行政院國家科學委員會專題研究計畫 成果報告
廢水生物處理系統生化指標與分析技術之建立
計畫類別: 個別型計畫 計畫編號: NSC91-2314-B-002-295- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 國立臺灣大學醫學院醫事技術學系暨研究所 計畫主持人: 何憲武 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 92 年 10 月 27 日
Membrane fluidity optimization regulates swarming through modulation of a two-component signal transduction thermosensor in Serratia marcescens
Hsin-Chih Lai1,2,*, Wen-Ching Yi1, Po-Chi Soo1, Yu-Sen Fang1, Jun-Rong Wei1,
Wei-Chun Chou1, Yu-Tze Horng1, Shi-Ming Lin3, Su-Jen Liaw1,2, Shen-Wu Ho1,2,
Simon Swift4 and Paul Williams5,6
1
School and Graduate Institute of Medical Technology, National Taiwan University College of Medicine, Taipei, Taiwan, R.O.C. 2Department of Laboratory Medicine, National Taiwan University Hospital and National Taiwan University College of Medicine, Taipei, Taiwan, R.O.C. 3 ********* 4Division of Molecular Medicine, Faculty of Medical and Health Sciences, University of Auckland, Auckland, New Zealand.5Institute of Infections and Immunity, University of Nottingham, Queen’s Medical Centre, Nottingham, NG7 2UH, United Kingdom and 6School of Pharmaceutical Sciences, University Park, University of Nottingham, Nottingham, NG7 2RD, United Kingdom.
*
For correspondence. Dr. Hsin-Chih Lai, Graduate Institute of Medical Technology,
College of Medicine, National Taiwan University, No.1. Chan-Der Street, Taipei 100,
Taiwan (R.O.C.).
Tel. +886 2 2397 0800 (ext 6931) Fax +886 2 2371 1574
Email: [email protected]
Running title: Swarming regulation in Serratia marcescens.
Keywords: Serratia marcescens, two-component system, thermosensor, swarming motility, fatty acids.
Abstract
Serratia marcescens swarming behavior is thermoregulated, which is characterized by continuous populational surface migration on rich swarming agar surface at 30°C, but not at 37°C. Here we report that environmental temperature shift and fatty acids (especially the saturated fatty acids, SFAs) act as the signal cues regulating S. marcescens swarming through an RssA-RssB two component signal transduction system. SFAs or temperature upshift specifically and negatively regulate S. marcescens swarming, but not swimming motility or the identified factors known to be involved in Serratia swarming, possibly through influencing pattern of cellular fatty acid profile. Fatty acid profile analysis strongly indicates that addition of SFAs at a constant temperature and temperature upshift may either directly or indirectly affect the signaling state of the RssA sensor protein by increasing the incorporation of the high-melting point fatty acids into membrane phospholipid, which subsequently inhibits S. marcescens swarming. The RssA-RssB two-component system mediates this response, as both partners are required to sense and transduce the signals. Expression of a fatty acid synthesis gene fabG and a hemolysin gene shlA is coordinately regulated with swarming by RssA-RssB. Biofilm formation, cell surface structure and extracellular polysaccharide are also affected in the rssA or rssB mutant. Similar phenomena of SFA inhibition are also observed in swarming regulation of Proteus mirabilis and Salmonella typhimurium. Thus, evidence suggests it may be common that membrane fluidity optimization through modulation of cellular fatty acid profile under different physiological culture conditions may subsequently affect two-component signaling pathway, which regulates the composition of cellular fatty acids, expression of virulence factor and multicellular swarming behaviours. This may provide an unidentified and common regulatory circuit for controlling swarming behaviour of not only S. marcescens, but also other swarming bacteria.
Introduction
More and more bacteria are identified to exhibit a form of cell differentiation and multicellular behaviour termed swarming migration. Swarming involves differentiation of vegetative cells into hyperflagellated swarm cells that undergo rapid and coordinated population migration across solid surfaces (Harshey 1994; Shapiro 1998, Fraser and Hughes 1999). Bacterial species reported to show swarming behaviours include Serratia (Alberti and Harshey 1990; Givskov et al., 1998; Liu et al., 2000), Proteus (Mobley and Belas 1995), Vibrio (McCarter and Silverman, 1990), Aeromonas (Kirov et al., 2002), Escherichia coli (Harshey and Matsuyama 1994), Salmonella enterica serovar typhimurium (Harshey and Matsuyama 1994), Pseudomonas (Kohler et al., 2000; Kinscherf and Willis 2002; Sanchez-Contreras et al., 2002), Burkholderia cepacia (Huber et al., 2001), Myxococcus xanthus (Jelsbak and Sogaard-Andersen 2000), a spirochete Treponema denticola (Lux et al., 2002), a nitrogen-fixation bacterium Sinorhizobium meliloti (Soto et al., 2002), a purple photosynthetic bacterium Rhodospirillum centenum (McClain et al., 2002), Bacillus (Senesi et al., 2002; Dixit et al., 2002; Ghelardi et al., 2002), and also some absolute anaerobic bacteria such as Clostridium species (Macfarlane et al., 2001).
We are interested in unraveling the underlying mechanism of S. marcescens swarming. Previous works have shown that when inoculated on LB medium solidified with 0.8% agar (LB swarming plate) at 30°C, S. marcescens shows a swarming phenomenon where a colony of short motile vegetative rods differentiate at the colony margin into elongated, aseptate and hyperflagellate swarm cells which migrate rapidly and coordinately away from the colony (Alberti and Harshey 1990; Liu et al., 2000). Phenotypically, the process of swarmer cell differentiation and swarming migration behavior may be divided into two separate phases in S. marcescens: (i) the lag period prior to onset of swarming behavior and the induction of swarmer cell differentiation at the colonial edge and (ii) active motileswarming migration (or translocation) from colonial edge. Individual swarmer cells by themselves do not have the ability to swarm. Rather, swarming behavior in Serratia is the result of a coordinated, multicellular effort of groups ofdifferentiated swarmer cells functioning through close cell-cell interactions(Eberl et al., 1999). Interestingly, Based on previous observation, swarming migration does not occurimmediately upon swarmer cell differentiation on LB swarming plates, a process that occurs 1.5 to 2 h postinoculation. Rather, swarming motilityis seen only after ca. 4-5 h (at 30°C) of incubation, long after differentiation and division have produced a visible colony of swarmer cells. Thus, the differentiated cell is only one partof the process. It requires a lag period prior to the commencement of swarming migration, during which propagation and close interaction among bacterial cells and environment occur. Serratia swarming, unlike that of Proteus where swarming colonies exhibit regular concentric zones of consolidation and swarm at a higher agar concentration (2.0%) surface (Allison and Hughes 1991), occurs at a lower agar concentration (0.8%) plate and is a continuous process in which populations of swarm cells continue in migration and result in irregular, dendritic patterns (Givskov et al., 1998; Liu et al., 2000).
Development of a Serratia surface expansion colony requires the sensingand integration of a variety of environmental, cell-to-cell, as well as intracellularsignals involving surface contact and local high population density. According to the current working hypothesis for the developmentof a swarming colony (Givskov et al., 1998), exposure of the cells to surfaceswith a certain viscosity is recognized by an unknown
sensor, andsignal transduction then progresses via the flhDC master operon (Liu et al., 2000) and chemotaxis system (Burkart et al., 1998).Stimulation of flhDC operon initiates swarmer cell differentiationthat involves development of characteristic traits such as cellelongation and multinucleation, and hyperflagellation (Eberl et al., 1996; Liu et al., 2000). The population density is recognized by a N-acyl-homoserine lactone(AHL)-dependentquorum-sensing system constituted by the swrI and swrR genes in Serratia liquefaciens (Eberl et al., 1996; Givskov et al., 1998) and spnI and spnR in S. marcescens (Horng et al., 2002). Expansion of the colony is dependent on a functional swrI or spnI gene; both are membersof the luxI family of autoinducer synthetase genes (Eberl et al., 1996; Horng et al., 2002). TheswrI gene product catalyzes the formation of N-butanoyl-L-homoserine lactone (BHL)(C4) and N-hexanoyl-L-homoserine lactone(C6 )(Eberl et al., 1996), and the spnI gene product mainly catalyzes the formation of C6 and 3-oxo-C6 AHLs (Horng et al., 2002). The signals produced then regulate the SwrR or SpnR activity respectively. Subsequently, production of biosurfactant was stimulated, leading to initiation of swarming or sliding (a flagellum-independent surface migration behaviour, Horng et al., 2002). The flagellar master operon and the quorum-sensing systemare global regulators which control two separated regulons involved in swarming regulation (Givskov et al., 1998).An additional layer of regulation of surface migration may also be exerted through S. marcescens RsmA (an E. coli CsrA homologue and a repressor), which functions as a global regulator and which, when over-expressed, also produces a non-spreadingcolony (Ang et al., 2001). RsmA may inhibit the Serratia spreading through inhibition of quorum-sensing system, which is also reported in P. aeruginosa (Pessi et al., 2001).
