各發現 1 個突變。P53 蛋白的免疫組織化學法表現 可分為 4 組:31 個翳狀贅肉標本(60.8%)為 0 (陰性), 9 個(17.6%)為+,5 個 (9.8%)為 ++,6 個(11.8%)為+++。每一組剛好各有 2 個突變樣本。那 2 個缺失 (deletion)突變,因導致框架移位(frame-shift),所以沒有 P53 蛋白表現(陰性),而 6 個取代(substitution)突變則有+到+++的蛋白表現。
結論:翳狀贅肉中的確有 p53 基因突變的存在,而其突變是屬於和氧化性壓力及 紫外線有關的突變形式。而 p53 基因突變的有無和蛋白質表現的強弱並無完全的 一致性。
關鍵詞
Abstract
Purpose. To investigate the spectrum of p53 gene mutations, and the relationship
between gene mutation and p53 protein levels in pterygium.
Methods. Pterygial samples were harvested from 51 patients undergoing pterygium
surgery. DNA samples for p53 mutation analyses were extracted from epithelial cells
and subjected to DNA sequencing for examination of mutations in exons 4, 5, 6, 7,
and 8 of the p53 gene. In situ levels of p53 protein were studied by
immunohistochemistry (IHC) and the percentage of positively stained cells quantified.
Ten normal conjunctiva samples were included in this study as controls.
Results. Mutations within the p53 gene were detected in 8 pterygial samples (15.7%)
with only one mutation found in each sample. All the mutations observed were point
mutations, with 6 being substitutions and 2 deletions. Three mutations were identified
in exon 6, two in exon 7 and a single mutation found in each of exons 4, 5, and 8. P53
protein levels were scored as 0 (negative) in 31 pterygial specimens (60.8%), + in 9
samples (17.6%), ++ in 5 samples (9.8%), and +++ in 6 samples (11.8%) by IHC. The
8 samples found to have p53 gene mutations were equally distributed among the
two deletion mutations, which caused a frame-shift to occur, were found in samples
negative for p53 immunostaining (score 0), while substitution mutations were found
in samples positively stained (score +, ++, and +++).
Conclusions. Mutations within p53 gene exons 4-8 were detected in pterygial
epithelium and the mutations showed no correlation with p53 protein levels as seen by
Key words:
Introduction
The p53 gene is a tumor suppressor gene with important functions in cell cycle
control and the prevention of tumor formation. Mutations in the p53 gene could lead
to tumor formation and have been found in various types of tumor cells.1 Previous
studies of the p53 gene have been conducted using either DNA sequencing or via
immunohistochemical (IHC) detection of the p53 protein.
Pterygium is a chronic condition characterized by the encroachment of a fleshy
triangle of conjunctival tissue into the cornea. Pterygium has long been considered a
degenerative condition; however, after abnormal levels of the p53 protein being found
in epithelium, some researchers feel that pterygium is a UV-related, uncontrolled cell
proliferation consistent with that of a tumor. 2-7
Nearly all previous reports of the p53 gene in pterygium have been conducted
usingIHC staining. The reported prevalence of p53 positive staining, or presumed p53
gene mutation, varies widely, from 7.9% to100%.2-9 In normal, unstressed cells, p53 is
a short-lived protein, which is maintained at low, often undetectable, levels in the cell.
Mutations in the p53 gene are believed to lead to an increased stability of its protein in
the cell, allowing its detection by antibodies recognizing several p53 epitopes. In
previous studies, over 85% of mutations detected in the p53 gene were miss-sense
immunostaining. However, other mutations, including non-in-frame deletions, splice
mutations, and non-sense mutations, were negative following IHC. 10-13 Therefore,
DNA sequencing is a far more accurate means to evaluate nature of p53 gene
mutations.12
To our knowledge, there have only been two previous studies using DNA
sequencing to identify p53 gene mutations in pterygium. In the study of Reisman et al,
they found that the p53 gene had undergone a mono-allelic deletion, but the remaining
allele remained wild-type.13 In the study of Shimmura et al, they reported that no
mutation was found in exons 5 through 8 of the p53 gene. 14 However, only 9 and 6
cases, respectively, were examined in these two studies.
