國立交通大學
生物科技學系
碩士論文
探討蛋白質四級結構對於細胞質亞硫酸基轉移酵素
之催化活性與功能的影響
The Effect of Quaternary Structure on the Activities and
Functions of Cytosolic Sulfotransferases
研究生:江漢平
指導教授:楊裕雄 教授
探討蛋白質四級結構對於細胞質亞硫酸基轉移酵素之催化
活性與功能的影響
The Effect of Quaternary Structure on the Activities and
Functions of Cytosolic Sulfotransferases
研 究 生:江漢平 Student: Han-Ping Chiang
指導教授:楊裕雄 教授 Advisor: Prof. Yuh-Shyong Yang
國立交通大學
生物科技學系
碩 士 論 文
A Thesis
Submitted to Department of Biological Science and Technology
National Chiao Tung University
in Partial Fulfillment of the Requirements
for the Degree of
Master of Science
in
Biologic Science and Technology
May 2007
Hsinchu, Taiwan, Republic of China
探討蛋白質四級結構對於細胞質亞硫酸基轉移酵素之催化活性與
功能的影響
學生: 江漢平
指導教授: 楊裕雄 教授
國立交通大學生物科技學系碩士班
摘要
亞硫酸基轉移酵素目前大部分已知為同型二聚體,在生理上負責將生
物分子亞硫酸化,藉此調控許多重要的生理反應。本論文主要在探討四級
結構對此酵素活性的影響,因此,藉由膠體過濾分析實驗的證實,我們成
功地製備人類酚亞硫酸基轉移酵素及脫氫表雄甾酮亞硫酸基轉移酵素之同
型二聚體及單體。此外,利用抗生素做大腸桿菌的菌株篩選及連續的親和
性管柱純化策略,我們得到了同時具備酚亞硫酸基轉移酵素及脫氫表雄甾
酮亞硫酸基轉移酵素之催化功能的異型二聚體。經由圓二色偏光光譜法來
觀察各式聚合體於二級及三級結構上的差異,並經由酵素動力分析、受質
抑制反應、與受質酵素結合能力的探討來幫助我們推論亞硫酸基轉移酵素
之催化機制。此外,熱穩定性、酸鹼值耐受度實驗的分析,能幫助我們研
究同型二聚體、單體、與異型二聚體在結構上的穩定性。經由對於同型二
聚體、單體、與異型二聚體之各種特性的分析,我們能瞭解四級結構對於
細胞質亞硫酸基轉移酵素的影響與重要性,更甚之,幫助我們推論藉由多
體聚合化產生具有新功能蛋白質的可能性。
The Effect of Quaternary Structure on the Activities and Functions of
Cytosolic Sulfotransferases
Student: Han-Ping Chiang Advisor: Prof. Yuh-Shyong Yang Department of Biological Science and Technology
National Chiao Tung University, Hsinchu, Taiwan, ROC
ABSTRACT
Sulfotransferase (SULT), mostly known as a homodimer in solution, belongs to an enzyme family that is responsible for the sulfonation of biomolecules which regulates many biological functions. In order to study the effects of quaternary structure on the activity of the enzyme, we prepared homodimer and monomer of SULT1A1 and SULT2A1 by site-directed mutagenesis and confirmed by size-exclusion liquid chromatography. Moreover, the bifunctional enzyme, SULT1A1/SULT2A1 heterodimer, was obtained from Escherichia coli by antibiotic selection and serially purified by affinity fusion tag. The secondary and tertiary structures of SULT in various combinations were investigated by circular dichroism spectrum. Enzyme kinetics, substrate inhibition and substrate binding affinity were also performed to deduce the SULT catalytic mechanism and actions. Thermal inactivation, pH tolerance, and conformational stabilities were determined by enzyme assay analysis. The above analyses toward homodimer, monomer, and heterodimer of SULT1A1 and SULT2A1, facilitate us to understand the importance of quaternary structure to cytosolic sulfotransferases, furthermore, assist us to understand how multimerization in proteins may produce novel biofunction.
Acknowledgement
從專題生到碩士班畢業,在 LEPE 也待四個年頭了 感謝楊裕雄老師的指導與栽培,讓我具備獨立思考、進行實驗的能力;更提供了一個非 常適合做研究的環境,讓我能充分的發揮所長,實現實驗的想法 這一路上受了很多學長姊的照顧與提攜 溫柔的靜玟學姊引領進門,帶我認識實驗室的大大小小 愛搞笑的陳蘋果傳授生化的知識與純化的技術,還幫我取了"江大哥"這名不符實的外號 運動全能的奇翰學長給了我很多中肯且受用的意見,也讓我看到了身為學長該有的擔當 良師兼益友的陸宜,因為你的指導與幫助,才讓我能順利拿到碩士學位;因為你傳授的 把妹技巧,才讓我順利追到中曦 擁有堅強實力的政哲學長,私底下是個很有趣的人,跟你做實驗很快樂,謝謝你不吝惜 敎授科學知識和生活智慧 同學們的互相打氣幫忙,是遇到挫折與挑戰時最好的定心丸 愛跳舞的小丸,總在我有難的時候伸出援手,雖然我很愛虧妳,但心裡其實真的很感謝 妳 愛笑的韋汝,雖然 10 次找妳有 9 次是跟妳要錢,但是每次看到妳笑心情都會很好 臉香香的美春,每次跟妳討論實驗都會有很大的收穫,如果妳能臉香香的話,一定到哪 裡都是搶破頭 學弟妹們的加入,讓我擺脫了LEPE 第(唯)一學弟的美稱 超愛吃的小胖,也是超熱心的小胖,你是最有資格繼承"好人"名號的男人,你讓我的 loading 減輕很多很多,但是我不希望看到你的 weighting 變多很多 一點都不宅的家煒,是個貼心的學弟,讓常飢腸轆轆的我們有一個美味的便當;是個無羽球團(來姐、嘉蔚、小丸、美春、Pupu、Peggy、win、大晃、Rich、朝陽、小胖和家煒) 和出遊團(嘉蔚、陸宜、大晃)是我排解研究壓力的良伴,希望將來還有機會能一起打羽 球,一起出去玩 要感謝的人真的很多,除了要謝天,我也要將這份感謝化為力量,來幫助需要這份力量 的人,謝謝大家 最後也是最重要的,我要感謝一直支持我的家人,你們是我能堅持下去的原動力,也是 我的避風港,爸爸、媽媽、逸平、大阿姨,還有中曦,你們都是我最重要的人,這份畢 業的榮耀希望能與你們一起分享 漢平 2007 / 7
CONTENTS PAGE
Abstract (Chinese) ...i
Abstract (English)...ii
Acknowledgement ...iii
CONTENTS ...v
CONTENTS OF TABLE AND FIGURES………. ...vi
Abbreviation and Symbol...viii
Introduction ...1
Experimental Procedures...4
Results and Discussion ... 11
Conclusion...17
CONTENTS OF TABLE AND FIGURES
PAGE
Table I Purification table of wild type and monomer mutant of SULT1A1 amd
SULT2A1... 21 Table II Calibration curve determination of molecular weight on Sephacryl S-200
HR... 22 Table III Physical and chemical parameters of wild type and monomer mutant of
SULT2A1... 23 Table IV Purification table of SULT1A1/SULT2A1 (heterodimer)……… 24 Table V Kinetic constants of pNPS with wild type and V270E of
SULT1A1……….. 25 Table VI Kinetic constants of PAPS and DHEA with wild type and V260E of
SULT2A1... 26 Table VII Kinetic constants of pNPS and DHEA with SULT1A1/SULT2A1
heterodimer... 27 Figure 1. Scheme for the determination of SULT1A1 activity……… 28 Figure 2. Scheme for the determination of SULT2A1 activity……… 29 Figure 3. The KTVE motif : sequence alignment of cytosolic sulfotransferases………. 30 Figure 4. SDS-PAGE for determination of wild type and monomer mutant of
Figure 5. Calibration curve of Sephacryl S-200 HR... 32 Figure 6. Superimpose at dimerization interface of SULT1A1 and SULT2A1………... 33 Figure 7. SDS-PAGE for determination of SULT1A1/SULT2A1 heterodimer………... 34 Figure 8. Circular dichroism specrtra of wild-type and V260E mutant of SULT2A1…. 35 Figure 9. pH profile of wild type and monomer mutant of SULT1A1………. 37 Figure 10. Thermal stability of wild type and monomer mutant of SULT1A1 and
Abbreviation and Symbol
Full name ε Absorption (exitinction) coefficient
A280 Absorption at 280 nm
A600 Absorption at 600 nm
ADT Androsterone
