?用?值分?方法比較臺灣野生?與?近緣分?群之形態特徵關係
14
0
0
全文
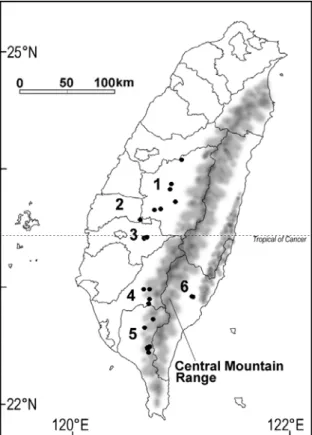
(2) March, 2007. Su et al.: Morphological comparisons of Taiwan wild tea plant and related taxa. 71. the same name, but he treated it as a synonym of C. sinensis. Although Suzuki (1937) had pointed that native wild tea plant was "Thea assamica affinis, sed foliis glabris" and Kitamura (1950) described that the forma was unique by 'Foliis majoribus angustioribus atroviridibus crassioribus', taxonomic uncertainties of this wild tea plant still remain. There had been several attempts to study C. sinensis f. formosensis based on numerical methods (Wu et al., 1970, 1972; Shih et al., 1972; Hu, 2004), however, all these studies aimed at exploring the variation among populations of C. sinensis f. formosensis, rather than solving the fundamental taxonomic problems. In the present study, the morphological variation among C. sinensis f. formosensis and its two related taxa, C. sinensis var. sinensis and C. sinensis var. assamica is summarized by applying multivariate numerical approaches. The aim is to detect infraspecific boundaries, to identify reliable distinguishing characters. A sound knowledge of taxonomy is a prerequisite for the success of any germplasm conservation program of the wild tea plant of Taiwan. MATERIALS AND METHODS Fig. 1. Distribution map of C. sinensis f. formosensis based on authors' samples and materials from TAI, TAIF and PPI herbaria. 1: Nantou County. 2: Yunlin County. 3: Chiayi County. 4: Kaohsiung County. 5: Pingtung County. 6: Taitung County.. several reports on tea improvements, the native wild tea plant was named as 'San-Cha' (means teas from mountains) to be distinguished from the imported tea plants (Shih, 1995; Li and Chang, 2003). Botanical nomenclatures of the native wild tea plant are somewhat complicated. Here we try to clarify it in brief. For its affinity to C. sinensis var. assamica, Masamune and Suzuki proposed Thea assamica Masters var. formosensis Masamune et Suzuki for the first time (Masamune, 1936). However, Masamune didn't publish the name validly, because there was no description provided. The correct name first published for this plant should be Thea formosensis Masamune et Suzuki (Suzuki, 1937). Later, this taxon was transferred as a forma under C. sinensis (Kitamura, 1950) and named as C. sinensis f. formosensis. In the following four decades, however, this name has been consistently neglected in all studies concerning the taxonomy of Camellia (Keng, 1950; Liu and Lu, 1967; Li, 1976; Ying, 1995) until the publication of Flora of Taiwan, second edition, in which Hsieh et al. (1996) followed Kitamura's treatment. Ming (2000) also mentioned. Materials In total, 165 specimens were selected for morphological study (Table 1). Sources of the specimens used in this investigation were deposited in the Herbarium of National Taiwan University (TAI), Herbarium of Taiwan Forest Research Institute (TAIF), Prof. Tzen-Yuh Chiang's Laboratory at the National Cheng Kung University (NCKU), and supplemented by the authors' own collection. Scientific names given on the specimen labels and annotated labels were tentatively used. Among 165 specimens, 72 were identified as C. sinensis var. sinensis, 41 as C. sinensis var. assamica and 52 as C. sinensis f. formosensis. Materials of C. sinensis f. formosensis were collected from all natural habitats we have known so far. Both C. sinensis var. sinensis and C. sinensis var. assamica are not native to Taiwan. In order to explore the variation of the two closely related taxa, several collections from China were made. Meanwhile, specimens from the Tea Research and Extension Station, which represented tea plants from India, Sri Lanka, Thailand and China, were included. Character measurement In this study, each collection was designated as an operational taxonomic unit (OTU). Specimen duplicates were treated as one OTU. Characters were.
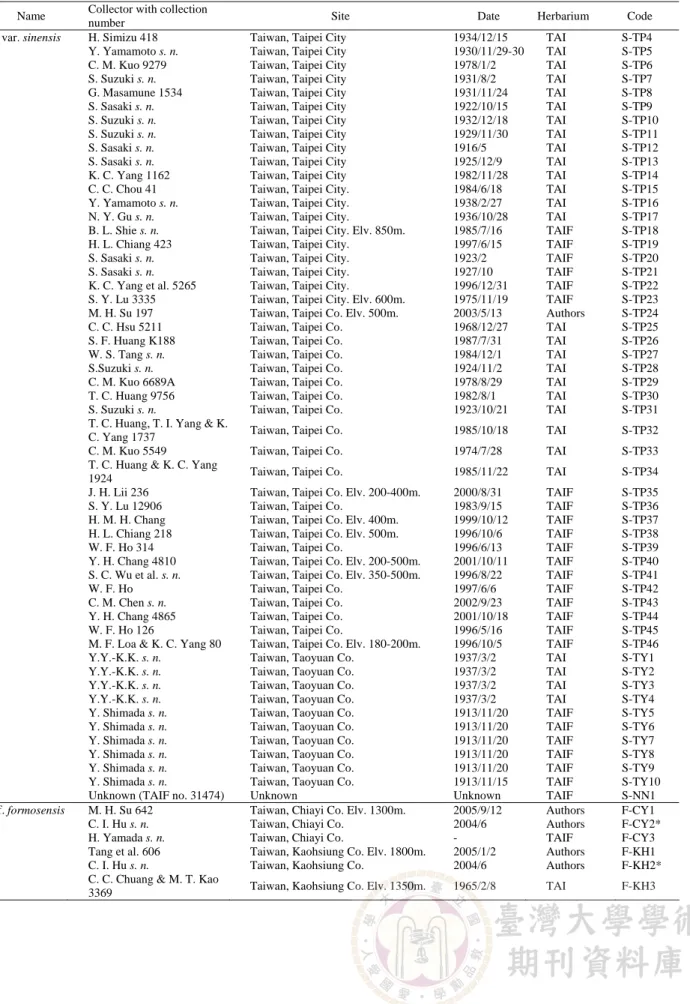
(3) 72. TAIWANIA. Vol. 52, No. 1. Table 1. List of voucher specimens included in the present study. Codes are comprised of an abbreviated scientific name, followed by a dash, and then the collection site with a serial number. Codes with an asterisk are the specimens collected from the germplasm banks of the Tea Research and Extension Station. TAI: Herbarium of National Taiwan University; TAIF: Herbarium of Taiwan Forest Research Institute; NCKU: Prof. Tzen-Yuh Chiang’s Laboratory at National Cheng Kung University. Name var. assamica. var. sinensis. Collector with collection number M. H. Su 669 K. S. Wang 5007 K. S. Wang 5010 K. S. Wang 5016 K. S. Wang 5017 K. S. Wang 5019 K. S. Wang 5021 K. S. Wang 5024 K. S. Wang 5026 K. S. Wang 5033 K. S. Wang 5034 K. S. Wang 5035 K. S. Wang 5036 K. S. Wang 5041 K. S. Wang 5042 K. S. Wang 5052 K. S. Wang 5053 K. S. Wang 5054 K. S. Wang 5058 K. S. Wang 5066 K. S. Wang 5072 K. S. Wang 5076 K. S. Wang 5079 K. S. Wang 5084 K. S. Wang 5091 M. H. Su 670 M. H. Su 667 M. H. Su 685 M. H. Su 610 M. H. Su 609 M. H. Su 608 M. T. Kao s. n. S. Hibino & S. Suzuki s. n. H. Keng, T. S. Liu & M. T. Kao s. n. S. Y. Lu 18521 S. Y. Lu 18291 T. S. Liu & H. Keng 2853 Y.Y.-K.K. s. n. Y.Y.-K.K. s. n. Y.Y.