行政院國家科學委員會專題研究計畫 成果報告
水稻穀粒發育之研究:穀粒蛋白質表現、功能與栽培溫度之
關係﹝3/3﹞
計畫類別: 個別型計畫 計畫編號: NSC92-2313-B-002-002- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學農藝學系暨研究所 計畫主持人: 盧虎生 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 12 月 15 日
Abstract
Proteins are essential to rice caryopsis development and quality formation. High
temperature is an important environmental factor which may decrease grain quality. In the present study rice caryopsis proteins were profiled by two-dimensional polyacrylamide gel electrophoresis, and differentially expressed proteins were analyzed by liquid chromatography/tandem mass spectrometry (LC\MS\MS). Expressions of more than 400 polypeptide spots were monitored during caryopsis development, and more than 70 differentially expressed polypeptides were analyzed by LC\MS\MS. We identified 54 proteins with known functions. Of these, 21 were involved with carbohydrate metabolism, 14 with protein synthesis and sorting, and 9 with stress responses. Waxy (Wx) proteins and glutelins were the most significant spots increased significantly during development. Allergen-like proteins, PPDK and NADH-SDH, also were expressed during development, implying their physiological roles in caryopsis. Expression of large isoforms of Wx proteins was correlated with the amylose content of rice caryopses. One protein with high GC content in its DNA sequence was correlated with the chalky trait of kernels. High temperature (35/30 ) ℃ decreased expression of Wx proteins, allergen-like proteins and elongation factor 1 β, but increased expression of small heat shock proteins (sHSP),
glyceraldehyde-3-phosphate dehydrogenase and prolamin. sHSP was positively correlated with the appearance of chalky kernels. During development, glutelins were phosphorylated and glycosylated, indicating these molecules were post-translationally modified. Possible functions of the expression of candidate proteins on grain quality are discussed.
Keywords: rice; proteomics; high temperature; grain quality; Wx protein; heat shock
中文摘要 蛋白質在水稻穎果發育與品質形成過程中扮演重要之角色,高溫為導致水稻 品質降低的主要環境因素之一。本研究利用雙向電泳技術建立水稻穎果蛋白質表 現之基本圖譜,再將其具有表現差異之蛋白質以逢機質譜方式鑑定其種類。本試 驗中可分離至少400個以上的蛋白質,其中70個具表現差異的蛋白質利用 LC\MS\MS的方式進行身分鑑定。並找出57個已知功能的蛋白質。其中包括21個與 碳水化合物生合成或代謝相關的酵素、14個與蛋白質合成或累積相關的酵素以及 9個與逆境反應有關的蛋白質。Waxy與glutelins為最主要隨著發育時期顯著增加 的蛋白質。Allergen-like proteins, PPDK and NADH-SDH等在穎果發育時期具 有重要生理功能的酵素亦被大量表現。其中Waxy基因之數個不同isoforms的表現 量與其質鏈澱粉含量具有相關性。其中一個蛋白質之基因具有高度重複之GC比 例,且其與心腹白之形成有關。高溫(35/30 ℃)會減少Waxy, allergen-like proteins與elongation factor 1β之表現,但會使低分子量之class 1 sHSP大 量表現,G-3-P dehydrogenase, prolamins與sHSP等基因的過量表現與心腹白的 形成具正相關。在穎果發育期間,glutelins有磷酸化與醣化修飾之反應,顯示 其具有轉譯後修飾之作用。穎果發育期間表現之重要蛋白質的功能及其與水稻穀 粒品質形成可能之關聯在文中有進一步之探討。
Introduction
Improving grain quality has been a major objective of rice culture management and breeding. Understanding rice kernel development should benefit the improvement of grain quality. Development of the rice caryopsis has been well characterized
histologically [1, 2]. The physiological processes in rice caryopses have been documented, and the biosynthesis of major macromolecules, such as starch and storage protein, also has been studied intensively [1]. However, the molecular basis for these processes still needs to be clarified, especially for the processes affecting grain quality.
After anthesis, the rice embryo and endosperm undergo cell division and
differentiation. The embryo grows faster than the endosperm and reaches maturity 10 days after anthesis (DAA). Prior to 10 DAA, the endosperm primarily undergoes cell division and cellularization [2]. From 10 DAA on, it commences significant cell enlargement and macromolecule (dry matter) accumulation. These processes continue until grain maturation [1, 3]. DNA endoreduplication of endosperm nuclei was
observed during the stage between cell division and enlargement [4], yet the
physiological importance of this phenomenon is unclear. Our previous study showed that rice caryopses are most sensitive to environmental stresses, such as high
temperature, during early development (before 15 DAA, the milky stage) [5]. However, the regulation and molecular basis of the responses to stresses need to be elucidated.
In rice, caryopsis proteins are important to development and grain quality. Most studies of rice caryopsis proteins have focused on the biosynthesis of starch and storage proteins. While most enzymes related to carbohydrate metabolism increase during kernel development, ADP-glucose pyrophosphorylase, Q-enzyme, and granule bound starch synthase (GBSS, Wx protein) are thought to play key roles in rice starch biosynthesis [6, 7]. Because amylose is a major factor affecting the physiochemical properties of starch and rice grain palatability, Wx protein has been the focus of studies on improving the quality of rice grains [8]. In different rice genotypes,
expression of GBSS gene mRNA is regulated by an alternative splicing mechanism at the polymorphic region at 5’ upstream of the GBSS gene of two alleles, Wxa
and Wxb,
during transcription and post transcription [9, 10]. The amount of Wx protein is positively correlated with the amylose content of rice kernels [11]. Four Wx protein isoforms have been found in rice kernels [12]. However, the regulation of translation, the function of each isoform and their relationship to amylose content are unknown. Protein content also is crucial to rice grain quality and nutritional value. Crude protein content is negatively correlated to the palatability of japonica type rice [13]. Most research on rice proteins has concentrated on storage proteins, especially glutelins and prolamins, which constitute 80% and 5% of the total storage protein in rice grains [14]. Both proteins are encoded by multigene families. During
development, glutelins accumulate in protein body II, and prolamins accumulate in protein body I [1, 14]. An ER luminal binding protein (BiP) facilitates the folding and assembly of prolamins in the formation of protein body I [15], and protein disulfide isomerase (PDI) might help the sorting process for both glutelins and prolamins [16]. Protein body I is less digestible than protein body II. Prolamin content is negatively correlated with the palatability of rice grains [1, 13]. For glutelins and prolamins, the
molecular basis of gene expression and protein accumulation has been studied thoroughly. However, for albumins and globulins much less is known about these processes. The effect of environmental factors on the accumulation of each class of storage protein is unclear.
During the major growing season in Taiwan, the temperature usually exceeds 30 ℃ during grain filling period. Similar temperature conditions may also occur in the rice growing areas of Australia and Japan [17, 18]. Rice grains are most vulnerable to high temperature during early development (before 15 DAA) [5, 19]. High temperature may damage the accumulation and packing of starch grains [18], decrease
carbohydrate and nitrogen content [19], and increase the frequency of chalky and abnormal grains [5, 17]. In addition, high temperature may decrease the amylose content of rice grains [20]. Furthermore, high temperature might affect the alternative splicing process at the 5’ splice site of the GBSS gene, especially for the Wxb allele
[21, 22]. There is little information on the effect of high temperature on rice grain storage proteins. In wheat high temperature (>35 ) increased the expression of ℃ Heat shock proteins (HSPs). The HSPs might modify the polymerization of glutelins during development, causing the grains to exhibit a weak dough property [23]. In transgenic rice, over-expression of the hsp101 gene enhanced the heat tolerance of Basmati rice plants in the vegetative stage [24]. But the effects of heat shock proteins on kernel development and quality formation have not been determined.
In general, research on the functions of rice caryopsis proteins has been conducted one biochemical pathway at a time, for example, the biosynthesis of starch or storage protein. However, recently developed proteomic technology allows for global
investigation of the functions of caryopsis proteins during development and provides integrated information, especially for responses to environmental factors. Proteomic studies of rice have profiled and identified the proteins associated with metabolic pathways in different organs [25, 26, 27, 28], responses to hormones [29, 30], and responses to water stress [31]. In this study, we made observations of the dynamic changes in protein expression patterns during rice caryopsis development; and determined which candidate proteins may be crucial to grain quality especially in a high temperature environment. Our proteomic analysis used two-dimensional
electrophoresis to monitor dynamic changes in protein expression patterns during rice caryopsis development. Proteins expressed differentially during development or in response to high temperature were selected and identified by liquid
chromatography/tandem mass spectrometry (LC/MS/MS). The possible function of each identified protein was investigated and discussed further.