Although previous works have unraveled many critical components involved in Serratia spp. swarming behaviour itself, many questions remain elusive. For example, it is not explained why swarming of S. marcescens is strictly temperature-dependent, i.e., when inoculated on a 0.8% LB swarming plate, S. marcescens swarms at 30°C, but this behavior was completely inhibited at 37°C. Furthermore, the potential physiological signals providing critical stimuli for initiation of swarming and the signal transduction system are also not characterized. We observed that S. marcescens under the situation of artificial over-expression of flhDC in the presence of excess amount of biosurfactant still took 4-5 h before initiation of swarming at 30°C and still did not initiate swarming at 37°C(data not shown). This observation together with the fact that cell elongation and flhDC over-expression may be independent of solid surface contact in S. liquefaciens and S. marcescens (Tolker-Nielsen et al., 2000; Lai et al., 1997) suggest that initiation of S. marcescens swarming may not purely be dependent on accumulation of surfactant, flhDC over-expression and formation of
swarmer cells. In addition, even though the importance of AHL signals is clearly identified in many swarming/sliding bacteria including Serratia spp. (Givskov et al., 1998; Horng et al., 2002), we and other colleagues have found that many S. marcescens strains, especially strains that show normal swarming behaviour, do not contain detectable spnRI, swrRI or smaRI AHL quorum-sensing systems (G. Salmond, personal communication; HC Lai, unpublished data). Furthermore, study from another swarming bacterium Salmonella typhimurium (Toguchi et al., 2000) failing to identify any specific swarming signals such as amino acids, pH changes, oxygen, iron starvation, increased viscosity, flagellar rotation, or quorum-sensing autoinducers leads us to consider that there must be some uncharacterized regulatory pathway controlling the behaviour of bacterial swarming in response to the versatile environmental conditions.
In this communication, through mini-Tn5 mutagenesis, we have screened a group of S. marcescens strains that swarmed well at 37°C. One of the mutants was selected for further physiological characterization. We describe a gene that, when mutated, lead to defect in haemolysin production and biofilm formation, and showed a “super-swarming” behaviour: decreased the length of the lag phase prior to swarming migration and swarmed not only faster, but also at higher agar concentration surface. In the process of characterization of this mutated gene and its flanking DNA sequences, we have identified a two-component regulatory system and a partial open reading frame with a protein predicted to be involved in fatty acid synthesis. Interestingly, this mutant appears tofunction to coordinate the initiation of swarming migration independent of flhDC flagellar swimming motility, AHL quorum-sensing system, and production of biosurfactant-- factors already characterized to be involved in the regulation of population surface migration behaviours in many bacterial species including S. marcescens (Horng et al., 2002; Liu et al., 2000) and S. liquefaciens (Lindum et al., 1998).
Here we present several lines of evidences to show that regulation of initiation of swarming is closely correlated to the homeoviscous acclimation of bacterial cell membrane, i.e., acclimatizing response of cell membrane lipid in response to different environmental conditions (Browse and Xin 2001). We present evidences that pattern of cellular fatty acid profile is affected by environmental temperature and also saturated fatty acids (SFAs) through a common mechanism. To swarm or not-to-swarm is determined by pattern of fatty acid profile which acts through a two-component signal transduction system. In addition, we demonstrate that expression of fabGSm, a NADPH-dependent 3-ketoacyl-ACP reductase, haemolysin production and composition of cellular fatty acid was further regulated by this two-component signal system. Evidence also suggested that cell surface topology and
extracellular polysaccharide (EPS) production which originally functions as a lubricant during swarming of many bacteria including S. typhimurium (Mireles et al., 2001) and P. mirabilis (Gygi et al., 1995) might also play a role in the regulation of Serratia swarming. A similar SFA effect on the swarming of P. mirabilis and S. typhimurium was also observed, suggesting that this regulation is a common phenomenon. Thus, a regulatory loop composed of the nutrients condition (especially the SFAs), environmental temperature, patterns of fatty acid profile, the two-component signal transduction system, fatty acid synthesis gene and cellular fatty acid synthesis, haemolysin production and amount of EPS produced provides a novel mechanism for the control of swarming behaviour in S. marcescens.
Results
A Serratia marcescens mutant defective in the temperature and temporal control of swarming migration
At 30°C, S. marcescens CH-1 cells exhibited typical swarming behaviour on LB medium containing 0.8% agar (LB swarming plate)(Figure 1A) and reached the edge of standard 9 cm diameter plates after 12h culture. In contrast, while growth dynamics of S. marcescens CH-1 were almost identical at 30 and 37°C, bacteria inoculated onto identical plates incubated at 37°C did not initiate swarming even after 48h culture and formed small defined colonies (Figure 1A).
To characterize any genetic determinants involved in the temperature-dependent regulation of swarming behaviour in S. marcescens CH-1, mini-Tn5 transposon mutagenesis was performed to screen for S. marcescens CH-1 colonies that swarmed at 37°C on modified LB swarming plates. A total of 17 colonies out of 6000 transconjugants were selected to show such a temperature-independent swarming behaviour. One of the mutants, named S. marcescens WC100 was selected for further characterization. When incubatedon a normal 0.8% LB swarming agar plate [the same result was obtained from either Bacto agar (Difco, USA) or Eiken-agar (Eiken, Japan); Eiken agar was chosen for subsequent experiments] at 30°C, S. marcescens WC100 showed a more progressive translocation acrossthe agar surface compared with its isogenic parent S. marcescens CH-1. This behavior resulted in a colony that moved out farther than CH-1 over a given time. Following the swarming fronts of S.
marcescens WC100 and CH-1 cells at 30°C bythe method of Gygi et al. (1995), the
swarming behavior and velocity of WC100 and CH-1 were monitored, and were shown in Figure1. Conspicuous differences were observed, including the difference in time spent during the initial lag phase, the swarming velocity after initiation of swarming, and the ability of cells to swarm on higher agar concentration in the swarming plates. Depending on the wetness of plates, CH-1 cells typicallyspent 4-5 h (at 30°C) in the initial lag phase prior tothe onset of swarming migration, while WC100 spent ca. 1 hour less in this phase. After initiation of swarming, S. marcescens WC100 swarms at a constitutively increasing velocity, which is significantly faster than that of CH-1 cells (Figure1C). The average velocity of S. marcescens CH-1 swarming increased from 1 mm/h (between 5 to 6 h) to 3.3 mm/h (between 7 to 8 h), while S. marcescens WC100 increased from 2 mm/h to 5 mm/h at the same period (Figure1C). By spending less time in the initial lag period and swarming faster than the CH-1 cells, WC100 thus translocates farther in a given time than do the CH-1cells. S. marcescens WC100 was further observed to swarm even if the agar concentration was increased up to 1% at 30°C, where CH-1 did not (Figure1B).
As the incubation temperature is shifted from 30 to 37°C, CH-1 cells typicallyare unable to initiate swarmingbehavior in the LB swarming plate (Figure 1A). WC100 still swarmed well and behaved similarly to that at 30°C in the time of swarming initiation, average swarming velocity (Figure 1C), and the ability of swarming at 1% agar concentration (data not shown), suggesting that the temperature-dependent regulation of swarming behaviour was abolished in the mutant. We refer to this unusual swarming phenotype as "super swarming". To see whether the mutation resulting in super-swarming extends to other phenotypes, some physiological assays including growth rate and production of the red pigment prodigiosin were performed. While there was no significant difference in the growth dynamics during growth on plates, WC100 cells were about 10% shorter under 1000x microscopy and produced less prodigiosin than CH-1 cells at 30 and 37°C (data not shown).