-K.K. s. n. M. H. Su 684 C. H. Tsou 2117 C. H. Tsou et al. 1957 M. S. An 3357 K. F. Wang 1-0560 J. H. Hu 282 J. H. Hu 242 T. M. Taing 00748 Z. Chen 09769 T. Makino s. n. M. H. Su 640 M. H. Su 644 M. H. Su 95 Y. Shimada s. n. A. T. Hsieh s. n. S. W. Chung 7520 S. P. Chien s. n. C. S. Kuoh 2952 C. M. Kuo 5457. Site Taiwan, Taipei Co. (Burma type) China, Yunan Prov. Elv. 1781m China, Yunan Prov. Elv. 970m China, Yunan Prov. China, Yunan Prov. China, Yunan Prov. China, Yunan Prov. China, Yunan Prov. China, Yunan Prov. China, Yunan Prov. China, Yunan Prov. Elv. 1200m. China, Yunan Prov. Elv. 1200m. China, Yunan Prov. Elv. 1200m. China, Yunan Prov. Elv. 1900m. China, Yunan Prov. Elv. 1900m. China, Yunan Prov. Elv. 1950m. China, Yunan Prov. China, Yunan Prov. Elv. 1470m. China, Yunan Prov. Elv. 1300m. China, Yunan Prov. Elv. 1020m. China, Yunan Prov. Elv. 1400m. China, Yunan Prov. Elv. 1380m. China, Yunan Prov. Elv. 1000m. China, Yunan Prov. Elv. 900m. China, Yunan Prov. Elv. 900m. Taiwan, Taipei Co. (Assam type) Taiwan, Taipei Co. (Manipur type) Taiwan, Nantou Co. (Sri Lanka type) Taiwan, Nantou Co. Elv. 1000m. Taiwan, Nantou Co. Elv. 1000m. Taiwan, Nantou Co. Elv. 1000m. Taiwan, Nantou Co. Taiwan, Nantou Co.. Date. Herbarium. Code. 2005/10/6 2004/12/1 2004/12/1 2004/12/1 2004/12/1 2004/12/1 2004/12/2 2004/12/2 2004/12/2 2004/12/2 2004/12/3 2004/12/3 2004/12/3 2004/12/3 2004/12/3 2004/12/4 2004/12/4 2004/12/4 2004/12/4 2004/12/4 2004/12/5 2004/12/5 2004/12/5 2004/12/5 2004/12/5 2005/10/6 2005/10/6 2005/11/15 2005/2/1 2005/2/1 2005/2/1 1955/2/12 1926/7/17. Authors NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU NCKU Authors Authors Authors Authors Authors Authors TAI TAI. A-BU1* A-CN1 A-CN2 A-CN3 A-CN4 A-CN5 A-CN6 A-CN7 A-CN8 A-CN9 A-CN10 A-CN11 A-CN12 A-CN13 A-CN14 A-CN15 A-CN16 A-CN17 A-CN18 A-CN19 A-CN20 A-CN21 A-CN22 A-CN23 A-CN24 A-IN1* A-IN2* A-SR1* A-NT1 A-NT2 A-NT3 A-NT4 A-NT5. Taiwan, Nantou Co.. 1955/7/20. TAI. A-NT6. Taiwan, Nantou Co. Elv. 400m. Taiwan, Nantou Co. Elv. 700m. Taiwan, Taitung Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taipei Co. (Thailand type) China, Fujian Prov. Elv. 500-1200m. China, Guangdong Prov. China, Guizhou Prov. Elv. 920m. China, Guizhou Prov. Elv. 650m. China, Hunan Prov. Elv. 500m. China, Hunan Prov. Elv. 880m. China, Jiangxi Prov. 180m. China, Sichuan Prov. Elv. 810m Japan, Tokyo City. Taiwan, Chiayi Co. Elv. 1100m. Taiwan, Chiayi Co. Elv. 800m. Taiwan, Hsinchu Co. Taiwan, Hsinchu Co. Taiwan, Miaoli Co. Taiwan, Nantou Co. Elv. 700-800m. Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City. 1986/3/2 1986/2/7 1955/8/10 1937/3/2 1937/3/2 1937/3/2 2005/11/15 2004/5/12 2004/8/6 2003/5/26 2003/7/15 2001/8/19 2002/6/2 2000/10/22 1996/10/5 1910/10 2005/9/12 2005/9/13 2003/5/8 1913/11/15 1929/12/14 2004/6/10 1984/6/18 1971/11/23 1974/7/6. TAIF TAIF TAI TAI TAI TAI Authors Authors Authors TAIF TAIF TAIF TAIF TAIF TAIF TAIF Authors Authors Authors TAIF TAIF TAIF TAI TAI TAI. A-NT7 A-NT8 A-TT1 A-TY1 A-TY2 A-TY3 A-TH1* S-CN1 S-CN2 S-CN3 S-CN4 S-CN5 S-CN6 S-CN7 S-CN8 S-JP1 S-CY1 S-CY2 S-HC1 S-HC2 S-ML1 S-NT1 S-TP1 S-TP2 S-TP3.
(4) March, 2007. Su et al.: Morphological comparisons of Taiwan wild tea plant and related taxa. 73. Table 1. Continued. Name var. sinensis. f. formosensis. Collector with collection number H. Simizu 418 Y. Yamamoto s. n. C. M. Kuo 9279 S. Suzuki s. n. G. Masamune 1534 S. Sasaki s. n. S. Suzuki s. n. S. Suzuki s. n. S. Sasaki s. n. S. Sasaki s. n. K. C. Yang 1162 C. C. Chou 41 Y. Yamamoto s. n. N. Y. Gu s. n. B. L. Shie s. n. H. L. Chiang 423 S. Sasaki s. n. S. Sasaki s. n. K. C. Yang et al. 5265 S. Y. Lu 3335 M. H. Su 197 C. C. Hsu 5211 S. F. Huang K188 W. S. Tang s. n. S.Suzuki s. n. C. M. Kuo 6689A T. C. Huang 9756 S. Suzuki s. n. T. C. Huang, T. I. Yang & K. C. Yang 1737 C. M. Kuo 5549 T. C. Huang & K. C. Yang 1924 J. H. Lii 236 S. Y. Lu 12906 H. M. H. Chang H. L. Chiang 218 W. F. Ho 314 Y. H. Chang 4810 S. C. Wu et al. s. n. W. F. Ho C. M. Chen s. n. Y. H. Chang 4865 W. F. Ho 126 M. F. Loa & K. C. Yang 80 Y.Y.-K.K. s. n. Y.Y.-K.K. s. n. Y.Y.-K.K. s. n. Y.Y.-K.K. s. n. Y. Shimada s. n. Y. Shimada s. n. Y. Shimada s. n. Y. Shimada s. n. Y. Shimada s. n. Y. Shimada s. n. Unknown (TAIF no. 31474) M. H. Su 642 C. I. Hu s. n. H. Yamada s. n. Tang et al. 606 C. I. Hu s. n. C. C. Chuang & M. T. Kao 3369. Site. Date. Herbarium. Code. Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City Taiwan, Taipei City. Taiwan, Taipei City. Taiwan, Taipei City. Taiwan, Taipei City. Elv. 850m. Taiwan, Taipei City. Taiwan, Taipei City. Taiwan, Taipei City. Taiwan, Taipei City. Taiwan, Taipei City. Elv. 600m. Taiwan, Taipei Co. Elv. 500m. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co.. 1934/12/15 1930/11/29-30 1978/1/2 1931/8/2 1931/11/24 1922/10/15 1932/12/18 1929/11/30 1916/5 1925/12/9 1982/11/28 1984/6/18 1938/2/27 1936/10/28 1985/7/16 1997/6/15 1923/2 1927/10 1996/12/31 1975/11/19 2003/5/13 1968/12/27 1987/7/31 1984/12/1 1924/11/2 1978/8/29 1982/8/1 1923/10/21. TAI TAI TAI TAI TAI TAI TAI TAI TAI TAI TAI TAI TAI TAI TAIF TAIF TAIF TAIF TAIF TAIF Authors TAI TAI TAI TAI TAI TAI TAI. S-TP4 S-TP5 S-TP6 S-TP7 S-TP8 S-TP9 S-TP10 S-TP11 S-TP12 S-TP13 S-TP14 S-TP15 S-TP16 S-TP17 S-TP18 S-TP19 S-TP20 S-TP21 S-TP22 S-TP23 S-TP24 S-TP25 S-TP26 S-TP27 S-TP28 S-TP29 S-TP30 S-TP31. Taiwan, Taipei Co.. 1985/10/18. TAI. S-TP32. Taiwan, Taipei Co.. 1974/7/28. TAI. S-TP33. Taiwan, Taipei Co.. 1985/11/22. TAI. S-TP34. Taiwan, Taipei Co. Elv. 200-400m. Taiwan, Taipei Co. Taiwan, Taipei Co. Elv. 400m. Taiwan, Taipei Co. Elv. 500m. Taiwan, Taipei Co. Taiwan, Taipei Co. Elv. 200-500m. Taiwan, Taipei Co. Elv. 350-500m. