國立交通大學
生物科技學系
博士論文
紅麴菌膽固醇抑制劑 (Monacolin K)、橘黴素及
轉位子相關基因之研究
Characterization of Monacolin K, Citrinin, and Non-LTR
Retrotransposon Related Genes in Monascus pilosus
研 究 生:陳煜沛
指導教授:曾慶平 博士
袁國芳 博士
摘要
紅麴菌可生產各種不同的二次代謝物,包括膽固醇抑制劑(monacolin K),橘 黴素(citrinin)及色素等。紅麴菌所生產的膽固醇抑制劑 monacolin K 與 Aspergillus terreus 產生的 lovastatin 為結構相同的物質。橘黴素則是一種腎肝毒素,亦是針 對格蘭氏陽性菌的抗菌劑,橘黴素除了紅麴菌會產生之外,於 Aspergillus 及 Penicillium 中也有發現。根據前人的研究指出,lovastatin 與 citrinin 基因群已分 別在 A. terreus 及 M. purpureus 真菌中發現。因此為了解紅麴菌 monacolin K 基因 群,我們從細菌人工染色體基因庫(Bacterial Artificial Chromosome library, BAC)
篩選出含有 monacolin K 基因群之 BAC clones,並完成定序及註解分析,
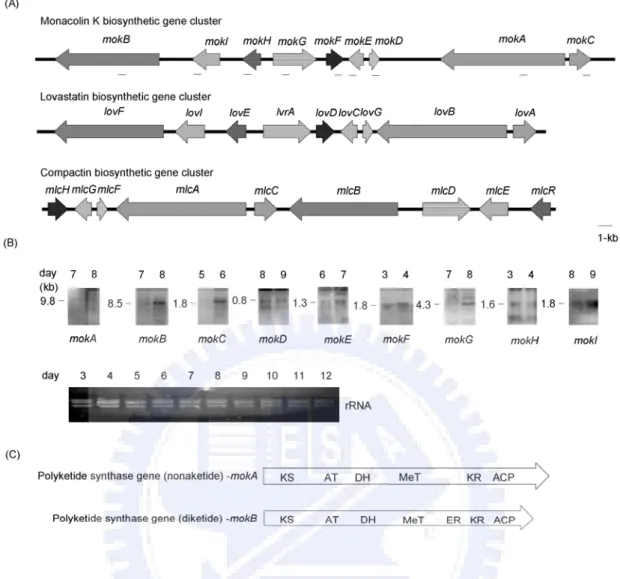
monacolin K 生合成基因群全長為 42 kb 包含 9 個基因,命名為 mokA-mokI,其與
已知 lovastatin 基因群具有 54%的相似度。然而為確認所選殖基因確實參與
monacolin K 的生合成,本研究首先利用核酸酶抑制劑(aurintricarboxylic acid)建 立具方便及效率性之紅麴基因轉殖技術,藉由基因轉殖篩選出 mokA 基因的突變
株,並證實此突變株不會產生monacolin K,進一步於紅麴菌中表現 mokH 基因,
確認轉型株中含有兩套 mokH 基因,分析轉型株的 monacolin K 產量及 RNA 表
現,發現其表現量皆高於野生株,因此結果明確說明所篩選的基因參與monacolin
K 的生合成。此外,選殖出 mokA 基因,並與 4’-phosphopantetheine transferase (PPTase) 於 Escherichia coli 中共同表現,發現 E. coli 中大量表現出色胺酸 (tryptophan)的前驅物 anthranilic acid,此結果顯示 mokA 基因於 E. coli 中表現出 的產物視同誘發物(elicitor),可調控誘導 anthranilic acid 在 E. coli 中表現。
再者,本研究於紅麴菌的細菌人工染色體基因庫定序中發現新的 non-LTR
逆轉位子,並命名為MRT,其序列長度約為 5.5 kb,包含有兩個開放讀架(ORFs)。
此兩個開放讀架與 gag 及 pol 基因相似,並且於 pol 相似基因的 3’端具有很多的
腺嘌呤。第一個開放讀架為517 個氨基酸的蛋白質,包含有多量半胱氨酸的鋅指
切酶、反轉錄酶、核糖核酸酶H 及 CCHC 區域。根據 MRT non-LTR 逆轉位子的
氨基酸序列所建立的親緣關係,可將其歸屬於 Tad1 群叢。南方雜交法則進一步
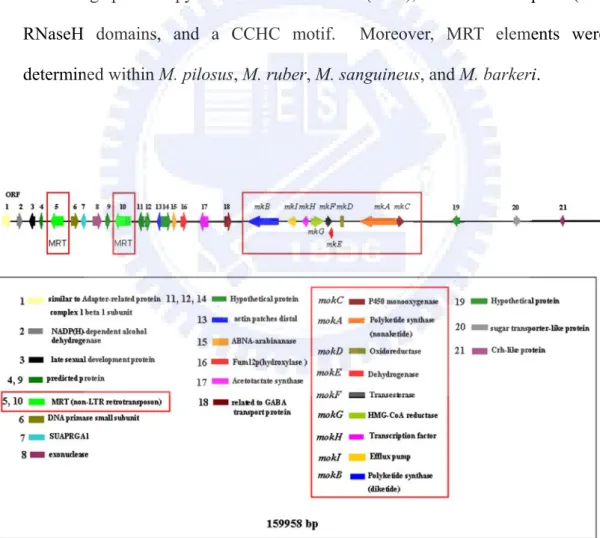
發現只有四種紅麴菌 M. pilosus,M. ruber,M. sanguineus 與 M. barkeri 具有 MRT non-LTR 逆轉位子。除此之外,利用 β-tubulin 基因所建立的親緣關係可歸群出 MRT non-LTR 逆轉位子之存在與否。
紅麴菌廣泛應用於食品發酵及保健藥物中,因此如何分辨紅麴菌中是否含有 橘黴素(citrinin)則相對性的重要,本研究分析了 18 株不同的紅麴菌種,發現只有 M. purpureus 與 M. kaoliang 具橘黴素的基因群,並且也只有此兩個菌種能偵測到 橘黴素的表現量,相反的其他物種包括 M. pilosus,M. ruber,M. barkeri,M. floridanus,M. lunisporas 及 M. pallens 皆缺乏橘黴素基因群,此研究結果顯示橘 黴素只有紅麴菌 M. purpureus 與 M. kaoliang 會產生,另外,利用 β-tubulin 基因 所建立的親緣關係則亦可歸群出橘黴素基因群之存在與否。
Abstract
Monascus species can produce various secondary metabolites with polyketide structures as monacolin K, citrinin, and pigments. Monacolin K which inhibits cholesterol synthesis is also known as lovastatin isolated from Aspergillus terreus. Citrinin, the hepato-nephrotoxic agent and antibiotic against gram-positive bacteria, has been identified in Aspergillus and Penicillium spp. In previous studies, lovastatin and citrinin biosynthetic gene clusters have been characterized in Aspergillus terreus and Monascus purpureus respectively. To explore the monacolin K biosynthetic gene cluster in M. pilosus BCRC38072 producing monacolin K, construction of a BAC library was carried out. The putative monacolin K biosynthetic gene cluster was found within a 42-kb region in the mps01 clone. The deduced amino acid sequences encoded by the nine genes designated as mokA–mokI, which share over 54% similarity with the lovastatin biosynthetic gene cluster in A. terreus, were assumed to be involved in monacolin K biosynthesis. In order to characterize the putative monacolin K biosynthetic gene cluste, the valid and convenient gene transformation in Monascus pilosus BCRC38072 was established using nuclease inhibitor, aurintricarboxylic acid (ATA). A gene disruption constructed to replace the central part of mokA, a polyketide synthase gene, in wild type M. pilosus BCRC38072 with a hygromycin B resistance gene through homologous recombination resulted in a mokA-disrupted strain. The disruptant did not produce monacolin K, indicating that mokA encoded the PKS responsible for monacolin K biosynthesis in M. pilosus BCRC38072. Moreover, the transformant containing two copies of the mokH gene-encoded transcription factor showed higher production of monacolin K than wild type strain. Real-time RT-PCR analysis also demonstrated that the transcripts of monacolin K biosynthetic genes in the
transformant were higher than those in wild type strain. These results suggested that mokA and mokH involved in the monacolin K biosynthesis. In addition, the mokA gene and sfp gene were coexpressed in Escherichia coli. The sfp gene obtained from Bacillus subtilis is a phosphopantetheinyl transferase required to convert the expressed apo-PKS to its holo form. Interestingly, anthranilic acid which is the precursor of tryptophan was found. This result revealed that that expression product of polyketide synthase (mokA) in E. coli was responsible for the elicitor to regulate anthranilate flux through the anthranilate synthase gene specific to the tryptophan biosynthetic pathway.
During the BAC (mps01) sequencing of M. pilosus BCRC38072, a new non-LTR retrotransposon, named MRT, was discovered. The entire nucleotide sequence of the MRT element was 5.5-kb long, including two open reading frames. These two ORFs showed homologies to gag-like and pol-like gene products, and an A-rich sequence at the 3’ end of pol-like gene. ORF1 encoded a protein of 517 amino acids and contained a cysteine-rich zinc finger motif. ORF2 encoded a protein of 1181 amino acids and contained apurinic/apyrimidinic endonuclease (APE), reverse transcriptase (RT), RNaseH domains, and a CCHC motif. The phylogenetic analyses demonstrated that the MRT element was classified into the Tad1 clade. The results of Southern hybridizations showed that MRT elements were distributed within M. pilosus, M. ruber, M. sanguineus, and M. barkeri. Also, the species of Monascus can be grouped by the presence or absence of MRT elements in the hybridization pattern according to phylogenetic subgroups established with the partial β-tubulin gene.