Materials and Methods Plant materials
Two rice cultivars, an indica type (Taichung Native 1, TN 1) and a japonica type (Tainung 67, TNG 67), were used in this study. The indica type cultivar is the source of a semi-dwarf gene in rice breeding program. The japonica cultivar is grown widely in Taiwan and has high yields. However, TNG 67 is sensitive to high temperature, which causes the formation of chalky grains [5]. Rice plants were grown with natural light in a phytotron. Control plants were grown at 30/25 ℃(day/night) while plants in the high temperature treatment experienced 35/30 ℃. Panicles were labeled at
anthesis when at least half the spikelets were flowering. Spikelets were harvested 3, 6, 9, 12, 15, and 30 (maturity) DAA and stored in liquid N2. Caryopses were removed
from the spikelets for the following analyses.
In addition, seeds of two low amylose type cultivars (TKwx 5 and TKwx 2), two high amylose type cultivar (IR 36 and KSC 7), three high chalky type cultivars (Pegonia, JY 1 and Arborio) and four premium quality cultivars (Basmati, M401, Jasmin and Koshihikari) were received from the Taichung District Agriculture Experimental Station (Taichung, Taiwan). These seeds were used to determine the relationship between kernel quality and the expression of candidate genes selected from the proteomic analysis.
Caryopsis weight and nuclei number
Caryopsis fresh weight and dry weight were monitored during development. Fresh weight was measured right after sampling and dry weight was measured after lyophilizing for 24 h. At least 15 caryopses were weighed on each sampling date. Caryopsis nucleus number was monitored to determine developmental status during the early growth stage. Caryopses were sampled 3, 6, 9 and 12 DAA and fixed in 950 g kg-1 ethanol. The number of nuclei was determined as described by Lur and Setter [32]. Samples were mixed with digestion buffer containing 2% (w/v) cellulose (No. 203019, Yakult, Tokyo, Japan), 0.1% (w/v) pectolyase Y23 (Seishin, Tokyo, Japan), 0.5% (w/v) macerozyme R10 (Yakult) and 13% (w/v) mannitol. Nuclei were stained with acetocarmine and counted using a haemocytometer under a microscope.
Glutelin and amylose analysis
During caryopsis development, glutelin and amylose content were measured to monitor macromolecule accumulation. Glutelin content was determined as described by Lookhard et al. [33] with the minor modifications described in our previous report [5]. Caryopsis samples were ground in five volume of 5% (v/v) acetic acid containing 5% (v/v) 2-mercaptoethanol, suspended for 30 min and centrifuged at 5,000 x g. The extraction was repeated three times and the supernatants were collected. Protein content was determined with a Bio-Rad Protein Assay kit (Bio-Rad Laboratories, Richmond, CA, USA), with BSA as the standard.
[5]. A 10 mg sample was mixed with 5 mL distilled water and then 10 mL 0.5 N NaOH were added. The solution was thoroughly mixed and shaken overnight. Amylose content was measured with an amylose autoanalyzer (BRAN+LUEBBE GmbH 2000, Norderstedt, UK), using amylose purified from cultivar TNG 67 as the standard. For each sampling stage of caryopsis development, there were four
replicates for the analysis of glutelin and amylose.
Protein extraction and 2D electrophoresis
Sample grains were dehusked and 30 mg were suspended with 2 ml lysis buffer (8.5 M urea, 1% (v/v) Triton X-100, 2.5% (w/v) DTT), centrifuged at 8,000 x g. The supernatant was collected. Proteins were precipitated with acetone at -20 ℃ for 2h, centrifuged at 12,000 x g, and vacuum dried. Protein content was determined with a BIO-RAD DC protein assay kit (Bio-Rad Laboratories) in accordance with the manufacturer’s instructions.
For IEF electrophoresis, proteins were resuspended with sample buffer containing 8.5 M urea, 1% (v/v) Triton X-100, 2.5 % (w/v) DTT, 1% (v/v) Pharmalyte, and tracking dye. IPG gel strips (13 cm, pH 3-10, Immobiline DryStrip, Amersham Biosciences, Uppsala, Sweden) were equilibrated overnight with 25 µl of the sample solution. IEF was conducted with a Multiphore II unit (Amersham Pharmacia Biotech), run at 3500 V for 8 h. In addition, IPG gels were used for SDS-PAGE with 15% (w/v) separation and 5% (w/v) stacking gels. Protein spots were visualized with a silver staining kit (Amersham Pharmacia Biotech).
Quantitative analysis of protein spots
For each caryopsis developmental stage, 2D-PAGE gels were recorded with a densitometer and analyzed with Bio-Rad PDQuest 7.1 software. The average
background reading value of our gels was 250. The densitometer value is negatively correlated with the density (abundance) of the protein spot. A ‘+‘ was used to represent the value range 250 – 220. The symbols +, ++, +++, ++++, and +++++ corresponded to the reading ranges 250-220, 219-190, 189-160, 159-130 and 129-100 respectively. During development, readings for spots 31, 35, and 41 were consistently near 250. Therefore, the average readings of these spots provided the reference for the ‘+’ range. At each developmental stage, the gels from at least five independent
samples were used to quantitatively measure the proteins.
Protein identification using tandem mass spectrometry
Protein spots were excised from 3 to 5 replicate gels. Excised spots were destained [35] and then subjected to in-gel reduction, alkylation and tryptic digestion [36]. The digestion mixture was loaded onto a 150 × 0.5 mm PE Brownlee C18 column with 5 µM particle diameter and 300 Å pore size (Perkin-Elmer). A gradient of 5% (v/v) to 65% (v/v) acetonitrile in 0.1% (v/v) formic acid over 60 min chromatographed peptides at a flow rate of 5 µl/min directly into the electrospray source of a Finnigan LCQ ion trap mass spectrometer. The mass spectrometer was programmed to acquire successive sets of three scan modes. The MS scan determined ion intensities from 395-1605 m/z. The most abundant ions were analyzed by zoom scan and MS/MS scan.
The resulting MS/MS spectra of the peptides were interpreted by database correlation with SEQUEST Blowser software [37, 38]. Protein identification was conducted using the SWISS-PROT (http://kr.expasy.org/sprot/), NCBI
(http://www.ncbi.nlm.nih.gov/), KOME (http://cdna01.dna.affrc.go.jp/cDNA/) and TIGR databases (http://tigrblast.tigr.org/tgi/).
Immunodetection of Waxy protein and heat shock protein
Immunoblotting was performed to confirm the expression of Wx protein and small molecular mass heat shock proteins (sHSP). Waxy protein antibody was received from Dr. Jaw-Shu Hsieh (Institute of Botany, Academia Sinica, Taiwan) and sHSP antibody was provided by Dr. Chu-Yung Lin (Department of Life Science, National Taiwan University). Caryopsis proteins were separated by 2D-PAGE and blotted onto nitrocellulose membranes. Immunodetection and analysis were conducted as described by Sambrook, Fritsch and Maniatis [39], using SuperSignal West Pico Chemiluminescent Substrate (PIERCE, Rockford, IL, USA).
Phosphorylation and glycosylation assays
After 2D-PAGE, selected gels were subjected to phosphorylation or glycosylation assays. The phosphorylation (Pro-Q Diamond Phosphoprotein Gel Stain Kit P-33300, Molecular Probes, Eugene, Oregon, USA) and glycosylation (Pro-Q Emerald 300 Glycoprotein Gel and Blot Stain Kit P-21857, Molecular Probes) assays were
performed according to the manufacturer’s instructions. Stained gels were visualized with an UV transilluminator at 300 nm. Spots showed positive phosphorylation signals were excised from gels than subjected to mass analysis to determine possible sites of phosphorylation, using the method as described in Tsay et al. [36].