A mutation in a locus encoding RssA, a member of the two-component family of sensory proteins, leads to the superswarming phenotype
Identification of the S. marcescens WC100 DNA flanking the mini-Tn5 insertion site was accomplished by conventional digestion and cloning ofthe DNA flanking the transposon followed by sequencing with primers designed from within the I end or O end of the transposon, respectively (de Lorenzo and Timmis, 1994). The result revealed that the mini-Tn5 insertiongiving rise to the super-swarming phenotype was locatedwithin a 3003-bp DNA fragment of the S. marcescens CH-1 genome (Figure 2). Sequencing revealed that mini-Tn5 had inserted in an 1179bp open reading frame (named ORF1), potentially encoding a 393-amino-acid (43.94 KDa) polypeptide with a calculated isoelectric point of 5.45. Upstream of orf1 was orf2 identified, potentially encoding 214-amino-acid in the same direction. Upstream of orf2 was a divergent partial orf3 identified, potentially encoding an N-terminus of 109 residues (Figure 2A).
The deduced protein sequences were compared with non-redundant protein sequence databases using blastn or blastx via the NCBI internet homepage. The mini-Tn5 insertion was found to be in the S. marcecens genome in a region that is highly homologous to the bacterial two-component signal transduction system region (Kottayil 2002). Computer-assisted homology searches using BLASTP (Altschul et al., 1997)of protein sequences stored in the DDBJ/EMBL/GenBank databanks indicate that Orf1 is homologous to elements in the two-component sensor family, including
Vibrio harveyi LuxQ(22% identity, 41% similarity), E. coli EvgS (22% identity,41%
similarity) and E. coli RcsC (19% identity, 39% similarity). Orf2 is also homologous to two-component response-regulator elements, including a 38 % identity and 58% similarity throughout the alignment to a putative two-component response regulator of
Ralstonia solanacearum (Salanoubat et al., 2002). Partial ORF3 shows high homology to members of the short-chain dehydrogenase/reductase family, including 62% identity out of the 109 N-terminal amino acids to YgfF of E. coli (Blattner et al., 1997). We have chosen to call the mutated gene (orf1) rssA, forregulation of Serratia swarming. orf2 was named rssB, and orf3 named fabGSm. The nucleotide sequence of 3003-bp encoding the three ORFs(rssA, rssB and the 5' portion of fabGSm) has been submitted tothe DDBJ/EMBL/GenBank databases under accession no. AF465237. Analysis of the deduced amino acid sequence of RssA, together with the domain search by MotifScan (Falquet et al., 2002) and hydrophobicity characteristic analysis by DAS (Cserzo et al., 1997) (Figure 2B), TMpred (Hofmann and Stoffel 1993), SOSUI (Mitaku and Hirokawa 1999) and PredictProtein (Columbia University, USA) from the ExPASy proteomics tools internet homepage (http://tw.expasy.org) indentified a hydrophobic region between P90and L112, which indicates thatit may form a membrane helix and localized in the inner membrane as a transmembrane protein (Figure 2C).The N-terminal sequence of RssA was predicted to stretch into the periplasmand was not predicted to contain signal peptide (Figure 2C).A HAMP domain which plays an active role in transmembrane signal transduction (Appleman and Stewart 2003) was identified between 90(D) and 158(A), a phosphoaccepter domain was found between 162(R) and 227(E) near the middle of the protein in the cytoplasm, and an ATPase domain was observed between 272(T) and 382(S) near the C-terminal (Figure 2C). RssA has strong motif similarity to many other sensory proteins thatare members of the two-component family of proteins (Hoch and Silhavy 1995). Specifically, as shown in Figure 2D, domains, suchas the H, N, D/F and G blocks that are associated with histidine kinases (Hoch and Silhavy 1995) are maintained withinRssA. Therefore, RssA may function as a typical sensory protein of histidine kinase in S. marcescens CH-1.
To confirm that the S. marcescens WC100 mutant phenotype was not due to artifact effect from introducing a copy of mini-Tn5 transposon, rssA in CH-1 was further mutated by insertion deletion through homologous recombination to form S.
marcescens CH-1∆A. The phenotypes of both S. marcescens WC100 and CH-1∆A
were characterized, including growth dynamics, prodigiosin synthesis, cell density and temperature-regulated swarming behaviours and haemolysin production (see later in the text). No difference was observed between WC100 and S. marcescens CH-1∆A. Plasmid pWC200(pACYC184::rssA) was further transformed into both WC100 and CH-1∆A at 37°C. In both strains, super-swarming behaviour was restored (data not shown), confirming that rssA was the gene responsible for the super-swarming phenotype. S. marcescens CH-1∆A was selected for further study. The effect of over-expression of rssA on swarmercell differentiation and behavior was examined
by transforming pWC200 into S. marcescens CH-1. When rssA is overexpressedfrom a multicopy pWC200 at 30°C, it delayed the initiation of CH-1 swarming for about 1 h, and also reduced its swarming velocity for about 40%. Plasmid-onlycontrols have no significant effect on the time of swarming (data not shown). The data suggest that RssA may function as a negative regulatorof swarming such that increased levels of the RssA protein increases the lag phase before swarming and result in a delayed, aberrant swarming phenotype.
RssA senses populational density and temperature for controlling initiation of swarming
Although CH-1 does not swarm at 37°C, the onset time of S. marcescens CH-1 swarming behavior is dependent on the population density at 30°C (Lai et al., 1997), such that thetime to swarming migration increased as the cell density decreased. One possible role for RssA in the regulation of initiation of swarming could be as a sensor of population cell-density, which, when defective, leads to super-swarming behaviour. If RssA senses the population density or degree of cell-to-cell contact, defect in RssA should alter the ability of cells to detect changes in the populationsize and may change the time of initiation of migration. To testthis, we compared the timing of swarming initiation of CH-1∆A cells to that of CH-1 at 30°C. In a series of experiments, we tested whether the initiationof CH-1∆A swarming behavior was still correlated with population density at 30°C. As similar to many other enterobacteria (Fraser and Hughes 1999), the flhDCSm is the master operon of flagellar regulon regulating the flagellar motility during swarming in S. marcescens (Liu et al., 2000), we chose to use the flhDCsm promoter activity as a reporter for S. marcescens swarming. In these experiments, the initial inoculum’s concentration of CH-1∆A(pPC300) and also the control strain S. marcescens CH-1(pPC300) was varied in 10-fold increments from 1 × 106 to 1 × 102 cells (CFU) delivered in 5-µl-aliquot droplets to the LB swarming agar surface. The light emission of swarming bacteria which indicates the promoter activity of flhDCSm master operon (Liu et al., 2000) was monitored hourly by X-ray film exposure following the growth of bacterial colonies on LB swarming plates and CH-1∆A(pPC300) swarming activity was shown in figure 3A. The number of each bacterial colony expressed as colony forming units (CFUs) was at the same time calculated by 10 times serial dilution and plate counts. Similar to CH-1(pPC300), at the earlier stage of CH-1∆A(pPC300) culture where no swarming was initiated, light emission could be detected from the whole colony. Once the cells started to swarm, light emission could only be detected at the swarming edge of the colony, where elongated cells are actively migrating outwards (Figure 3). Thus the light emission pattern reflected the real time colonial
swarming behavior. Although initiation of CH-1∆A(pPC300) swarming is stilldensity dependent, it started to swarm earlier than CH-1(pPC300) cells at a much lower cell density. CH-1∆A(pPC300) only requires ca. 50-fold fewer cells to commence migration (3.4 × 106 versus 1.7 × 108 cells to initiate swarming) (Figure 3B). Further experiments showed that CH-1∆A(pPC300) also behaved similarly in a cell-density dependent swarming phenomenon at 37°C (data not shown). In summary, although initiation of S. marcescens CH-1∆A swarming is still density-dependent, it reproducibly initiates swarming at a lower cell-density, suggesting rssA indeed is involved in the cell-density dependent regulation of swarming behaviour. The phenomenon that the rssA mutant swarming behaviour was not affected by temperature-upshift (from 30 to 37°C) further suggested that RssA also acts as a temperature-dependent sensor regulating the swarming behaviour of S. marcescens CH-1.
Swimming motility and biosurfactant production are not affected in rssA mutant Factors already identified to be essential for or involved in the regulation of swarming process in Serratia (Liu et al., 2000; Horng et al., 2002; Givskov et al., 1998; Harshey 1994) were examined to see whether expression of any of these factors were aberrant, leading to the super-swarming behaviour in S. marcescens CH-1∆A. These include the swimming motility, amount of flagellum produced and production of surfactant.