Monascus species has been widely used in food fermentation, and has shown a highly promising application in medicine development. Therefore, it is important to identify the non-citrinin producing Monascus strains. Eighteen Monascus strains
were investigated for the distribution of mycotoxin citrinin biosynthesis genes. These results showed that the genotype of citrinin genes was observed only in M. purpureus and M. kaoliang, while the phenotype of citrinin productivity was detected in M. purpureus and M. kaoliang. In contrast, the PCR and Southern blot results suggested that citrinin biosythesis genes were absent or significantly different in M. pilosus, M. ruber, M. barkeri, M. floridanus, M. lunisporas, and M. pallens. These results clearly indicated that the highly conserved citrinin gene cluster in M. purpureus and M. kaoliang carried out the citrinin biosynthesis. In addition, according to the phylogenetic subgroups established with the β-tubulin gene, the citrinin gene cluster can respectively group the species of Monascus.
Content 中文摘要 ……… ii Abstract ……… iv Catalog ……… vii Overview ……… 1 References ……… 16 Chapter 1 Improving the Genetic Transformation of Filamentous Fungus Monascus pilosus Using Aurintricarboxylic Acid (ATA) Abstract ……… 22
Introduction ……… 23
Materials and methods ……… 24
Results and discussion ……… 27
References ……… 33
Table ……… 36
Figure ……… 37
Chapter 2 Cloning and Characterization of Monacolin K Biosynthetic Gene Cluster from Monascus pilosus Abstract ……… 44
Introduction ……… 45
Materials and methods ……… 46
Results ……… 53
References ……… 64
Table ……… 68
Figure ……… 71
Chapter 3 Overexpression of mokA Encoding the Polyketide Synthase of Monascus pilosus in Escherichia coli Abstract ……… 85
Introduction ……… 86
Materials and methods ……… 87
Results ……… 90
Discussion ……… 92
References ……… 95
Figure ……… 98
Chapter 4 Exploring the Distribution of Citrinin Biosynthesis Related Genes among Monascus Species Abstract ……… 106
Introduction ……… 107
Materials and methods ……… 108
Results ……… 111 Discussion ……… 115 References ……… 119 Table ……… 122 Figure ……… 129 Chapter 5
Characterization of MRT, a New Non-LTR Retrotransposon in
Monascus spp.
Abstract ……… 136
Introduction ……… 137
Materials and methods ……… 138
Results and discussion ……… 140
References ……… 147
Table ……… 151
Table
Chapter 1
Improving the Genetic Transformation of Filamentous Fungus Monascus
pilosus Using Aurintricarboxylic Acid (ATA)
Table 1 The number of transformants based on various transformation
methods………... 36
Chapter 2
Cloning and Characterization of Monacolin K Biosynthetic Gene Cluster from Monascus pilosus
Table 1 Summary of genes identified in BAC mps01 obtained from M.
pilosus BCRC38072………... 68 Table 2 Real-time RT-PCR analysis of relative expression of transformant
T-mokH1 to M. pilosus BCRC38072………. 70
Chapter 4
Exploring the Distribution of Citrinin Biosynthesis Related Genes among
Monascus Species
Table 1 Strains used in this work for determination of citrinin………... 122 Table 2 Primers used to amplify citrinin related genes fragments………….. 124 Table 3 Strains used in this work for determination of monacolin K………. 126 Table 4 The concentration of citrinin produced by Monascus species……... 128
Chapter 5
Characterization of MRT, a New Non-LTR Retrotransposon in Monascus spp.
Figure
Overview
Figure 1 Biosynthesis of erythromycin from Saccharopolyspora erythraea 3
Figure 2 Biosynthesis of doxorubicin from Streptomyce peucetius……….. 3
Figure 3 Lovastatin gene cluster from Aspergillus terreus………... 4
Figure 4 Lovastatin biosynthesis pathway from A. terreus………... 6
Figure 5 Hexaketide and heptaketide expressed in A. nidulans by harboring lovB gene from A. terreus………... 7
Figure 6 Compactin gene cluster from Penicillium citrinum……….... 8
Figure 7 Compactin biosynthesis pathway from P. citrinum……….... 9
Figure 8 Citrinin biosynthesis pathway from M. purpureus………... 10
Figure 9 Citrinin gene cluster from Monascus purpureus………... 11
Figure 10 Annotation of BAC DNA mps01 predicted by BLAST and VectorNTI………... 15
Chapter 1 Improving the Genetic Transformation of Filamentous Fungus Monascus pilosus Using Aurintricarboxylic Acid (ATA) Figure 1 Morphology of M. pilosus………...………...………… 37
Figure 2 Map and Stability of pMS-1.5hp Plasmid………... 38
Figure 3 Submerged cultures of the wild type BCRC38072 and the T7 transformant………...………...………... 40
Figure 4 Analyses of Southern Hybridization and Fluorescence of M. pilosus transformants………...………...……….... 41
Cloning and Characterization of Monacolin K Biosynthetic Gene Cluster from Monascus pilosus
Figure 1 Identification of the monacolin K biosynthetic gene cluster of
Monascus pilosus BCRC38072………...………... 71 Figure 2 Deduced amino acid sequences alignment of transcription factors
from the mokH, lovE, and mlcR genes………...………... 73 Figure 3 Phylogenetic tree of PKSs from M. pilosus BCRC38072 and
various organisms………...………...………... 74 Figure 4 Disruption of the mokA gene in M. pilosus BCRC38072 76 Figure 5 Identification of monacolin K produced by M. pilosus
BCRC38072………...………...………... 78 Figure 6 mokH gene expression vector pMSmokH for genetic
transformation in M. pilosus BCRC38072………... 80 Figure 7 Analysis of the mokH gene containing gpd promoter and trpC
terminator………...………...………... 81 Figure 8 Southern hybridization analyses of copy number of the mokH
gene in the genomes of M. pilosus BCRC38072 and transformants 82 Figure 9 Submerged cultures of the wild type BCRC38072 and the
T-mokH1 transformant………...………...………... 83
Chapter 3
Overexpression of mokA Encoding the Polyketide Synthase of Monascus
pilosus in Escherichia coli
Figure 1 Structures of natural and “unnatural” polyketide products……… 98 Figure 2 Expression of MokA and Sfp in E. coli………...………. 99 Figure 3 HPLC traces monitored at a UV wavelength of 254 nm with E. 100
coli extract
Figure 4 Identification of anthranilic acid produced by E. coli with
coexpression of pETmkA and pCDFsfp………...………….... 101 Figure 5 Real-time RT-PCR analysis of relative expression of E. coli
harboring the MokA and Sfp expression plasmids to control…… 104
Chapter 4
Exploring the Distribution of Citrinin Biosynthesis Related Genes among
Monascus Species
Figure 1 The citrinin gene cluster of M. purpureus BCRC33325
(IFO30873) ………...………...………...……... 129 Figure 2 The Southern hybridization analyses of the citrinin-related genes 130 Figure 3 Phylogenetic tree of PKSs from Monascus and various organisms 132 Figure 4 Sequence analyses of the pksCT KS domain………... 133 Figure 5 Phylogeny of the Monascus species based on the partial β-tubulin
gene………...………...………...………... 134
Chapter 5
Characterization of MRT, a New Non-LTR Retrotransposon in Monascus spp.
Figure 1 Structure of the MRT element………...………... 153 Figure 2 Deduced Amino acid sequences alignment of the MRT elements
ORF1………...………...………...………... 154 Figure 3 Multiple alignment of deduced amino acid sequences of the MRT
elements with related organisms………...………... 155 Figure 4 Comparison of 3’ conserved region of the MRT elements from
Figure 5 Phylogenetic tree of non-LTR retrotransposons from M. pilosus
and various organisms………...………...……….... 159 Figure 6 Southern hybridizations analyses of the MRT elements………… 161 Figure 7 Phylogeny of Monascus species based on the partial β-tubulin
Overview
Introdution
Monascus is a filamentous fungus which belongs to the class Ascomycetes and the family Monascaceae. Thirteen Monascus species have been reported. Asexual sporulation is a common reproductive mode for Monascus species. Sexual reproduction, which produces ascospores, has been observed (Carels and Shepherd 1975). Monascus species are known as producers of various secondary metabolites with polyketide structures as monacolins, pigments, and citrinin (Endo 1979; Shimizu et al., 2005). It has been applied in the food industry for thousands of years in Asia and discovered that Monascus produces several bioactive substances. These bioactive substances are mainly the secondary metabolites including substances for reducing hypertension, substances of anti-putrefaction bacteria such as monascidin, anti-cancer substances, substances for lowering blood sugar, ergosterol, anti-oxidants, and inhibitors of cholesterol synthesis such as monacolin. Moreover, the pigments produced by the various species of Monascus have been extensively used as natural food colorants (Babitha et al., 2007). Monascus can produce yellow, orange, and red pigments. The red pigment is regarded as the most important among these pigments because it can be used as substitutes for nitrites in meat products. It has enormous commercial value in terms of increasing the production of secondary metabolites. Therefore, Monascus has been valued as a functional health food.