bis-tris propane 1, 3-bis [tris (hydroxymethyl) methylamino] propane
CD Circular dichroism
DHEA Dehydroepiandrosterone
EDC Ethyl (dimethyl aminopropyl) carbodiimide
kcat Turnover number
kDa Kilodalton
Ki Inhibitory constant of substrate inhibition
Km Michaelis constant
MALDI-TOF Matrix-assisted laser desorption inonzation time-of-flight MES 2-[N-morpholino]ethanesulfonic acid
MU 4-methylumbilliferone
MUS 4-methylumbilliferyl sulfate
PAGE Polyacrylamide gel electrophoresis
PAP Adenosine 3', 5'-diphosphate
PAPS 3'-phosphoadenosine 5'-phosphosulfate
pNP 4-nitrophenol
pNPS 4-nitrophenyl sulfate
RP-HPLC Reversed phase high performance liguid chromatography
SDS Sodium dodecyl sulfate
SULT Sulfotransferase
Vmax Maximum velocity
Introduction
Sulfotransferases (SULTs) are a large family of enzymes that catalyze the transfer of sulfuryl group from the universal donor, 3’-phosphoadenosine 5’-phosphosulfate (PAPS) to numerous endogenous and exogenous compounds [1]. Cytosolic sulfotransferases sulfonate small molecules including steroid hormones, bioamines, chemical carcinogens, therapeutic drugs, and neurotransmitters [2]. Therefore, sulfonation is not only considered to be deactivation and secretion processes of biological signal molecules and xenochemicals, but also be involved in maintaining physiological homeostasis such as dehydroepiandrosterone (DHEA), androsterone (ADT), testosterone, estradiol, and many endogenous steroids [3]. Cytosolic sulfotransferases consist of around 300 amino acid residues, and most of then are present as dimer in solution. It was confirmed previously study performing the cross-linking by ethyl (dimethyl aminopropyl) carbodiimide (EDC) following the separation by C4 reversed phase high performance liquid chromatography (RP-HPLC) column and analyzed by matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF) [4]. It was found that the cross-linked peptides were located between one subunit to the other subunit both nearby the carboxyl terminus, and they also expose to the surface of each subunit. Therefore the mutants V269E and V260E in human SULT1E1 and SULT2A1, respectively, converted the homodimers into monomers. It was further shown that the mutant P269T and
E270V performed simultaneously in mouse SULT1E1, a monomer naturally in solution, forced the formation of the dimer. It was concluded that the general motif KXXXTVXXXE (KTVE motif) is responsible for the dimerization of cytosolic sulfotransferases.
Interestingly, sulfotransferases appear to be capable of forming not only homodimers but also heterodimers. It had been reported the sulfonation activity of
N-hydroxy-2-acetylaminofluorene is associated with three different dimers of
sulfotransferases from rat liver cytosol: rat SULT1A2 homodimer, rat SULT1C1 homodimer, and rat SULT1A2/SULT1C1 heterodimer [5]. However, protein-protein interaction of sulfotransferases has not been well studied and the functional significance of the dimerization process is not known at the present time.
To date, despite the dimerization motif of cytosolic sulfotransferases had been identified [4], the effect of quaternary structure on the catalysis and function of cytosolic sulfotransferases was still far from known and the protein-protein interaction of cytosolic sulfotransferases has not been well studied. The functional significance of the dimerization process is not known at the present time. In this report, we showed that the mutants V270E and V260E of human SULT1A1 and SULT2A1 respectively were sufficient to convert the homodimer to monomer by interfering the hydrophobic KTVE motif and confirmed by size-exclusion chromatography. To investigate the effects of the quaternary structure on the catalysis and function, the studies of the conformational change, catalytic kinetics, substrate
inhibition, substrate binding, thermal and conformational stability are all examined.
In addition to the studies of homodimer and monomer, we established a system to express and purify the heterodimer form Escherichia coli. In this experiment, human SULT1A1 and SULT2A1 were cloned into expression vectors, pET-41a and pET-21b respectively. To take advantages of the different antibiotic genes and the different affinity fusion tags on these two vectors, we obtained the bifunctional enzyme, SULT1A1/SULT2A1 heterodimer, by performed double selections in clone and purification stages. Furthermore the differences between homodimer, monomer, and heterodimer of SULT1A1 and SULT2A1 are clarified and this may be a model to study the effect of quaternary structure among cytosolic sulfotransferases.
Experimental Procedures
Materials. 4-methylumbelliferyl sulfate (MUS), 4-methylumbelliferone (MU),
dehydroepiandrosterone (DHEA), Tris, 4-Nitrophenyl sulfate (pNPS), 3’-phosphoadenosine 5’-phoshpate (PAP), 3’-phosphoadenosine 5’-phosphosulfate (PAPS), β-Naphthol, 2-mercaptoethanol, phenylmethylsulfonylfluoride (PMSF), and bis-tris propane, were purchased from Sigma (St. Louis MO, USA). Sodium dodecyl sulfate (SDS) was obtained from J.T. Baker (Phillipsburg, NJ, USA). PfuTurbo DNA polymerase and DpnI restriction enzyme were purchased from Stratagene (La Jolla, CA). Glutathione S-transferase Sepharose fast flow, and HisTrap affinity column was obtained from Amersham-Pharmacia Biotech Asia Pacific (Hong Kong). All other chemicals were of the highest purity commercially available.