In this study, we used DNA sequencing to detect mutations of the p53 gene in
exons 4 to 8 in 51 pterygium specimens. P53 protein is reportedly found only in the
thin layer of epithelium, 2-9 therefore, laser capture micro-dissection (LCM) was used
to remove the sub-epithelial fibrovascular layer to then permit isolation of only the
epithelial cells. P53 protein levels in these cells were also evaluated by IHC. Finally,
Patients and methods
Pterygial samples were harvested from 51 patients undergoing pterygium surgery.
All specimens were formalin-fixed and paraffin-embedded. Sections (3 µm thick)
were cut, mounted on glass, and dried overnight at 37℃ for IHC and DNA sequence
analysis.
DNA sequence analysis
Mutations in exons 4, 5, 6, 7, and 8 of the p53 gene were determined by direct
sequencing. DNA was extracted from the paraffin-embedded pterygium tissues. As
pterygial epithelial cells are difficult to capture by LCM directly, we removed the
sub-epithelial fibrovascular layer first by LCM and left the epithelium intact as shown
in Fig. 1. DNA lysis buffer was applied to lyse the epithelial cells on a glass slide and
then the DNA solution transferred into an eppendorf tube for traditional proteinase K
digestion and phenol-chloroform extraction. Finally, the DNA was precipitated by
ethanol with the addition of linear polyacrylamide to increase the amount of DNA
available15. Target sequences were amplified in a 50 µl reaction mixture containing 20
pmol of each specific primer, 2.5 units of Taq polymerase (TAKARA Shuzo, Shiga),
0.5 mM dNTPs, 5 µl PCR reaction buffer, and 1 µl genomic DNA as the template.
amplification of long DNA fragments and, therefore the PCR products amplified
ranged from only 200 to 400 bp. β-actin, was used as an internal control. The
primers used for p53 gene sequencing were E4S (5’acctggtcctctgactgctc3’), E4S 1
(5’cagcagctcctacaccggcg3’) and E4AS (5’aggcattgaagtctcatgga3’) for p53 exon 4,
E5S (5’tgccctgactttcaactctg3’), and E5AS (5’gctgctcaccatcgctatc3’) for p53 exon 5,
E6S (5’ctgattcctcactgattgct3’) and E6AS (5’agttgcaaaccagacctcagg3’) for p53 exon 6,
E7S (5’cctgtgttatctcctaggttg3’) and E7AS (5’gcacagcaggccagtgtgca3’) for p53 exon 7,
and E8S (5’gacctgatttccttactgcc3’) and E8AS (5’tctcctccaccgcttcttgt3’) for p53 exon 8.
An initial cycle was performed for 5 min at 94℃, followed by 35 cycles each for 40
sec at 94℃, 40 sec at 54℃, and 1 min at 72℃. The PCR products were sequenced by
an auto-sequencing system (Applied Biosystems 3100 Avant Genetic Analyzer). All of
the p53 mutations identified were confirmed by direct sequence analysis of both
strands.
IHC analysis of p53 protein levels
All sections were de-paraffinized in xylene, re-hydrated through a graded series
of alcohols, and washed in phosphate-buffered saline. This buffer was used for all
subsequent washes. IHC using the streptavidin-biotin-peroxidase method was
(Dako, CA, USA, diluted 1:100), which recognizes the N-terminus of the human p53
protein (amino acids 19 to 26). In addition, the antibody reacts with both wild-type
and many mutant p53 proteins. The IHC results were scored for the percentage of
cells with positive reactivity: score 0, being no positive staining in the nuclei of any
cells; score +, from 1% to 10% of cells viewed immunoreactive; score ++, from 11% -
50%; score +++, more than 50% positive cells. The normal conjunctival samples,
collected from 10 patients without pterygium or pinguecula when they underwent
cataract or vitreoretinal surgery, were included as normal controls.
This study was conducted with the approval of the Human Study Committee of
the China Medical University Hospital and National Cheng Kung University Hospital.
Results
There were 31 males and 20 females in the pterygium group (ranging from 50 to
83 years of age; average of 64.2), and 6 males and 4 females in the normal control
group (ranging from 55 to 81 years of age; average of 68.3).