Polyketide biosynthesis produced by microorganisms
Polyketide are secondary metabolites extensively produced by bacteria and fungi (Pfeifer and Khosla, 2001). The biosynthesis process of polyketide in
microorganisms is similar to bacterial and mammalian fatty acid synthesis. Bacteria produced polyketide are known to be classified into type I and type II polyketide synthase (PKS) systems according to the biosynthesis of polyketides (Pfeifer and Khosla, 2001). Modular system (type I PKS) contains multifunctional enzymesthat are organized into modules. A module consisting of many motifs includes keto-synthase (KS), acyltransferase (AT), keto-reductase (KR), dehydratase (DH), enoyl rductase (ER), and acyl carrier protein (ACP) (Marsden et al., 1998) (Figure 1). Iterative system (type II PKS) contains each active sites encoded by separate enzymes. In addition, there is a significant characterization of type II polyketide compound with the aromatic group such as antibiotics doxorubicin (Pfeifer and Khosla, 2001) (Figure 2). However, the biosynthetic pathway of polyketides produced by fungi is different from bacteria. Fungal PKS defined as iterative type I PKS makes polyketides similar to bacterial type II PKS, but active sites of PKS are encoded by one enzyme instead of separate enzymes (Pfeifer and Khosla, 2001). According to the reduction of polyketide structures, fungal PKSs are classified into three types, highly-reduced (HR), partially-reduced (PR), and non-reduced (NR) (Nicholson et al., 2001). The structural diversity of polyketide also revealed the variety of biological activity. A number of polyketides have been used as drugs for human being diseases (Pfeifer and Khosla, 2001). For example, monacolin K produced by fungal iterative type I PKS has been used for reducing cholesterol synthesis.
Figure 1. Biosynthesis of erythromycin from Saccharopolyspora erythraea.
Monacolin K (Lovastatin) and Compactin biosynthesis
Monacolin K, the cholesterol synthesis inhibitor, is first isolated from the medium of Monascus ruber (Endo, 1979) and found the same substance from Aspergillus terreus as lovastatin (Hendrickson et al., 1999; Kennedy et al., 1999). The structure of monacolin K shares similarity with HMG-CoA; therefore, monacolin K competitively inhibits HMG-CoA reductase with HMG-CoA during cholesterol synthesis resulting in reduction of cholesterol synthesis. In previous studies, the lovastatin biosynthetic gene cluster has been proposed in A. terreus (Figure 3). Two polyketide synthases, transesterase, enoyl reductase, and P450 monooxygenase have been proved to involve in the structural biosynthesis of lovastatin (Hutchinson et al., 2000; Sorensen et al., 2003a; Sorensen et al., 2003b).
Figure 3. Lovastatin gene cluster from Aspergillus terreus. lovB, polyketide synthase; lovF, polyketide synthase; lovA, P450 monooxygenase; lovG, oxidoreductase; lovC dehydrogenase; lovD, transesterase; lvrA, HMG-CoA reductase; lovE, transcription factor and lovI, efflux pump.
The lovB encoding the polyketide synthase is responsible for nonaketide of lovastatin biosynthesis. According to the previous study, lovastatin can not be determined in the BX102 mutant, which is defective in lovB (Hendrickson et al., 1999). The cosmid library constructed from wild-type A. terreus is used to
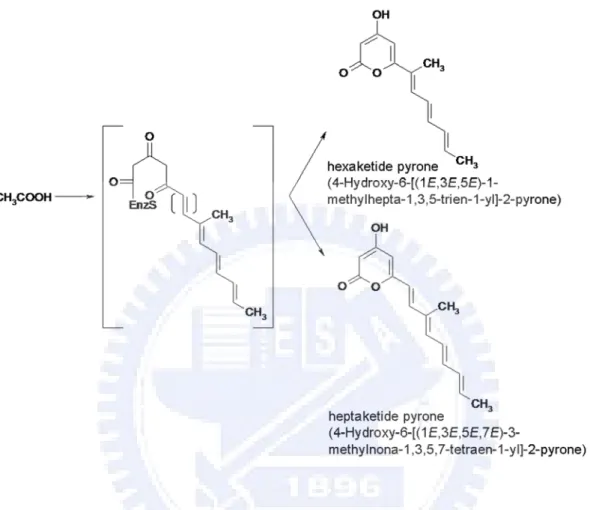
complement the BX102 mutant. One transformant is identified with cosmid containing lovB gene for lovastatin producer (Hendrickson et al., 1999). The disruption of lovC blocks in lovastatin production and causes accumulation of the new compound, 4-Hydroxy-6-[(1E,3E,5E)-1-methylhepta-1,3,5-trien-1-yl]-2-pyrone. Coexpression of lovB and lovC in A. nidulans results in the production of dihydromonacolin L. Furthermore, the disruption of the lovD gene generates accumulation of monacolin J which is the precursor of lovastatin. Consequently, LovD could join together the two polyketide, 2-methylbutyryl and monacolin J, into lovastatin (Figure 4). The lovE encodes protein with binuclear Zn+2 finger motif characteristic of eukaryotic transcription factors (Kennedy et al., 1999). Transformation of an extra copy of the lovE gene-encoded transcription factor into the wild type strain resulted in a 7- to 10-fold overproduction of lovastatin (Hutchinson et al., 2000). The lovI mutant does not produce lovastatin or its known precursors. The deduced amino acid of lovI resembles known transmembrane metabolite transport proteins of microorganisms. In addition, lovB has successfully expressed in Aspergillus nidulans, and produces two novel polyketides, hexaketide pyrone (4-Hydroxy-6-[(1E,3E,5E)-1-methylhepta- 1,3,5-trien-1-yl]-2-pyrone) and heptaketide pyrone (4-Hydroxy-6-[(1E,3E,5E,7E)- 3-methylnona-1,3,5,7-tetraen-1-yl]-2-pyrone) (Figure 5).
Fugure 4. Lovastatin biosynthesis pathway from A. terreus. (A) lovB encodes nonaketide synthase is involved in the synthesis of monacolin J. (B) lovF encodes diketide synthase is involved in the synthesis of 2-methylbutyric acid. (C)
2-methylbutyryl and monacolin J join together into lovastatin by LovD.
Figure 5. Hexaketide and heptaketide expressed in A. nidulans by harboring lovB gene from A. terreus.
Compactin is also the cholesterol synthesis inhibitor isolated from Penicillium citrinum (Abe et al., 2002). The structure of compactin differs from that of monacolin K, in which the C-6 position of the nonaketide-derived backbone lacks a methyl group derived from S-adenosyl-L-methionine (SAM). Recently, the compactin gene cluster is characterized (Abe et al., 2002a) (Figure 6). An extensive comparison analysis of these nine genes indicates compactin gene cluster
shared high similarity with the genes involved in lovastatin biosynthesis.
Figure 6. Compactin gene cluster from Penicillium citrinum. mlcA, polyketide synthase; mlcB, polyketide synthase; mlcC, P450 monooxygenase; mlcF, oxidoreductase; mlcG dehydrogenase; mlcH, transesterase; mlcD, HMG-CoA reductase; mlcR, transcription factor and mlcE, Efflux pump.
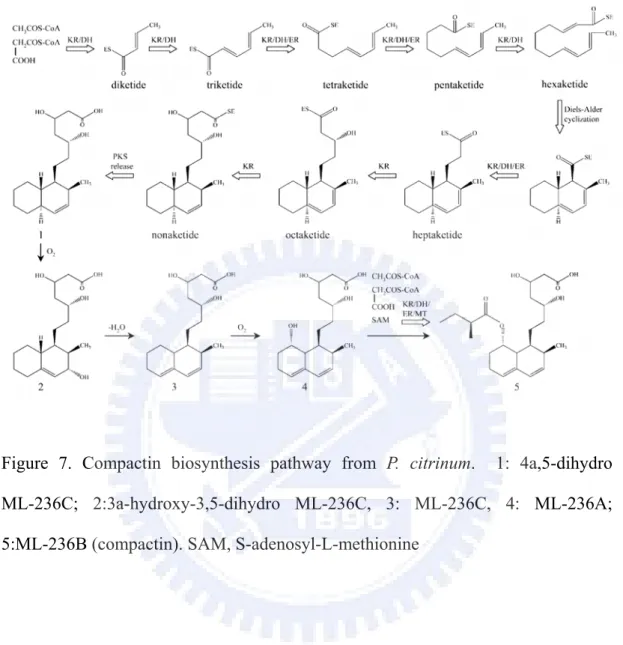
The polyketide synthases (mlcA and mlcB ) and regulatory (mlcR) have been proved to be involved in compactin biosynthesis (Abe et al., 2002a; Abe et al., 2002b; Abe et al., 2002c) (Figure 7). The mlcA encoding polyketide synthase is responsible for nonaketide of compactin biosynthesis. As such, the disruption of mlcA results in the phenotype of lost compactin productivity (Abe et al., 2002a). The mlcB encoding polyketide synthase is responsible for diketide of compactin biosynthesis. The mlcB mutant demonstrates the accumulation of ML-236A (Figure 7). The mlcR that encodes the Zn(II)2Cys6 binuclear DNA binding protein is a regulatory of citrinin biosynthesis. Transformation of an extra copy of the mlcR gene-encoded transcription factor into the wild type strain results in a 10-15% increase of compactin (Abe et al., 2002b). Furthermore, introduction of the cosmid, which contains seven of the nine compactin biosynthetic genes, into P. citrinum can increase the yield of compactin of transformants (Abe et al., 2002c). It is interesting to compare the monacolin K, lovastatin, and compactin gene clusters from Monascus, Aspergillus, and Penicillium. They will provide useful
information to study evolution of secondary metabolite.