Reverse transcriptase PCR
The RNA of differentiatially expressed proteins was analyzed by reverse transcriptase PCR (RT-PCR). Total RNA was isolated from 1 g of caryopsis endosperm, 9 DAA. First–strand cDNA synthesis was conducted with 1 µg of total RNA following the manufacturer’s instructions (Advantage TM RT-for-PCR Kit, Bio-Rad). PCR amplification of selected genes was performed using primers searched and obtained from the rice genomic database. The selected genes and primers were: heat shock protein 17.9 (5’-ATGTCGCTGATCCGCCGCA-3’ and
5’-ATTTGCTCCTTCACGACAGACA-3’); heat shock protein18.0 (5’-CACAATACACAATGTCGCTGAT-3’ and
5’-TCTCCACCGTGCATGTCACA-3’); G-3-P dehydrogenase
(5’-ATGGGCAAGATTAAGATCGGA-3’ and 5’-CTACTGGGTCTTGGCCATGT-3’); allergen-like protein (5’-ATGGCGTCCAAGGTTGAGCT-3’ and
5’-GCTAGTTCTTGAGGAGGTAGT -3’); elongation factor 1 beta (5’-ATGGCCGTGACCTTCACCGA-3’ and
5’-AATGCTTCCAGAACAACCAGT-3’); nucleoside diphosphate kinase (5’-ATGGAGCAGTCCTTCATCATGA-3’ and
5’-CTAAGACTCATAGATCCAAGGGT-3’); glutelin (5’-CTAGTCGTGCTGATTCATACAA-3’ and
5’-TTCGGACTCATTCGATAATCCT-3’); prolamine 7 (5’-ATGAAGATCATTTTCGTCTTTGCT-3’ and
5’-AACTTCCGAACCACTATACTGT-3’); Wx (GBSS) gene
(5’-GACGTCCAGATCGTTCTTCT-3’ and 5’-TCAAGGAGCAGCCACGTTCT-3’). For amplification, there were 30 cycles of denaturation for 30 sec at 94 ℃, annealing for 45 sec at 57 ℃, and extension for 1 min at 72 by Perkin℃ -Elmer 3700 Thermal Cycler. PCR products were visualized by 1% agarose gel electrophoresis. Fragments of RT-PCR cDNA were sequenced at Academia Seneca, Taiwan.
Results
Growth Analysis
To provide basic physiological information for the proteomic analysis of caryopsis development, we measured the dry weight, number of endosperm nuclei, amylose, glutelin and urea-soluble protein contents during the early stages of kernel
development (Fig. 1). From 6 to 15 DAA, dry weight and endosperm nuclei number increased linearly. Glutelin and amylose content increased rapidly from 6 to 12 DAA, but the increase had slowed by 15 DAA. Urea-soluble protein, which was used for proteomic analysis, accumulated relatively slowly from 6 to 12 DAA, but increased rapidly thereafter. This rapid increase was coincident with the rapid accumulation of storage proteins after 12 DAA (Fig. 2). Caryopsis water content decreased gradually from 60%, 6 DAA, to near 20%, 15 DAA. In addition the effect of high temperature on amylose content of caryopsis of TNG 67 at 15 DAA was also determined. The amylose contents are 12.3 ± 0.5 % and 15.6 ± 0.4 % in the caryopsis for high temperature treatment and the control respectively. High temperature caused the decrease of amylose content in the caryopsis grown under high temperature. We determined the level of DNA endoreduplication in the caryopsis and endosperm. The endosperm nuclei C-value changed from 3C to 6C at 3 DAF, and to 12C at 15 DAF. Some nuclei reached 24C at 15 DAA. DNA endoreduplication is an indicative of endosperm cell differentiation during early developmental stage [4].
2-D Electrophoresis and Protein Profiling
Urea extraction buffer extracted large amounts of rice caryopsis proteins for use in 2-D electrophoresis. In the sample collected 12 DAF, silver staining revealed nearly 400 protein spots in the gel (Fig. 2). However, more than 1200 spots could be monitored with a computer-equipped image analysis system. We found proteins in a variety of function categories. Resolution of the present 2-D system is comparable to other rice 2-D profiling studies on leaf or embryo [25, 27, 28]. Thus, the present system can be applied to profile caryopsis proteins.
Gel regions with glutelins frequently exhibited negative staining, especially on gels with samples older than 12 DAA. This may result from the high amounts of glutamic acid and aspartic acid in glutelins [40] and the binding of the amino acids and Ag+. The rapid accumulation of glutelins after 12 DAA may inhibit the reduction of silver ions in the protein complex zone on the gel [41].
Protein Identification and Function
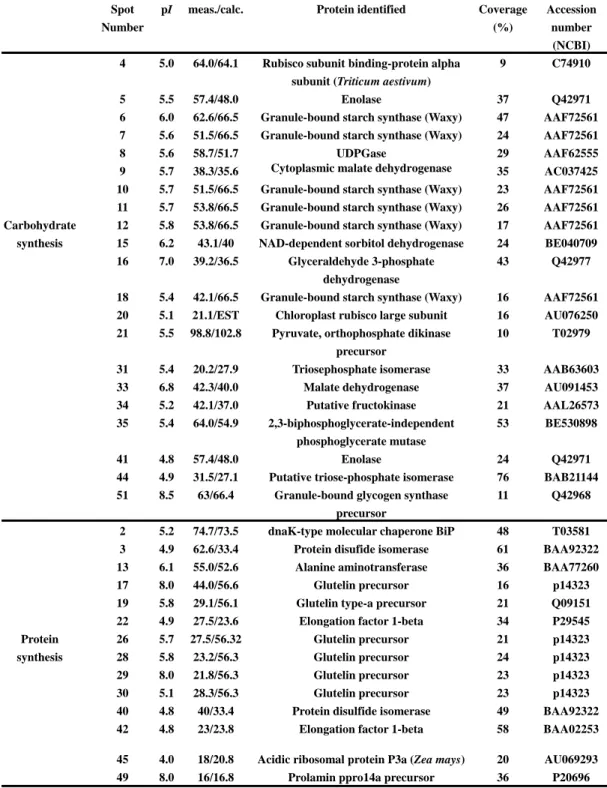
Protein spots exhibiting differential expression on gels from different caryopsis developmental stages were chosen for LC/MS/MS identification. Out of more than 70 selected spots, 54 proteins and their annotated functions were identified from the survey of gene banks (Table 1). The efficiency of identification of protein functions is comparable or even higher than that of other proteomic studies on rice embryo [27, 28]. Functionally, 21 proteins were related to carbohydrate metabolism, and 14 proteins were involved in protein biosynthesis, including glutelins and prolamins, the major storage proteins in rice seeds. Most of the identified glutelins were
αsubunit-type glutelin polypeptides, as reported in other studies [40, 42]. Nine proteins were related to stress response; most of them were heat shock proteins. Another 9 proteins had a specific function that we could not categorize. Finally, we could not assign any function to one protein (no. 25).
Several proteins that exhibited differential expression during caryopsis development, including the Wx protein isoforms and protein no. 25, were studied in greater detail to clarify their functions and their possible relationship to rice quality.
Changes in Protein Expression during Development
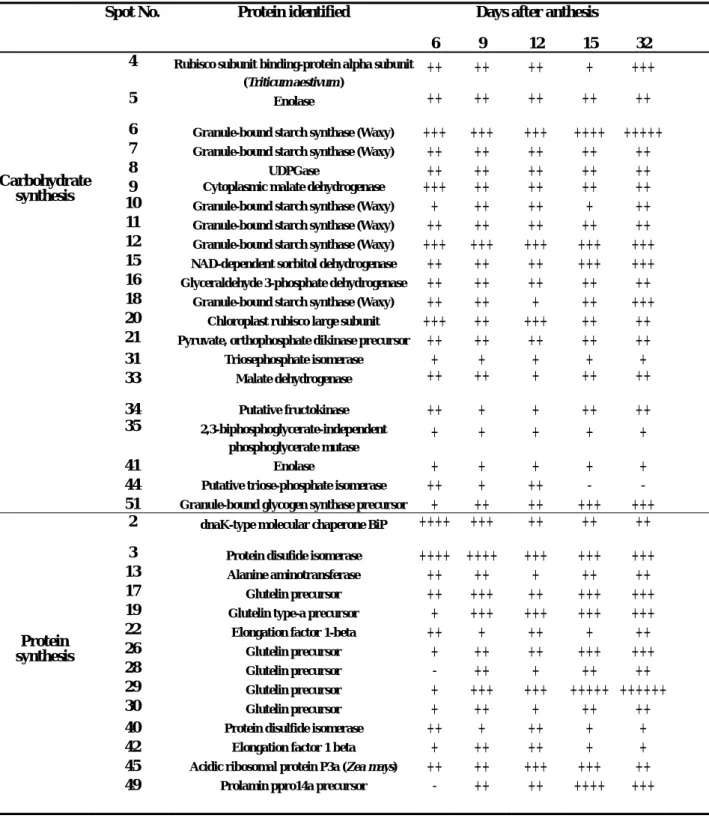
The abundance of most identified proteins involved in carbohydrate metabolism increased after 6 DAF (Table 2). There were six Wx proteins (granule bound starch synthase) and these could be classified into two major groups based on molecular mass. The amount of high molecular mass (Mr) (represented by spot 6) and low Mr
(spot 12) isoforms increased during development. At maturation, high Mr Wx
isoforms accounted for a considerable portion of the seed proteins, especially in high amylose content cultivars (Fig. 4). Interestingly, pyruvate, orthophosphate dikinase (spot 21), a C4 photosynthesis related enzyme, was identified in the gel. This protein was expressed stably until maturation.