DNA sequencing
P53 mutations were detected in 8 of the 51 (i.e. 15.7%) pterygial samples and
each of these 8 had only one form of mutation. The spectrum of mutations identified
in the exons examined of the p53 gene are shown in Table 1. Slightly more than one
third of the mutations occurred in exon 6 (37.5%) and one fourth (25%) in exon 7. All
eight mutations were point mutations, with 6 (75%) of the 8 mutations being
substitutions and 2 (25%) being deletions. The 6 substitutions were miss-sense and the
two deletions resulted in a frame-shift.
IHC analysis
Thirty-one (60.8%) pterygial specimens were scored as 0 for IHC i.e. no
immunoreactive cells visible, 9 (17.6%) were scored +, 5 (9.8%) were ++, and 6
(11.8%) were +++. The p53 positive rate was 21.6%, if score 0 and + were considered
level at 10%). If only score 0 was considered to be negative and +, ++, and +++ to be
positive (setting cutoff level at 1%), the p53 positive rate was 39.2%.
P53 staining was limited to the nuclei of cells in the epithelial layer. No
substantial staining was visible in the sub-epithelial fibrovascular layers. In normal
conjunctiva group, all specimens were negative.
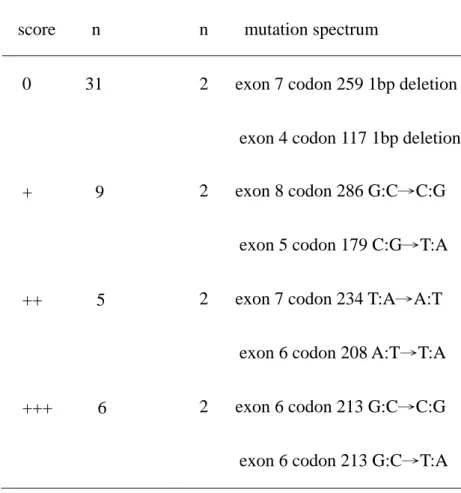
Concordance between IHC and DNA sequencing
The relationship between p53 IHC protein level and gene mutation spectrum is
shown in Table 2. Mutations were detected in each IHC score group. Deletion
mutations were found in the group with no p53 protein IHC reactivity (IHC score 0),
and substitution mutations found in groups with p53 IHC reactivity (IHC scores +, ++,
Discussion
To the best of our knowledge, there have only been 10 studies to date looking at
p53 in pterygium.2-9, 13, 14 Eight of these studies used IHC to examine p53 protein
levels to detect p53 gene mutations, 2-9 and the other two studies used Western transfer
analysis and enzyme-linked immunosorbent assays. 13, 14 In the 8 IHC studies, the
reported p53 positive rate, or the presumed prevalence of p53 gene mutations, varied
widely, i.e. 7.9%, 36.8%, 37.5%, 38.1%, 50%, 53.8%, 60% and 100%. 2-9 Although
the biological causes of such variation in the prevalence of p53 positive staining in
these reports are unclear, we believe that the different cutoff levels are largely to
blame. In our series, 21.6% of all the specimens showed p53 staining at a cutoff level
of 10% and the positive rate would double to 39.2% at a cutoff level of 1%. It has
been reported that p53 immunoreactivity in less than 10% of cells was commonly
associated with a high proliferative activity, not with p53 gene mutation.16 In our
series, setting the cutoff level at 10% (21.6%) is near the prevalence of p53 gene
mutation by the DNA sequencing (15.7%) than setting at 1% (39.2%).
DNA sequencing was reported to be more accurate than IHC for detection of p53
gene mutations. 12 IHC simply reflects the levels of protein available for
immunoreactivity in cells and mutations in a gene may not affect the immunoreactivty
the resultant p53 protein produced and showed strong positive immunostaining.
Samples with deletion mutations, which resulted in stop codons or truncated proteins,
did not react with the antibody and therefore, little, if any, staining was observed. The
wild-type p53 protein was usually not immunostained due to the short half-life (6-20
min); however,sometimes the wild-type p53 protein was immunostained. 12, 17- 19
Hence, drawing conclusions about the gene simply from examination of the protein
levels viewed by IHC is perilous. In our results, miss-sense mutations resulted in
positive staining by IHC, whereas deletion mutations resulted in negative staining.
Because there were specimens with mutations in the p53 negative staining group and
specimens without mutations in the p53 positive staining group, our results suggest
IHC is not the most appropriate method to assess the prevalence of p53 gene mutation
in pterygium.