Figure 7. Compactin biosynthesis pathway from P. citrinum. 1: 4a,5-dihydro ML-236C; 2:3a-hydroxy-3,5-dihydro ML-236C, 3: ML-236C, 4: ML-236A; 5:ML-236B (compactin). SAM, S-adenosyl-L-methionine
Citrinin biosynthesis
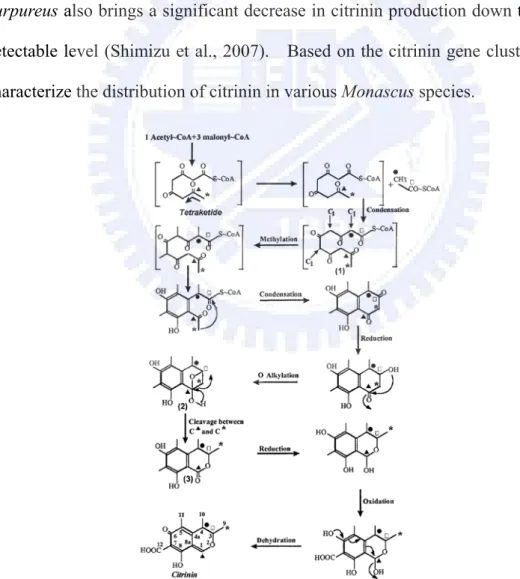
Citrinin is a polyketide produced by fungal species including the genus Aspergillus, Penicillium and Monascus. It is identified as a contaminant in several foods and resulted in accumulation in the mitochondria inducing apoptosis at the cellular level. The biosynthesis of citrinin synthesized by the iterative type I polyketide synthase (PKS) originates from a tetraketide arising from the condensation of one acetyl-CoA molecule with three malonyl-CoA molecules in Monascus (Hajjaj et al., 1999) (Figure 8).
The citrinin biosynthetic gene cluster has been proposed in M. purpureus BCRC33325 (IFO30873) in Shimizu’s studies (2005) (Figure 9). pksCT is predicted to be responsible for the synthesis of the polyketide skeleton. Also included are a dehydrogenase (orf1), a transcriptional activator (ctnA), an oxygenase (orf3), an oxidoreductase (orf4), and a transporter gene (orf5). The pksCT encoding polyketide synthase is responsible for such biosynthesis. Therefore, the disruption of pksCT results in the phenotype of lost citrinin productivity (Shimizu et al., 2005). The ctnA that encodes the Zn(II)2Cys6 binuclear DNA binding protein is a major activator of citrinin biosynthesis. The ctnA-disrupted strain of M. purpureus also brings a significant decrease in citrinin production down to a barely detectable level (Shimizu et al., 2007). Based on the citrinin gene cluster, we can characterize the distribution of citrinin in various Monascus species.
Figure 9. Citrinin gene cluster from Monascus purpureus. orf1, dehydrogenase; ctnA, transcriptional activator; orf3, oxygenase; orf4, oxidoreductase; pksCT, polyketide synthase; and orf5, transporter.
Retrotransposon
Retrotransposons are integral components of eukaryotic genome. The copy-and-paste mechanism of retrotransposons results in diversity of organisms and plays an important role in evolutionary history and population dynamics (Daboussi and Capy, 2003). Most retrotransposons are present in ancestral species and evolve with time. However, the distribution of retrotransposon is present in some species and lost in others, possibly due to genetic drift (Engels, 1981). Analyses of this distribution could exploit retrotransposons behavior in the various species, and allow us to group the species in the phylogenetic subgroups previously established with other characters (Cizeron et al. 1998; Blesa et al., 2001).
Retrotransposons can be divided into two major classes: long terminal repeat (LTR) and non-LTR retrotransposons. Approximately 30 transposable elements corresponding to both classes are recognized in fungi (Daboussi and Capy, 2003). Non-LTR retrotransposon, also known as long interspersed nuclear elements (LINE), contains a significant feature lacking in long terminal repeats. A mechanism of non-LTR retrotransposon for reverse transcription and integration of a new copy has been suggested in which reverse transcript becomes inserted at the nicks in the chromosome during DNA repair or recombination by host-encoded proteins (Flavell, 1995; Finnegan, 1997). Moreover, a relatively simple insertion mechanism of
non-LTR retrotransposon is proposed. Non-LTR retrotransposon inserts itself into the chromosome using the 3’ end of the broken DNA digested by the endonuclease domain of retrotransposon’s product to prime cDNA synthesis, a process termed target-primed reverse transcription (TPRT) (Chaboissier et al., 2000; Bibillo and Eickbush, 2002; Eickbush, 2002). Additionally, it has been suggested that DNA repair of a host’s mechanism may involve the retrotransposition for the integration of the 5’ end of R2Bm non-LTR retrotransposon (Fujimoto et al., 2004). Most non-LTR retrotransposons in fungi contain two open reading frames. The first one is considered to be the gag gene containing cysteine-rich nucleotide-binding domains though it does not show a conserved protein sequence. The second one encodes several conserved domains including endonuclease, reverse transcriptase, RNaseH domains, and zinc finger motifs (Daboussi and Capy, 2003). Extensive analysis of non-LTR retrotransposon in fungi has revealed the presence of numerous mutations resulting from the repeat-induced point mutation (RIP) process (Selker et al., 1987). The genome of N. crassa shows that most of the repetitive sequences (81%) have been mutated by RIP (Galagan et al., 2003). Methylation-induced premeiotically (MIP) occurs during the sexual phase of the fungal life cycle and leads to the methylation of cystosine residues (Faugeron et al., 1990). The occurrence of MIP also suggests that this process is directed against repetitive sequences (Goyon et al., 1996).
Five non-LTR retrotransposons have been characterized in fungi including Tad1-1 in Neurospora crassa (Cambareri et al., 1994), MGR583 in Magnaporthe grisea (Hamer et al., 1989), CgT1 in Colletotrichum gloeosporioides (He et al., 1996), marY2N in Tricholoma matsutake (Murata et al., 2001), and Mars in Ascobollus immerses (Goyon et al., 1996) though the entire element is not described for the last species. Non-LTR retrotransposons are classified into eleven clades
(Malik et al., 1999). Based on sequence, structure, and phylogenetic analyses, the elements from genomes of fungi belong to the Tad1 clade. Recently, two non-LTR retrotransposons of yeast, Zorro in Candida albicans (Goodwin et al., 2001) and Ylli in Yarrowia lipolytica (Casaregola et al., 2002), have been characterized. These two non-LTR retrotransposons belong to the L1 clade of elements known from human beings and mice. So far, no attempt was made to analyze the non-LTR retrotransposon in Monascus. It will provide useful information for the phylogenetic characterization in Monascus.
The specific aims of this study:
Monascus has been used in Chinese fermented foods for thousands of years. Moreover, many secondary metabolites have been identified such as monacolins, citrinin, GABA (gamma-aminobutyric acid), ergosterol, and various pigment compounds. However, only a few attempts have been made at genetic investigation of Monascus species. In the present study, a BAC (Bacterial Artificial Chromosome) library of the strain M. pilosus BCRC38072 about 12,000 clones was constructed for the genomic research. According to the BAC clones, monacolin K, citrinin, and non-LTR retrotransposon related genes were examined. The evolutionary relationship was further explored among different Monascus species. Therefore, the following specific aims were studied.
1. The efficiency of genetic transformation is depending on different Monascus strains. Although the genetic transformation of M. ruber and M. purpureus has been developed (Yang and Lee, 2008; Kim et al., 2003), no attempt has been made on genetic transformation of M. pilosus BCRC38072. Here, a method of genetic transformation of high efficiency was established using nuclease
inhibitor, aurintricarboxylic acid (ATA). The results demonstrated that ATA can improve the frequency of genetic transformation in M. pilosus. This valid and convenient method shows that ATA can be applied to fungi for efficient genetic transformation.
2. Although the lovastatin biosynthetic gene cluster in A. terreus has been characterized (Kennedy et al., 1999), the structural genes responsible for monacolin K (lovastatin) biosynthesis in Monascus are still unclear. Firstly, to explore the monacolin K biosynthetic gene cluster, a BAC library from M. pilosus BCRC38072 producing monacolin K was constructed (Figure 10). The nine genes were analyzed and found to share high similarity with the genes involved in the lovastatin biosynthetic gene cluster of Aspergillus terreus and compactin biosynthetic gene cluster of Penicillium citrinum. Moreover, the mokA encoding polyketide synthase and mokH encoding transcription factor were futher disrupted and overexpressed in M. pilosus BCRC38072 respectively. 3. Only a few attempts have been made at heterologous expression of fungal
polyketide synthase. mokA encoding polyketide synthase was expressed in Escherichia coli for development of novel polyketide. However, an unexpected product, anthranilic acid which is the precursor of tryptophan, was observed. These results revealed that expression product of polyketide synthase (mokA) in E. coli was responsible for the elicitor to improve the activity of anthranilate synthase.