Site-Directed Mutagenesis. The QuickChang site-directed mutagenesis kit from Stratagene
was used for the generation of Val270→Glu270 (V270E)and Val260→Glu260 (V260E) mutations to convert homodimers SULT1A1 and SULT2A1 respectively, into monomers. Wild-type SULT1A1 and SULT2A1 cDNAs incorporated in the pET-41a and pET-21b expression vector respectively, was used as the template in conjunction with specific mutagenic primers. The V270E sense mutagenic oligonucleotide primer sets was 5’-caccttcaccgtggcgcagaatg-3’ for SULT1A1; V260E sense mutagenic oligonucleotide primer sets was 5’-tcacttcacagaggcccaagctg-3’ for SULT2A1. The amplification conditions were 35 cycles of
1 min at 95 °C, 1 min at 55 °C, and 7 min at 68 °C. The reaction contained 100 ng of DNA template, 0.4 μM of sense and antisense primers, 200 μM of each of the four deoxynucleotide triphosphates (dNTPs), 1×PfuTurbo buffer, and 2.5 units of PfuTurbo DNA polymerase. The PCR products were treated with DpnI restriction enzyme for 1 hour at 37 °C followed by transforming into DH5α. Primers for mutagenesis were purchased from Mission Biotech, and mutated cDNA sequences were confirmed by the same company.
Preparation of Wild-type and Monomer Mutant of SULT1A1 and SULT2A1. The recombinant
SULT1A1 was cloned into an expression vector, pET-41a, and transformed into Escherichia
coli BL21 (DE3) [6] were grown in 0.5 liter of LB medium supplemented with 50 μg/ml
kanamycin. When growth to A600 nm = 0.8, induction with 1 mM isopropyl
β-D-thiogalactopyranoside overnight at 25 ℃, the cells were collected by centrifugation and
homogenized in an ice-cold lysis buffer containing 20 mM Tris, pH 8.0, 150 mM NaCl, 125 mM sucrose, and 10% glycerol using a Fisher Scientific Sonic Dismenbrator. The supernatant collected was fractionated by the glutathione sepharose. The glutathione S-transferase fusion protein was eluted by 15 mM glutathione contained in lysis buffer. Concentrated and excluded glutathione of aliquot protein and then treated with 20 units of bovine thrombin for 3 hours at 4 . ℃ Reload the treated protein solution and collected the unbind protein.
expression vector, and the antibiotic was ampicillin (50 μg/ml). The cells were collected by centrifugation and homogenized in a lysis buffer. The supernatant was charged onto a HisTrap affinity column, and fractioned by lysis buffer contained 0~500 mM imidazole.
A homogeneous protein was obtained as determined by SDS-polyacrylamide gel electrophoresis [7]. Protein concentration of the homogeneous form of wild type and monomer mutant SULT1A1 and SULT2A1 were estimated on the basis of absorbency at 280 nm (1.52 and 2.38 ml/mg/cm respectively) [8] using a UV/Vis spectrophotometer (Hitachi UV/Vis-3300, Japan).
Preparation of SULT1A1/SULT2A1 Heterodimer. The recombinant SULT1A1 and SULT2A1
which cloned into pET-41a and pET-21b respectively, were transformed into Escherichia coli BL21 simultaneously, and grown in 0.5 liter of LB medium supplemented with 50 μg/ml ampicillin and 50 μg/ml kanamycin. When growth to A600 nm = 0.8, induction with 1 mM
isopropyl β-D-thiogalactopyranoside overnight at 25 ℃. The cells were collected by
centrifugation and homogenized in a lysis buffer. First, the aliquot protein was fractionated by the glutathione sepharose, and was eluded by 15 mM glutathion contained in lysis buffer. Concentrated and excluded glutathione of aliquot protein. Second, aliquot protein was separated to two parts by HisTrap affinity column. The elution buffers were lysis buffer contained 50 mM and 500 mM imidazole respectively. Collected the solution which eluted by
lysis buffer contained 500 mM imidazole, and then the aliquot protein was concentrated and excluded imidazole by lysis buffer.
A SULT1A1/SULT2A1 heterodimer was obtained as determined by SDS-polyacrylamide gel electrophoresis. Protein concentration of the heterodimer was estimated on the basis of absorbency at 280 nm (1.81 ml/mg/cm).
Size-Exclusion Chromatography and Calibration Curve Preparation. Aliquots of 1ml of
various samples were applied on a Sephacryl S-200 HR column that was equilibrated with 20 mM Tris buffer pH 7.5 containing 150 mM NaCl. Proteins were eluted with the same buffer at a flow rate of 1.0 ml/min. Molecular weights were estimated against calibration standards consisted of ribonuclease A (15.6 kDa), chymotrypsinogen A (19.4 kDa), ovalbumin (47.6 kDa), and albumin (62.9 kDa). Molecular weights of wild type (homodimer) and monomer mutant of SULT1A1, SULT2A1, and SULT1A1/SULT2A1 heterodimer were estimated by gel filtration chromatography [4].
Native-Polyacrylamide Gel Electrophoresis (Native-PAGE). Native-PAGE was
performed according to the Laemmli’s method with 7 % polyacrylamide on separating gel and 5% on the stacking gel. Samples for native-PEGE were mix with a loading buffer (12 mM Tris–HCl, pH 6.8, 5% glycerol (v/v), 2.88 mM of 2-mercaptoethanol, and 0.02% bromphenol
blue (w/v)). Enzyme amount set was 1.18 pmol of each lane.
Circular Dichroism Spectroscopy Measurement. CD measurements were carried out on a
Jasco J715 spectropolarimeter and processed data with J-700 standard analysis version 1.33.00 (Jasco, Japan). CD spectra of enzymes at a concentration of 1 mg/ml in 20 mM phosphate buffer (pH 7.0) were measured in a 1 cm light path length quartz cell in the near-UV range (240-320 nm) and 1 mm light path in the far-UV (200-240 nm) at room temperature.
SULT1A1 and SULT2A1 Enzymatic Assay. The wild type and monomer mutant SULT1A1
assay was carried out via the formation of free pNP (ε = 10500 cm-1 M-1 at pH 7.0). PAPS was
generated first from pNPS and PAP as described previously [8, 9], and then used by the same enzyme for the sulfonation of β-naphthol (Figure 1). The reaction mixture with final volume 1 ml consisted of 100 mM bis-tris propane at pH 7.0, 1 mM pNPS, 50 μM β-naphthol, and 2 μM PAP. Sufficient enzyme was added for detection of an absorption change at 400 nm. The reaction was carried out for 2 min in cuvette with path lengths of 1 cm at 25 °C in a Hitachi UV/Vis-3300 spectrophotometer. Under these conditions, the reaction was linear with respect to time and enzyme concentration. A unit of activity represents the amount of enzyme required to catalyze the formation of 1 μmol of pNP / min in this assay. Specific activity is
given in units per milligram of protein.
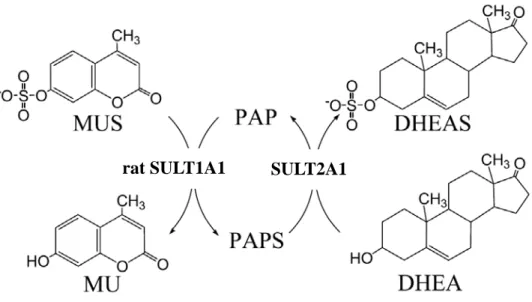
The activities of wild-type and monomer mutant of SULT2A1 were determined according to the change of fluorescence based on a coupled-enzyme assay method [10]. The fluorescence of MU at 460 nm was measured upon excitation at 360 nm. The reaction mixture with a final volume of 1 ml consisted of 100 mM potassium phosphate buffer at pH 7.0, 5 mM 2-mercaptoethanol, 20 μM PAPS, 2 mM MUS, 5.4 μg K65ER68G [11] of rat SULT1A1, SULT2A1 and 5 μMof DHEA at 37 ℃ (Figure 2).A linear response was obtained when 1.49 to 14.9 nM (0.1 to 1 μg) SULT2A1 was added in the standard assay condition [10]. In the reaction condition, regeneration of PAPS catalyzed by K65ER68G of rat SULT1A1 is much faster than the production of PAP catalyzed by SULT2A1 for approximately 40 to 50 folds.