Three important proteins involved in protein folding/sorting were identified: Bip (spot 2), heat shock protein 70 (spot 1) and a protein disulfide isomerase (spot 3). These proteins appeared at 6 DAA and accumulated until maturation. Six glutelin
polypeptides and a prolamin polypeptide were also identified. The amount of these proteins increased drastically after 6 DAA. The large amount of glutelin proteins could cause negative staining in gels subjected to silver staining. After 15 DAA, glutelin could account for more than 20% of the total caryopsis proteins appearing in the gel.
Effect of Temperature on the Expression of Caryopsis Proteins
The protein content of kernels of japonica cultivar TNG 67 grown at high temperature was lower than that of control kernels. Compared to TNG 67, protein expression in indica cultivar TN 1 was relatively stable (Fig. 3). When grown at high temperature, TNG 67, which is heat sensitive, produced lower amounts of protein no. 25 and formed chalky grains. Thus far, the functional annotation of protein 25 has not been reported. The kernels of TNG 67 and TN 1 grown at high temperature had lower levels of protein 52 than kernels from control plants. Protein 52 was identified as an allergen-related protein (Table 1). In TNG 67 grown at high temperature, expression of protein 22, identified as elongation factor 1ß, was lower than in control kernels 12 DAA.
However, high temperature treatment increased the abundance of some proteins. Three (no. 47, 53 and 54) were identified as low Mr heat shock proteins (sHSP, Table 1). Expression of sHSP in the high temperature treatment was confirmed by
immunoanalysis (Fig. 4). High temperature had little effect on the expression of high Mr heat shock proteins. In addition, high temperature treatment increased the amount of glyceraldehyde-3-phosphate dehydrogenase (no. 16) in TNG 67 kernels at 12 DAA, and increased expression of prolamine (no. 49), especially in TN 1.
sHSP Expression and Grain Quality
Reports of the relationship between heat induced sHSP and wheat dough quality led us to examine the correlation between sHSP and kernel quality in rice [23]. The presence of sHSP in the gel was confirmed by immunoblotting for the caryopsis endosperm of high temperature grown TNG 67 (Fig. 4). In addition to spots no.53 and no.47, an additional sHSP spot was identified by the immunoblotting (Fig. 4, left button box, below the spot no. 53). High temperature treatment stimulated the
expression of all three sHSP during the grain filling stage. In general the caryopses of TNG 67 grown at high temperature exhibit the chalky trait. Thus, we examined the relationship between sHSP expression and chalky endosperm using grains from 10 cultivars exhibiting different levels of chalkiness. sHSP were less expressed in Koshihikari, Jasmin, KSC 7 and TK 9 (so-called premium quality rice varieties) than in Pegonia, Arborio, JY 1 and TNG 67 (chalky varieties) (Fig. 4).
Waxy Protein Isoforms and Amylose Content
Wx protein is the major enzyme involved in amylose biosynthesis in rice. More than 6 isoforms were identified in the gels so we investigated the relationship between the expression of each isoform and the amylose content of rice kernels. Immunoblotting analysis detected high amounts of Mr isoforms (Fig. 5, button boxes). Wx protein was remarkably expressed in the kernels of varieties with high amylose content (TN 1 and IR 36). In gluten rice varieties (TKwx 5 and TKwx 2), which had less than 5%
amylose content, there was little or no expression of Wx protein isoforms. In the varieties tested, the abundance of high Mr Wx protein isoforms (represented by spot no. 6) was positively correlated with amylose content. However, the expression of low Mr isoforms (represented by spot 12) was not correlated with amylose content in the varieties tested.
Protein 25 and the Chalky Trait
Because high temperature treatment resulted in lower amounts of protein 25 in the caryopses of heat-sensitive cultivar TNG 67, we analyzed the relationship between protein 25 expression and the chalky trait in 6 cultivars. In the 6 tested cultivars, protein no. 25 expression was higher in the translucent than in the chalky regions of the grains (Fig. 6, B). We cloned the gene and identified its full-length DNA sequence and the amino acid sequence using the rice genomic database (Fig. 6, A). The DNA sequence of the correspondent gene has 74% GC content and is located on
chromosome 7. So far, the function of the sequence has not been determined.
Expression of protein no. 25 commenced at 9 DAA and gradually increased 2.5-fold by maturation (Fig. 6, C). Thus, expression of the gene coding for protein no. 25 appears to be related to the formation of chalkiness in rice grains.
Glycosylation and Phosphorylation of Caryopsis Proteins
During development, seed proteins may be modified at post-translational level [43]. Using recently developed commercial kits, we examined the status of
and phosphorylation of caryopsis proteins are shown in Fig. 7. In the phosphorylation assay, only the regions in the gel with Wx proteins and glutelin yielded positive signals for protein expression at 12 DAA (Fig. 7, A). Four Wx protein isoforms and seven protein spots in the glutelin protein region exhibited signals for phosphorylation staining. The four Wx isoforms detected corresponded to the high Mr Wx isoforms (Fig. 1). Spot no 24, identified as a DNA fragment, gave a strong phosphorylation signal in the gel and can be used as an internal check to confirm that phosphorylation is being detected. Possible phosphorylation sites of glutelins and Wx isoforms were further analyzed by LC\MS\MS. A phosphopeptide (274ASLQEQEQGQVQpSR287)
with the phosphorylation site at Ser-286 of was detected in gutelins (Fig. 7, C). No significant positive signal of phosphorylation could be detected in Wx isoforms by our present LC/MS/MS system. In the protein glycosylation assay, seven proteins in the glutelin region gave positive signals (Fig. 7, B).
RT-PCR analysis
Regulation of temperature responsive proteins at the RNA level was analyzed using RT-PCR (Fig. 8). Expression of small HSP, G-3-P dehydrogenase and prolamin were enhanced by high growing temperature. High temperature lowered the gene
expression of allergen-like protein and elongation factor 1 beta. In general, the trends of the expressions at RNA level were in accordance with that at the protein level.
Discussion
2D Electrophoresis and Profiling of Rice Caryopsis Proteins
Two-dimensional gel electrophoresis has been used to study proteins important to carbohydrate or storage protein biosynthesis in rice caryopses or embryo [11, 25, 27, 28, 42]. In comparison, the system used in our study presented a more comprehensive protein expression profile. Most major proteins that have been reported in other studies could be displayed clearly in our gels. Some of these proteins may affect rice grain quality. In rice grains, Wx protein content was positively correlated with amylose content, which in turn affects the physical and chemical properties, and, hence, palatability of rice [8]. Glutelin expression is positively correlated to the protein content and nutrient value of rice grains [40]. Thus, the protein profile of a rice kernel is indicative of its physiological/biochemical status and its
physicochemical properties. In our system, many important, quality-related proteins could be identified and monitored simultaneously on the gels (Fig. 1). In addition, we demonstrated that protein expression patterns vary with temperature (Fig. 3) and among genotypes that differ in grain quality (Fig. 5). This profiling method may be able to display polymorphisms among different genotypes/varieties, and even to reveal the quality characteristics of different genotypes/varieties.
Dynamic Expression and Function of Caryopsis Proteins
Several studies have used proteomic analysis to examine rice caryopsis proteins [28]. Our study provides additional information on the dynamic changes in protein
expression during development. Protein expression patterns changed markedly from 1 to 15 DAA (Table 2). During this time, the rice caryopsis undergoes active mitotic division followed by the commencement of the rapid accumulation of dry matter [1, 2]. Environmental factors can have a profound effect on grain quality during this stage [5, 19]. We have provided a global description of protein expression during this stage. After 6 DAA, the regions of the gel exhibiting the greatest changes in protein
expression were those with Wx protein isoforms and glutelins. Changes in the
expression of these proteins can be used to mark the physiological development stages of rice caryopses. In addition, we found that proteins extracted 3 DAA, which were distributed mostly in the low pI region of the gel, were fairly unstable and decreased the reproducibility of the gel pattern. We are working on ways to circumvent this problem. Similar proteomic approach for different organs of caryopsis such as embryo is worthy of further study to specify the dynamic expressions and functions of
proteins at organ level [27, 28].
Starch is the major storage carbohydrate in rice kernels. Expression of most of the identified proteins involved with starch biosynthesis increased after 6 DAA. Changes in the expression of these proteins were consistent with the changes in their
corresponding activities reported in previous studies [1, 6]. It is interesting that we found two classes of Wx protein isoforms. The higher molecular mass isoforms seem to be the same as those reported by Nakamura et al. [12]. During development, the four large isoforms increased in abundance simultaneously. This increase was coincident with the increase in amylose. Thus, these large isoforms may be the main enzymes involved with amylose synthesis in rice kernels (Fig. 5). Regulation of gene expression of Wx protein at the transcription level has been studied extensively [9, 10,
21], but regulation at the translation level is still unknown. We found that only the large isoforms might be phosphorylated (Fig. 7), which suggests translational modification occurs in the rice caryopsis during development. Because the isoforms have different pI values, further research is needed to determine the phosphorylation status, activity of each isoform and the dynamic changes that occur during
development.