4. Monascus has been valued as a functional health food. Therefore, it is important to identify the non-citrinin producing Monascus strains or generally eliminate the production of citrinin in Monascus. Various Monascus species were analyzed for the presence of the citrinin genes and production of citrinin. The citrinin genes were observed only in M. purpureus and M. kaoliang.
Likewise, the citrinin productivity was detected in M. purpureus and M. kaoliang.
5. During the whole genome sequencing of M. pilosus BCRC38072, two repetitive sequences (mps01-1 and mps01-2) were observed in a ca. 160kb BAC, mps01 (Figure 10). These two repetitive sequences were new non-LTR retrotransposon named MRT (Monascus Retrotransposon). The entire nucleotide sequence of the MRT element was 5.5-kb long, including two open reading frames, ORF1 containing a cysteine-rich zinc finger motif and ORF2 containing apurinic/apyrimidinic endonuclease (APE), reverse transcriptase (RT), RNaseH domains, and a CCHC motif. Moreover, MRT elements were determined within M. pilosus, M. ruber, M. sanguineus, and M. barkeri.
Reference
(1) Abe, Y.; Suzuki, T.; Ono, C.; Iwamoto, K.; Hosobuchi, M.; Yoshikawa, H. Molecular cloning and characterization of an ML-236B (compactin) biosynthetic gene cluster in Penicillium citrinum. Mol. Genet. Genomics. 2002a, 267, 636-646.
(2) Abe, Y.; Ono, C.; Hosobuchi, M.; Yoshikawa, H. Functional analysis of mlcR, a regulatory gene for ML-236B (compactin) biosynthesis in Penicillium citrinum. Mol. Genet. Genomics. 2002b, 268, 352-361
(3) Abe, Y.; Suzuki, T.; Mizuno, T.; Ono, C.; Iwamoto, K.; Hosobuchi, M.; Yoshikawa, H.; Effect of increased dosage of the ML-236B (compactin) biosynthetic gene cluster on ML-236B production in Penicillium citrinum. Mol. Genet. Genomics. 2002c, 268, 130-137.
(4) Babitha, S.; Soccol, C. R.; Pandey, A. Solid-state fermentation for the production of Monascus pigments from jackfruit seed. Bioresour. Technol. 2007, 98, 1554-1560.
(5) Bibillo, A.; Eickbush, T.H. The reverse transcriptase of the R2 non-LTR retrotransposon: continuous synthesis of cDNA on non-continuous RNA templates. J. Mol. Biol. 2002, 316, 459-473.
(6) Blesa, D.; Gandía, M.; Martínez-Sebastián, M.J. Distribution of the bilbo non-LTR retrotransposon in Drosophilidae and its evolution in the Drosophila obscura species group. Mol. Biol. Evol. 2001, 18, 585-592.
(7) Cambareri, E.B.; Helber, J.; Kinsey, J.A. Tad1-1, an active LINE-like element of Neurospora crassa. Mol. Gen. Genet. 1994, 242, 658-665.
(8) Carels, M.; Shepherd, D. Sexal reproductive cycle of Monascus in submerged shaken culture. J. Bacteriol. 1975, 122, 288-294.
retrotransposon L1 family in the dimorphic yeast Yarrowia lipolytica. Mol. Biol. Evol. 2002, 19, 664-677.
(10) Chaboissier, M.C., Finnegan, D.; Bucheton, A. Retrotransposition of the I factor, a non-long terminal repeat retrotransposon of Drosophila, generates tandem repeats at the 3' end. Nucleic Acids Res. 2000, 28, 2467-2472.
(11) Cizeron, G.; Lemeunier, F.; Loevenbruck, C.; Brehm, A.; Biémont., C. Distribution of the retrotransposable element 412 in Drosophila species. Mol. Biol. Evol. 1998, 15, 1589-1599.
(12) Daboussi, M.J.; Capy, P. Transposable elements in filamentous fungi. Annu. Rev. Microbiol. 2003, 57, 275-299.
(13) Eickbush, T.H. Repair by retrotransposition. Nat. Genet. 2002, 31, 126-127. (14) Endo, A. Monacolin K, a new hypocholesterolemic agent produced by a
Monascus species. J. Antibiot. (Tokyo) 1979, 32, 852-854.
(15) Engels, W.R. Estimating genetic divergence and genetic variability with restriction endonucleases. Proc. Natl. Acad. Sci. USA. 1981, 78, 6329-6333. (16) Faugeron, G., Rhounim, L.; Rossignol, J.L. How does the cell count the number
of ectopic copies of a gene in the premeiotic inactivation process acting in Ascobolus immersus? Genetics. 1990, 124, 585-591.
(17) Finnegan, D.J. Transposable elements: how non-LTR retrotransposons do it. Curr. Biol. 1997, 7, 245-248.
(18) Flavell, A.J. Retroelements, reverse transcriptase and evolution. Comp. Biochem. Physiol. 1995, 110, 3-15.
(19) Fujimoto, H.; Hirukawa, Y.; Tani, H.; Matsuura, Y.; Hashido, K.; Tsuchida, K.; Takada, N.; Kobayashi, M.; Maekawa, H. Integration of the 5' end of the retrotransposon, R2Bm, can be complemented by homologous recombination. Nucleic Acids Res. 2004, 32, 1555-1565.
(20) Galagan, J.E.; Calvo, S.E. et al. (77 co-authors). The genome sequence of the filamentous fungus Neurospora crassa. Nature. 2003, 422, 859-868.
(21) Goodwin, T.J.; Ormandy, J.E.; Poulter, R.T. L1-like non-LTR retrotransposons in the yeast Candida albicans. Curr. Genet. 2001, 39, 83-91.
(22) Goyon, C., Rossignol, J.L.; Faugeron, G. Native DNA repeats and methylation in Ascobolus. Nucleic. Acids. Res. 1996, 24, 3348-3356.
(23) Hajjaj, H.; Klaebe, A.; Loret, M. O.; Goma, G.; Blanc, P. J.; Francois, J. Biosynthetic pathway of citrinin in the filamentous fungus Monascus ruber as revealed by 13C nuclear magnetic resonance. Appl. Environ. Microbiol. 1999, 65, 311-314.
(24) Hamer, J.E.; Farrall, L.; Orbach, M.J.; Valent, B.; Chumley, F.G. Host species-specific conservation of a family of repeated DNA sequences in the genome of a fungal plant pathogen. Proc. Natl. Acad. Sci. USA. 1989, 86, 9981-9985.
(25) He, C.; Nourse, J.P.; Kelemu, S.; Irwin, J.A.; Manners, J.M. CgT1: a non-LTR retrotransposon with restricted distribution in the fungal phytopathogen Colletotrichum gloeosporioides. Mol. Gen. Genet. 1996, 252, 320-331.
(26) Hendrickson, L.; Davis, C. R.; Roach, C.; Nguyen, D. K.; Aldrich, T.; McAda, P. C.; Reeves, C. D. Lovastatin biosynthesis in Aspergillus terreus: characterization of blocked mutants, enzyme activities and a multifunctional polyketide synthase gene. Chem. Biol. 1999, 6, 429-439.
(27) Hutchinson, C. R.; Kennedy, J.; Park, C.; Kendrew, S.; Auclair, K.; Vederas, J. Aspects of the biosynthesis of non-aromatic fungal polyketides by iterative polyketide synthases. Antonie Van Leeuwenhoek. 2000, 78, 287-295.
(28) Kennedy, J.; Auclair, K.; Kendrew, S. G.; Park, C.; Vederas, J. C.; Hutchinson, C. R. Modulation of polyketide synthase activity by accessory proteins during
lovastatin biosynthesis. Science. 1999, 284, 1368-72.
(29) Kim, J.G.; Choi, Y.D.; Chang, Y.J.; Kim, S.U. Genetic transformation of Monascus purpureus DSM1379. Biotechnol. Lett. 2003, 25, 1509-1514.
(30) Malik, H.S; Burke, W.D.; Eickbush, T.H. The age and evolution of non-LTR retrotransposable elements. Mol. Biol. Evol. 1999, 16, 793-805.
(31) Marsden, A.F.; Wilkinson, B.; Cortés, J.; Dunster, N.J.; Staunton, J.; Leadlay, P.F. Engineering broader specificity into an antibiotic-producing polyketide synthase. Science. 1998, 279, 199-202.
(32) Murata, H.; Miyazaki, Y.; Yamada, A. marY2N, a LINE-like non-long terminal repeat (non-LTR) retroelement from the ectomycorrhizal homobasidiomycete Tricholoma matsutake. Biosci. Biotechnol. Biochem. 2001, 65, 2301-2305.
(33) Nicholson, T.P.; Rudd, B.A.; Dawson, M.; Lazarus, C.M.; Simpson, T.J.; Cox, R.J. Design and utility of oligonucleotide gene probes for fungal polyketide synthases. Chem. Biol. 2001, 8, 157-178.
