探討SARS冠狀病毒類木瓜蛋白酶對第一型干擾素訊號傳遞路徑之影響; Functional Characterization of SARS Coronavirus Papain-Like Protease in Type I Interferon Signaling Pathway
104
0
0
全文
(2) 致謝 誠摯的感謝指導教授 林振文老師,在碩士研究期間的指導與教 誨。也非常感謝 高銘欽老師、張清堯老師以及蕭乃文老師對此論文 的細心指導,指正並提共許多寶貴意見,使我獲益良多,得以完成這 本碩士論文。 同時感謝醫技所 鄭如茜老師、莊淨媛老師、楊翠青老師、黃蕙 君老師以及所裡各位師長在課業上的指導。 此外,也感謝實驗室 佳芳、明宏、盈如、冠傑、韋華及學弟妹 們在課業上與實驗上的協助,以及所有給我鼓勵和關懷的朋友們。 最後,謹將本論文獻給我親愛的父母、姊姊、哥哥以及男友,給 我許多鼓勵並支持我,讓我順利的完成碩士學業。.
(3) 中文摘要 嚴重急性呼吸道症候群冠狀病毒(SARS Coronavirus,SARS CoV)造 成致命性的非典型肺炎。SARS CoV 為正股單股 RNA 套膜病毒,可轉譯 成兩個大片段、非結構性的多聚蛋白,pp1a 與 pp1ab。類木瓜蛋白酵素 (PLpro)位在 nsp3 的位置中,具有辨識 LNGG 序列並水解之能力,以加工 水解多聚蛋白 pp1a 成為 nsp1、nsp2、nsp3。近來,類木瓜蛋白酵素被證 實具有對干擾素所引起的 15kDa 蛋白質-多汎素鏈 (ISG15-polyubiquitin chain)以及對 Ubiquitin-AMC 基質具有去汎素化 (de-ubiquitining)的酵素 活性。本研究先藉由基因工程技術表現 SARS PLpro 重組蛋白,再分析 SARS PLpro 之表現對於細胞干擾素反應,並藉由 2D-GE/MS 深入探討 SARS PLpro 對第一型干擾素路徑之訊息傳送之影響,以期了解 SARS PLpro 之致病機制,開發出有效之抗 SARS 病毒策略。在重組蛋白方面已 能表現出大量 SARS PLpro 蛋白,且在以具有 LNGG 辨識序列之 HRP 為 受質時,可切割 HRP 使之失去活性、呈色程度下降。再將此具活性之序 列轉殖成細胞表現載體以送入細胞表現後,在細胞內訊號傳遞途徑試驗 發現,SARS CoV PLpro 蛋白表現會抑制第一型干擾素所誘導的訊息途徑 表現,使 ISRE 訊息表現程度下降,但是無明顯抑制 LPS 所誘導的 NFκB 之訊息表現。以 RT-PCR 測定 ISRE 其下游基因 PKR 之表現,發現 SARS PLpro 表現細胞在經過第一型干擾素刺激後,使其 PKR 之表現較同樣經. I.
(4) 第一型干擾素刺激對照組細胞表現程度為低。再利用酵素連結免疫吸附 分析法,分析表現 SARS PLpro 細胞與對照組細胞經第一型干擾素 Interferon-β 刺激前後,SARS PLpro 表現細胞表現 Interferon-α 明顯偏低。 最後藉由二維電泳技術,分析表現 SARS PLpro 蛋白之細胞與對照組細 胞,經過第一型干擾素刺激前後之蛋白質體變化,再經由質譜儀分析技 術鑑定差異性蛋白質點。SARS PLpro 表現細胞經過 Interferon-α 處理後有 表現增加之蛋白質,如:Heat shock cognate71 kDa protein、Heat shock 27 kDa protein、Chloride intracellular channel protein 1、Ran-binding protein 1、 Peroxiredoxin-6 等,而表現減少的蛋白質,如:NADH dehydrogenase iron-sulfur protein 3 等。透過蛋白質點分析及架構出細胞內蛋白表現途徑 之改變,從中了解 SARS PLpro 機轉,以便將來開發 SARS 診斷及治療策 略。. II.