Interestingly, pyruvate orthophosphate dikinase (PPDK, spot no. 21), a C4 plant enzyme, is expressed during development [44]. The prominent and consistent expression of this protein lends support to the hypothesis that it may be involved in CO2 fixation in the dark or amino acid synthesis in rice caryopses [44, 45]. In
addition, PPDK expression was closely and horizontally in line with several other spots that may be PPDK isoforms with different pI. We also identified a
NADP+-dependent sorbitol dehydrogenase (NAD-SDH; spot no. 15). This may be the first report of the existence and developmental expression of this protein in rice. NAD-SDH has ketose activity and might be involved with sucrose degradation, sorbitol synthesis and starch biosynthesis in maize endosperm [46]. In fruit,
NAD-SDH might play an important role in the translocation of carbon sources [47]. The function of this protein in rice caryopses needs to be clarified.
Glutelins and prolamins are the most prominent storage proteins in rice and have been studied thoroughly [1]. We found that glutelins were expressed as early as 3 DAA and the timing was earlier than that of prolamins. In addition to glutelins and prolamins, Bip and protein disulfide isomerase (PDI), two proteins crucial to folding/sorting, were identified. Both proteins were expressed abundantly and constantly throughout caryopsis development (Fig. 1, Table 2). Bip might fold and sort prolamins into protein body I, while PDI is involved in the segregation of preglutelins and prolamins in rice caryopses [43]. We found that BiP was expressed about 6 DAA. The timing of BiP expression paralleled the expression of prolamins. PDI was expressed as early as 3 DAA, which corresponds with the expression of PDI mRNA [16]. Temporally, PDI expression was similar to that of some glutelin precusors, proteins no. 17 and 29, which were expressed as early as 3 DAA (data not shown). This supports the idea that PDI is crucial to glutelin accumulation. Our system provides a tool for observing the coordinated expression of functionally related proteins during development, and is especially valuable for detecting the effects of environmental conditions and genetic differences, such as mutants, on protein expression.
We identified several proteins related to stress responses, including GSH-dependent dehydroascorbate reductase and heat shock proteins. In general, these proteins are expressed at about the same level throughout caryopsis development. This indicates that they have essential, basic functions, such as maintaining the physiological stability of seed molecules as seed moisture content gradually decreases during maturation. In addition, we found that high temperature treatment increased the abundance of several low molecular mass heat shock proteins (discussed in the following section).
The expression of some allergen-like proteins was monitored. In general, they were expressed consistently during development, as represented by protein no. 52. There are very few reports documenting the physiological function of allergen-like proteins in rice caryopsis [26, 50]. We found that high temperature treatment decreased
expression of the allergen-like proteins (see below).
The Effect of Temperature on Protein Expression and Grain Quality
During the grain filling stage, high temperature can affect the starch and crude protein contents of rice grains, which leads to changes in grain quality [17, 18]. However, the effects of high temperature on carbohydrate and storage protein biosynthesis in rice grains have not been studied in depth. Our gel profiles documented the effects of high temperature on the abundance of Wx proteins, glutelins and prolamins simultaneously. All three crucially affect grain quality. Responses were variety dependent. Based on changes in the abundance of Wx proteins and prolamins, the japonica variety TNG 67 was more sensitive to high temperature, while the indica variety TN 1 was more sensitive to low temperature. These responses are in accord with the evolutionary origin and geographic distribution of each variety. Increase of prolamins by high temperature can decrease palatability of rice grains [1, 13]. And the temperature effect on Wx protein expression may affect the starch characteristics and grain quality [8, 13]. As shown in the present case, high temperature treatment leads to the reduction of Wx protein expression in TNG 67 (Fig. 3). The decrease of the expression of Wx protein may result in a lower enzyme activity of GBSS and amylose content in the grains grown at high temperature [7, 20]. Since amylose content is associated with the starch characteristics of rice grains [8], the lower amylose content can affect starch viscosity characteristics of the grains grown at high temperature [20]. Nevertheless, although we observed a decrease of amylose content and Wx protein expression in the caryopsis under high temperature treatment, no significant effect could be detected at the transcription level (Fig. 8). It is known that low temperature can regulate
alternative splicing of Wx gene expression [21], enhance the enzyme activity and cause an increase in amylose content [7]. Further research is needed to assess the effect of high temperature fluctuation on the coordination between transcription and translation of the Wx locus.
Early stage is a transition process of caryopsis development. Endosperm cell mitosis decreases gradually, nuclei DNA endoreduplicates and macromolecules accumulate rapidly. Elongation factor 1β may connect the regulation between cyclin dependent kinase of the mitotic cycle and protein translation [49]. We found that high
temperature decreased expression of elongation factor 1β (spots 22, 42; Figs. 3, 8). We suspect that elongation factor 1β might mediate the effects of temperature on early caryopsis development.
A family of proteins allergenic to animals exists in rice kernels. Some of the allergenic proteins belong to the alpha-amylase/trypsin inhibitor family [50]. The physiological function of these proteins in rice kernel development is unclear. We found that high temperature decreased expression of allergen like protein and its RNA, but increased the incidence of chalky trait (spot 52), suggesting these proteins may affect grain structure. A similar phenomenon was reported in a study using protein mutant line [48].
The function of protein no. 25 has not been documented in the rice bioinformatics database. We found that expression of this protein was much lower in chalky endosperm than in translucent endosperm. Protein no. 25 was not homologous with friabilin, which is a marker protein for hardness in wheat grain [51]. Based on
hydropathy data, protein no. 25 might be located in the cytoplasmic region of the cell. We have cloned this gene and are conducting experiments, including in situ
hybridization, to determine the possible functions of this protein.
In rice seedlings, expression of glyceraldehyde-3-phosphate dehydrogenase RNA is enhanced by submergence, drought and ABA treatment [52]. It has been suggested that the gene for glyceraldehyde-3-phosphate dehydrogenase is involved with
physiological responses to anaerobic environments. Expression of this protein and its RNA was enhanced by high temperature in rice kernels, especially in the sensitive cultivar TNG 67 (Figs. 3, 8). Our examination of rice roots indicates the existence of several isoforms of this enzyme (data not shown). The physiological function of this gene and its possible isoforms in rice caryopses merits further research.
Heat shock proteins
Transgenic expression of a high Mr HSP gene (hsp 101) enhanced the tolerance of rice plants to high temperature stress [24]. In our study, although several classes of HSP proteins were monitored, high temperature treatment significantly stimulated expression of sHSP RNA and proteins (Fig. 4 and Fig. 8). However, the function of sHSP in rice caryopses has not been reported. We found that sHSP were expressed differentially in caryopses grown at high 35/30 and low temperatures 25/20 . ℃ ℃ Temperatures of 35/30 are common in subtropical rice growing areas, such as ℃ Taiwan, during the grain filling stage. High temperature may be one of major factors that decrease rice grain quality [5, 17, 18]. In wheat, high temperature treatment increases HSP 70, which is associated with a decrease in wheat dough quality [23]. High temperature also induces expression of a sHSP gene that might increase pectin depolymeration and affect tomato juice viscosity [53]. In E. coli, expression of rice sHSP could change the protein aggregation pattern and confer thermotolerance [54]. We suspect that sHSP might play a similar role in rice caryopses grown at high temperatures. The sHSP could protect seed proteins from heat injury. However, sHSP might also affect the aggregation of macromolecules, such as starch grains or storage proteins, during grain filling, and change the physicochemical properties of rice grains. Research is being conducted to determine the function of sHSP on the development and quality of rice kernels, such as in situ localization and RNAi transformation.
Phosphorylation and Glycosylation of Caryopsis Proteins
There are few reports on post-translational regulation of rice seed proteins. Using recently developed kits, we made preliminarily observations on the phosphorylation and glycosylation status of rice caryopsis proteins. Phosphorylation at Ser—286 of a polypeptide of glutelins was confirmed using LC\MS\MS analysis. This may be the first report revealing the evidence of phosphorylation for rice glutelins. The
phosphorylation may be related to the expression of protein kinases, such as a Ca2+-dependent protein kinase [55] or a SNF type protein kinase [56], during early development. Although phosphorylation signal for Wx isoforms was detected by method of gel staining, we have been unable to identify the phosphorylation sites by our present method [36]. High possibility of phosphorylation for Wx proteins is predicated by protein database (NetPhos 2.0 Server,
http://www.cbs.dtu.dk/services/NetPhos/). Further sensitive or sophisticate analysis is needed to confirm the phosphorylation of Wx isoforms, as a recent report on the
phosphorylation of starch branching enzymes in wheat [57]. In legume seeds,
glycosylation is related to storage protein stability during accumulation. However, few reports have documented the glycosylation status and functions of kernel storage proteins in rice [43]. We have been performing analysis of dynamic changes of phosphorylation and glycosylation of these proteins to explore possible functions of the post translational modification on caryopsis development and quality formation. In conclusion, we report a first proteomic profiling of developmental changes for the expression of rice caryopsis proteins. Relative or coordinate developmental changes of important proteins crucial to storage proteins and carbohydrates accumulations during growth or in response to environmental stresses can be monitored by the profiling. Several proteins related to grain quality and high temperature response were also identified. The results should benefit the understanding of functions of rice caryopsis proteins and improvement of rice grain quality under temperature stress.