Mutations in the p53 gene were found in 15.7% of our cases, determined via
DNA sequencing. The result was very different from the previous studies of Reisman
et al and Shimmura et al. 13, 14 Reisman et al found that the p53 gene had undergone a
mono-allelic deletion, and the remaining allele remained wild-type in 9 American
patients, and Shimmura et al reported that no mutation was found in exons 5 to 8 in 6
Japanese. 13, 14 Since there were mutations in 8 of our 51 Taiwanese patients, such
LCM, which presumably minimized the chances of the epithelium containing many
fibroblast or other types of cells that may have contaminated the samples in the
previous studies.
LCM under direct microscopic visualization enables rapid one-step procurement
of selected human cell populations from a histological section. This method has made
micro-dissection of selected areas much easier, so that accurate study of specific target
lesions is possible. As p53 staining was previously reported to be found only in the
epithelial layer of pterygium and not in the sub-epithelial fibrovascular layer, the p53
gene mutation was presumed to be in the epithelial cells. 2-7 Hence, if DNA or RNA
was extracted from whole excised pterygium, including epithelial cells and many
other normal cells, it would be difficult to get specific and accurate results for
epithelial cells alone. However, using the LCM method, we selectively
micro-dissected epithelial cells from pterygial samples, which permitted confident
examination of p53 gene mutations in the epithelial cells alone.
All the mutations in our cases were point mutations. These included single base
substitutions (including miss-sense mutations, non-sense mutations, silent mutations
and single base changes at intronic sites), deletions, and insertions. 20,21 Most of the
mutations (6/8, 75%) in our series were substitutions that led to miss-sense changes
resulted in frame-shift mutations and negative staining by IHC. No insertion
mutations were found in our series.
Pterygium formation has been reported to be related to the dose of UV
irradiation.22 UV irradiation mainly produces DNA lesions between adjacent
pyrimidines, and C to T transitions on dipyrimidine sites or CC to TT tandem
mutations in the p53 gene are considered as the UV-related skin cancer molecular
signatures.23 In our series, there was one case with a C to T transition, but no CC to
TT tandem mutations.
In humans, different DNA damage is repaired by different DNA repair systems.
24
Single base changes are normally reversed by the base excision repair (BER)
system and tandem mutations repaired by the nucleotide excision repair (NER)
system.25 Hence, the presence of a single base change of C to T transition or tandem
mutation of CC to TT is reported to be related to perturbations in the different DNA
repair systems.23 CC to TT tandem mutations are usually associated with deficiency in
the NER, such as in patients with xeroderma pigmentosum (XP). 23 Thirty-six to
sixty-one percent of XP patients with skin cancer were reported to be CC to TT
tandem mutations, which are very rare (<10%) in non-XP patients. 23 In non-XP
patients with skin cancer, 50-60% of the mutations seen are C to T transitions. 23 In
but no CC to TT tandem mutations, suggesting that pterygium patients have defective
BER systems. In the study of Kau et al, polymorphism of HOGG1, one of the BER
genes, was reported to be associated with pterygium formation. 26 In our unpublished
study, polymorphisms of XPA and XPD, two NER genes, were not associated with
pterygium formation, while polymorphism of XRCC1, one of the BER genes, was
reported to be associated with pterygium formation.
Mutational ‘hot spots’ in UV-related cutaneous squamous cell carcinoma (SCC)
and basal cell carcinoma (BCC), including codons 177, 196, 245, 248 and 278, have
been reported. 23 None of the ‘hot spots’ were found altered in our studies. However, a
mutation at codon 213 was detected in 2 cases in our study. If we considered codon
213 as a ‘hot spot’ of p53 gene mutations in pterygium, pterygium has the same ‘hot
spot’ as malignant melanoma, breast cancer, and colon cancer.23 Further evaluation of
correlations between potential ‘hot spots’ of pterygium with SCC and BCC are
necessary.
There were two limitations in our study. Firstly, mutational analysis in this study
was confined to exons 4 through 8, instead of sequencing the entire 11 exons of the
p53 gene. We concede that other mutations may occur outside of the field examined,
however, several studies have shown that tumor cells with mutations outside exons 5
contain domains involved in the induction of apoptosis, which may be associated with
tumor behavior.28 Hence, we studied exons 4 to 8 in our series, which, we believe
included nearly all possible relevant mutations in pterygium. Secondly, since we
removed the fibrovascular layer by LCM and then studied the whole epithelium layer,
there was the possibility of false negative due to average of all cells. Hence, the
sequence analysis of all 51 pterygial samples has been rechecked by sense and
antisense sequencing to avoid the false negative, and the sequences of p53 gene were
repeatedly analyzed by three independent experiments. No different results were
found from these analysis.