Kinetic Constants Determination. Measurement of the kinetic constants of each substrate was
performed by varying the concentration of one substrate, while keeping the other substrate at a fixed and near saturating concentration. The apparent Km and Vmax were determined using
nonlinear regression by Sigmaplot 2001, V7.0 and Enzyme Kinetics Module, V1.1
pH effects on activity. To examine the pH effects of wild type, monomer mutant and
heterodimer of SULT1A1 and SULT2A1, activities of enzymes were measured described in “Experimental Procedures”, but using the different buffer systems in different pH (100 mM
MES at pH 6 ~ 7, bis-tris propane at pH 7 ~ 9, glycine at pH 9 ~ 10).
Thermal stability Studies. To examine the thermal stability of wild type, monomer mutant and
heterodimer of SULT1A1 and SULT2A1, aliquot of them were preincubated at desired temperatures for 15 minutes and then performed activity measurements as described in “Experimental Procedures”.
Substrate Binding. The dissociation constants (Kd) and stoichiometry of PAP, pNP, and
DHEA towards homodimer, monomer, and heterodimer of SULT1A1 and SULT2A1 were determined with a spectrofluorimeter. The decrease in intrinsic fluorescence of protein at 340 nm was observed upon excitation at 290 nm when an aliquot amount of PAP was added into the mixture consisted of 100 mM potassium phosphate buffer at pH 7.0, 100 nM of aliquot enzyme and varying concentrations of PAP, pNP, and DHEA at 37 ℃ with a final volume of 2 ml in a quartz cuvette of 1 cm square cross-section.
Results and Discussion
Preparation of wild type and monomer mutant of SULT1A1 and SULT2A1. Previous
studies have delineated the dimerization interface of cytosolic sulfotransferase naturally existed in solution, and the individual subunits form the homodimer through the highly conserved sequence KXXXTVXXXE (KTVE motif) [4] near the carboxyl terminus of each subunit are shown in Figure 3. The KTVE motif of each monomer appears to form a zipper-like and anti-parallel interface, and promote the formation of the homodimer. The main feature of the interface can be described as the complementary hydrophobic interaction and backbone hydrogen bonds in the central portion of motif that are reinforced by an ion pair at each end of the motif [4]. For the reason, residue Val270 and Val260 of human SULT1A1 and SULT2A1 respectively were selected and mutated to Glu. Using bacterial expression system, wild type and monomer mutant of SULT1A1 and SULT2A1 were purified through GSTrap and HisTrap affinity columns respectively to get homogeneous enzymes determined by SDS-PAGE in Figure 4. The purification table is shown in TableⅠ. The molecular weights of wild type and monomer mutant of SULT1A1 and SULT2A1 were estimated by gel filtration using Sephacryl S-200 HR. The retention volume (Ve) and the parameter, Kav of the set of molecular weight standards are shown in Table Ⅱ. The retention volumn of the blue dextran 2000 was 38.1 ml and took as V0. Kavs of standards could be estimated by the
equation : Kav = ( Ve – V0 ) / ( Vt – V0 ), and Vt meant the gel filtration column volume. The calibration curve was plotted as the independent variable was log (MW) versus the dependent variable Kav as shown in Figure 5. By means of the calibration curve, the apparent molecular weight of wild type and mutant SULT1A1 and SULT2A1 were estimated as described in
Table Ⅲ. The calculated molecular weights by calibration were similar to the theoretical
molecular weights of wild type and mutant of SULT1A1. Similar result was obtained for SULT2A1 (Table Ⅲ). The apparent molecular weights of wild type SULT1A1 and SULT2A1 were approximately 2-fold than those of V270E and V260E mutants respectively. These data indicated that the interface of dimerizaion was easily disrupted through site-directed mutagenesis to form monomeric SULT1A1 and SULT2A1 because glutamic acid might break hydrophobic zipper-like KTVE motif as predicted in previous studies [4]. Following the purification, the protein yields of wild type were much better than those of monomer mutants of SULT1A1 and SULT2A1, even although they expressed similar total activities in the crude extract (TableⅠ). The low recoveries toward SULT1A1 and SULT2A1 mutants indicated that monomer is unstable under the processes of purification.
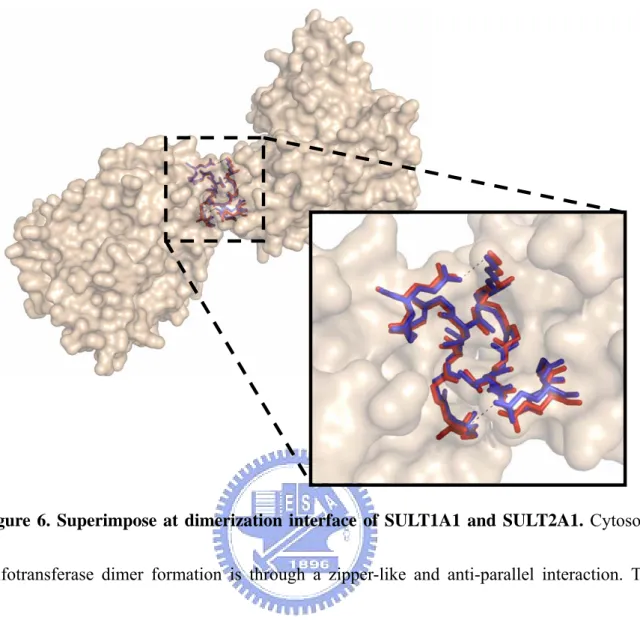
Preparation of SULT1A1/SULT2A1 heterodimer. The sequences of cytosolic sulfotransferase indicate that the KTVE motif is highly conserved among dimeric sulfotransferase (Figure 3). The crystal structures of SULT1E1, SULT1A1, SULT1C2, and
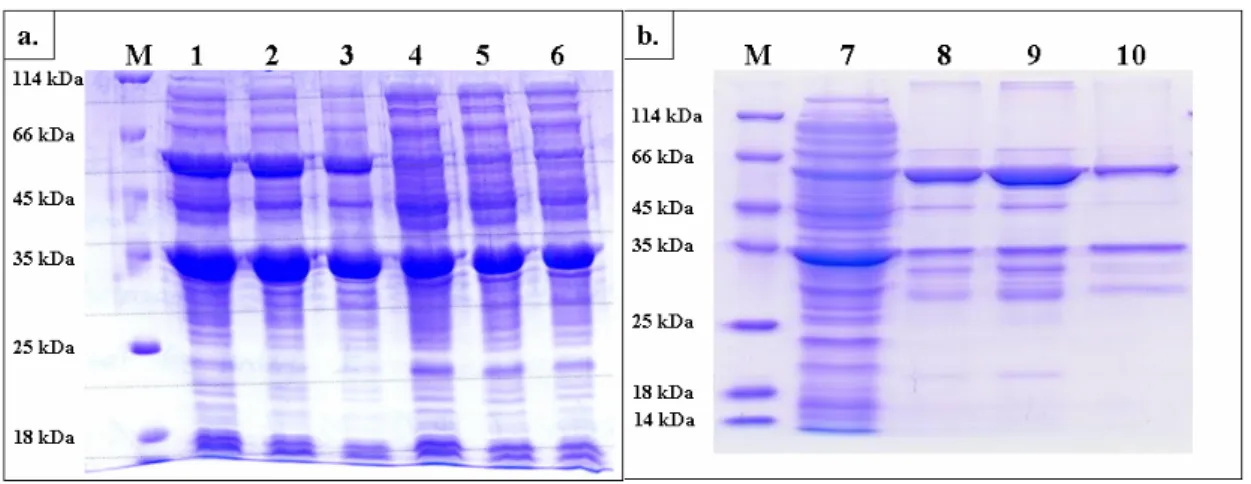
SULT1C3 also confirmed the conservation of dimerization interface [12, 13]. Figure 6 illustrates the very similar structure at dimerization interface by the superimposition of SULT1A1 and SULT2A1. According to these characteristics of cytosolic sulfotransferases, we predict the formation of SULT1A1/SULT2A1 heterodimer during the coexpression of the two recombinant proteins in Escherichia coli BL21 (DE3). The expression vectors of SULT1A1 and SULT2A1 were pET41a and pET21b respectively. To take advantage of the different antibiotic genes, kanamycin and ampicillin, on pET41a and pET21b respectively, we easily selected the colony that contained the two plasmids and could express SULT1A1 and SULT2A1 simultaneously as shown in Figure 7a. Following the expression, there are three possible dimmers, SULT1A1 homodimer, SULT2A1 homodimer, and SULT1A1/SULT2A1 heterodimer. Vector pET41a and pET21b provided two fusion tags, GST (glutathione
S-transferase) and Histag, to SULT1A1 and SULT2A1 respectively, for us to perform the