The authors are grateful to Professor Jaw-Shu Hsieh (Department of Agronomy, National Taiwan University) for providing antibody of Wx protein, and to Dr.
Chu-Yung Lin (Department of Life Science, National Taiwan University) for providing antibody of sHSP. This research was funded by National Science Council, Taiwan, Republic of China. 計畫成果自評 本研究內容與原定計畫目標一致,本試驗為三年期的研究計劃,目前已順利 將 proteomics 技術應用於水稻榖粒在不同溫度環境因子下之蛋白質雙向電泳圖 譜的差異分析。初步已完成 80 個具有表現差異的蛋白質分子之序列分析與比 對,並得知各分子在水稻基因組中的位置及其可能的功能。 目前已知在日本型與印度型品種,在各榖粒發育期於不同溫度環境下的蛋白 質差異表現,藉由 proteomics 的技術將各具有差異表現的蛋白質一一進行功能 分析,提供水稻基礎的蛋白質圖譜分析研究,判定各蛋白質在水稻榖粒發育中所 扮演的角色,以供作將來育種與分子操作上的重要參考指標。本試驗也以蛋白質 磷酸化與醣化分析的技術,成功應用在水稻穎果發育期間具有轉譯後修飾之蛋白 質的分析與功能探討,此技術亦可應用於植物學領域上,許多基因轉譯後修飾作 用之研究。 本研究室目前正針對上述結果進行整理,並繼續分析未分析的蛋白質,目前 已將部份成果選錄並發表於Proteomics期刊(SCI),水稻榖粒proteomics的分 析,除可供本研究室進一步研究其與溫度環境間的關係,也可提供給相關研究人 員做進一步與稻米品質相關的探討。
References
[1] Tanaka, K., Kasai, Z., Ogawa, M., in: Matsuo, T., Kumazawa, K., Ishii, R., Ishihara, K., Hirata, H. (Eds.), Science of the Rice Plant, Volume Two: Physiology, Food and Agriculture Policy Research Center, Tokyo, Japan. 1995, 97-118.
[2] Brown, R. C., Lemmon, B. E., Olsen, O. A., J. Plant Res. 1996,109, 301-313. [3] Ellis, J. R., Gates, P. J., Boulter, D., Ann. Bot. 1987, 60, 663-670.
[4] Ramachandran, C., Raghavan, V., Ann. Bot. 1989, 64, 459-468. [5] Huang, J. J., Lur, H. S., J. Agric. Assoc. of China. 2000, 1, 370-389. [6] Nakamura, Y., Yuki, K., Plant Sci. 1992, 82, 15-20.
[7] Umemoto, T., Nakamura, Y., Ishikura, N., Phytochemistry 1995, 40, 1613-1616. [8] Larkin, P. D., McClung, A. M., Ayres, N. M., Park, W. D., Euphytica 2003, 131,
243-253.
[9] Cai, X. L., Wang, Z. Y., Xing, Y. Y., Zhang, J. L., Hong, M. M., Plant J. 1998, 14, 459-465.
[10] Hirano, H. Y., Eiguchi, M., Sano, Y., Mol. Biol. Evol. 1998, 15, 978-987. [11] Sano, Y., Theor. Appl. Genet. 1984, 68, 467-473.
[12] Nakamura, T., Yamamori, M., Hirano, H., Hidaka, S., Phytochemistry 1993 33, 749-753.
[13] Matsue, Y., Odahara, K., Hiramatsu, M., Jap. J. Crop Sci. 1995, 64, 601-606. [14] Yamagata, H., Sugimoto, T., Tanaka, K., Kasai, Z., Plant Physiol. 1982, 70,
1094-1100.
[15] Muench, D. G., Wu, Y., Zhang, Y., Li, X., Boaton, R. S., Okita, T. W., Plant Cell
Physiol. 1997, 38, 404-412.
[16] Takemoto, Y., Coughlan, S. J., Okita, T. W., Satoh, H. et al., Plant Physiol. 2002,
128, 1212-1222.
[17] Tashiro, T., Wardlaw, I. F., 1991, Aust. J. Plant Physiol. 18, 259-265.
[18] Zakaria, S., Matsuda, T., Tajima, S., Nitta, Y., Plant Prod. Sci. 2002, 5, 160-168. [19] Tashiro, T., Wardlaw, I. F., Aust. J. Agric. Res. 1991, 42, 485-496.
[20] He, G. C., Kogure, K., Suziki, H., Japan. J. Crop Sci. 1990, 59, 340-345. [21] Larkin, P. D., Park, W. D., Plant Mol. Biol. 1999, 40, 719-727.
[22] Umemoto, T., Terashima, K., Funct. Plant Biol. 2002, 29, 1121-1124.
[23] Blumenthal, C., Stone, P. J., Gras, P. W., Bekes, F. et al., Cereal Chem. 1998, 75, 43-50.
[24] Katiyar-Agarwal, S., Agarwal, M., Grover, A., Plant Mol. Biol. 2003, 51, 677-686.
[25] Komatsu, S., Kjiwara, H., Hirano, H., Theor. Appl .Genet. 1993, 86, 935-942. [26] Koller, A., Washburn, M. P., Lange, B. M., Andon, N. L. et al., Proc. Natl. Acad.
Sci. 2002, 18, 11969-11974.
[27] Woo, S. H., Fukuda, M., Islam, N., Takaoka, M. et al., Electrophoresis 2002, 23, 647-654.
[28] Fukuda, M., Islam, N., Woo, S. H., Yamagishi, A. et al., Electrophoresis 2003, 24, 1319-1329.
[29] Rakwal, R., Komatsu, S., Electrophoresis 2000, 21, 2492-2500.
[30] Komatsu, S., Konishi, H., Shen, S., Yang, G., Mol. Cell. Proteomics 2003, 2, 2-10.
[31] Salekdeh, G. H., Siopongco, J., Wade, L. J., Ghareyazie, B., Bennett, J.,
Proteomics 2002, 2, 1131-45.
[33] Lookhart, G. L., Juliano,B. O., Webb, B. D., Cereal Chem. 1991, 68, 396-400. [34] Juliano, B. O., Cereal Chem. 1971, 16, 334-340.
[35] Gharahdaghi, F, Weinberg, C. R., Meagher, D. A., Imai, B. S., Mische, S. M.,
Electrophoresis 1999, 20, 601-605.
[36] Tsay, Y. G., Wang, Y. H., Chiu, C. M., Shen, B. J., Lee, S. C., Anal. Biochem. 2000, 287, 55-64.
[37] Eng, J. K., McCormick, A. L. Yates, J.R., J. Am. Soc. Mass Spect., 1994, 5, 976–989.
[38] Chittum, H. S., Lane, W. S., Carlson, B. A., Roller, P. P. et al., Biochemistry 1998,
37, 10866–10870.
[39] Sambrook, J., Fritsch, E. F., Maniatis, T. (Eds.), Molecular Cloning: A laboratory manual, 2nd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 1989.
[40] Wen, T. N., Luthe, D. S., Plant Physiol. 1985, 78, 172-177. [41] Rabilloud, T., Electrophoresis 1990, 11, 785-794.
[42] Abe, T., Gusti, R. S., Ono, M. Sasahara, T., Genes Genet. Syst. 1996, 71, 63-68. [43] Shewry, P. R., Napier, J. A., Tatham, A. S., Plant Cell 1995, 7, 945-956.
[44] Sadimantara, G. R., Abe, T., Suzuki, J., Hirano, H., Sasahara, T., J. Plant Physiol. 1996, 149, 285-289.
[45] Imaizumi, N., Ku, M. S. B., Ishihara, K., Samejima, M., Plant Mol. Biol. 1997,
34, 701-716.
[46] Doehlert, D. C., Plant Physiol. 1987, 84, 830-834.
[47] Oura, Y., Yamada, K., Shiratake, K., Yamaki, S., Phytochemistry 2000, 54, 567-572.
[48] Schaeffer, G. W., Sharpe, F. T., Theor. Appl. Genet. 1997, 95, 230-235.