(34) Pfeifer, B. A.; Khosla, C. Biosynthesis of polyketides in heterologous hosts. Microbiol. Mol. Biol. Rev. 2001, 65, 106-118.
(35) Selker, E.U.; Cambareri, E.B.; Jensen, B.C.; Haack, K.R. Rearrangement of duplicated DNA in specialized cells of Neurospora. Cell. 1987, 51, 741-752. (36) Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T.
Polyketide synthase gene responsible for citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453-3457.
(37) Shimizu, T.; Kinoshita, H.; Nihira, T. Identification and in vivo functional analysis by gene disruption of ctnA, an activator gene involved in citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2007, 73, 5097-5103.
Transformations of cyclic nonaketides by Aspergillus terreus mutants blocked for lovastatin biosynthesis at the lovA and lovC genes. Org. Biomol. Chem.
2003a, 1, 50-59.
(39) Sorensen, J. L.; Vederas, J. C. Monacolin N, a compound resulting from derailment of type I iterative polyketide synthase function en route to lovastatin. Chem. Commun. 2003b, 13, 1492-1493.
(40) Yang, Y.J.; Lee, I. Agrobactrium tumefaciens-mediated transformation of Monascus ruber. J. Microbiol. Biotechnol. 2008, 18, 754-758.
Chapter 1
Improving the Genetic Transformation of Filamentous
Fungus Monascus pilosus Using Aurintricarboxylic Acid
(ATA)
Abstract
The filamentous fungus, Monascus pilosus, was genetically transformed with a reporter plasmid, pMS-1.5hp, using aurintricarboxylic acid (ATA) treatment to obtain a high red pigment-producing mutant. The transformation efficiency of Monascus pilosus was higher with the ATA-treatment than with either a non- restriction-enzyme-mediated integration (REMI) or a REMI method. This valid and convenient random mutagenesis method shows that ATA can be applied in fungi for efficient genetic transformation.
Key words: Monascus pilosus; restriction-enzyme-mediated integration (REMI); aurintricarboxylic acid (ATA).
Introduction
Monascus spp. are filamentous fungi that are known as producers of various secondary metabolites with polyketide structures as pigments, monacolins and citrinin (Endo et al., 1986; Shimizu et al., 2005). The production of red pigments by various species of Monascus has been extensively used as natural food colorants (Babitha et al., 2007a). Monacolin K, also known as lovastatin for reducing serum cholesterol levels in human beings, was first isolated from the medium of Monascus ruber (Endo et al., 1979; Tobert, 2003). The development of the transformation system of Monascus should enable the function of polyketide biosynthesis to be examined by constructing knock-out mutants, or high-economic polyketides, such as monacolins and pigments, to be produced.
The preparation of fungal protoplasts is usually the typical approach for delivering the transforming DNA into filamentous fungus. The protoplasts are mixed with a combination of CaCl2 and polyethylene glycol (PEG), which cultivate
the transforming DNA. Restriction-enzyme-mediated integration (REMI) (Kahmann and Basse, 1999) and the Agrobacterium tumefaciens-mediated transformation (ATMT) (de Groot et al., 1998) methods have been established to improve the frequency of transformation substantially. The REMI method has been adopted in various filamentous fungi (Cantone and Vandenberg, 1999; Mullins and Kang, 2001; Thon et al., 2000). The method is based on the addition of a restriction enzyme into the mix of protoplasts and the transforming DNA, such that the specific restriction enzyme generates a site for inserting DNA into genomic locus (Mullins and Kang, 2001). The process increases the efficiency of transformation; however, adding different restriction enzymes and concentrations yields different efficiencies of transformation (Cantone and Vandenberg. 1999; Jin et al., 2005).
Aurintricarboxylic acid (ATA) is an inhibitor for the inhibition of the activity of DNaseI, S1 nuclease, exonuclease III and RNAse A (Hallick et al., 1977). It has been demonstrated to be useful in protecting plasmid DNA from nuclease degradation and increasing the transfer efficiency of DNA in the respiratory tissues and the skin of mice, pigs and macaques (Glasspool-Malone and Malone, 1999; Glasspool-Malone et al., 2000; Glasspool-Malone et al., 2002; Walther et al., 2005). In this study, the use of ATA was investigated for the genetic transformation in M. pilosus. The combination of CaCl2 and PEG was employed to analyze the
efficiency of genetic transformation between ATA-treatment and REMI. The transformation plasmid, including the promoter of heat shock protein 90 (hsp 90) from M. pilosus, was constructed. Additionally, the fusion of the hygromycin B resistance gene (HPH) with enhanced green fluorescent protein (EGFP) was expressed in M. pilosus.
Materials and methods
Strain used and growth conditions
M. pilosus BCRC38072, a monacolin K-producing strain that was isolated from red rice (anka) collected from a traditional local market, was used in this study. To prepare the protoplasts, the strain was incubated on PDB (DIFCO 254920, Detroit, Michigan) agar for one week, and spore suspensions were obtained by washing cultured PDA plates.
Construction of transformation vector
Biosciences Clontech, Palo. Alto, CA) was digested by BglII-XhoI and blunted using an End-It DNA End-repair kit (Epicentre, Madison, WS) to obtain the linear plasmid. The heat shock protein 90 (hsp 90) promoter from M. pilosus BCRC38072 (GenBank accession no. DQ983312) was amplified by the primer set (Mps9259F:
AGTGGCAGCCAACCCTCACC and Mps7782R: CGGGCTGATAGAGCAGATAGATAGATG). The 1.5kb PCR product was
phosphorylated and introduced into the linear plasmid pHygEGFP to obtain the plasmid pMS-1.5hp.
Protoplasts isolation
The preparation of protoplast developed from Neurospora crassa (Vollmer and Yanofsky, 1986) was modified for the genetic transformation of M. pilosus. The conidia from the one-week culture of M. pilosus were incubated in 100 ml of Vogel medium at 30oC for 16~18 hr. The mycelia were harvested on miracloth (Millipore, MA, USA) and washed in the MA digestion solution (0.1 M maleic acid, pH 5.5 and 1.2 M (NH4)2SO4). The mycelia were digested for 4~5 hr using 100 mg of Yatalase
(Takara, Shiga, Japan), 100 mg of lysing enzyme (Sigma, St.Louis, Mo., USA) and 100 μl of β-glucuronidase (Sigma, St.Louis, Mo., USA) in 50 ml MA digestion solution. To remove the undigested mycelia, protoplasts were harvested by passing them through miracloth and by centrifugation at 1000 rpm for 10 min (Sorvall, CT, USA). The protoplasts were maintained in 80 % STC (1 M sorbitol, 50 mM Tris pH 8.0, 50 mM CaCl2) and 20 % PTC (40 % PEG 4000, 50 mM Tris pH 8.0, 50 mM
CaCl2), and dimethylsulfoxide (DMSO) was added to a final concentration of 1 %.
Transformation of M. pilosus BCRC38072
with 5 μg DNA-linearized and ATA (1 mM) or restriction enzyme (10 units). The mixtures were incubated on ice for 30 min. One milliliter PTC was added and mixed gently. Following incubation at room temperature for 20 min, the protoplast mixtures were added to 15 ml of SYP medium (1 M sorbitol, 0.1 % yeast extract, 0.1 % peptone and 2 % agar) that contained 60 μg hygromycin B/ml. The transformants were incubated at 30 oC.
Manipulations of nucleic acid
Fungal genomic DNA was isolated by liquid nitrogen treatment according to the method developed by Bingle et al. (1999). Southern hybridization was performed using the DIG system (Roche Diagnostics, Mannheim, Germany). The probe of hph gene was DIG-labeled by PCR amplification using the PCR DIG probe synthesis kit (Roche Diagnostics, Mannheim, Germany). The primer set of hph gene was pMS-f: GAACTGCCCGCTGTTCTGCA and pMS-r: ACATCGCCTCGCTCCAGTCA.
Fluorescence microscopy
The hyphae of transformants were compared by fluorescence microscopy to that of the wild type in EGFP expression. Fluorescence photomicrography of the specimens was conducted using a fluorescence microscope (Leica RXA, Mannheim, Germany). The red filter (Chroma, VT, USA) with peak transmission of excitation at 450 to 490 nm and peak transmission of suppression at 525 to 575 nm was used to detect EGFP.
Measurement of red pigment
and spore suspensions were obtained by washing cultured PDA agar plates with distilled water. Mycelia and media were harvested after incubating 3~10 days at 25
oC with constant agitation in liquid medium (7 % glycerol, 3 % glucose, 3 %
monosodium glutamate, 1.2 % polypetone, 0.2 % NaNO3 and 0.1 % MgSO4.7H2O).
The aliquots of M. pilosus and the transformant culture were cleared of cells, and were filtered by a 0.2 mm filter. The supernatants were analyzed at 500 nm by the spectrophotometer for the determination of red pigment.