There were lower signals in figure 2B at positions 339, 343, 344, and 355, which
were not present in the normal DNA of figure 2A. The lower signals in figure 2B may
caused by various factors, including insufficient DNA in the sequence reaction,
degraded template, old or mishandled reagents, incorrect thermal cycling conditions
or electrokinetic injection failure. In this study, DNA for p53 mutation analysis was
extracted from paraffin-embedded pterygium tissues, base on our knowledge, it is
very difficult to extract high quality DNA from paraffin sections. Therefore, we
consider that the lower signals in figure 2B at positions 339, 343, 344, and 355 were
caused by insufficient DNA.
that p53 gene inactivation was predominately through p53 mutations and caused
pterygium formation.3, 6 However, some reports, such as Shimmura et al, Onur et al
and Reisman et al do not support the above conclusion, because few p53 mutations
were detected in pterygia tissues.8, 13, 14 They suggested that the inactivation of p53
was not through gene mutation, but rather through repressed expression of the gene.13
Our results indicated that only 15.7% (8 of 51) had p53 mutations. As p53 mutation
did not occur in all pterygium patients, p53 mutation was not the only cause of
pterygium formation. We suggest that besides p53 gene mutations, there may be other
mechanisms leading to loss of p53 function involved. Silencing of the p53 gene by
alterations or mutations in its regulatory regions, methylation of important regions of
the gene, or defects on other controlling factors required for p53 gene expression may
play a role.13 Moreover, there may be other tumor suppressor genes or oncogenes
involved in pterygium formation.
In conclusion, p53 gene mutations were detected in some pterygial epithelium.
All mutations were single base substitutions and could be found in both p53
immunoreactive and non-reactive groups. This is the first paper to investigate the
Table1. Mutation spectrum in exons 4 to 8 of p53 gene
exon mutation
n (%) spectrum effect
4 1 (12.5%) codon 117 1bp deletion frameshift
5 1 (12.5%) codon 179 C:G→T:A CAT(His)→TAT(Tyr)
6 3 (37.5%) codon 208 A:T→T:A GAC(Asp)→GTC(Val)
codon 213 G:C→C:G CGA(Arg)→CCA(Pro)
codon 213 G:C→T:A CGA(Arg)→CTA(Leu)
7 2 (25%) codon 234 T:A→A:T TAC(Tyr)→AAC(Asn)
codon 259 1bp deletion frameshift
8 1 (12.5%) codon 286 G:C→C:G GAA(Glu)→CAA(Gln)
Table 2. Mutation spectrum and immunohistochemical analysis of p53 in the 51 pterygia immunohistochemistry score n p53 gene mutation n mutation spectrum
0 31 2 exon 7 codon 259 1bp deletion
exon 4 codon 117 1bp deletion
+ 9 2 exon 8 codon 286 G:C→C:G
exon 5 codon 179 C:G→T:A
++ 5 2 exon 7 codon 234 T:A→A:T
exon 6 codon 208 A:T→T:A
+++ 6 2 exon 6 codon 213 G:C→C:G
Figure 1
Fig. 1. Representative results of laser capture micro-dissection (LCM) of pterygium.
(A) and (C) illustrate a specimen before micro-dissection. The epithelium and
sub-epithelial fibrovascular layer are shown. (B) and (D) illustrate a specimen after
Figure 2
Fig. 2. Representative result from p53 DNA sequencing. The mutation at codon 208
of the patient is indicated by an arrow (B), and the normal DNA sequence is shown in
References
1. Nigro JM, Baker SJ, Preisinger AC, et al. Mutations in the p53 gene occur in
diverse human tumor types. Nature 1989; 342:705-708.
2. Tan DTH, Lim AS, Goh HS, Smith DR. Abnormal expression of the p53 tumor
suppressor gene in the conjunctiva of patients with pterygium. Am J Ophthalmol
1997; 123:404-405.