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co. Taiwan, Taipei Co. Elv. 180-200m. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Taiwan, Taoyuan Co. Unknown Taiwan, Chiayi Co. Elv. 1300m. Taiwan, Chiayi Co. Taiwan, Chiayi Co. Taiwan, Kaohsiung Co. Elv. 1800m. Taiwan, Kaohsiung Co.. 2000/8/31 1983/9/15 1999/10/12 1996/10/6 1996/6/13 2001/10/11 1996/8/22 1997/6/6 2002/9/23 2001/10/18 1996/5/16 1996/10/5 1937/3/2 1937/3/2 1937/3/2 1937/3/2 1913/11/20 1913/11/20 1913/11/20 1913/11/20 1913/11/20 1913/11/15 Unknown 2005/9/12 2004/6 2005/1/2 2004/6. TAIF TAIF TAIF TAIF TAIF TAIF TAIF TAIF TAIF TAIF TAIF TAIF TAI TAI TAI TAI TAIF TAIF TAIF TAIF TAIF TAIF TAIF Authors Authors TAIF Authors Authors. S-TP35 S-TP36 S-TP37 S-TP38 S-TP39 S-TP40 S-TP41 S-TP42 S-TP43 S-TP44 S-TP45 S-TP46 S-TY1 S-TY2 S-TY3 S-TY4 S-TY5 S-TY6 S-TY7 S-TY8 S-TY9 S-TY10 S-NN1 F-CY1 F-CY2* F-CY3 F-KH1 F-KH2*. Taiwan, Kaohsiung Co. Elv. 1350m.. 1965/2/8. TAI. F-KH3.
(5) 74. TAIWANIA. Vol. 52, No. 1. Table 1. Continued. Name f. formosensis. Collector with collection number T. Kiang & M. T. Kao KT439 M. T. Kao 7448 T. C. Huang 4952 A. Tanimura s. n. S. Sasaki s. n. S. Y. Lu 18256 S. Y. Lu 18664 Y. H. Lai 83 C. P. Lin s. n. C. H. Tsou 2134 C. H. Tsou 2132 C. H. Tsou 2137 C. H. Tsou 2139 M. H. Su 687 M. H. Su 683 M. Hasimoto s. n. S. Sasaki s. n. S. Taniguchi s. n. S. Suzuki 3245 S. Sasaki s. n. M. Hasimoto s. n. S. Sasaki s. n. M. T. Kao 6668 B. J. Wang 15069 S. Sasaki s. n. M. H. Su 575 M. H. Su 269 M. H. Su 270 M. H. Su 497 M. H. Su 498 M. H. Su 646 M. H. Su 645 M. H. Su 544 M. H. Su 545 E. Matuda s. n. E. Matuda K. C. Yang et al. 4583 K. C. Yang et al. 4527 S. W. Chung 7090 M. H. Su 659 M. H. Su 661 M. H. Su 660 M. H. Su 655 M. T. Kao 6612 K. Mori 1901 Tanimura s. n.. Site. Date. Herbarium. Code. Taiwan, Kaohsiung Co.. 1971/5/12. TAI. F-KH4. Taiwan, Kaohsiung Co. Taiwan, Kaohsiung Co. Taiwan, Kaohsiung Co. Taiwan, Kaohsiung Co. Taiwan, Kaohsiung Co. Elv. 1150m. Taiwan, Kaohsiung Co. Elv. 1500m. Taiwan, Kaohsiung Co. Elv. 750-800m. Taiwan, Kaohsiung Co. Elv. 650m. Taiwan, Nantou Co. Elv. 1200m. Taiwan, Nantou Co. Elv. 1200m. Taiwan, Nantou Co. Elv. 1200m. Taiwan, Nantou Co. Elv. 1200m. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Nantou Co. Taiwan, Pingtung Co. Elv. 1400m. Taiwan, Pingtung Co. Taiwan, Pingtung Co. Elv. 1300m. Taiwan, Pingtung Co. Elv. 1200m. Taiwan, Pingtung Co. Elv. 1200m. Taiwan, Pingtung Co. Elv. 1100m. Taiwan, Pingtung Co. Elv. 1100m. Taiwan, Pingtung Co. Elv. 1100m. Taiwan, Pingtung Co. Elv. 1100m. Taiwan, Pingtung Co. Taiwan, Pingtung Co. Taiwan, Pingtung Co. Elv. 850m. Taiwan, Pingtung Co. Elv. 750-1100m. Taiwan, Pingtung Co. Elv. 800-1000m. Taiwan, Taitung Co. Elv. 1000m. Taiwan, Taitung Co. Elv. 1100m. Taiwan, Taitung Co. Elv. 1100m. Taiwan, Taitung Co. Elv. 1100m. Taiwan, Taoyuan Co. Taiwan, Yunlin Co. Unknown. 1968/12/10 1968/12/10 1935/1/12 1936/3/8 1986/1/30 1986/3/11. TAI TAI TAI TAI TAIF TAIF. F-KH5 F-KH6 F-KH7 F-NN8 F-KH9 F-KH10. 1996/12/2. TAIF. F-KH11. 2004/5/25 2005/3/30 2005/3/30 2005/3/30 2005/3/30 2005/11/15 2005/11/15 1966/1/13 1935/11/8 1931/7/12 1935/11/8 1935/11/8 1966/1/16 1935/10/8 1966/4/23 1988/12/25 1922/11/30 2004/4/14 2003/9/20 2003/9/20 2004/1/24 2004/1/24 2005/9/27 2005/9/27 2004/3/10 2004/3/10 1919/7/11 1912/11/7 1995/12/2. TAIF Authors Authors Authors Authors Authors Authors TAI TAI TAI TAI TAI TAI TAI TAI TAIF TAIF Authors Authors Authors Authors Authors Authors Authors Authors Authors TAI TAI TAIF. F-KH12 F-NT1 F-NT2 F-NT3 F-NT4 F-NT5 F-NT6 F-NT7 F-NT8 F-NT9 F-NT10 F-NT11 F-NT12 F-NT13 F-NT14 F-NT15 F-NT16 F-PT1 F-PT2 F-PT3 F-PT4 F-PT5 F-PT6 F-PT7 F-PT8 F-PT9 F-PT10 F-PT11 F-PT12. 1995/12/3. TAIF. F-PT13. 2004/5/29. TAIF. F-PT14. 2005/9/28 2005/9/28 2005/9/28 2005/9/28 1966/1/11 1906/11/5 1935/1/12. Authors Authors Authors Authors TAI TAIF TAI. F-TT1 F-TT2 F-TT3 F-TT4 F-TY1 F-YL1 F-NN1. chosen with respect to variation among taxa mentioned in literature and based on personal observations on specimens. Finally, a total of 35 characters were scored, including 17 vegetative and 18 floral characters (Table 2). For each specimen, five mature, healthy-look leaves were scored and averaged. The measurement on floral characters was averaged from one to three flowers, depending on the condition of the specimens. We also measured the angles between the midrib and one major lateral vein. at two different positions (Fig. 2), because the usual curved lateral veins cannot be expressed by a single value (often measured at the base in most studies). Upon further examination, it was found that one vegetative and seven floral characters were constant (not informative) and should be eliminated from the following analyses. Finally, 27 (16 vegetative and 11 floral) characters were selected. Since not all specimens were in the flowering stage, the data for the numerical analyses were divided into three.