[49] Pomerening, J. R., Valente, L., Kinzy, T. G., Jacobs, T. W., Mol. Gen. Genom. 2003, 269, 776-788.
[50] Adachi, T., Izumi, H., Yamada, T., Tanaka, K., Plant Mol. Biol. 1993, 21, 239-248.
[51] Giroux, M. J., Talbert, L., Habernicht, D. K., Lanning, S., Hemphill, A., Martin, J. M., Crop Sci. 2000, 40, 370-374.
[52] Arumugam, Pillai, M., Lihuang, Z. Akiyama, T., Theor. Appl. Genet. 2002, 105, 34-42.
[53] Ramakrishna, W., Deng, Z., Ding, C. K., Handa, A. K., Ozminkowski, R. H.,
Plant Physiol. 2003, 131, 725-735.
[54] Yeh, C. H., Chen, Y. M., Lin, C. Y., Plant Physiol. 2002, 168, 661-668.
[55] Tsutomu, K., Nobuaki, H., Tadashi, B., Kazuo, S., Hiroaki, S., Gene 1993,
129,183-189.
[56] Takano, M., Kajiya-Kanegae, H., Funatsuki, H., Kikuchi, S., Mol. Gen. Genet. 1998, 260, 388-394.
[57] Tetlow, I. J., Wait, R., Lu, Z., Akkasaeng, R., Caroline, G. et al., Plant Cell, 2004, 16, 694–708
Table 1. Functional categorization of caryopsis proteins isolated from 2D gels. meas./calc.: measured value/theoretical value of molecular mass.
Spot Number
pI meas./calc. Protein identified Coverage (%)
Accession number
(NCBI) 4 5.0 64.0/64.1 Rubisco subunit binding-protein alpha
subunit (Triticum aestivum)
9 C74910
5 5.5 57.4/48.0 Enolase 37 Q42971 6 6.0 62.6/66.5 Granule-bound starch synthase (Waxy) 47 AAF72561 7 5.6 51.5/66.5 Granule-bound starch synthase (Waxy) 24 AAF72561 8 5.6 58.7/51.7 UDPGase 29 AAF62555 9 5.7 38.3/35.6 Cytoplasmic malate dehydrogenase 35 AC037425 10 5.7 51.5/66.5 Granule-bound starch synthase (Waxy) 23 AAF72561 11 5.7 53.8/66.5 Granule-bound starch synthase (Waxy) 26 AAF72561 12 5.8 53.8/66.5 Granule-bound starch synthase (Waxy) 17 AAF72561 15 6.2 43.1/40 NAD-dependent sorbitol dehydrogenase 24 BE040709 16 7.0 39.2/36.5 Glyceraldehyde 3-phosphate
dehydrogenase
43 Q42977
18 5.4 42.1/66.5 Granule-bound starch synthase (Waxy) 16 AAF72561 20 5.1 21.1/EST Chloroplast rubisco large subunit 16 AU076250 21 5.5 98.8/102.8 Pyruvate, orthophosphate dikinase
precursor
10 T02979
31 5.4 20.2/27.9 Triosephosphate isomerase 33 AAB63603 33 6.8 42.3/40.0 Malate dehydrogenase 37 AU091453 34 5.2 42.1/37.0 Putative fructokinase 21 AAL26573 35 5.4 64.0/54.9 2,3-biphosphoglycerate-independent
phosphoglycerate mutase
53 BE530898
41 4.8 57.4/48.0 Enolase 24 Q42971 44 4.9 31.5/27.1 Putative triose-phosphate isomerase 76 BAB21144 Carbohydrate
synthesis
51 8.5 63/66.4 Granule-bound glycogen synthase precursor
11 Q42968
2 5.2 74.7/73.5 dnaK-type molecular chaperone BiP 48 T03581 3 4.9 62.6/33.4 Protein disufide isomerase 61 BAA92322 13 6.1 55.0/52.6 Alanine aminotransferase 36 BAA77260 17 8.0 44.0/56.6 Glutelin precursor 16 p14323 19 5.8 29.1/56.1 Glutelin type-a precursor 21 Q09151 22 4.9 27.5/23.6 Elongation factor 1-beta 34 P29545 26 5.7 27.5/56.32 Glutelin precursor 21 p14323 28 5.8 23.2/56.3 Glutelin precursor 24 p14323 29 8.0 21.8/56.3 Glutelin precursor 23 p14323 30 5.1 28.3/56.3 Glutelin precursor 23 p14323 40 4.8 40/33.4 Protein disulfide isomerase 49 BAA92322 42 4.8 23/23.8 Elongation factor 1-beta 58 BAA02253 45 4.0 18/20.8 Acidic ribosomal protein P3a (Zea mays) 20 AU069293 Protein
synthesis
49 8.0 16/16.8 Prolamin ppro14a precursor 36 P20696
Spot Number
pI meas./calc. Protein identified Coverage (%)
Accession number (NCBI) 1 4.8 78.1/73.6 Heat shock protein 70B (Dunaliella
salina)
2 AY072744
14 4.0 46.5/71.1 dnaK-type molecular chaperone hsp70 10 S53126 27 5.9 28.9/23.6 GSH-dependent dehydroascorbate
reductase
16 BAA90672
38 5.5 46.0/71.6 dnaK-type molecular chaperone hsp70 7 S53126 39 4.7 39.5/60.9 Mitochondrial chaperonin-60 30 AAN05528 46 5.0 17.9/13.2 Thioredoxin h-type 38 Q42443 47 5.8 16/16.9 Class 1 heat shock protein 49 P27777 53 5.5 16.0/17.9 17.9 kDa heat shock protein 34 AY034057 Stress protein
54 5.4 15.3/18 18 kDa heat shock protein (Zea mays) 30 X54076 23 4.5 15.9/17.1 Pollen allergen-like protein (Arabidopsis
thaliana)
59 AAM65899
24 5.7 15.2/19.8 DNA fragment 31 L12252 32 5.3 57.4/59.1 ATP synthase beta chain 23 Q01859 36 5.3 28.3/61.5 Peptidylprolyl cis-trans isomerase, A
novel heat shock and calmodulin binding PPIase (Triticum aestivum)
23 CAA60505
37 4.8 48.6/73.5 Endosperm luminal binding protein 33 AAB63469 43 4.5 29.5/107.8 Putative chloroplast inner envelope
protein
15 AAG13554
48 6.8 10.7/16.8 Nucleoside diphosphate kinase 54 Q07661 50 9.0 10.7/15.2 Oleosin 30 Q42980 Others
52 4.5 17.1/16.6 Allergen-like protein (Oryza sativa) 25 AU198169
Table 2. Expression of caryopsis proteins during development. The symbols +, ++, +++, ++++, and +++++ correspond to the reading value ranges 250-220, 219-190, 189-160, 159-130 and 129-100, respectively. For each
developmental stage, at least 5 gels from independent samples were used for quantitative measurement of protein spots.