Results and discussion
Preparation of protoplasts and sensitivity test of M. pilosus BCRC38072 to hygromycin B
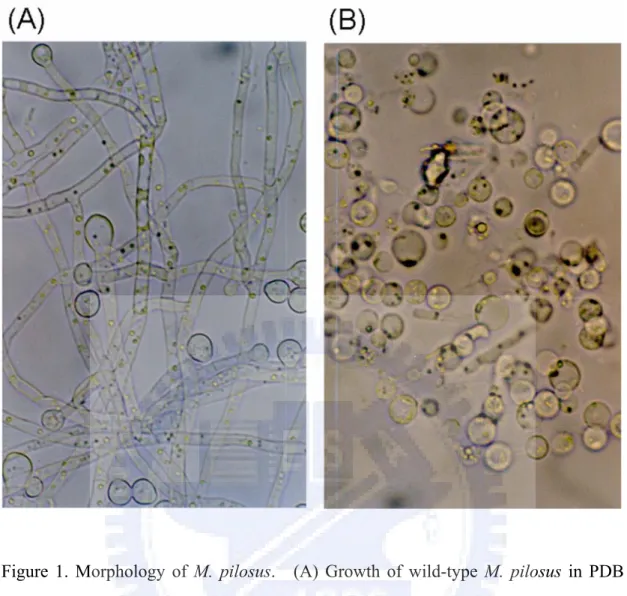
The mycelia of M. pilosus BCRC38072 were initially white, and then became reddish orange. The size of conidia was 10~13 × 8~10 μm and the ascospore was ellipsoid with dimensions of 4.6~6.3 × 3.3~4.2 μm. The conidia obtained after cultivation for seven days was transferred into the Vogel medium and was over 90 % germinated at 30 oC for 16~18 hr. The result revealed that a mixture of 100 mg Yatalase, 100 mg lysing enzyme and 100 μl β-glucuronidase was sufficient to convert hyphae (Figure 1A) into protoplasts (Figure 1B) when digested for 4~5 h in 50 ml of digestion solution, after which the protoplasts were used for genetic transformation. The protoplast yield for M. pilosus was 1.0~3.0×107 ml-1.
The inhibition of the growth of M. pilosus BCRC38072 was determined by plating conidia on a PDA plate with various hygromycin B concentrations - 0, 20, 30, and 40 μg/ml. The growth of M. pilosus BCRC38072 was completely inhibited at 30 μg/ml. However, the false-positive colonies were commonly involved in the
genetic transformation of fungi (Mort-Bontemps and Fèvre, 1997; Shimizu et al., 2006). To ensure the absence of false-positive colonies, high concentration of hygromycin B (60 μg/ml) was used to select resistant colonies in genetic transformation experiments.
Evaluation of efficiency of genetic transformation using the expression vector pMS-1.5hp
Hsp90, the 90kDa heat shock protein, is a major molecular chaperon which is distributed in all eukaryotes. It is the most abundant protein and occupies 1~2 % of all cellular proteins in the cytosols of eukaryotic cells (Csermely et al., 1998). Therefore, the construction of the expression vector using the hsp90 promoter from M. pilosus may promote the production of hygromycin-resistance protein. In this study, based on the Bacterial Artificial Chromosome (BAC) library constructed from M. pilosus BCRC38072 (Chen et al., 2007), the primer set was designed to amplify the promoter of the hsp90 gene (GenBank accession no. DQ983312). The human cytomegalovirus (CMV) promoter of the expression vector pHygEGFP, including a fusion of the hygromycin B resistance gene (HPH) with enhanced green fluorescent protein (EGFP), was replaced with the promoter of the hsp90 gene from M. pilosus BCRC38072 to yield the plasmid pMS-1.5hp (Figure 2A). Furthermore, a time course of pMS-1.5hp plasmid stability was analyzed in lysates from M. pilosus pretreated with 0, 0.1 and 1.0 mM ATA. We observed that 0.1 mM ATA protected the plasmid in 1 mg ml-1 of protein lysate, whereas the stability of the plasmid was decreased in 3.5 mg ml-1 of protein lysate (Figure 2B, C). These results indicate that the presence of higher ATA concentrations (1.0 mM) can prolong the stability of plasmid DNA. The expression vector was utilized to achieve the over-expression of the dual-function marker for the genetic transformation in M. pilosus BCRC38072.
The REMI method was established to substantially improve the frequency of transformation and adopted for use in various filamentous fungi (Cantone and Vandenberg, 1999; Mullins and Kang, 2001; Thon et al., 2000). The REMI method increases the efficiency of transformation; however, different efficiencies of transformation are achieved by adding different restriction enzymes at different enzyme concentrations (Cantone and Vandenberg, 1999; Jin et al., 2005). To understand the efficiency of genetic transformation with ATA treatment (1.0 mM), three procedures, including non-REMI, REMI and ATA-treatment, were carried out using the PEG-based protoplasts method (Vollmer and Yanofsky, 1986). The non-REMI method using FspI-linearized pMS-1.5hp had the lowest efficiency of genetic transformation (17 ± 2 transformants per 5 μg of linear plasmid), while the FspI-REMI had an efficiency of genetic transformation (48 ± 4 transformants per 5 μg of linear plasmid) that was 2.8 times greater than the non-REMI treatment (Table
1). The HpaI-REMI had a seven times higher efficiency of genetic transformation
(119 ± 11 transformants per 5 μg of linear plasmid) than the non-REMI treatment. Interestingly, the ATA-treatment yielded a 9.4 times higher efficiency of genetic transformation (159 ± 24 transformants per 5 μg of linear plasmid) when compared to the non-REMI transformation and a 3.3 and 1.3 times higher efficiency of genetic transformation compared to the FspI-REMI and HpaI-REMI respectively. These results suggest that adding ATA to the protoplasts increases the efficiency of genetic transformation in M. pilosus BCRC38072. This result is consistent with the results reported for the transformation of the respiratory tissues and skin of mice, pigs and macaques (Glasspool-Malone and Malone, 1999; Glasspool-Malone and Malone, 2002; Glasspool-Malone et al., 2000; Glasspool-Malone et al., 2002; Walther et al., 2005). The ATA effect on genetic transformation is likely a result of ATA’s ability to significantly reduce nuclease activity in cells, which prevents the transforming DNA from being degraded (Glasspool-Malone et al., 2002). The results of this study are also consistent with the findings of Cantone and Vandenberg, who
indicated that adding ATA results in a high efficiency of genetic transformation in the pathogen Paecilomyces fumosoroseus (Cantone and Vandenberg, 1999).
Characterization of the high red pigment transformant
Monascus produces various pigments that have enormous commercial value in terms of increasing the production of pigments (Babitha et al., 2007a). Previous studies have shown that high temperature, high concentration of NaCl, and improvement in oxygen supply can increase the yield of pigments (Babitha et al., 2007b; Hajjaj et al., 1999). In this study, 30 transformants were screened and one of these that had the highest production of red pigment was characterized. The result showed that random mutagenesis with efficient genetic transformation using ATA can obtain the transformant with high red pigment. The transformant T7 was compared with the wild type during submerged fermentation of 10 days. The red pigment production of the T7 transformant was 4 to 5-fold higher than the corresponding values of the wild type, while the cultured cell mass revealed no significant difference (Figure 3). After 5 days of cultivation, the red pigment increased in the T7 transformant. The maximum red pigment level was reached after 8 days of cultivation; however, the red pigment was slowly produced by the wild type and reached the maximum level during the 10th day of cultivation.
Confirmation of M. pilosus transformants
The high red pigment-producing transformant and seven other transformants of M. pilosus BCRC38072 were selected for Southern hybridization and fluorescence microscopy analyses. Their identities were also verified by transferring the stably grown colonies onto the new plates with hygromycin. The eight transformants were identified by Southern hybridization using HindIII and PvuII restriction
enzymes (Figure 4A, B). HindIII cuts at one site in the pMS-1.5hp plasmid, while PvuII does not cut within the plasmid. The results of Southern hybridization showed that the T5 transformant had a single copy integration, while T6 and T8 revealed the two integration events (two copies). Using the HindIII restriction enzyme, 6kb and 10kb fragments were observed in the transformants labeled as T1, T3 and T4, while 4kb and 6kb fragments corresponded to the transformants that were labeled as T2 and T7 (Figure 4A). However, only one fragment (>10 kb), corresponding to the transformants T1, T2, T3, T4 and T7, was detected when PvuII was used as the restriction enzyme (Figure 4B). These results suggested that the plasmid had integrated as a tandem repeat in the transformants T1, T2, T3, T4 and T7. They were attributed to the rearrangement of the integrated plasmid. Further studies on the loci of gene disruption and the mechanism of gene regulation will clarify the molecular mechanisms by which the biosynthesis of red pigment occurs. To confirm that the transformants expressed EGFP from the integrated DNA, the expression of EGFP was detected (Figure 4C, D) using a fluorescence microscopy. Fluorescence photomicrography of the specimens was conducted and fluorescence was clearly observed in the transformants when the samples were excited with a red filter. The red filter, with a peak transmission of excitation at 450 to 490 nm and a peak transmission of suppression at 525 to 575 nm, was used to detect EGFP. These results indicated that the EGFP protein was extensively expressed in the mycelia and spores. Moreover, the highest red pigment-producing mutation was stably inherited through the next generation after several rounds of cultivation.
In conclusion, this study demonstrates that the nuclease inhibitor aurintricarboxylic acid (ATA) can be adopted to stably transform M. pilosus. As was shown, the addition of ATA increased the efficiency of genetic transformation in M. pilosus. A major advantage of ATA is that different restriction enzymes need not be tested. Due to the fact that the transformant with high red pigment can be isolated by random mutagenesis with efficient genetic transformation using ATA,
this valid and convenient method could be used to screen the transformant with high monacolin K, as well as other high-economic polyketides.