purification of heterodimer through two affinity columns, GSTrap and Histrap, sequentially to obtain the homogeneous SULT1A1/SULT2A1 heterodimer shown in Figure 7b. The crude extract (Lane 7) was separated by GSTrap at first to separate SULT2A1 homodimer from SULT1A1 homodimer and SULT1A1/SULT2A1 heterodimer (Lane 8), which were further fractionated two portions by HisTrap chromatography, eluted with 50 mM (Lane 9) and 500 mM (Lane 10) imidazole respectively. Table Ⅳ summarizes the activities of SULT1A1 and SULT2A1 obtained in each step of the purification. According to Figure 7 and Table Ⅳ, we
are able to confirm that SULT1A1/SULT2A1 heterodimer through antibiotic and affinity tag double selection was obtained.
Circular dichroism measurement. Analysis by circular dichroism spectrum reveals the differences between wild type and monomer mutant of SULT1A1 and SULT2A1 whether in the secondary or the tertiary structure. The CD spectrum in the range of far-UV (200-240 nm) indicated the unfastened secondary structure of SULT1A1 monomer (Figure 8a), and showed the slight difference between homodimer and monomer in the 205-225 nm of SULT2A1 (Figure 8c), where is the absorption of α-helix and β-sheet. It suggests that the single mutation of SULT1A1 and SULT2A1, caused the change in their secondary structure. Furthermore, the change of tertiary structure of SULT1A1 and SULT2A1 were also monitored by the CD spectrum in the range of near-UV (240-320 nm). The variation at 260-300 nm indicates that environment of aromatic acid in SULT1A1 (Figure 8b) and SULT2A1 (Figure 8d), are different in monomeric and dimeric sulfotransferase. CD data indicates that single mutation of V270E and V260E, caused the structure change at secondary and tertiary level. It may reveal that there is a bit difference between the homodimer and monomer although it may affect the whole protein structure or partial quaternary protein structure located near the KTVE motif.
Kinetic analysis. Kinetic constants of wild type and monomer mutants of SULT1A1 and
SULT2A1 were shown in Table Ⅴand Table Ⅵ, respectively. The apparently Km values of
pNPS were similar for homodiemr and monomer of SULT1A1, but the catalytic rate (kcat) of
dimer was higher than that of monomer at approximately 2 folds. Similar kinetics data were observed in homodimer and monomer of SULT2A1, and further, substrate inhibition was found in both of them. Km and Ki were not apparently different in monomer or homodimer
sulfotransferase. The kinetic constants of pNPS and DHEA with SULT1A1/SULT2A1 heterodimer (Table Ⅶ) are similar to those obtained from a monomer eznymer. These results indicated that both the homodimer and heterodimer might have competent catalytic ability in each subunit in both SULT1A1 and SULT2A1. These results raise an interesting question regarding how many nucleotide cofactors (PAP or PAPS) are needed in sulfotransferase catalyzed reaction. Previous studies indicated that there was only one PAP bound per homodimer for rat SULT1A1 [11], and one PAP or PAPS bound per homodimer for human SULT1E1 [14].
pH profile and thermal stability. Buffers used for a pH-dependence experiment may also
affect activity of sulfotransferase (Figure 9). The optima pH toward wild type and monomer mutant of SULT1A1 was pH 6.5, and they have identical specific activities at pH 7.0, with similar tendencies after at pH 7 ~ 10 (Figure 9a); the optima pH for SULT2A1 was also 6.5,
and wild type and monomer mutant have identical specific activities at pH 7.5 (Figure 9b). Interestingly, the activities of SULT1A1 have acute degradation both of them from pH 6.5 to 6.0, but the wild type still have 38 % remnant and the monomer mutant almost lost its activity was barely 3 %. Thermal inactivation of SULT1A1 and SULT2A1 were studied over the temperature range 25 ~ 50 ℃ after the incubation at each desired temperature for 15 min. The severe decreased activities toward wild type and monomer mutant of SULT1A1 are at the temperature of 45 and 40 ℃, respectively shown in Figure 10a, and there were similar results toward SULT2A1, 40 and 35 ℃, respectively in Figure 10b. The results of pH and thermal stability reveal the monomer was more unstable than homodimer both of SULT1A1 and SULT2A1 while the enzymatic activities drop off severely in the rigorous conditions. It might be attributed to the effects of quaternary structure but it is not clear to know that whether the protein-protein interaction forces the homodimer more stable than monomer.
Conclusion
We obtained the monomers of SULT1A1 and SULT2A1 by site-directed mutagenesis to disrupt the KTVE motif and confirmed by gel filtration. Preliminary results showed no significant different in catalytic properties between homodimer and monomer of SULT1A1 and SULT2A1. That is, each enzyme unit is fully active in ether monomeric of dimeric form. However, the current study found that each enzyme unit also binds to one PAP, in contract to previous reports [11, 14]. These results indicate that other factors such as redox [15, 16] and α, β isoform [11, 16] of sulfotransferase may play a role in the cofactor binding. The pH tolerance and thermal inactivation indicate that monomer is more labile than homodimer, and it may be the result of protein-protein interaction. To investigate the structure diversity that may influence the ligand binding between homodimer and monomer, crystallization of the enzymes in different form will by proceeded in the further.
Furthermore, the SULT1A1/SULT2A1 heterodimer was prepared by double-selection of antibiotic and affinity tag. We will continue to investigate the heterodimeric effect of sulfotransferase and try to understand the importance in quaternary structure for cytosolic sulfotransferases.