3. Dushku N, Reid TW. P53 expression in altered limbal basal cells of pingula,
pterygia, and limbal tumors. Curr Eye Res 1997; 16:1179-1192.
4. Weinstein O, Rosenthal G, Zirkin H, et al. Overexpression of p53 tumor
suppressor gene in pterygia. Eye 2002; 16:619-621.
5. Tan DTH, Tang WY, Liu YP, et al. Apoptosis and apoptosis related gene
expression in normal conjunctiva and pterygium. Br J Ophthalmol 2000;
84:212-216.
6. Dushku N, Hatcher SLS, Albert DM, Reid TW. P53 expression and relation to
human papillomavirus infection in pinguecula, pterygia, and limbal tumors. Arch
Ophthalmol 1999; 117:1593-1599.
7. Ueda Y, Kanazawa S, Kitaoka T, et al. Immunohistochemical study of p53, p21
and PCNA in pterygium. Acta Histochem 2001; 103:159-65.
Ophthalmol 1998; 8:157-161.
9. Chowers I, Pe’er J, Zamir E, et al. Proliferative activity and p53 expression in
primary and recurrent pterygia. Ophthalmology 2001; 108:985-988.
10. Skilling JS, Sood A, Niemann T, et al. An abundance of p53 null mutations in
ovarian carcinoma. Oncogene 1996; 13:117-123.
11. Wang YC, Chen CY, Chen SK, et al. High frequency of deletion mutations in
p53 gene from squamous lung cancer patients in Taiwan. Cancer Res 1998;
58:328-333.
12. Sjorgen S, Inganas M, Norberg T, et al. The p53 gene in breast cancer: prognostic
value of complementary DNA sequencing versus immunohistochemistry. J Natl
Cancer Inst 1996; 88:173-182.
13. Reisman D, McFadden JW, Lu G. Loss of heterozygosity and p53 expression in
pterygium. Cancer Letters 2004; 206:77-83.
14. Shimmura S, Ishioka M, Hanada K, Shimazaki J, Tsubota K. Telomerase activity
and p53 expression in pterygia. Invest Ophthalmol Vis Sci 2000; 41:1364-1369.
15. Gaillard C and Strauss F. Ethanol precipitation of DNA with linear
polyacrylamide as carrier. Nucleic Acids Research 1990; 18:378.
16. Levine AJ. P53, the cellular gatekeeper for growth and division [review]. Cell
17. Skilling JS, Sood A, Niemann T, et al. An abundance of p53 null mutations in
ovarian carcinoma. Oncogene 1996; 13:117-123.
18. Elledge RM. Assessing p53 status in breast cancer prognosis: where should you
put the thermometer if you think your p53 is sick? J Natl Cancer Inst 1996;
88:141-143.
19. Kastan MB, Onyekwere O, Sidransky D, Vogelstein B, Craig RW. Participation
of p53 protein in cellular response to DNA damage. Cancer Res 1991;
51:6304-11.
20. Beroud C, Soussi T. The UMD-p53 database: New mutations and analysis tools.
Hum Mutat 2003; 21:176-181.
21. Soussi T, Beroud C. Significance of TP 53 mutations in human cancer: A critical
analysis of mutations at CpG dinucleotides. Hum Mutat 2003; 21:192-200.
22. Threlfall TJ, English DR. Sun exposure and pterygium of the eye: a
dose-response curve. Am J Ophthalmol 1999; 128:280-287.
23. Giglia-Mari G, Sarasin A. TP53 mutations in human skin cancers. Hum Mutat
2003; 21:217-228.
24. Wood RC, Mitchell M, Sgouros J, Lindahl T. Human DNA repair genes. Science
2001; 291:1284–1288.
Horikawa T. UV-induced skin damage. Toxicology 2003; 189:21-39.
26. Kau HC, Tsai CC, Hsu WM, Liu JH, Wei YH. Genetic polymorphism of hOGG1
and risk of pterygium in Chinese. Eye 2004; 18:635-639.
27. May P, May E. Twenty years of p53 research: structural and functional aspects of the p53 protein. Oncogene 1999; 18:7621-7636.
28. Zhu J, Zhou W, Jiang J, Chen X. Identification of a novel p53 functional domain that is necessary for mediating apoptosis. J Biol Chem 1998; 273:13030-13036.