(Continue on next page)
Spot No. Protein identified Days after anthesis
6 9 12 15 32
4 Rubisco subunit binding-protein alpha subunit (Triticum aestivum )
++ ++ ++ + +++
5 Enolase ++ ++ ++ ++ ++ 6 Granule-bound starch synthase (Waxy) +++ +++ +++ ++++ +++++ 7 Granule-bound starch synthase (Waxy) ++ ++ ++ ++ ++ 8 UDPGase ++ ++ ++ ++ ++ 9 Cytoplasmic malate dehydrogenase +++ ++ ++ ++ ++ 10 Granule-bound starch synthase (Waxy) + ++ ++ + ++ 11 Granule-bound starch synthase (Waxy) ++ ++ ++ ++ ++ 12 Granule-bound starch synthase (Waxy) +++ +++ +++ +++ +++ 15 NAD-dependent sorbitol dehydrogenase ++ ++ ++ +++ +++ 16 Glyceraldehyde 3-phosphate dehydrogenase ++ ++ ++ ++ ++ 18 Granule-bound starch synthase (Waxy) ++ ++ + ++ +++ 20 Chloroplast rubisco large subunit +++ ++ +++ ++ ++ 21 Pyruvate, orthophosphate dikinase precursor ++ ++ ++ ++ ++ 31 Triosephosphate isomerase + + + + + 33 Malate dehydrogenase ++ ++ + ++ ++ 34 Putative fructokinase ++ + + ++ ++ 35 2,3-biphosphoglycerate-independent phosphoglycerate mutase + + + + + 41 Enolase + + + + + 44 Putative triose-phosphate isomerase ++ + ++ - - Carbohydrate
synthesis
51 Granule-bound glycogen synthase precursor + ++ ++ +++ +++ 2 dnaK-type molecular chaperone BiP ++++ +++ ++ ++ ++ 3 Protein disufide isomerase ++++ ++++ +++ +++ +++ 13 Alanine aminotransferase ++ ++ + ++ ++ 17 Glutelin precursor ++ +++ ++ +++ +++ 19 Glutelin type-a precursor + +++ +++ +++ +++ 22 Elongation factor 1-beta ++ + ++ + ++ 26 Glutelin precursor + ++ ++ +++ +++ 28 Glutelin precursor - ++ + ++ ++ 29 Glutelin precursor + +++ +++ +++++ ++++++ 30 Glutelin precursor + ++ + ++ ++ 40 Protein disulfide isomerase ++ + ++ + + 42 Elongation factor 1 beta + ++ ++ + + 45 Acidic ribosomal protein P3a (Zea mays) ++ ++ +++ +++ ++ Protein
synthesis
Spot No. Protein identified Day after anthesis
6 9 12 15 32
1 Heat shock protein 70B (Dunaliella salina) ++ ++ + + +++ 14 dnaK-type molecular chaperone hsp70 ++ ++ ++ ++ ++ 27 GSH-dependent dehydroascorbate reductase +++ +++ +++ +++ ++ 38 dnaK-type molecular chaperone hsp70 ++ + + + + 39 Mitochondrial chaperonin-60 + - + + + 46 Thioredoxin h-type ++ ++ ++ ++ ++ 47 Class 1 heat shock protein - + + + ++ 53 17.9 kDa heat shock protein + + - - - Stress enzyme
54 18 kDa heat shock protein (Zea mays) - + + - - 23 Pollen allergen-like protein
(Arabidopsis thaliana)
++ + - - -
24 DNA fragment ++ +++ +++ ++++ +++++ 32 ATP synthase beta chain ++ ++ ++ ++ ++ 36 Peptidylprolyl cis-trans isomerase, A novel
heat shock and calmodulin binding PPIase (Triticum aestivum)
+ + ++ - -
37 Endosperm luminal binding protein ++ + + + ++ 43 Putative chloroplast inner envelope protein ++ + ++ + + 48 Nucleoside diphosphate kinase + ++ ++ +++ ++ 50 Oleosin - + ++ +++ +++ 52 Allergen-like protein (Oryza sativa) ++ + + + + Others
Figure Legends:
Figure 1. Changes in fresh weight, dry weight, the number of nuclei, urea-soluble protein content, glutelin content and amylose content during early caryopsis development. Caryopses of TN 1 cultivar were used for the analysis. Values are means ± SE of four replicates.
Figure 2. A 2D-PAGE protein expression profile of rice caryopses 6 days after anthesis. Caryopses of TN 1 cultivar were used for the analysis. Arrows and numbers indicate the corresponding annotations for the spots.
Figure 3. Effect of temperature on the expression of caryopsis proteins during development. Arrows indicate proteins that were more or less abundant in caryopses subjected to high temperature treatment than in caryopses in the control treatment. Small boxes and numbers indicate the gel regions corresponding to Wx proteins (spot 6), glutelins (spots 19 and 29) and prolamins (spot 49). Intensity of spots in these regions significantly increased during development.
Figure 4. sHSP expression in rice cultivars with and without the chalky trait. Grains of the cultivars Pegonia, Arborio, TNG 67, M401 and JY 1 have the chalky trait. TK 9, Jasmin, basmati, KSC 7 and Koshihikari are premium quality cultivars that do not exhibit this trait. Twenty mature grains of each cultivar were randomly sampled, dekusked, milled and subjected to 2D-PAGE analysis. To confirm sHSP expression, heat-treated samples of TNG 67 were used for immunoanalysis (bottom left and right boxes). Circles and numbers indicate the corresponding sHSP spots. Figure 5. Expression of Wx proteins in cultivars with different amylose contents. TN 1 and IR 36 are indica type cultivars that have high amylose content. TNG 67 and Koshihikari are japonica type cultivars whose amylose content is medium. TKwx 5 and TSwx 2 are japonica and indica type cultivars, respectively, that have low amylose content (wx type variety). The gel of the TN 1 sample was subjected to immunodetection to confirm the expression of Wx protein isoforms. Arrows and numbers indicate the spots corresponding to the Wx protein isoforms.
Figure 6. Characteristics of protein no. 25 gene. (A) DNA sequence. (B) The 2D expression patterns of chalky and translucent kernels of TN1, TNG67, KSC 7, M401, Basmati and JY 1. (C) Change of abundance during development.
Figure 7. Phosphorylation (A) and glycosylation (B) gel profiles of caryopsis proteins, and detected phosphorylation polypeptede of glutelin (C). Caryopses 32 DAA were used for the analysis. Proteins were separated on 2D gels. The phosphorylation and glycosylation status of proteins on gels and mass analysis for phosphorylation site were determined as described in the Materials and Methods. Upper and lower boxes in (A) and (B) indicates the regions of Wx protein isoforms and glutelins, respectively. Arrow indicates the spot identified as a DNA fragment (spot 24). We used this
fragment as an internal reference for the phosphorylation analysis.
Figure 8. Expression of putative temperature responsive protein genes was analyzed by reverse transcription PCR. HSP17.9 and HSP18: heat shock protein with Mr of 17.9 and 18 respectively; prolamin 7: prolamin gene no. 7; G3PDH: G-3-P
dehygrogenase; NDK: nucleoside diphosphate kinase 1; GBSS: granule bound starch synthase; EF1B: elongation factor 1 β; ALP: allergen like protein. Actin was used to check the RNA and amplification conditions of the RT-PCR reaction internally.
(Fig. 2.) p I 4.6 5 .6 6.1 6.9 8.6 12 1 86 69 53 40 28 21 17 M. W .
(Fig. 3.) TNG67 25/20 ℃ 6 DAF TNG67 35/30 ℃ 6 DAF TNG67 25/20 ℃ 12 DAF TNG67 35/30 ℃ 12 DAF TN1 25/20 ℃ 6 DAF TN1 35/30 ℃ 6 DAF TN1 25/20 ℃ 12 DAF TN1 35/30 ℃ 12 DAF 6 9 12 14 19 25 29 47 49 52 54 53 52 52 52 52 52 52 52 47 47 47 47 47 47 53 53 53 53 53 53 54 54 54 54 54 54 54 14 14 14 14 14 14 14 25 25 25 25 25 25 25 6 12 12 6 6 12 12 6 6 12 6 12 6 12 19 29 19 29 19 29 19 29 19 29 19 29 19 29 49 49 49 49 49 49 49 47 53
(Fig. 4.)
Pegonia
Arborio
TNG 67
TK 9
Jasmin
Koshihikari
TNG 67 35/30
℃
TNG 67 25/20
℃
JY 1
M 401
Basmati
KSC 7
(Fig. 5.)
TSw
x2
TKw
x5
TN 1
TNG 67
Koshihikari
IR36
High
amylose
(>28 %)
Middle
amylose
(16-22 %)
Low
amylose
(0-2 %)
Western
blotting
12 6TN 1
12 6 12 6 12 6 12 6 12 6 12 6(Fig. 6.)
translucent chalky
(A)
1 atgatttgcc tccgagccgc cttccccttg acctcctcct tctcctcctc ccctctccgc 61 cgcctcgccc tcaagccctc ctcctcccgc gccgccgccg ccgccgccat gtcgtccgcc 121 cccgccaccg ccatcgccgc gccggtcgag cacatcgtgc tcatcaaggt ccgccccgag 181 gcggcggcgt cgggcgccgc cgcggcgatg gtgtcctcgc tgcaggcgct gtccagcgtg 241 gtgcccgggc tgtcgtacat ccacgtcggc cccgtgctcc gcctccggtc gccggcggcg 301 gaggcgctgg ggccgaccca cgtcctccac tcccgctacg ccaccaagcc cgacctggcg 361 gcgtacgcgg cgcacccggc gcacgtcgcc gccgtgcagg ggcacgtcct ccccaacgcg 421 ctcgacacca ccgccatcga ctgggtcaac gccgcgctcg cgccgtcccc ggtcaccccg 481 ggctccgccg tgcggctcac gctggcgaag gtgaaggaag gtgtggaggt gccccagctc 541 gtggagaagg tcgccgcggc gacggcggcg gcgggcgagg cgaagggcgc cagggtgagc 601 ttcggggaga acttctcccc cgcgcgggct aaggggtacc agttcgggat ggtggcggtg 661 ttcgacagcg tggaggagct cgacgcggtg gagggggacg ggaaggtgca ggaggccaag 721 gcggcggtga ggccactgct cgacgaggtg ctcgtgctgg acttcgtcgt cggacctgcc 781 gccgtggagg ctccggcgcc ggccaacctc tga
(B) (C) TN1 TNG67 KSC7 M401 Basmati JY1
(Fig. 7.)