References
1. W.B. Jakoby, and D.M. Zigeler, The enzymes of detoxication. J. Biol. Chem. 265 (1990), 20715-20718.
2. C.N. Falany, Sulfation and sulfotransferases. Introduction: changing view of sulfation and the cytosolic sulfotransferases. FASEB J. 11 (1997), 1-2.
3. C.N. Falany, Enzymology of human cytosolic sulfotransferases. FASEB J. 11 (1997), 206-216.
4. E.V. Petrotchenko, L.C. Pedersen, C.H. Borchers, K.B. Tomer, and M. Negishi, The dimerization motif of cytosolic sulfotransferases. FEBS Lett. 490 (2001), 39-43.
5. C.C. Kiehlbauch. Y.F. Lam, D.P. Ringer, Homodimeric and heterodimeric aryl sulfotransferases catalyze the sulfuric acid esterification of
N-hydroxy-2-acetylaminofluorene. J. Biol. Chem. 270 (1995), 18941-18947.
6. X. Chen, Y.S. Yang, Y. Zheng, B.M. Martin, M.W. DuVel, W.B. Jakoby, Tyrosine-ester sulfotransferase from rat liver: bacterial expression and identiWcation, Protein Expr.
Purif. 3 (1992) 421-426.
7. U.K. Laemmli, Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227 (1970) 680-685.
8. S.C. Gill, P.H. von Hippel, Calculation of protein extinction coeYcients from amino acid sequence data, Anal. Biochem. 182 (1989) 319-326.
9. E.S. Lin, and Y.S. Yang, Nucleotide binding and sulfation catalyzed by phenol sulfotransferase, Biochem. Biophys. Res. Commun. 271 (2000) 818-822.
10. W.T. Chen, M.C. Liu, Y.S. Yang, Fluorometric assay for alcohol sulfotransferase. Anal.
Biochem. 339 (2005), 54-60.
11. Y.S. Yang, A.D. Marshall, P. McPhie, W.X. Guo, X. Xie, X. Chen, W.B. Jakoby, Two phenol sulfotransferase species from one cDNA: nature of the differences. Protein Expr.
Purif. 8 (1996), 423-429.
12. S. Shevtsov, E.V. Petrotchenko, L.C. Pedersen, M. Negishi, Crystallographic analysis of a hydroxylated polychlorinated biphenyl (OH-PCB) bound to the catalytic estrogen binding site of human estrogen sulfotransferase Environ. Health Perspect. 111 (2003), 884-888.
13. N.U. Gamage, S. Tsvetanov, R.G. Duggleby, M.E. McManus, J.L. Martin, The structure of human SULT1A1 crystallized with estradiol. An insight into active site plasticity and substrate inhibition with multi-ring substrates J. Biol. Chem. 280 (2005), 41482-41486. 14. H. Zhang, O. Varlamova, F.M. Vargas, C.N. Falany, T.S. Leyh, Sulfuryl transfer: the
catalytic mechanism of human estrogen sulfotransferase. J. Biol. Chem. 273 (1998), 10888–10892.
15. A.D. Marshall, J.F. Darbyshire, A.P. Hunter, P. McPhie, W.B. Jakoby, Contol of activity through oxidative modification at the conserved residue Cys66 of aryl sulfotransferase Ⅳ.
J. Biol. Chem.. 272 (1997), 9153-9160.
16. T.M. Su, Y.S. Yang, Mechanism of posttranslational regulation of phenol sulfotransferase : expression to two enzyme forms through redox modification and nucleotide binding. Biochemistry 42 (2003), 6863-6870.
17. K. Nagata, Y. Yamazoe, Pharmacegenetics of sulfotransferase. Annu. Rev. Pharmacol.
a Total activity represented the amounts of products catalyzed by enzymes of each step per minute. The reaction conditions were described in
“Experimental Procedures”.
Total Activity a Total Protein Sepcific Activity Yield nmol / min mg nmol / min / mg %
Crude Extract 41143 1129.8 41 100 1 Homogeneus Protein 10012 11.3 887 24.3 22 Crude Extract 54857 976.2 56 100 1 Homogeneus Protein 1905 2.6 719 3.5 13 Crude Extract 1432 695.7 2 100 1 Homogeneus Protein 621 11.2 56 43 23 Crude Extract 756 537.4 1 100 1 Homogeneus Protein 100 1.8 56 13.2 56 SULT2A1 wild type V270E wild type V260E Step Fold Enzyme SULT1A1
Purification table of wild type and monomer mutant of SULT1A1 amd SULT2A1 Table Ⅰ
Table Ⅱ
Calibration curve determination of molecular weight on Sephacryl S-200 HR
MW log (MW) Ve a Kav b
Albumin 67000 4.83 51.77 0.17
Ovabumin 43000 4.63 57.26 0.23
Chymotrypsinofen A 25000 4.40 72.83 0.42
Ribonuclease A 13700 4.14 81.14 0.52
a Ve represented the retention volume of eluted protein under the condition as indicated under
“Experimental Procedures”.
b Kav was calculated by the equation: Kav = (Ve – V0) / (Vt – V0). The Vt was the column
volume as 120 ml and the V0 was the retention volume of blue dextran 2000 (2000 kDa), whose retention volume was 38.1 ml.
c The molecular weights were determined by Sephacryl S-200 HR according to the calibration
curve described above.
b The Ve indicated the retention volume as described above and determined by Sephacryl
S-200 HR.
a The molecular weights were determined by the ProtParam on the EXPASY server
(http://tw.expasy.org/tools/protparam.html).
Physical and chemical parameters of wild type and monomer mutant of SULT2A1.
Molecular weight Ⅰa Ve b Molecular weight Ⅱ c
Wild type 68331 52.53 63093 V270E 34195 62.69 37350 Wild type 68994 52.41 63485 V260E 34541 63.33 36137 SULT1A1 SULT2A1 Table Ⅲ
b The steps of GSTrap and HisTrap represented the protein was separated by these two affinity column as described in “Experimental Procedure”.
a Heterodimer is composed of SULT1A1 and SULT2A1. In the procedure of purification, we recorded the activities simultaneously performed by
SULT1A1 and SULT2A1 respectively.
Total Activity Total Protein Sepcific Activity Yield
nmol / min mg nmol / min / mg %
Crude extract
17524
1680
9
100
1
GSTrap b1743
4.1
421
9.95
47
Histrap b15
0.1
223
0.09
25
Crude extract3520
1680
2
100
1
GSTrap b27
4.1
7
0.78
4
Histrap b2
0.1
24
0.05
12
Enzyme SULT1A1 a SULT2A1 a Step FoldPurification table of SULT1A1/SULT2A1 (heterodimer) Table Ⅳ
Table Ⅴ
Kinetic constants of pNPS with wild type and V270E of SULT1A1
kcat Km kcat / Km kcat Km kcat / Km
(min-1) (nM) (min-1) (mM)
Wild type 132 ± 4 25.6 ± 3.5 5.2 402 ± 9 6.5 ± 0.3 61.8
V270E 82 ± 2 23.8 ± 2.9 3.4 199 ± 11 5.9 ± 0.8 33.7
PAP a p NPS b
a The reaction mixture with a final volume of 1 ml consisted of 100 mM bis-tris propane at pH
7.0, 50 μM β-naphthol, 2.5 mM pNPS, and PAP (0.005 ~ 1 μM), plus 2.5 μg of enzyme (wild type or V270E mutant of SULT1A1) at 25 ℃. The kinetic constants were obtained using nonlinear regression by SigmaPlot 2001, V7.0 and Enzyme Kinetics Module, V1.1 (SPSS Inc., Chicago, IL).
b The reaction mixture with a final volume of 1 ml consisted of 100 mM bis-tris propane at
pH 7.0, 50 μM β-naphthol, 2 μM PAP, and pNPS (0.05 ~ 20 mM), plus 2.5 μg of enzyme (wild type or V270E mutant of SULT1A1) at 25 ℃. The kinetic constants were obtained using nonlinear regression by SigmaPlot 2001, V7.0 and Enzyme Kinetics Module, V1.1 (SPSS Inc., Chicago, IL).