We first checked whether the swimming motility was significantly activated in CH-1∆A. Swimming assay and monitoring of average swimming velocity using CH-1 and CH-1∆A cells at both 30 and 37°C were performed. The result in Figure 4A and 4B shows that CH-1 and CH-1∆A swimmed in a similar scale at both 30 and 37°C. Therefore, this mutation does not appear to affect flagellar rotation and motility. Further monitor of flhDCSm promoter activity by pPC300 (Figure 4C) and SDS-PAGE analysis for quantification of flagellum production (data not shown) also failed to show difference between CH-1 and CH-1∆A at 37°C, suggesting that the flhDCSm regulon is not affected by rssA mutation.
We further supposed the CH-1∆A super-swarming phenotype may be due to over-production of biosurfactant reducing the surface friction of swarming plates. Drop-collapsing test (Horng et al., 2002) was performed to qualitatively assay the surface tension of stationary phase culture suspensions from CH-1∆A and CH-1 cells at both temperatures. Again, even after repeated tests, no difference was observed (Figure 4D). For confirmation of the assay, TLC (Thin Layer Chromatography) (Horng et al., 2002) was performed to quantify the biosurfactant production to see whether there is a minor difference between the two strains. Still, no difference was observed (Figure 4E), suggesting that biosurfactant production is not affected in
CH-1∆A.
As CH-1∆A was screened from modified LB agar medium that contains a lower NaCl concentration [0.04% (w/v)] compared with the normal LB agar salt concentration [1% (w/v)], together with the fact that the predicted gene products of almost all transposon–disrupted mutant genes from the screened super-swarming mutants are closely associated with cell envelope components (data not shown), we then hypothesized that cell envelope integrity might be less compact in the rssA mutant. To see whether this is possible, we grew CH-1 and CH-1∆A for overnight in normal LB broth media before transferring the bacterial cells into normal [1% NaCl], high [1.5% NaCl] and low [0.04% NaCl]-osmolarity LB broth media at both 30 and 37°C and monitored their growth dynamics by measuring the optical density (A600nm).
S. marcescens CH-1∆A was shown to grow with a generation time similar to that of
the CH-1 cells under all the conditions assayed (data not shown). This result suggests that the RssA was not involved in osmolarity regulation in CH-1. Taken together, Our results suggest that some novel factors other than the factors already identified are involved in the regulation of S. marcescens CH-1 swarming behaviour, which, when defective, leading to the super swarming phenotype.
Biofilm formation and haemolysin production are regulated by RssA in a temperature-dependent way
Previous works have shown that biofilm formation is closely related to swarming in many bacterial species including Salmonella typhimurium (Mireles et al., 2001), P. aeruginosa (Deziel et al., 2001) and B. cepacia (Huber et al., 2001). We reasoned that biofilm formation might also be aberrant in the CH-1∆A super-swarming mutant. To validate this, the microtiter well assay (O’Tolle and Kolter, 1998) which monitors the ability of S. marcescens to attach to the wells of microtiter dishes was used to quantify the biofilm formation of CH-1 and CH-1∆A at both 30 and 37°C. We found that biofilm-forming ability of CH-1 was higher at 30°C(Figure 5A), with an average absorbance value of 0.34 was detected, and was reduced at 37°C(Figure 5A), where an average value of optical density at 0.28 was detected (Figure 5B). Biofilm formation for CH-1∆A is significantly defective (figure 5A), reaching only about 0.20 (59% of CH-1 level) at 30°C and 0.12 (43% of the CH-1 level) at 37°C (Figure 5B). The data suggest that biofilm-forming ability of S. marcescens CH-1 is altered under different environmental temperatures; with 30°C being more proficient than 37°C, and CH-1∆A is less proficient in binding to the microtiter plate surface at both temperatures, confirming that biofilm formation is related to swarming behaviour under the control of RssA.
during swarming of CH-1∆A may be aberrant compared with CH-1. To confirm this, swarming assays were performed at 30 and 37°C for CH-1 and CH-1∆A on LB swarming plates and the swarming edge of the bacterial colonies were observed closely under 400x and 1000x phase contrast microscopy at both temperatures. Compared with CH-1∆A, CH-1 did not swarm at 37°C and the cells looked significantly non-active near the colonial edge, where the way of cell alignment was similar to the sliding edge of S. marcescens SS-1 (Horng et al., 2002). Similar cell alignment and motility were observed near the swarming edge of CH-1 and CH-1∆A at 30°C (data not shown). No other significant differences were observed between the two strains, including colonial texture organization (from short, vegetative and still cells in the colonial center, gradually outwards to the vegetative, actively turning cells in the middle, then to the medium-elongated, also actively moving cells near the swarming edge, finally to the very much elongated swarming cells in the swarming fronts) and the ratio of pigment to non-pigment producing cells under the 400x phase contrast microscopy (data not shown).
The cell-surface associated haemolysin is identified to be a dominant virulence factor in S. marcescens (Kurz et al., 2003). Previous works have shown that expression of some virulence factors including haemolysin is closely coupled to swarming in P. mirabilis (Fraser et al., 2002). It is also shown that production of hemolysin is under the control of bacterial two-component systems in Bordetella pertussis (Stibitz 1994) and Staphylococcus aureus (Fournier et al., 2001). We decided to determine whether production of haemolysin was affected in CH-1∆A. To this end, haemolysin activity was assayed in CH-1 and CH-1∆A. Equivalent number of CH-1 and CH-1∆A cells were harvested from LB seeding agar plates at 30 and 37°C at an hourly interval following the growth. It was clearly observed that the haemolysin activity of CH-1∆A was significantly higher than that of CH-1 at both 30 (320 %) and 37°C (250%) (Figure 5C). To see whether the regulation occurs at the transcriptional level, a recombinant plasmid, pSA400 (PshlA::luxCDABE) was constructed as a reporter for the promoter activity of PshlA. A comparison of the light emission patterns from CH-1∆A(pSA400) and CH-1(pSA400) showed an average 4 to 8 fold increase in shlA promoter activity in CH-1∆A(Figure 5D), indicating the negative regulation effect of RssA on the promoter activity of shlA. Together, these data indicate that in S. marcescens CH-1, a complicated two-component regulatory network existing in S. marcescens, in which biofilm formation and haemolysin virulence factor gene, together with some uncharacterized swarming-related genes are components.
dose-dependent way
Although CH-1 does not swarm on LB swarming plate at 37°C, interestingly we found that CH-1 did swarm well on defined MGM plate (M9 minimal growth medium containing 0.8% Eiken agar) and MGM plate containing casamino acids(1%), not only at 30, but also 37°C (Figure 6A), suggesting that higher environmental temperature is not a must condition for inhibition of S. marcescens CH-1 swarming. We further observed that CH-1 started to swarm almost immediately (within 1 hour, at a much lower colonial cell density) after it was transferred from saturated LB broth culture to MGM plate or MGM-casamino acids (1%) plate at both temperatures. This observation is striking when compared with the cell-density dependent CH-1 swarming behaviour on LB swarming plate, indicating that absence of some contents in the minimal media lead to the immediate CH-1 swarming. Further swarming assays on MGM-casamino acids (1%)/yeast extract (1%) plate showed that while CH-1∆A swarmed, CH-1 swarming was completely inhibited at 37°C (Figure 6A). These observations suggested that some components (or the metabolic molecules derived from the components) contained in yeast extract inhibited CH-1 swarming at 37°C, and existence of the component(s) did not inhibit CH-1∆A swarming. We hypothesized that some amino acids contents in excess might inhibit the CH-1 swarming at 37°C. Using MGM-casamino acids (1%) swarming plate as the basal medium, the 20 single amino acids including the branch chain amino acids isoleucine, leucine and valine, and glutamine, asparagines, histidine, lysine, phenylanaline, threonine, tryptophan, valine, tyrosine, serine, proline, glycine, glutamine, glutamate, cysteine, aspartate and alanine at the final concentration of 1% was separately incorporated, followed by swarming assay at 30 and 37°C. Comparatively, a similar swarming pattern was observed on the swarming scale between CH-1 and CH-1∆A when compared with those on MGM-casamino acids (1%) plates (data not shown), suggesting that individual amino acid in excess did not have effect on the inhibition of swarming of CH-1. While changes of other culture conditions including the concentration of glucose, addition of other mono- or di-saccharides, iron starvation, medium pH…etc. did not lead to change of CH-1 swarming behaviour (data not shown), we continued to determine whether exogenous saturated or unsaturated fatty acids with different chain length have some effect on the CH-1 swarming. Saturated fatty acids (SFAs) such as lauric acid (C12:0), myristic acid (C14:0), palmitic acid (C16:0) and stearic acid (C18:0) and unsaturated fatty acids (UFAs) such as myristoleic acid (14:1 ∆9), palmitoleic acid (16:1 ∆9) and oleic acid (C18:1 ∆9) at a final concentration of 0.01% (w/v) were added separately into MGM-casamino acids (1%) plates followed by swarming assays for CH-1 and CH-1∆A at 37°C. We have found that while there were not so significantly different swarming-inhibition effect
from the UFAs tested, S. marcescens CH-1 swarming was completely inhibited by addition of lauric acid and myristic acid, and slightly inhibited by the presence of palmitic acid and stearic acid (Figure 6A). All fatty acids tested did not show significant inhibitory effect on CH-1∆A swarming (Figure 6A). To further confirm whether this regulatory phenomenon was dose-dependent, myristic acid at the concentration of 0.01%, 0.005%, 0.0025% and 0.00125% (w/v) was added into MGM-casamino acids plates, followed by swarming assays at 37°C. The experiments showed that the time spent before initiation of swarming of CH-1 was delayed as the concentration of myristic acid was increased and that swarming was completely inhibited by myristic acid at the concentration of 0.01% (w/v) (Figure 6B). A similar myristic acid inhibitory effect was also observed at 30°C on LB swarming plates or MGM-casamino acids plates, and lauric acid also showed a similar inhibitory trend (Figure 6A). Taken together, these data indicate that, in addition to temperature shift, initiation of swarming is regulated by the availability of SFAs in a concentration-dependent manner under a constant temperature.