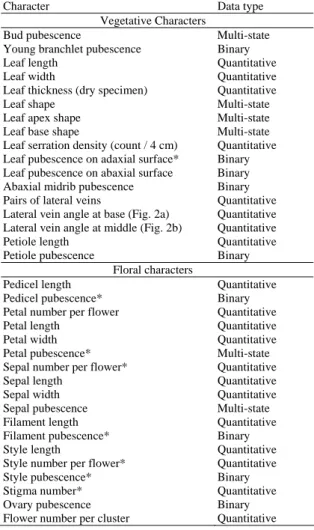
(6) March, 2007. Su et al.: Morphological comparisons of Taiwan wild tea plant and related taxa. 75. Table 2. List of morphological characters examined for multivariate analyses. The asterisk (*) denotes constant characters that are excluded from the data analysis. The angles measured between midrib and lateral vein are shown in Fig. 2. Character. Data type Vegetative Characters. Bud pubescence Young branchlet pubescence Leaf length Leaf width Leaf thickness (dry specimen) Leaf shape Leaf apex shape Leaf base shape Leaf serration density (count / 4 cm) Leaf pubescence on adaxial surface* Leaf pubescence on abaxial surface Abaxial midrib pubescence Pairs of lateral veins Lateral vein angle at base (Fig. 2a) Lateral vein angle at middle (Fig. 2b) Petiole length Petiole pubescence Floral characters Pedicel length Pedicel pubescence* Petal number per flower Petal length Petal width Petal pubescence* Sepal number per flower* Sepal length Sepal width Sepal pubescence Filament length Filament pubescence* Style length Style number per flower* Style pubescence* Stigma number* Ovary pubescence Flower number per cluster. Multi-state Binary Quantitative Quantitative Quantitative Multi-state Multi-state Multi-state Quantitative Binary Binary Binary Quantitative Quantitative Quantitative Quantitative Binary Quantitative Binary Quantitative Quantitative Quantitative Multi-state Quantitative Quantitative Quantitative Multi-state Quantitative Binary Quantitative Quantitative Binary Quantitative Binary Quantitative. categories: (1) only vegetative characters (165 OTUs × 16 characters); (2) only floral characters (62 OTUs × 11 characters); and (3) all characters (62 OTUs × 27 characters). Cluster analysis Similarity matrices were generated using the coefficient proposed by Gower (1971). Gower's similarity coefficient (GSC) was designed to deal with mixed type of characters, and was thus widely used (Schultze-Motel and Meyer, 1981; Zaharof, 1988; Cheng, 1990; Ward, 1993; Gugerli, 1997; St-Laurent et al., 2000; Muvaffak et al., 2001; Binns et al., 2002; Bayly et al., 2003; Mckenzie et al., 2004). This similarity matrix was then used to perform a cluster analysis using the Unweighted Pair Grouping Method Based on Arithmetic Averages (UPGMA) (Sokal and Michener, 1958) with the software MVSP v3.01 (Kovach Computing Service, 1999).. b a Fig. 2. Scheme of a leaf showing the angles (a) formed between the midrib and a lateral vein, and (b) created by the midrib and the interception of the tangent to the middle portion of a lateral vein.. Nonlinear principal component analysis To further explore the pattern of variation in measured characters and to find those characters which are decisive to distinguish taxa, a nonlinear principal components analysis (NLPCA, de Leeuw, 1982) was undertaken. Similar to principal components analysis, NLPCA can be used for transforming attributes of a dataset into a new set of uncorrelated attributes (principal components), while still retaining as much of the variability of the dataset as possible. It can handle variables of different types (nominal, ordinal and numerical) simultaneously, and deal with nonlinear relationships between variables. NLPCA is performed by the program CATPCA implemented in the software SPSS v13.0 (SPSS Inc.). In addition, Cronbach's Alpha (Cronbach, 1951) was calculated for each of the components extracted. If Alpha value of a specific component is high, it would be interpreted as indicating that the component has a strong one-dimensional structure, or, the dimension is reliable to account for the total variance. Generally, an Alpha value of 0.70 or greater is considered to be reliable (Bland and Altman, 1997).. RESULTS Vegetative characters The UPGMA phenogram based on vegetative characters showed two discrete clusters (Fig. 3), namely Group I-1 (GSC = 0.62) and Group I-2 (GSC = 0.67). Group I-1 was composed entirely of C. sinensis f. formosensis from central and southern Taiwan. Within this cluster, there did not appear to be any regional patterns. Group I-2, however, contained all the samples of C. sinensis var. sinensis and C. sinensis var. assamica, with C. sinensis f. formosensis from eastern Taiwan. Despite of that, samples of eastern C. sinensis f. formosensis formed a consistent subgroup (Group I-2-1; GSC = 0.87) within Group I-2. In contrast, all samples of C. sinensis var. sinensis and C. sinensis var. assamica overlapped extensively and together they formed a large subgroup (Group I-2-2; GSC = 0.71)..
(7) 76. TAIWANIA. F-PT12 F-PT13 F-PT6 F-NT12 F-NT3 F-NT2 F-NN1 F-KH12 F-NT9 F-NT7 F-KH10 F-NT16 F-KH7 F-NT8 F-KH9 F-KH6 F-PT10 F-NT15 F-NT11 F-NN8 F-PT4 F-KH3 F-PT7 F-PT5 F-TY1 F-KH5 F-KH2 F-PT2 F-PT9 F-KH1 F-YL1 F-NT4 F-NT1 F-NT14 F-NT13 F-PT14 F-KH4 F-NT10 F-CY3 F-PT11 F-PT8 F-PT3 F-PT1 F-KH11 F-NT5 F-NT6 F-CY2 F-CY1 F-TT2 F-TT3 F-TT4 F-TT1 A-TH1 A-NT3 A-CN18 S-TP16 A-NT7 S-TP40 S-TY4 S-TP11 S-TP10 S-TP8 S-TY3 S-CN5 S-TP32 S-TP15 S-CN2 A-NT4 A-TT1 A-TY3 A-TY2 A-CN20 A-NT5 A-CN3 S-TP26 S-TP12 S-TP28 S-TP24 S-HC1 S-TP22 S-TP7 S-TP18 S-CY2 A-SR1 A-CN7 S-TY7 S-CN3 A-CN5 A-NT1 A-IN2 A-CN19 A-TY1 A-NT8 A-CN24 A-CN13 A-CN16 A-CN2 S-TP37 S-NN1 S-TP33 S-TP31 S-TY5 S-TP19 S-TP5 A-CN22 S-TP39 S-TP36 S-CY1 S-TY6 S-HC2 S-TP42 S-TY9 A-CN8 S-TP41 S-TP29 S-TP17 S-TP2 A-CN23 S-TP3 S-CN8 S-CN6 S-CN4 A-CN9 A-CN10 A-CN11 A-CN1 S-TY8 S-TY10 S-TP38 S-TP21 S-TP35 S-TP20 S-TP45 S-TP14 S-TP43 S-NT1 S-TP30 S-TP1 A-NT6 S-TP44 S-TP46 A-NT2 S-TY1 S-TP25 S-TP27 S-TP13 S-TP4 S-TP23 S-CN7 A-CN15 S-TP9 A-CN17 A-CN21 A-CN14 A-CN12 S-TP6 S-TP34 S-ML1 A-CN6 S-TY2 S-JP1 A-CN4 S-CN1 A-IN1 A-BU1. 0.4. 0.5. 0.6. 0.7. 0.8. 