Table Ⅵ
Kinetic constants of PAPS and DHEA with wild type and V260E of SULT2A1
kcat Km kcat / Km kcat Km Ki kcat / Km
(min-1) (μ M) (min-1) (μ M) (μ M)
Wild type 8.6 ± 0.8 3.9 ± 0.4 2.2 8.2 ± 0.7 4.7 ± 0.3 4.3 ± 0.3 1.7 V260E 4.4 ± 0.7 4.2 ± 0.6 1.1 4.2 ± 0.5 5.6 ± 0.5 4.9 ± 0.5 0.8
PAPS a DHEA b
a The reaction mixture with a final volume of 1 ml consisted of 100 mM potassium phosphate
buffer at pH 7.0, 5 mM 2-mercaptoethanol, 5 μM DHEA, 2 mM MUS, 5.4 μg K65ER68G of rat SULT1A1, and PAPS (0.5 ~ 20 μM), plus 1 μg of enzyme (wild type or V260E mutant of SULT2A1) at 37 ℃. The kinetic constants were obtained using nonlinear regression by SigmaPlot 2001, V7.0 and Enzyme Kinetics Module, V1.1 (SPSS Inc., Chicago, IL).
b The reaction mixture with a final volume of 1 ml consisted of 100 mM potassium phosphate
buffer at pH 7.0, 5 mM 2-mercaptoethanol, 20 μM PAPS, 2 mM MUS, 5.4 μg K65ER68G of rat SULT1A1, and DHEA (0.3 ~ 50 μM), plus 1 μg enzyme (wild type or V260E mutant of SULT2A1) at 37 ℃.
Table Ⅶ
Kinetic constants of pNPS and DHEA with SULT1A1/SULT2A1 heterodimer
kcat Km kcat / Km kcat Km Ki kcat / Km
(min-1) (mM) (min-1) (μM) (μM)
192 ± 7 5.3 ± 0.5 36.2 2.4 ± 0.4 1.6 ± 0.5 17.2 ± 6.0 1.5 DHEA b
SULT1A1/SULT2A1
p NPS a
a The reaction mixture with a final volume of 1 ml consisted of 100 mM bis-tris propane at pH
7.0, 50 μM β-naphthol, 2 μM PAP, and pNPS (0.05 ~ 20 mM), plus 2.5 μg of enzyme at 25 ℃. The kinetic constants were obtained using nonlinear regression by SigmaPlot 2001, V7.0 and Enzyme Kinetics Module, V1.1 (SPSS Inc., Chicago, IL).
b The reaction mixture with a final volume of 1 ml consisted of 100 mM potassium phosphate
buffer at pH 7.0, 5 mM 2-mercaptoethanol, 20 μM PAPS, 2 mM MUS, 5.4 μg K65ER68G of rat SULT1A1, and DHEA (0.3 ~ 50 μM), plus 1 μg enzyme at 37 ℃.
SULT1A1 SULT1A1
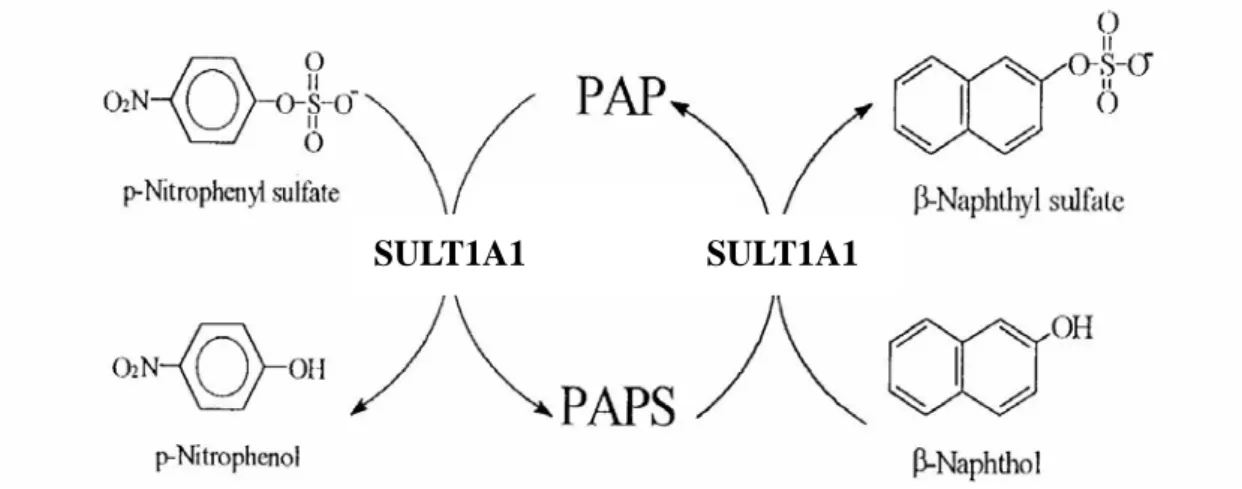
Figure 1. Scheme for the determination of SULT1A1 activity. The assay depends on the
formation of p-nitrophenol (ε = 10500 cm-1 M-1 at pH 7.0) and PAPS from
p-nitrophenulsulfate and PAP. In turn, PAPS is used by SULT1A1 for the sulfurylation of a
second phenol, β-naphthol to achieve optimum rate even at very low concentrations of PAP since the nucleotide is recycled at this coupled reaction.
rat SULT1A1 SULT2A1
Figure 2. Scheme for the determination of SULT2A1 activity. This assay was based on the
regeneration of PAPS from PAP catalyzed by a recombinant rat SULT1A1 (K65ER68G) using MUS as the sulfuryl group donor. In coupled enzyme assay, rat SULT1A1 represented an auxiliary enzyme and the product, MU, was used as a fluorescent indicator of enzyme turnover.
Figure 3. The KTVE motif : sequence alignment of cytosolic sulfotransferases. The
conserved residues, KxxxTVxxxE, are highlighted as red. Sulfotransferases are listed according to the classification used in a review article [17] (data not show). The sequence source was from NCBI database (http://www.ncbi.nlm.nih.gov), and using Weblogo to generate the sequence logos (http://weblogo.berkeley.edu).