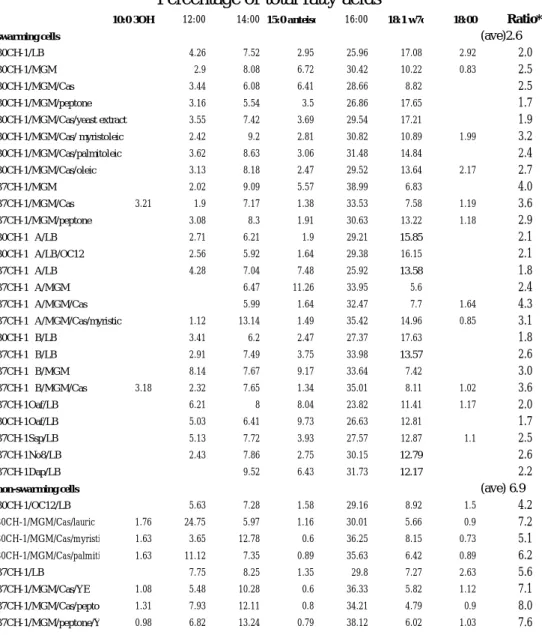
Swarming behaviours are closely related to “swarming” and “non-swarming” cellular fatty acid profiles
The findings that SFAs inhibit CH-1 swarming and fabGSm identified upstream of rssA and rssB was predicted to encode a protein with high amino acid identity to members of short-chain dehydrogenase/reductase family, which basically were NADPH-dependent 3-ketoacyl-ACP-reductases involved in the fatty acid synthesis during the elongation step (Magnuson et al., 1993), together with the fact that the membrane fluidity determined by composition of membrane fatty acids is optimized by a DesK-DesR two component pathway in B. subtilis (Aguilar et al., 2001), we reasoned that the cellular fatty acid profile of CH-1 might be affected by environmental nutrients contained in the culture media, which may then affect the conformation of RssA and subsequently regulate the signal transduction of RssA-RssB two-component pair, leading to either swarming or non-swarming phenotypes. To this end, cellular fatty acid profile expressed as percentage of total fatty acids of CH-1 cultured at 37°C on MGM plates, MGM casamino acids(1%) plates, MGM-peptone(1%) plates (swarming initiated), LB plates, MGM-casamino acids(1%)/yeast extracts (1%) plates, and MGM-peptone(1%)/yeast extracts (1%) plates (swarming inhibited) was determined by fatty acid extraction followed by fatty acid analysis by MIDI gas chromatography system. The major changes observed in fatty acid profile from cells grown on non-swarming conditions were the significant increase in the composition of lauric acid (12:0) and then myristic acid (14:0) and palmitic acid (16:00) (Figure 6C; Table 1). For cells grown in the swarming condition,
a concomitant increase in unsaturated fatty acids (especially 18:1 w7c) and branch-chain fatty acids (especially 15:0 anteiso) were observed (Figure 6C; Table 1). The results highlight the importance of exogenous fatty acids influcing the composition of cellular fatty acids and swarming behaviour at a constant temperature in S. marcescens.
The changes in fatty acid profile under different nutrient conditions at 37°C that coincide with the swarming or non-swarming behaviour of CH-1 reinforced our hypothesis of a direct correlation between fatty acid profile and swarming regulation. To this end, we collected CH-1 swarming cells and non-swarming cells cultured under different growth conditions for fatty acid profile analysis. CH-1 swarming cells were collected from LB swarming plates (30°C), MGM-casamino acids (1%)/yeast extract(1%) plates (30°C), MGM-casamino acids(1%)-myristoleic acids (0.01%) (30°C), MGM-casamino acids(1%)-oleic acids (0.01%)(30°C), MGM-casamino acids(1%)-palmitoleic acids (0.01%)(30°C), MGM plates and MGM-casamino acids (1%) plates (30 and 37°C). For CH-1 non-swarming cells, cells were collected from MGM-casamino acids(1%)-lauric acid (0.01%) plate (30°C), MGM-casamino acids(1%)-myristic acid (0.01%) plates(30°C), MGM-casamino acids(1%)-palmitic acid (0.01%) plates(30°C), LB swarming plates(37°C), and MGM- casamino acids(1%)/myristic acid(0.01%) plates(37°C). The results were summarized in Table 1. We found a significant trend that for CH-1 cells growing at non-swarming conditions including either higher incubation temperature or nutrients containing SFAs, the ratio of high melting point fatty acids to low melting point fatty acids (12:0+14:0+16:0+18:0/10:0 3OH+15:0 anteiso+18:1 w7c) is much higher than that from cells growing at swarming conditions (Table 1), with the percentage of 12:0 showing the most significant difference (Figure 6C). On the contrast, CH-1 cells growing at swarming conditions, including lower incubation temperature or media that do not contain SFAs, contain higher percentage of UFAs and branch-chain fatty acids, with 18:1 w7c and 15:0 anteiso contents showing the most significant difference (Figure 6C).
The fatty acid profiles of the two super-swarming mutants CH-1∆A and CH-1∆B (see later in the text) inoculated at either on LB swarming plates (30 or 37°C), MGM, MGM-casamino acids, or MGM-casamino acids-myristic acid(0.01%) (37°C), similar to some other super-swarming mutants including no. 3, 6, and O3 on LB swarming plates at 30 or 37°C, also contained a similar swarming fatty acid profile (Table 1). In conclusion, we have presented a clear evidence that S. marcescens CH-1 cellular fatty acid profile is affected by environmental temperature and fatty acids, and that the cellular fatty acid profile is closely related to swarming phenotypes, where swarming and non-swarming cells show a different pattern, with swarming cells containing
relatively more UFAs and branch-chain fatty acids and non-swarming cells containing more SFAs. We classify the two fatty acid patterns into “swarming” and “non-swarming” profiles for subsequent studies.
Expression of fabGsm is activated by myristic acid
Effect of myristic acid on fabGSm promoter activity was evaluated. A recombinant plasmid pSA401 (PfabGSm::luxCDABE) in which the fabGSm promoter ligated in front of the luxCDABE reporter was constructed. pSA401 was electroporated into CH-1 followed by monitoring the bioluminescent activity in LB broth culture with and without 0.01% (w/v) myristic acid at 30°C. While grown in LB broth, fabGSm promoter activity in CH-1 reached its peak soon at about 30 minutes while cells were still growing at lag phase after it was transferred from LB over-night broth culture into fresh LB broth containing myristic acid 0.01% (w/v), decreased rapidly thereafter as cells are growing into log phase and maintained at a very low level as cells were grown into stationary phase (Figure 7A). Addition of myristic acid activates fabGSm promoter activity up to 2-fold, followed by a similar decrease pattern as cultured in LB broth (Figure 7A). The data suggested that fabGSm promoter responds very sensitively to myristic acid in LB medium.