0.9. Vol. 52, No. 1. Table 3. Loadings of the 16 vegetative characters on the first three components from NLPCA. Eigenvalues, percentage of variance explained and cumulated, and Cronbach's Alpha are given for each component. I-1. Character Bud pubescence Young branchlet pubescence Leaf length Leaf width Leaf thickness Leaf shape Leaf apex shape Leaf base shape Leaf serration density Leaf pubescence on abaxial surface Abaxial midrib pubescence Pairs of lateral veins Lateral vein angle at base Lateral vein angle at middle Petiole length Petiole pubescence Eigenvalue Variance explained (%) Variance cumulative (%) Cronbach's Alpha. I-2-1. I-2-2 I-2. Component 2 0.29 -0.32 -0.55 -0.71 -0.10 0.10 -0.02 -0.02 0.73 -0.23 -0.32 -0.27 0.16 0.13 -0.35 -0.30 2.03 12.70 55.90 0.54. 3 -0.05 0.03 -0.09 0.17 0.25 0.47 -0.26 0.60 0.03 0.36 0.06 0.13 0.64 0.41 -0.18 0.04 1.51 9.40 65.30 0.36. 3. 2. 1. Fig. 3. Phenogram of UPGMA cluster analysis based on vegetative characters. The coefficient was defined as the Gower's similarity coefficient.. 1. Component 2. The results of NLPCA on the vegetative characters are presented in Table 3. The first three components accounted for 65.3 % of the total variance in the dataset. The first component alone accounted for 43.2% of the total variance and was far more important than other components. The first component with a Cronbach's alpha value of 0.91 was the only one considered to be reliable. The component loadings correspond to the correlation coefficients between characters and the derived components. Characters with high loadings were bud pubescence, young branchlet pubescence, abaxial midrib pubescence and petiole pubescence (over 0.9), followed by leaf length, leaf apex shape and pairs of lateral veins (over 0.7). The plot by the first two components (Fig. 4) shows a similar grouping with the cluster analysis. However, the eastern samples of C. sinensis f. formosensis were placed onto the intermediate positions in NLPCA. It didn't group these eastern samples with C. sinensis var. sinensis and C. sinensis var. assamica absolutely.. 1 -0.93 0.91 -0.77 -0.55 0.13 -0.33 -0.74 0.14 0.34 0.67 0.90 -0.79 -0.39 -0.64 -0.50 0.91 6.90 43.20 43.20 0.91. 0. -1. -2. -3 -2. -1. 0. 1. 2. Component 1. Fig. 4. Ordination plot of NLPCA based on vegetative characters. : central and southern C. sinensis f. formosensis. : eastern C. sinensis f. formosensis. {: C. sinensis var. assamica. c: C. sinensis var. sinensis..
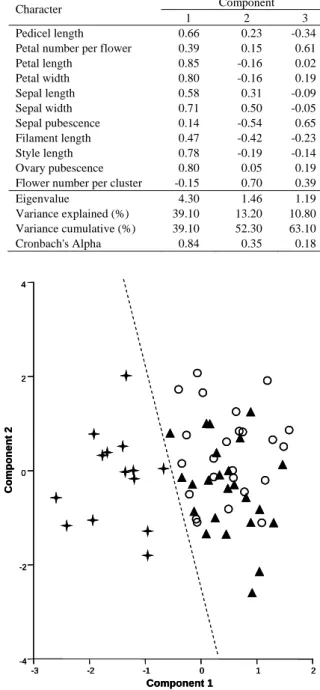
(8) Su et al.: Morphological comparisons of Taiwan wild tea plant and related taxa. Floral characters Cluster analysis of OTUs based on floral characters revealed two main groups (i.e. Group II-1 and Group II-2; Fig. 5), similar to the result obtained based on vegetative characters (Fig. 3). However, the members within each cluster were different. The OTUs of C. sinensis f. formosensis from eastern Taiwan, previously clustered within Group I-2, are now located amongst Group II-1. Consequently, Group II-1 encompassed all samples of C. sinensis f. formosensis, while Group II-2 consisted of a mix of C. sinensis var. sinensis and C. sinensis var. assamica. A further subdivision of the two main groups into geographical or taxonomic subgroups could not be made. The results of NLPCA on floral characters are shown in Table 4. The first three principal components accounted for 39.1%, 13.2% and 10.8% of the total variance, respectively. Only the first principal component was meaningful (Cronbach's alpha = 0.84) for grouping samples discretely. Characters with high loadings on the first principal component were petal length, petal width and ovary pubescence (over 0.8), followed by sepal width and style length (over 0.7). C. sinensis f. formosensis also can be separated by a line on the plot of the first two components (Fig. 6), a result just the same with that of the cluster analysis. F-NT11 F-NT6 F-CY2 F-TT3 F-PT12 F-KH1 F-PT2 F-KH11 F-PT13 F-TT4 F-PT7 F-PT6 F-NT15 F-CY1 A-IN1 A-CN13 A-CN7 S-TY2 A-CN24 A-CN4 S-CN8 S-TP32 S-TP17 A-CN15 S-TP25 A-CN10 S-TP20 S-TP27 A-CN14 S-TP11 A-CN23 A-CN6 A-CN6 A-CN22 A-CN21 A-CN9 S-TY8 S-TP8 A-CN18 A-CN3 A-CN11 A-CN19 A-CN12 A-CN8 S-ML1 S-NN1 S-TP5 A-CN17 S-TP13 S-TP13 A-CN2 S-TP44 S-TP31 S-TP10 S-TP2 A-NT8 S-TP37 S-TP38 S-TP22 S-TP4 S-CY1 A-NT1 S-TP21 A-BU1 0.6. 0.68. 0.76. 0.84. 0.92. II-1. 77. Table 4. Loadings of the 11 floral characters on the first three components from NLPCA. Eigenvalues, percentage of variance explained and cumulated, and Cronbach's Alpha are given for each component. Character. Component 2 0.23 0.15 -0.16 -0.16 0.31 0.50 -0.54 -0.42 -0.19 0.05 0.70 1.46 13.20 52.30 0.35. 1 0.66 0.39 0.85 0.80 0.58 0.71 0.14 0.47 0.78 0.80 -0.15 4.30 39.10 39.10 0.84. Pedicel length Petal number per flower Petal length Petal width Sepal length Sepal width Sepal pubescence Filament length Style length Ovary pubescence Flower number per cluster Eigenvalue Variance explained (%) Variance cumulative (%) Cronbach's Alpha. 3 -0.34 0.61 0.02 0.19 -0.09 -0.05 0.65 -0.23 -0.14 0.19 0.39 1.19 10.80 63.10 0.18. 4. 2. Component 2. March, 2007. 0. -2. II-2. 1. Fig. 5. Phenogram of UPGMA cluster analysis based on floral characters. The coefficient was defined as the Gower's similarity coefficient.. -4 -3. -2. -1. 0. 1. 2. Component 1. Fig. 6. Ordination plot of NLPCA based on floral characters. : C. sinensis f. formosensis. {: C. sinensis var. assamica. c: C. sinensis var. sinensis.. All characters Cluster analysis of all the floral and vegetative characters produced a phenogram with two groups of OTUs (Group III-1 and Group III-2, Fig. 7) that corresponded to the separation based on floral characters. OTUs of C. sinensis f. formosensis were.