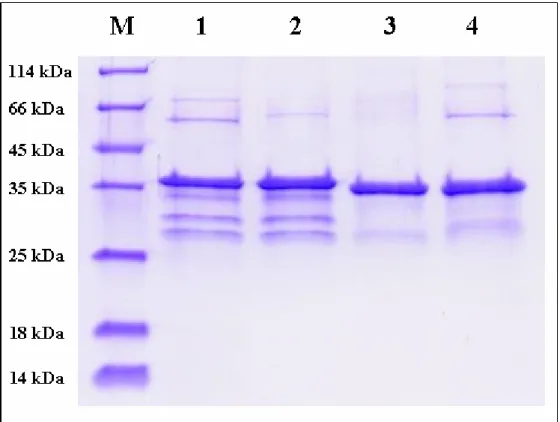
Figure 4. SDS-PAGE for determination of wild type and monomer mutant of SULT1A1
and SULT2A1. Purification of wild type (lane 1) and monomer mutant (lane 2) of
SULT1A1; wild type (lane 3) and monomer mutant (lane 4) of SULT2A1, the molecular weight of SULT1A1 and SULT2A1 were 38 kDa (SULT1A1 plus the remnant segments connect to SULT1A1 after thrombin digestion) and 34 kDa respectively.
log (MW)
4.1 4.2 4.3 4.4 4.5 4.6 4.7 4.8 4.9Ka
v
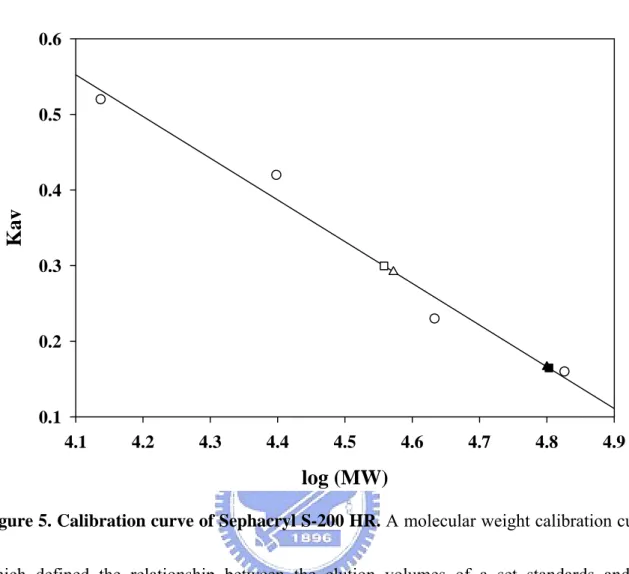
0.1 0.2 0.3 0.4 0.5 0.6Figure 5. Calibration curve of Sephacryl S-200 HR. A molecular weight calibration curve,
which defined the relationship between the elution volumes of a set standards and the logarithm of their respective molecular weights, was determined with Sephacryl S-200 HR. Calibration standards (circle) used were ribonuclease A (15.6 KDa), chymotrypsinogen A (19.4 KDa), ovalbumin (47.6 KDa), and Albumin (62.9 KDa). The wild type and V270E mutant of SULT1A1 were closed and open triangle, respectively; the wild type and V260E mutant of SULT2A1 were closed and open square respectively. The Kav is obtained from the equation Kav = (Ve – V0) / (Vt – V0) as described above.
Figure 6. Superimpose at dimerization interface of SULT1A1 and SULT2A1. Cytosolic
sulfotransferase dimer formation is through a zipper-like and anti-parallel interaction. The structure of SULT1A1 (blue) at this interface is very similar to SULT2A1 (red). The superimposition was constructed using PyMOL base on the two structures SULT1A1 (PDB code : 2D06) and SULT2A1 (PDB code : 1J99).
Figure 7. SDS-PAGE for determination of SULT1A1/SULT2A1 heterodimer. (a)
Coexpression of SULT1A1 (34 kDa + 36 kDa, fusion of glutathione S-transferase) and SULT2A1 (34 kDa). Lane 1, 2, and 3 indicated the incubation time were 30 hr, 16 hr, and 8 hr respectively with 1 mM IPTG at 25 ℃. Lane 4, 5, and 6 indicated the incubation time were 30 hr, 16 hr, and 8 hr respectively without 1 mM IPTG at 25 ℃. (b) Purification steps of SULT1A1/SULT2A1 heterodimer. Lane 7 was crude extract; lane 8 shown the protein was purified by GSTrap affinity column; lane 9 and lane 10 indicated the proteins were separated by HisTrap (Ni 2+) affinity column, and the elution buffer was lysis buffer contained 50 mM and 500 mM imidazole respectively.
[θ ] (10 4 de g c m 2 /dmol) -4 -2 0 2 W a v e le n g th (n m ) 2 4 0 2 6 0 2 8 0 3 0 0 3 2 0 [θ ] (10 4 de g cm 2 / dmol ) -6 -4 -2 0 a W a v e le n g th (n m ) 2 0 0 2 1 0 2 2 0 2 3 0 2 4 0 [θ ] (10 6 de g cm 2 / dmol ) -4 -3 -2 -1 0 1 b c [θ ] (10 6 de g cm 2 /dmol) -4 -3 -2 -1 0 1 d
Figure 8. Circular dichroism specrtra of wild-type and V260E mutant of SULT2A1. CD
measurements were carried out on a Jasco J715 spectropolarimeter and processed data with J-700 standard analysis version 1.33.00 (Jasco, Japan). CD spectra of enzymes at a concentration of 1 mg / ml in the phosphate buffer (pH 7.0) were measured in a (a, c) 1 mm light path length quartz cell in the far-UV range (200-240 nm) and (b, d) 1 cm light path in the near-UV (240-320 nm) at room temperature individually. The closed and open diamonds indicated the wild type and monomer mutant of SULT1A1 (a, b) and SULT2A1 (c, d), respectively.
pH 5 6 7 8 9 10 11 Specif ic act ivity ( n mol/min/mg) 0 1000 2000 3000 4000 a
pH
5 6 7 8 9 10 b 11Sepecific activty (nmol/min/mg)
020 40 60 80 100
Figure 9. pH profile of wild type and monomer mutant of SULT1A1 and SULT2A1. (a)
The enzymatic assay for SULT1A1 was carried out under the condition of 2 μM PAP, 50 μM β-naphthol, and 2.5 mM pNPS at 25 ℃, with different buffer systems, 100 mM MES at pH 6 ~ 7, bis-tris propane at pH 7 ~ 9, and glycine at pH 9 ~ 10. (b) The enzymatic assay for SULT2A1 was carried out under the condition of 20 μM PAPS, 5 mM 2-mercaptoethanol, 5 μM DHEA, 2 mM MUS, 5.4 μg K65ER68G of rat SULT1A1 at 37 ℃, with different buffer systems, 100 mM MES at pH 6 ~ 6.5, potassium phosphate at pH 6.5 ~ 7.5, bis-tris propane at 7.5 ~ 9, and glycine at pH 9 ~ 10. The results of wild type and monomer mutant were closed and open diamond respectively
a Temperature (oC) 25 30 35 40 45 50 Res idu al Activity (%) 0 20 40 60 80 100 Temperature (oC) 25 30 35 40 45 50 Residual Activity (%) 0 20 40 60 80 100 b
Figure 10. Thermal stability of wild type and monomer mutant of SULT1A1 and
SULT2A1. The experiments were performed as described in “Experimental Procedures”. The
results of wild type (closed diamonds) and monomer mutant (open diamonds) of SULT1A1 were shown in (a), and SULT2A1 were (b).