References
(1) Babitha, S.; Soccol, C.R.; Pandey, A. Solid-state fermentation for the production of Monascus pigments from jackfruit seed. Bioresour. Technol.
2007a, 98, 1554-1560.
(2) Babitha, S.; Soccol, C.R.; Pandey, A. Effect of stress on growth, pigment production and morphology of Monascus sp. in solid cultures. J. Basic Microbiol. 2007b, 47, 118-126.
(3) Bingle, L.E.H.; Simpson, T.J.; Lazarus, C.M. Ketosynthase domain probes identify two subclasses of fungal polyketide synthase genes. Fungal Genet. Biol. 1999, 26, 209-223.
(4) Cantone, F.A.; Vandenberg, J.D. Genetic Transformation and Mutagenesis of the Entomopathogenic Fungus Paecilomyces fumosoroseus. J. Invertebr. Pathol. 1999, 74, 281-288.
(5) Chen, Y.-P.; Tseng, C.-P.; Liaw, L.-L.; Wang, C.-L.; Yuan, G.-F. Characterization of MRT, a New Non-LTR Retrotransposon in Monascus spp. Bot. Bull. Acad. Sin. 2007, 48, 377-385.
(6) Csermely, P.; Schnaider, T.; Sőti, C.; Prohászka, Z.; Nardai, G. The 90-kDa molecular chaperone family: structure, function, and clinical applications. A comprehensive review. Pharmacol. Ther. 1998, 79, 129-168.
(7) de Groot, M.J.; Bundock, P.; Hooykaas, P.J.; Beijersbergen, A.G. Agrobacterium tumefaciens-mediated transformation of filamentous fungi. Nat. Biotechnol. 1998, 16, 839-842.
(8) Endo, A. Monacolin K, a new hypocholesterolemic agent produced by a Monascus species. J. Antibiot. (Tokyo) 1979, 32, 852-854.
tissue transfection by aurintricarboxylic acid. Hum. Gene. Ther. 1999, 10, 1703-1713.
(10) Glasspool-Malone, J.; Malone, R.W. Enhancing direct in vivo transfection with nuclease inibitors and pulsed electrical fields. Methods Enzymol. 2002, 346, 72-91.
(11) Glasspool-Malone, J.; Somiari, S.; Drabick, J.J.; Malone, R.W. Efficient nonviral cutaneous transfection. Mol. Ther. 2000, 2, 140-146.
(12) Glasspool-Malone, J.; Steenland, P.R.; McDonald, R.J.; Sanchez, R.A.; Watts, T.L.; Zabner, J.; Malone, R.W. DNA transfection of macaque and murine respiratory tissue is greatly enhanced by use of nuclease inhibitor. J. Gene. Med.
2002, 4, 323-332.
(13) Hajjaj, H.; Blanc, P.J.; Groussac, E.; Goma, G.; Uribelarrea, J.L.; Loubiere, P. Improvement of red pigment/citrinin production ratio as a function of environmental conditions by Monascus ruber. Biotechnol. Bioeng. 1999, 64, 497-501.
(14) Hallick, R.B.; Chelm, B.K.; Gray, P.W.; Orozco, E.M., Jr. Use of aurintricarboxylic acid as an inhibitor of nuclease during nucleic acid isolation. Nucleic Acids Res. 1977, 4, 3055-3064.
(15) Jin, X.; Mo, M.-H.; Wei, Z.; Huang, X.W.; Zhang, K.Q. Transformation and mutagenesis of the nematode-trapping fungus Monacrosporium sphaeroides by restriction enzyme-mediated integration (REMI). J. Microbiol. 2005, 43, 417-423.
(16) Kahmann, R.; Basse, C. REMI (Restriction Enzyme Mediated Integration) and its impact on the isolation of pathogenicity genes in fungi attacking plants. Eur. J. Plant Pathol. 1999, 105, 221-229.
monoïca to hygromycin-B resistance. Curr. Genet. 1997, 31, 272-275.
(18) Mullins, E.D.; Kang, S. Transformation: a tool for studying fungal pathogens of plants. Cell. Mol. Life Sci. 2001, 58, 2043-2052.
(19) Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T. Polyketide synthase gene responsible for citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453-3457.
(20) Shimizu, T.; Kinoshita, H.; Nihira, T. Development of transformation system in Monascus purpureus using an autonomous replication vector with aureobasidin A resistance gene. Biotechnol. Lett. 2006, 28, 115-120.
(21) Thon, M.R.; Nuckles, E.M.; Vaillancourt, L.J. Restriction enzyme-mediated integration used to produce pathogenicity mutants of Colletotrichum graminicola. Mol. Plant-Microbe Interact. 2000, 13, 1356-1365.
(22) Tobert, J.A. Lovastatin and beyond: the history of the HMG-CoA reductase inhibitors. Nat. Rev. Drug Discov. 2003, 2, 517-526.
(23) Vollmer, S.J.; Yanofsky, C. Efficient cloning of genes of Neurospora crassa. Proc. Natl. Acad. Sci. USA. 1986, 83, 4869-4873.
(24) Walther, W.; Stein, U.; Siegel, R.; Fichtner, I.; Schlag, P.M. Use of the nuclease inhibitor aurintricarboxylic acid (ATA) for improved non-viral intratumoral in vivo gene transfer by jet-injection. J. Gene Med. 2005, 7, 477-485.
Table 1. The number of transformants based on various transformation methods. The plasmid pMS-1.5hp was used for genetic transformation.
Transformation methods
REMI Experiment Linear plasmid
(non-REMI)a
Treated by ATA (1mM)b
FspI (10 units)c HpaI (10 units)d
M. pilosuse 17 ± 2 159 ± 24 48 ± 4 119 ± 11
a. The plasmid pMS-1.5hp was linearized by FspI restriction enzyme.
b. The FspI-linearized pMS-1.5hp was mixed with 1 mM ATA and protoplasts.
c. The FspI-linearized pMS-1.5hp was mixed with 10 units FspI restriction enzyme and protoplasts.
d. The HpaI-linearized pMS-1.5hp was mixed with 10 units HpaI restriction enzyme and protoplasts.
e. Number of transformants per 5 ug linear plasmid. The values were based on three determinations.
Figure 1. Morphology of M. pilosus. (A) Growth of wild-type M. pilosus in PDB medium was observed under microscope. (B) The mycelia of M. pilosus were digested by 2 mg Yatalase/ml, 2 mg lysing enzyme/ml and 2 μl β-glucuronidase/ml. They were converted into protoplasts after 4~5 hr digestion. The photographs were taken at a magnification of X1000.
Figure 2. Map and Stability of pMS-1.5hp Plasmid. (A) Map of pMS-1.5hp plasmid used to overexpress a fusion protein of the hygromycin B resistance gene (HPH) with enhanced green fluorescent protein (EGFP). The pMS-1.5hp plasmid was modified
from the pHygEGFP plasmid. The small black bar represents the probe for analyzing integrations by Southern hybridization. A time course of the stability of the pMS-1.5hp plasmid in the M. pilosus BCRC38072 lysate was analyzed by including total protein of 1 mg ml-1 (B) or 3.5 mg ml-1 (C). Five μg of the pMS-1.5hp plasmid was incubated at 37oC with the lysates for 5, 15, 30, 45, and 60 min, containing 0, 0.1, and 1.0 mm ATA. The plasmid DNA was separated on 1.0% agarose gel by electrophoresis. Lane 1, untreated control plasmid without lysate; lanes 2-6, plasmid DNA with lysate protein containing 0 mm ATA incubated for 5, 15, 30, 45, and 60 min; lanes 7-11, plasmid DNA with lysate protein containing 0.1 mm ATA incubated for 5, 15, 30, 45, and 60 min; lanes 12-16, plasmid DNA with lysate protein containing 1.0 mm ATA incubated for 5, 15, 30, 45, and 60 min.
Figure 3. Submerged cultures of the wild type BCRC38072 and the T7 transformant were incubated on a 200 rpm rotary shaker for 10 days at 25 oC: (△) dry cell mass of the wild type; (□) dry cell mass of the transformant; (▲) red pigment production of the wild type; (■) red pigment production of the transformant. The red pigment level was detected by measuring the absorbance at 500 nm using a spectrophotometer.
Figure 4. Analyses of Southern Hybridization and Fluorescence of M. pilosus transformants. Southern hybridization analyses of the genomes of eight transformants digested with HindIII (A) or PvuII (B) and hybridized with an hph probe. Lanes 1-8 show DNA from the transformants (T1-T8); lane 9 shows DNA from wild-type M. pilosus BCRC38072; lane 10 shows the plasmid, pMS-1.5hp. Southern hybridization was performed using a DIG labeling system. Enhanced green fluorescent protein (EGFP) was expressed in the transformed M. pilosus. The conidia (C) and hyphae (D) of the transformant were analyzed by fluorescence (right panel) and bright field microscopy (left panel). The small bars represent distances of 50 μm and 100 μm in the conidia (C) and hyphae (D) respectively. The photograph was taken at a magnification of 1000X with an exposure time of 1 to 2 s.