RssA and RssB down-regulate fabGSm expression
To see whether fabGSm expression was regulated by RssA, a recombinant plasmid pSA401 (PfabGSm::luxCDABE) in which the fabGSm promoter ligated in front of the luxCDABE reporter was constructed. pSA401 was electroporated into CH-1 and CH-1∆A followed by monitoring the bioluminescent activity in LB broth culture containing 0.01% (w/v) myristic acid at 30°C. Compared with CH-1 fabGSm promoter activity which decreased rapidly, fabGSm promoter in CH-1∆A was maintained at a much higher level for longer time for 2 hours followed by slower reduction (figure 7A). Northern blot hybridization using partial fabGSm as the probe further confirmed this phenomenon (Figure 7B). Briefly, a clear trend was observed that when growing in a culture condition containing myristic acid, the fabGSm promoter activity in CH-1 was activated and was rapidly down-regulated. Such an effect was not observed when
rssA was mutated, suggesting that expression of fabGSm was inhibited by RssA in
response to effect of myristic acid.
We hypothesized that the reason why CH-1∆A swarmed under all the culture conditions tested may be due to blockage of phosphorelay transfer to the transcriptional regulator RssB, albeit the fatty acid profile varied under different culture conditions. To test whether rssB was involved in regulation of swarming behaviour and fabGSm expression, we constructed a strain S. marcescens CH-1∆B, which contains a streptomycin resistance gene (SmR) cassette interrupting the rssB
gene. After confirmation of the mutated rssB genotype by Southern blot hybridization (data not shown), CH-1∆B was subject to swimming assay and the swimming velocity and flhDCSm promoter activity following the growth in LB broth culture were measured. While there is no significant difference observed compared with S.
marcescens CH-1(Figure 4B; 4C), CH-1∆B was further assayed for surfactant
production, where still no difference was observed compared with that of CH-1 (data not shown). CH-1∆B was tested for swarming motiltiy under different physiological culture conditions including LB swarming plates, MGM plates, MGM-casamino acid(1%) plates, MGM-gelatin(1%) plates, MGM-gelatin(1%)/yeast extract(1%) plates, and MGM-casamino acids plates containing a series of 0.01% (w/v) fatty acids (C12, C14, C16, C18, C14:1 ∆9, C16:1 ∆9, and C18:1 ∆9) at both 30 and 37°C. CH-1∆B showed an unique “super-swarming” phenotype under all conditions tested, i.e., similar to CH-1∆A, CH-1∆B swarmed on all plates tested at both temperatures. CH-1∆B swarming behaviour on LB swarming plates was shown in Figure 1A. Although CH-1∆B only showed slight decrease in biofilm formation (Figure 5A), it was observed to swarm much faster than CH-1 (when swarmed) but slower than CH-1∆A. The result of comparison of average swarming velocity among CH-1, CH-1∆A and CH-1∆B on LB swarming plates was shown in Figure 1C. On 1% LB swarming plates at 30°C, CH-1∆B migrated farther than CH-1, but less than CH-1∆A within a given time of 9 hr incubation (Figure 1B). The data suggest that RssB may act as a negative regulator involved in the regulation of CH-1 swarming.
Expression of fabGSm may also be regulated RssB. To confirm this, myristic acid at the concentration of 0.01% (w/v) were added into LB broth and the intensity of light emission from S. marcescens CH-1(pSA401) and CH-1∆B(pSA401) was monitored following the growth at 30°C. Similar to CH-1∆A, effect of myristic acid on the fabGSm expression was aborted when rssB was mutated (Figure 7A). Northern blot analysis using partial fabGSm as the probe further confirmed the observation (Figure 7B). In conclusion, we have shown a strong evidence that RssA and RssB are both involved in a signal transduction regulatory pathway negatively regulating swarming and fabGSm expression in response to environmental nutrients, especially SFAs.
Previous work has shown that overexpression of response regulators in the absence of their cognate kinases could result in constitutive expression of the target gene(s) controlled (Powell and Kado 1990). This suggested that high concentrations of unphosphorylated response regulator could bind in vivo to target promoter and caused unregulated transcription. While we reasoned that phosphorylated RssB may act as an negative regulator, an over-expression of rssB without the assistance of RssA might rapidly down-regulate transcription of fabGSm and inhibit CH-1∆A
swarming at 30°C, pSA402(pBCSK::rssB) was transformed into S. marcescens CH-1∆A followed by swarming assay and monitoring the expression of fabGSm. Initiation of swarming in CH-1∆A(pSA402) was significantly delayed for about 2h compared with CH-1∆A containing control plasmid (data not shown). Further experiments using pSA401 as the reporter for fabGSm expression showed that fabGSm promoter activity was decreased up to 2-fold when rssB was over-expressed. These experiments demonstrate that high production of RssB down-regulates fabGSm expression and inhibits swarming without the assistance of RssA at 30°C. This result therefore agrees with the observation that unphosphorylated response regulators can still regulates transcription when they are overexpressed (Powell and Kado, 1990).
RssA-RssB regulate cellular fatty acid composition
As expression of fabGSm was regulated by RssA-RssB in response to different environmental culture conditions, and FabGSm was predicted to be involved in the cellular fatty acid synthesis, we reasoned that pattern of cellular fatty acid profile is regulated by the RssA-RssB two-component system. The fatty acid composition of CH-1, CH-1∆A and CH-1∆B grown on LB plates at 30 and 37°C was analysed by gas chromatography and results were shown in Table 1. Although CH-1, CH-1∆A and CH-1∆B all show a swarming fatty acid profile at 30°C, the fatty acid composition of these 3 strains was different from each other (Table 1). The fatty acid profile of CH-1 cultured at 37°C, which showed a typical non-swarming profile, is also significantly different from those of CH-1∆A and CH-1∆B at 37°C, which showed a swarming profile (Table 1). Further experiments analyzing the fatty acid profile of the three strains also showed vary from each other when cultured on MGM media at 37°C (Table 1). These data indicate that in accordance with swarming, cellular fatty acid profile is regulated by RssA-RssB two component system.
3-oxo-C12 AHL signal inhibits swarming and modulates fatty acid profile
To study the role of quorum sensing in the regulation of swarming motility in S. marcescens, we began work with identification of the quorum sensing signals produced by S. marcescens CH-1. Unexpectedly, we discovered that this strain was unable to produce AHLs capable of activating biosensors based upon LuxR, CviR, AhyR or LasR in T-streak assays and in well assays after dichloromethane extraction of spent culture supernatants (data not shown). To exclude the possibility that inhibitory compounds were present in the crude dichloromethane extract we subjected the extract to fractionation by both HPLC and TLC and assayed using well assays or biosensor overlays respectively. In both cases no AHL activity was detected (data not shown). As further evidence for the absence of a LuxRI/AHL-based quorum sensing system in S. marcescens CH-1, DNA/DNA hybridisation in a Southern blot failed to
identify any homologous DNA to the spnTIR genes of S. marcescens SS-1 (Horng et al., 2002).
Previously we have found that long-chain AHLs have inhibitory effect on the sliding behaviour of S. marcescens SS-1 (Horng et al., 2002). Although S. marcescens CH-1 did not produce detectable AHLs, we hypothesized that long chain AHL signals produced from other sources may inhibit CH-1 swarming. To confirm this, CH-1 was subject to swarming assay on LB swarming plates containing a range of synthetic AHLs incorporated into the medium at the final concentration of 10 µM at 30°C. Figure 8A shows while all the short chain AHLs tested did not inhibit swarming, a clear and inhibitory effect of some specific long chain AHLs upon swarm plate motility was observed. Among the long chain AHLs tested, 3-oxo-C10, and especially 3-oxo-C12 and 3-oxo-C14 inhibit the CH-1 swarming most significantly, and addition of these signals did not significantly inhibit the swarming of CH-1∆A (data not shown), suggesting that the inhibitory effect of the long chain AHLs might act through change of membrane fatty acid profile and RssA-RssB signaling system. To see whether this is possible, the long chain AHL signal 3-oxo-C12 at the concentration of 10 µM was incorporated into the LB swarming media followed by analyzing fatty acid profiles of CH-1 and CH-1∆A that are seeding-plate cultured at 30°C. Results in Table 1 showed that a clear difference was observed on the fatty acid profile between the two strains, which was concordant with swarming behaviours. While CH-1 showed a swarming fatty acid profile in the absence, and a non-swarming fatty acid profile in the presence of 3-oxo-C12 at 30°C, CH-1∆A showed a similar swarming fatty acid profile irrespective the presence of 3-oxo-C12. This observation suggests that although CH-1 does not bear AHL quorum sensing system, 3-oxo-C12 does regulate CH-1 swarming and such a regulation might act through affecting the cellular fatty acid profile.