(9) 78. TAIWANIA. F-PT12 F-TT4 F-TT3 F-PT13 F-PT6 F-PT7 F-NT15 F-NT11 F-KH11 F-PT2 F-KH1 F-NT6 F-CY2 F-CY1 A-CN18 A-CN3 S-TP11 S-TP10 A-IN1 A-CN7 S-TP32 S-TY2 A-CN4 S-TY8 S-TP8 S-TP25 A-CN15 A-CN21 A-CN14 A-CN17 A-CN12 S-TP20 S-TP20 S-TP27 S-TP13 S-ML1 A-CN6 S-NN1 S-TP5 S-CN8 A-CN22 A-CN9 S-TP17 A-CN8 A-CN19 A-CN13 A-CN23 A-CN10 A-CN24 A-CN24 A-CN11 A-CN2 S-TP31 S-TP44 S-TP37 S-TP22 S-TP38 S-TP4 A-NT8 S-TP2 S-CY1 A-NT1 S-TP21 A-BU1 0.52. 0.6. 0.68. 0.76. 0.84. 0.92. III-1. Vol. 52, No. 1. Table 5. Loadings of the 27 vegatative and floral characters on the first three components from NLPCA. Eigenvalues, percentage of variance explained and cumulated, and Cronbach's Alpha are given for each component. Character. III-2. 1. Fig. 7. Phenogram of UPGMA cluster analysis based on vegetative and floral characters. The coefficient was defined as the Gower's similarity coefficient.. contained entirely within Group III-1, while C. sinensis var. sinensis and C. sinensis var. assamica were dispersed throughout Group III-2. The NLPCA character loadings, percentage, and variance explained and cumulated for the first three components are given in Table 5. The first component accounted for 36.8% of the total variance observed, and was highly interpretable (Cronbach's alpha = 0.93). It had high contributing component loadings from bud pubescence, young branchlet pubescence, abaxial midrib pubescence, petiole pubescence and ovary pubescence (over 0.88), and leaf length, leaf apex shape, leaf pubescence on abaxial surface, pairs of lateral veins, petal length, petal width and sepal width (over 0.65). Figure 8 shows that the first principal component effectively separates C. sinensis f. formosensis from C. sinensis var. sinensis and C. sinensis var. assamica. The second component was just on the level of reliability (Cronbach's alpha = 0.71), with negative loading on leaf width, and positive loading on leaf serration density. The results of this study also showed that characters with high component loadings based on all characters were in agreement with those only based on vegetative or floral characters. The plot of the first two components based on all characters (Fig. 8) shows a clearer separation of C. sinensis f. formosensis from the other. Bud pubescence Young branchlet pubescence Leaf length Leaf width Leaf thickness Leaf shape Leaf apex shape Leaf base shape Leaf serration density Leaf pubescence on abaxial surface Abaxial midrib pubescence Pairs of lateral veins Lateral vein angle at base Lateral vein angle at middle Petiole length Petiole pubescence Pedicel length Petal number per flower Petal length Petal width Sepal length Sepal width Sepal pubescence Filament length Style length Ovary pubescence Flower number per cluster Eigenvalue Variance explained (%) Variance cumulative (%) Cronbach's Alpha. 1 -0.95 0.89 -0.67 -0.46 0.15 -0.55 -0.65 -0.26 0.26 0.73 0.89 -0.70 -0.25 -0.52 -0.48 0.89 0.59 0.27 0.75 0.71 0.49 0.66 -0.20 0.29 0.63 0.91 -0.21 9.95 36.80 36.80 0.93. Component 2 3 0.19 0.10 -0.21 -0.15 -0.62 -0.08 -0.81 0.05 -0.18 -0.38 -0.05 -0.13 -0.03 -0.06 -0.16 -0.20 0.71 0.11 -0.40 0.03 -0.21 -0.15 -0.49 0.07 -0.25 0.69 -0.19 0.63 -0.35 -0.32 -0.21 -0.15 -0.23 0.30 -0.52 -0.02 0.00 0.24 -0.09 0.17 -0.20 0.25 -0.29 0.03 -0.11 0.49 -0.02 0.51 0.01 0.24 -0.17 -0.14 -0.49 -0.15 3.11 2.12 11.50 7.90 48.30 56.20 0.71 0.55. related taxa than only on vegetative or floral characters. It suggests that both vegetative and floral characters should be taken into consideration for distinguishing these taxa.. DISCUSSION Morphological distinctiveness of C. sinensis f. formosensis The results of clustering analyses based on the floral (Fig. 5) and all characters (Fig. 7) showed almost a similar clustering pattern. In both phenograms all C. sinensis f. formosensis samples were grouped into a single cluster and clearly separated from C. sinensis var. sinensis and C. sinensis var. assamica. Although the phenogram based on vegetative characters alone showed an inconsistency position of C. sinensis f. formosensis from eastern Taiwan (Group I-2, Fig. 3), the plot of the first two components based on vegetative characters indicated the eastern C. sinensis f. formosensis is intermediate morphologically rather.
(10) March, 2007. Su et al.: Morphological comparisons of Taiwan wild tea plant and related taxa. 2. Component 2. 1. 0. -1. -2 -2. -1. 0. 1. Component 1. Fig. 8. Ordination plot of NLPCA based on vegetative and floral characters. : C. sinensis f. formosensis. {: C. sinensis var. assamica. c: C. sinensis var. sinensis.. than closed to the other two taxa (Fig. 4). Taken together, the present study has shown that clear morphological differences existed between C. sinensis f. formosensis and two other closely related taxa, and it seems not proper to treat it as the same as C. sinensis var. sinensis as considered by Ming (2000). The present results show that reproductive organs provide more informative characters for the classification of tea plant than do vegetative structures. This is in close agreement with that previously reported by Banerjee (1992a). In general, reproductive characters have been considered more useful than vegetative features in plant systematics (Stuessy, 1990). In both cluster analysis and NLPCA, the currently recognized varieties C. sinensis var. sinensis and C. sinensis var. assamica integrated considerably. Economic tea plants are heterogeneous with many overlapping morphological attributes. Most vegetative characters show a continuous variation and a high degree of plasticity, and hence, cannot be separated into discrete groups to identify various taxa (Banerjee, 1992a). For the improvement of tea quality, it did happen that artificial hybridizations on the two taxa in the history. 79. (Banerjee, 1992b). In this study, some materials of C. sinensis var. sinensis and C. sinensis var. assamica were collected from the wild in China or the germplasm banks, which were thought to be genetically independent. Other materials were sourced from tea gardens which might possibly be hybrids. These hybrids have intermediate characteristics that may confuse their identification. However, the individuals of C. sinensis f. formosensis formed a clearly defined group, and were never embedded in the group of C. sinensis var. sinensis and C. sinensis var. assamica. Hu (2004) used 15 leaf characters measured on 132 tea germplasms to evaluate inter-taxa variation among C. sinensis var. sinensis, C. sinensis var. assamica and C. sinensis f. formosensis. In the scatterplot of PCA (Hu, 2004, Fig. 4), all individuals of the three taxa showed two distinct groups. Individuals from C. sinensis f. formosensis were dispersed throughout both groups. This is incongruent with present study. Two reasons may explain these inconsistent results. First, characters considered as diagnostic in the present study such as bud pubescence, young branchlet pubescence, abaxial midrib pubescence and petiole pubescence were not used by Hu. Only 7 out of the 15 characters (Hu, 2004, Table 10) adopted by Hu were used in the present study, but these characters were not significantly different among taxa in both studies. Second, Hu transformed nominal characters to ordinal variables for PCA analysis, and this would produce results different from those derived from NLPCA with the same characters. A comparison between PCA and NLPCA showed that the NLPCA would gain more loadings and led to a better performance than PCA (Ellis et al., 2006). Taxonomic rank of C. sinensis f. formosensis As mentioned before, there has been controversial regarding the appropriate taxonomic rank of C. sinensis f. formosensis. Current study showed that C. sinensis f. formosensis could be clearly discerned from C. sinensis var. sinensis and C. sinensis var. assamica in both cluster analysis and NLPCA. In contrast, extensive overlap was found between C. sinensis var. sinensis and C. sinensis var. assamica, even though these two varieties have long been recognized as distinct species (Chang, 1984) or infraspecies (Kitamura, 1950; Sealy, 1958; Ming, 2000). Therefore, it is quite probable that C. sinensis f. formosensis might deserve the species rank. Further work, perhaps including molecular approaches, may be necessary to resolve these taxonomic questions..