We continue to see whether expression of fabGSm was affected by 3-oxo-C12. The light emission assay using pSA401 as a reporter was performed in LB broth culture containing 10 µM 3-oxo-C12 at 30°C. fabGSm promoter activity in CH-1 was only slightly affected by the presence of 3-oxo-C12 (Figure 8B; C). Compared with CH-1, CH-1∆A and CH-1∆B, we also did not observe a significant effect of 3-oxo-C12 on fabGSm promoter activity in mutant strains (Figure 8B; C). A similar result was obtained using pSA400 as a reporter for monitoring the effect of 3-oxo-C12 on shlA expression (data not shown). In brief, although 3-oxo-C12 does inhibit CH-1 swarming and affect cellular fatty acid profile, it seems not to regulate fabGSm and shlA through RssA-RssB signaling pathway.
The detailed cellular surface topography of CH-1, CH-1∆A and CH-1∆B growing in the log phase was studied using atomic force microscopy (AFM) by systematically zooming in on target cells and regions within the cell envelope. A larger scan area (3 x 3 µm2) was first made in order to select the desired cells for higher-resolution images. For each CH-1, CH-1∆A and CH-1∆B strain, over twenty individual vegetative cells were examined. Although some variations exist in the calculation of exact cellular size, a clear trend was observed that on an average, the calculated RMS values of length and width of CH-1, CH-1∆A and CH-1∆B were 1981 x 734 nm, 1518 x 555 nm and 1644 x 680 nm, respectively, confirming our previous observation that average cell length and width of both CH-1∆A and CH-1∆B was shorter than that of CH-1 cells under 1000x light microscopy. The cellular surface topography of both CH-1∆A and CH-1∆B changed when cultured in LB broth at 37°C (Figure 9A), and characterization of these strains were shown in Figure 9B. CH-1 showed a mean valey-to-peak distance (Rmean) of 11.900nm, with a maximal value (Rmax) of 25.900nM, a surface roughness (Ra) of 2.652nm, and a root-mean-square (Rq) 3.344nm. CH-1∆A and CH-1∆B showed an Rmean of 9.075 and 7.775nm, an Rmax of 17.700 and 14.675nm, an Ra of 1.999 and 1.545 nm, and an Rq of 2.240nm and 1.948nm, respectively. These data showed that Rssa and RssB are either directly or indirectly involved in determination of S. marcescens surface topology.
Evidence suggests that extracellular polysaccharide is reduced in rssA mutant As we used modified LB swarming plates which contained glycerol for selecting the super-swarming mutants, and glycerol was reported to stimulate extracellular polysaccharide (EPS) production in S. marcescens strains (Aucken et al., 1997), together with the fact that although not yet been identified in S. marcescens, EPS is shown to play an important role in stimulating swarming in many other bacterial species: (i) a colony migration factor for P. mirabilis swarming (Gygi et al., 1995); (ii) involved in E. coli swarming under the control of a two-component signaling system (Takeda et al., 2001), and (iii) important for V. parahaemolyticus swarming by a functionally uncharacterized three-gene-operon scrABC (Boles and MaCarter, 2002), we reasoned that production of EPS may be altered in CH-1∆A or CH-1∆B, leading to the super swarming phenotype. Measurement of EPS production (both the bound form and unbound form) (von Bodman et al., 1998) was performed under the bacterial growth conditions of CPG (Dolph et al., 1988) or LB broth, and seeding plate cultures. Unexpectedly but consistently from all culture conditions tested, while there was no significant difference in the amount of bound-form EPS produced among the 3 strains, we found that CH-1∆A produced less (about 40%) un-bound form EPS than CH-1 and CH-1∆B at 3 hr after inoculation at both temperatures, and the results from LB
seeding plates at 30°C were shown in Figure 10, suggesting that production of EPS is, instead of increased, reduced in CH-1∆A and CH-1∆B.
rssA-rssB are conserved among Serratia marcescens
To see whether rssA and rssB was conserved among S. marcescens strains, a total of 67 S. marcescens strains collected from National Taiwan University Hospital, one strain (S. marcescens NewCDC) isolated from Germany, and three strains (S. marcescens 1324E, S1220 and 4444) isolated from U.K. were subject to PCR
amplification using the primer pairs (5’CCATCATCGTCACCTTGCTGTTTACC3’/GAGCGACAGTTCCACATCCTTTT
CCA3’)and(5’TGCTGGATCTCACGCTGCCG3’/5’CCGGTTGACAGCCTTGACG C3’) designed from within the open reading frame region of rssA and rssB for amplifying rssA and rssB, respectively. We have detected both rssA and rssB DNA fragments from 68 out of the 71 S. marcescens strains tested (data not shown), suggesting that the rssA-rssB gene pair was conserved among S. marcescens strains.
Long chain SFAs regulate swarming of P. mirabilis and S. typhimurium
To see whether swarming of another two bacterial strains, P. mirabilis P19 and S. typhimurium LT2 was also inhibited by SFAs, SFAs and UFAs at the concentration of 0.01%(w/v) was incorporated into 2% Eiken agar LB plates and 0.5% Eiken agar LB plates followed by swarming assay of P. mirabilis and S. typhimurium, respectively, at 37°C. We found that a similar inhibitory effect of SFAs on swarming of both bacteria was observed and that UFAs showed either stimulate or inhibit the swarming of both bacteria (data not shown). The effect of myristic acid and myristoleic acid on swarming of both bacteria was shown in Figure 11. Both myristic acid and myristoleic acid significantly inhibited swarming of both bacteria. The results suggested a common regulatory effect of fatty acids on swarming bacterial species.
Discussion
Bacteria that differentiate and demonstrate multicellular behaviour as part of the regulated expression of gene networks required for the complex processes underlying morphological and physiological changes are commonly observed (Shimkets 1990; Matsuyama and Matsushita 1993; Shapiro 1995; Shapiro 1998 Rice et al., 1999; Alavi and Belas 2001). Regulation of these multicellular behaviours usually involves interaction between cells and/or cells to environment signals. Examples include Gram-positive bacterial antimicrobial peptide production (Kleerebezem and Quadri 2001), sporulation in Bacillus (Ryan and Shapiro 2003), light emission in Vibrio (Meighen 1999), biofilm formation in Pseudomonas (Costerton et al., 1999; Costerton 2001), production of nitrogen-fixing cells in cyanobacterium (Adams 2000), and also populational surface migration of many bacterial species (Velicer and Yu 2003; Horng et al., 2002; Romling 2001; Macfarlane et al., 2001; Fraser and Hughes 1999; Eberl et al., 1999; Harshey 1994; Shapiro 1998; McCarter and Silverman 1990).
Although a large body of information concerning the swarming mechanisms has been accumulated in Serratia and related bacterial species, the molecular mechanism(s) of swarming is far from understood. This is because the wide spectrum effects of physiology in swarming bacteria make it difficult to study the mechanisms directly. Furthermore, a potential specific and conserved regulator governing initiation of swarming has not been clearly identified, albeit a regulator of swarming behaviour of P. mirabilis, RsbA, has been reported by Belas et al., (1998). The identification of such a swarming regulator gene is thus an essential step in understanding the underlying mechanism of swarming. In this paper we present strong evidence that determination of S. marcescens swarming is controlled by acclimation of membrane fluidity, which is affected by environmental factors including temperature and nutrients, especially the provision of SFA substrates. We propose that the change of membrane fluidity is sensed by a pair of two-component regulatory proteins, RssA and RssB, which subsequently govern the expression of hemolysin gene shlA and fabGSm gene coding for the NADPH-dependent 3-ketoacyl-ACP reductase, an enzyme involved in fatty acid synthesis (Rawlings and Cronan 1992). In P. aeruginosa, FabG is also reported to be involved in rhamnolipid synthesis and 3-oxo-homoserine lactone acyl chain length determination (Campos-Garcia et al., 1998; Hoang et al., 2002). Although kinase-response regulator pairs of this type were frequently reported as governors of a wide variety of pathways in response to a myriad of environmental signals (Dutta et al., 1999; Hoch et al., 2000), we have for the first time shown the requirement for temperature- and SFAs- dependent regulation of swarming via these genes. The genetic studies shown in this work indicate that RssA-RssB may form a signal transduction pathway controlling the fatty acid
profile-dependent regulation of fabGSm gene expression and subsequently fatty acid profile, shlA hemolysin gene expression and initiation of swarming.
We hypothesized that when S. marcescens cells are growing into stationary phase at 30°C, no phosphorelay signaling is transferred between the RssA-RssB pair so RssB is mostly in a unphosphorylated form (inactive repressor) and at 37°C, the phosphorelay occurs and RssB is mostly phosphorylated (active repressor):(i) at 30°C, RssB in stationary phase cells is potentially in a unphosphorylated form in the presence or absence of RssA, expression of fabG Sm and shlA is not down-regulated, and bacteria swarm; when rssB is mutated, CH-1∆B thus showed a swarming, albeit “slow-swarming” phenotype. (ii) at 37°C, compared with CH-1 whose RssB is potentially phosphorylated to become an active repressor, fabGSm expression is inhibited and cells do not swarm, both CH-1∆A and CH-1∆B show a decrease in fabGSm expression and super-swarming phenotype, although CH-1∆A swarms faster and CH-1∆B slower. On the basis that no response regulator has yet been identified to be active in the unphosphorylated form (Hoch, 2000), we propose that RssB may act as an active repressor binding to the promoter of fabGSm when phosphorylated at 37°C, and binds to nearby promoter DNA region at 30°C (Lai et al., unpublished data). The role of RssA would then be to act as a phosphatase that selectively dephosphorylates RssB at 30°C. However, after a temperature upshift RssA would function as a specific kinase phosphorylating RssB, the cognate response regulator, which inhibits fabGSm expression and also swarming initiation on LB swarming plate. The results in this report strongly suggest that the sensor protein RssA is a bifunctional enzyme having both kinase and phosphatase activities. These two opposite activities of the sensor protein have been demonstrated in different two-component systems (for a review see Dutta et a., 1999). We have further shown that the transcriptional activity of the fabGSm promoter and subsequently the “swarming” or “non-swarming” fatty acid profile can be regulated by not only temperature shift, but also SFAs at a constant temperature. Temperature downshift or SFA deficiency may mediate activation of RssA phosphatase activity, leading to dissociation of unphosphorylated RssB, and subsequently RssB binding to another fabGSm promoter DNA region (Lai et al., unpublished data). Currently experiments are being performed to confirm these questions.
A provisional model accounting for our results is shown in Figure 12. We envisage that RssA could assume different signaling states under varying growth temperatures and nutrient conditions which lead to different membrane fatty acid composition for the sake of homeostasis. This could be accomplished by regulating the ratio of kinase to phosphatase activities, such that a kinase-dominant state is present at high growth temperature or SFA-rich nutrients. RssA possesses a single
transmembrane domain and either the periplasmic domain or cytoplasmic domain would function to propagate a conformational change that is sufficient to significantly alter its activity. This conformational change could be governed by the physical state of the membrane lipid bilayer. Lipids in biological membranes are usually maintained in the fluid, liquid-crystalline state (Vigh et al., 1998). The correct physical state of membrane lipids is required for optimal membrane structure and function. Temperature markedly affects membrane lipid composition, and changes in lipid composition are thought to occur in order to maintain an appropriate liquid crystalline state. The major way in which bacteria, generally lacking cholesterol, maintain this functional membrane physical state is by changing their fatty acid composition (Vigh et al., 1998). As the growth temperature decreases, the proportion of low-melting-point fatty acids in the membrane lipids increases. The phenomenon of membrane fluidity affected by environmental temperature is also shown in the cyanobacterium Synechocystis, where low-temperature signals are shown to induce the desaturation of fatty acids in the cell membrane, thus changing the membrane fluidity (Sakamoto and Murata 2002). Membranes at 30°C are normally in a less-liquid crystalline form and will undergo a transition to a more-fluid phase state when the temperature increases (Cronan and Rock 1996; Vigh et al., 1998). This change from a non-fluid (ordered) to a liquid state (less ordered) might cause activation of the kinase activity, resulting in autophosphorylation of a conserved histidine (His 188) contained in the transmitter domain of RssA. The phosphoryl group of His188 could be directly transferred to RssB, which down-regulates transcription of fabGSm or shlA. Activation of fabGSm results in a synthesis of “non-swarming fatty acid profile”. This metabolic pathway, therefore, generates a regulatory loop where SFAs or temperature upshift stimulates fabGSm transcription by favouring RssB phosphorylation and promotes phosphorylated RssB binding to its binding site. In B. subtilis, cells respond to a decrease in ambient growth temperature by desaturating the fatty acids of their membrane lipids and by increasing the proportion of anteiso-branched fatty acids (Aguilar et al., 2001). This pathway, termed the Des pathway, responds to a decrease in growth temperature by enhancing the expression of the des gene coding for an acyl-lipid desaturase (Aguilar et al., 2001). The Des pathway is also uniquely and stringently regulated by a two-component system composed of a membrane-associated kinase, DesK, and a soluble transcriptional activator, DesR. It is suggested that activation of this transduction pathway might be mediated by a decrease in membrane fluidity provoked by a temperature downshift (Aguilar et al., 2001). It is also found in B. subtilis that a decrease in the content of isoleucine-derived fatty acids in the membrane at a constant temperature could mimic a drop in growth temperature and
that both stimuli could induce UFAs synthesis (Cybulski et al., 2002). Although so far based on our experimental results there is no evidence the S. marcescens uses branch-chain amino acids for maintaining the membrane fluidity, changes of fatty acid profile in response to environmental temperature, nutrients or chemical signals is a common phenomenon. Therefore, this adaptive response could be important in providing an appropriate degree of membrane fluidity for bacterial survival under ever-changing physiological conditions. Here so far as we know, we have for the first time connecting the synthesis of fatty acids to be regulated by the two-component system and phenomenon of membrane fluid optimization to the regulation of swarming.
It is a common phenomenon that bacterial two-component systems are proved to be important for regulation of bacterial physiological behaviours under the regulation of physical or chemical environmental signals. Examples include the PhoP/PhoQ system of S. enterica governing transcription of some 25 loci in response to the extracellular concentration of Mg2+ (Soncini and Groisman 1996), the regulation of he arc system of E. coli by redox Quinone signals (Georgellis et al., 2001), the interaction between two autoinducers (AI-1 and AI-2) and two cognate membrane-bound two-component hybrid sensor kinases called LuxN and LuxQ respectively and regulation of light emission in Vibrio harveyi (Freeman et al., 2000). Furthermore, mostly regulation of swarming, although the signals that interact with bacterial two-component systems and thus the mechanism remain uncharacterized, regulation of P. mirabilis swarming by the putative RsbA sensor kinase (Belas et al., 1998), P. aeruginosa swarming, virulence factor expression and biofilm formation by GacA-GacS (Brinkman et al., 2001; Parkins et al., 2001), E. coli swarming and capsule synthesis by RcsC-YojN-RcsB (Takeda et al., 2001), and the S. enterica swarming by the putative two-component signaling components are also reported (Toguchi et al., 2000).
Temperature shift is a common environmental signal directing metabolic and phenotypic changes in bacteria. Pathogenic bacteria such as E. coli, Listeria, Shigella and Bordetella frequently express virulence characteristics preferentially at 37°C as part of the adaptation required to colonize their hosts (Mekalanos, 1992). Many bacteria use the two-component signal transduction system for adaptation of the environmental temperature shift. These include regulation of BvgA/BvgS by temperature, MgSO4 and nicotinic acid in Bordetella pertussis (Manetti et al., 1994)
and E. coli (Utsumi et al., 1994), regulation of des gene expression by DesK/DesR induced by temperature shift in B. subtilis (Aguilar et al., 2001), cold-regulated genes under control of the cold sensor kinase Hik33 in Synechocystis (Suzuki et al., 2001), the temperature-dependent biosynthesis of the P. syringae phytotoxin coronatine by