(11) 80. TAIWANIA. Morphological identification of C. sinensis f. formosensis Based on the results of morphological study and NLPCA, several characters were found useful to distinguish among these tea plants (Table 6). The surface features of buds and ovaries with the highest component loadings clearly separated C. sinensis f. formosensis from C. sinensis var. sinensis and C. sinensis var. assamica by the first principal component. The buds of C. sinensis var. sinensis and C. sinensis var. assamica are densely covered with silver-yellowish hairs, whereas those of C. sinensis f. formosensis are glabrous or partly covered with sparse hairs. The surface features of buds have been previously used to identify C. sinensis f. formosensis by Suzuki (1937) and Kitamura (1950). The surface of ovaries was also a reliable and stable character. Ovaries of C. sinensis f. formosensis are glabrous while those of C. sinensis var. sinensis and C. sinensis var. assamica are pubescent. The importance of reproductive characters in the taxonomy of Camellia has been previously reported (Hsieh et al., 1996). Based on herbarium specimens, the flowering time of C. sinensis f. formosensis extends from September to January. After that period identification can only be based on vegetative characters. In most cases, young branches, abaxial midribs and petioles of C. sinensis f. formosensis are glabrous, while those of C. sinensis var. sinensis and C. sinensis var. assamica are hairy. There are, however, some inconsistencies that do not fit with the above delineation. The samples of C. sinensis f. formosensis from eastern Taiwan share some characteristics with C. sinensis var. sinensis and C. sinensis var. assamica such as sparsely hairy young branches, abaxial midribs and petioles. Other characters with higher component loadings included leaf pubescence on abaxial surface, pairs of lateral veins and petal size. However, variation of these characters was continuous with. Vol. 52, No. 1. some degree of overlap across taxa and was not considered to be the most taxonomically discriminating. The angles between midrib and lateral veins have been considered by local technical personals to be useful characters for distinguishing. They feel the angles were usually wider in C. sinensis f. formosensis. However, component loadings of these characters were low, indicating that the use of these characters to discriminate among taxa is not reliable. Phytogeography of C. sinensis f. formosensis The remarkable floristic similarity between Taiwan and southeastern China has long been recognized (Li, 1957; Hsieh, 2003). Migrations between Taiwan and mainland China were facilitated by the presence of the Taiwan Strait land bridge that had connected Taiwan and mainland China several times during the glacial ages. The close morphological affinity between C. sinensis f. formosensis and C. sinensis implied that this forma may originated in mainland China. There was a marked difference in distribution patterns between the western and eastern populations of C. sinensis f. formosensis (Fig. 1). C. sinensis f. formosensis is distributed almost continuously throughout the western side of Taiwan, while there is only one population on the southeastern flank of the Central Mountain Range. The eastern population was likely the result of post-glacial range expansion of the populations in western Taiwan. The little separation between the western and eastern populations of C. sinensis f. formosensis in vegetation characters indicated that the vegetative characters of this plant displayed a stronger response to environmental variables than did floral characters. Future research should explore different approaches, including the use of different molecular markers, to establish the post-glacial migration patterns of the Taiwan wild tea plant and determine its origin.. Table 6. Useful characters to distinguish C. sinensis f. formosensis from C. sinensis var. sinensis and C. sinensis var. assamica. An asterisk (*) denotes characters that are (nearly) decisive. Character. C. sinensis f. formosensis. Bud pubescence* Young branchlet pubescence Abaxial midrib pubescence Petiole pubescence Leaf pubescence on abaxial surface Pairs of lateral veins Ovary pubescence * Petal length Petal width. glabrous or sparsely covered with hairs at margins glabrous except for eastern populations glabrous except for eastern populations glabrous except for eastern populations glabrous 8-14, mostly > 10 glabrous 0.7-1.3 cm 0.6-1.1 cm. C. sinensis var. sinensis and C. sinensis var. assamica densely covered with silver-yellowish hairs nearly all hairy nearly all hairy nearly all hairy mostly hairy 5-12, mostly < 10 pubescent 1.1-2.1 cm 0.7-2.0 cm.
(12) March, 2007. Su et al.: Morphological comparisons of Taiwan wild tea plant and related taxa. ACKNOWLEDGEMENTS We greatly appreciate TAI, TAIF, PPI, Tea Research and Extension Station and Prof. Tzen-Yuh Chiang for their support and permission to examine the related specimens. Financial support was provided by the National Science Council of the Republic of China (NSC-95-2422-H-002-002).. LITERATURE CITED Banerjee, B. 1992a. Botanical classification of tea. In: Willson, K. C. and M. N. Clifford (eds.), Tea: Cultivation to consumption. Chapman & Hall, London, UK. pp. 25-51. Banerjee, B. 1992b. Selection and breeding of tea. In: Willson, K. C. and M. N. Clifford (eds.), Tea: Cultivation to consumption. Chapman & Hall, London, UK. pp. 53-86. Bayly, M. J., A. V. Kellow, P. J. D. Lange, K. A. Mitchell, K. R. Markham, P. J. Garnock-Jones and P. J. Brownsey. 2003. Geographic variation in morphology and flavonoid chemistry in Hebe pubescens and H. bollonsii (Scrophulariaceae), including a new infraspecific classification for H. pubescens. N. Z. J. Bot. 41: 23-53. Binns, S. E., B. R. Baum and J. T. Arnason. 2002. A taxonomic revision of Echinacea (Asteraceae: Heliantheae). Syst. Bot. 27: 610-632. Bland, J. M. and D. G. Altman. 1997. Statistics notes: Cronbach's alpha. Brit. Med. J. 314: 572. Chang, H.-T. 1984. A revision on the tea resource plants. Acta Sci. Nat. Univ. Sunyatseni 23: 1-12. Cheng, W.-T. 1990. Populational studies on Formosan Lilium (Liliaceae) I. A cluster analysis of variants in L. longiflorum Thunb. Taiwania 35: 198-205. Chou, C.-S. 1717. Chu-Lo-Hsien-Chi. The 56th year of the Kangxi Emperor, Ching Dynasty. Reprinted in 1962 by Bank of Taiwan, Taipei, Taiwan. p. 286. Cronbach, L. J. 1951. Coefficient alpha and the internal structure of tests. Psychometrika 16: 297-334. de Leeuw, J. 1982. Nonlinear principal components analysis. In: Caussinus, H., P. Ettinger, and R. Tomassone (eds.), COMPSTAT Proceedings in Computational Statistics, Physica Verlag, Vienna, Austria. pp. 77-89. Ellis, R. N., P. M. Kroonenberg, B. D. Harch and K. E. Basford. 2006. Non-linear principal components analysis: an alternative method for. 81. finding patterns in environmental data. Environmetrics 17: 1-11. Gower, J. C. 1971. A general coefficient of similarity and some of its properties. Biometrics 27: 857-871. Gugerli, F. 1997. Hybridization of Saxifraga oppositifolia and S. biflora (Saxifragaceae) in a mixed alpine population. Plant Syst. Evol. 207: 255-272. Hasimoto, M. 1967. Studies on the wild tea plants from Taiwan. Sci. Rep. Fac. Agric. Meijo Univ. 4: 17-20. Hsieh, C.-F. 2003. Composition, endemism and phytogeographical affinities of the Taiwan Flora. In: Huang, T.-C. et al. (eds.), Flora of Taiwan, 2nd ed. 6: 1-14. Editorial Committee, Dept. Bot., NTU, Taipei, Taiwan. Hsieh, C.-F., L.-K. Ling and K.-C. Yang. 1996. Camellia, Theaceae. In: Huang, T.-C. et al. (eds.), Flora of Taiwan, 2nd ed. 2: 667-674. Editorial Committee, Dept. Bot., NTU, Taipei, Taiwan. Hu, C.-Y. 2004. Studies on the variations in leaf characters and DNA sequences of tea germplasm in Taiwan. M. S. Thesis. National Taiwan University, Taipei, Taiwan. 146pp. Huang, S.-J. 1724. Report on the Topography, Culture, Society, Resources, Inhabitants and History of Taiwan (Taihai Shichalu). The 61st year of the Kangxi Emperor, Ching Dynasty. Reprinted in 1957 by Bank of Taiwan, Taipei, Taiwan. p. 62. Keng, H. 1950. The Theaceae of Taiwan. Taiwania 1: 223-268. Kitamura, S. 1950. On tea and camellias. Acta Phytotaxon. Geobot. 14: 56-63. Lai, J.-A., W.-C. Yang and J.-Y. Hsiao. 2001. An assessment of genetic relationships in cultivated tea clones and native wild tea in Taiwan using RAPD and ISSR markers. Bot. Bull. Acad. Sin. 42: 93-100. Li, H.-L. 1957. The genetic affinities of the Formosan flora. Proceedings of the Eighth Pacific Science Congress 4: 189-195. Li, H.-L. 1976. Theaceae. In: Li, H.-L. et al. (eds.), Flora of Taiwan 2: 592-619. Epoch Publ. Co., Taipei, Taiwan. Li, T.-C. and C.-K. Chang. 2003. Tea germplasm resources in Taiwan. Tea Research and Extension Station, Taoyuan, Taiwan. p. 173. Liu, T.-S. and F.-Y. Lu. 1967. Studies in the Taiwan Theaceae based on the morphological characters of leaves. Tech. Bull. Exp. For. Nat. Taiwan Univ. 52: 1-55..
(13) 82. TAIWANIA. Masamune, G. 1936. Short Flora of Formosa. The Editorial Department of "Kudo", Taihoku (Taipei), Taiwan. p. 140. Mckenzie, R. J., J. M. Ward, J. D. Lovis and I. Breitwieser. 2004. Morphological evidence for natural intergeneric hybridization in the New Zealand Gnaphalieae (Compositae): Anaphalioides bellidioides × Ewartia sinclairii. Bot. J. Linn. Soc. 145: 59-75. Ming, T.-L. 2000. Monograph of the Genus Camellia. Yunnan Science and Technology Press, Kunming, China. pp. 128-134. Muvaffak, A., M. Dogan and C. C. Bilgin. 2001. A numerical taxonomic study of the genus Acantholimon Boiss. (Plumbaginaceae) in Ankara Province (Turkey). Isr. J. Plant Sci. 49: 298-300. Schultze-Motel, J. and D. Meyer. 1981. Numerical taxonomic studies in the genera Triticum L. and Pisum L. Genet. Resour. Crop Evol. 29: 241-250. Sealy, J. 1958. A revision of the genus Camellia. Royal Horticultural Society, London, UK. 239pp. Shih, C., Y.-S. Chen, S.-K. Yang, C.-Y. Shih and T.-L. Liau. 1972. Investigation of the wild tea plants in Nantou, Taiwan. Taiwan Agric. Quart. 8: 193-201. Shih, Y.-S. 1995. Translated documents of tea agricultural articles during Japanese Colonized Period. Tea Research and Extension Station, Taoyuan, Taiwan. 521pp. Sokal, R. R. and C. D. Michener. 1958. A Statistical Method for Evaluating Systematic Relationships. Univ. Kansas Sci. Bull. 38: 1409-1438. St-Laurent, L., B. R. Baum, K. Akpagana and J. T. Arnason. 2000. A numerical taxonomic study of Trema (Ulmaceae) from Togo, West Africa. Syst. Bot. 25: 399-413. Stuessy, T. F. 1990. Plant Taxonomy: The Systematic Evaluation of Comparative Data. Columbia University Press, New York, USA. 514pp. Suzuki, S. 1937. Taiwan Zyumoku Benran, Taihoku (Taipei), Taiwan. p. 262. Ward, J. M. 1993. Systematics of New Zealand Inuleae (Compositae - Asteraceae) - 2. A numerical phenetic study of Raoulia in relation to allied genera. N. Z. J. Bot. 31: 29-42. Wu, C.-T., J.-H. Fong and J.-M. Tsay. 1972. Observation on the morphological characters of the wild tea plants on Mt. Meiyuan of Taiwan (II). Taiwan Agric. Quart. 8: 133-159.. Vol. 52, No. 1. Wu, C.-T., Y.-H. Chia, J.-H. Fong and J.-M. Tsay. 1970. Observation on the morphological characters of the wild tea plants on Mei-yuan Mountain of Taiwan. Taiwan Agric. Quart. 6: 1-10. Ying, S.-S. 1995. Systematic study on Theaceae of Taiwan. Memoirs of the College of Agriculture, National Taiwan University 35: 131-170. Zaharof, E. 1988. A phenetic study of Fritillaria (Liliaceae) in Greece. Plant Syst. Evol. 161: 23-34..
(14) March, 2007. Su et al.: Morphological comparisons of Taiwan wild tea plant and related taxa. 83. 利用數值分類方法比較臺灣野生茶與兩近緣分類群之形態特徵關係 蘇夢淮(1)、鄒稚華(2)、謝長富(1,3) (收搞日期:2006 年 8 月 30 日;接受日期:2006 年 12 月 22 日). 摘. 要. 臺灣野生茶 (Camellia sinensis f. formosensis) 乃泛指自生於臺灣中海拔山區的茶類植 物,臺灣植物誌將其中名稱為臺灣山茶。在分類史上,臺灣野生茶曾經被處理成數個不 同分類階層的歸屬。為了提供分類學者一個更客觀的看法,我們利用數值分類方法中的 群聚分析 (cluster analysis) 與非線性主成份分析 (nonlinear principal component analysis), 計算了 165 份標本中的 16 個營養及 11 個花部特徵的測量值,並依據分析結果來探究臺 灣野生茶與關係密切的茶 (C. sinensis var. sinensis) 和阿薩姆茶 (C. sinensis var. assamica) 的區別。將營養及花部特徵獨立或合併進行群聚分析的結果幾乎一致地指出臺灣野生茶 在形態特徵上的獨立性。相反地,茶與阿薩姆茶的形態差異卻無法被解析出來。非線性 主成份分析則顯現休眠芽與子房的毛被狀態是區分臺灣野生茶以及茶與阿薩姆茶的最 有效特徵。參照目前分類學家對茶與阿薩姆茶這兩群植物的分類處理,我們建議臺灣野 生茶應該被處理成種的階層會比較恰當。 關鍵詞:野生茶、臺灣山茶、臺灣、數值分類。. ___________________________________________________________________________ 1. 國立臺灣大學生態學與演化生物學研究所,106台北市羅斯福路4段1號,臺灣。 2. 中央研究院植物暨微生物學研究所,115 台北市研究院路 2 段 128 號,臺灣。 3. 通信作者。Email: [email protected].
(15)
數據


+7




相關文件
if no candidates for expansion then return failure choose leaf node for expansion according to strategy if node contains goal state then return solution. else expand the node and
• bool CanIntersect() returns whether this shape can do intersection test; if not, the shape must provide.
茶捲葉蛾多危害成葉,幼蟲分散後隨即吐絲將 2 片葉黏在一起,棲於內面 取食,隨著幼蟲長大,再將附近
(A) The scene where the heroine of Guardian: The Lonely and Great God places a maple leaf between the pages of a book was shot in Canada.. (B) The maple leaf on the Canadian
service table 服務桌、side table 旁桌,都是擺在工作區附近,作為分菜或放置 備用餐具的桌面。folding leaf table 摺疊桌、round table 圓桌、square table
• The function that describes the histogram (the shape of the distribution) is called the probability density function or pdf, of the random variable.. Is there a difference between
awkeotsang)是一種臺灣特有藤本植 物,在分類上屬於桑科、榕屬。適合生長在濕潤涼爽的環境,臺灣 的野生愛玉分布在中高海拔
Partial Volume Segmentation with Voxel Histograms; Higher Order Statistics for Tissue Segmentation; III Quantification; Two-dimensional Shape and Texture