(5) 英文摘要 Severe acute respiratory syndrome coronavirus (SARS-CoV) causes a life-threatening atypical pneumonia. SARS CoV is a positive-sense single strand RNA enveloped virus, that encodes two large nonstructural polypeptides, pp1a and pp1ab. Papain-like protease (PLpro) that resides within nsp3 is involved in the proteolytic processing on the LNGG sites at the boundary regions between nsp1, nsp2, and nsp3. Recently, PLpro has been demonstrated to have the de-ubiquitining enzymatic ability on the ISG15-polyubiquitin chains and ubiquitin-AMC substrates.. In this study, we. intend to generate recombiment SARS PLpro protease and to analyse its cellular response. Then, 2D gel electrophoresis/mass spectrometry was used to differentiate proteomic profiling among mock cells and PLpro-expressing cells treated with type I interferon. In vivo signaling pathway assay indicated that type I interferon-induced signal transduction pathway was inhibited by SARS-CoV PLpro, while the LPS-induced NF-κB signaling was not significantly inhibited in PLpro-expressing cells. The real time RT-PCR assay indicated the expression of PKR gene was blocked by SARS PLpro in the interferon treated cells. In addition, the Enzyme-linked immunoassay revealed that expression of the interferon-α, in the interferon-β treated cell was inhibited by SARS PLpro. The 2D gel and MALDI-TOF/MS proteomic profiling were performed and the different expressed protein spots were identified.. Up-regulated. proteins. in. interferon-α. treated. SARS. PLpro-expressing cell were heat shock cognate71 kDa protein、heat shock 27 kDa protein、chloride intracellular channel protein 1、Ran-binding protein 1、 peroxiredoxin-6, while down-regulated protein was NADH dehydrogenase III.
(6) iron-sulfur protein 3. In this study, we provide the bio-informatics about the proteomics change and biochemical mechanism in the SARS PLpro expressing cells which will be helpful for developing diagnostic and therapeutic approaches to the SARS infection.. IV.
(7) 圖目錄 圖 4-1 構築 pET 表現系統以作為細胞外表現 PLpro 蛋白.....................61 圖 4-2 pET 系列表現 SARS PLpro 蛋白...................................................62 圖 4-3 pET 系列重組 SARS PLpro 蛋白之表現與純化............................63 圖 4-4 SARS PLpro 活性試驗(Horseradish Peroxidase, HRP ).................64 圖B.4-5 SARS PLpro 切割 HRP 活性試驗...................................................65 圖 4-6 構築 pcDNA3.1 His C 作為細胞表現 PLpro 蛋白表現系統.........66 圖 4-7 以 Anti His-tag 做免疫螢光染色之細胞內蛋白表現確認............67 圖 4-8 Anti HSV-tag 免疫螢光染色之細胞內蛋白表現確認....................68 圖 4-9 西方墨點法確認細胞內表現 PLpro...............................................69 圖 4-10 Single-reporter assay(Luciferase assay)測試 ISRE 訊息...............70 圖 4-11 Single-reporter assay(Luciferase assay)測試 NFκB 訊息..............71 圖 4-12 即時定量聚合酶連鎖反應偵測 Interferon α 誘發之 下游基因表現................................................................................72 圖 4-13 酵素連結免疫吸附分析法分析 interferon α 表現程度...............73 圖 4-14-1 二維電泳....................................................................................74 圖 4-14-2 二維電泳之差異蛋白質點……………………………………75. V.
(8) 表目錄 表 4-1 2D 膠體間相互比對之位置相似率..............................................76 表 4-2 以質譜儀分析出差異之蛋白質點...............................................77. VI.
(9) 目錄 中文摘要.................................................................................................I 英文摘要...............................................................................................III 圖目錄....................................................................................................V 表目錄...................................................................................................VI. 第一章. 緒論. 第一節. 研究背景與動機………………………………………..1. 第二節. 研究的重要性…………………………………………..3. 第三節. 研究目的………………………………………………..7. 第四節. 研究策略………………………………………………..8. 第二章. 文獻查證. 第一節 流行病學...........................................................................10 第二節 臨床症狀………………………………………………...11 第三節 病毒之遺傳物質與結構………………………………...13 第四節 類木瓜酵素 Papain-like protease……………………….15 第五節 Ubiquitining and de-ubiquitining…………….………….18.
(10) 第三章. 研究方法. 第一節. 研究材料........................................................................20. 第二節. 實驗方法. 標的一:SARS PLpro 重組蛋白之表現與其活性分析......................23 標的二:構築 SARS PLpro 表現細胞.............................................33 標的三:SARS PLpro 表現細胞經干擾素刺激前後之訊息分析........37 標的四:以二維電泳分析 SARS PLpro 表現細胞經干擾素刺 激前後之蛋白質表現......................................................44 標的五:以質譜儀技術分析二維電泳之蛋白質點,並探討其 影響之訊息及蛋白網絡架構...........................................47. 第四章. 研究結果. 標的一:SARS PLpro 重組蛋白之表現與其活性分析....................48 標的二:構築 SARS PLpro 表現細胞...........................................50 標的三:SARS PLpro 表現細胞經干擾素刺激前後之訊息分析......52 標的四:以二維電泳分析 SARS PLpro 表現細胞經干擾素刺 激前後之蛋白質表現....................................................55 標的五:以質譜儀技術分析二維電泳之蛋白質點,並探討其 影響之訊息及蛋白網絡架構..........................................56. 第五章. 討論...............................................................................57. 參考文獻............................................................................................78 附錄....................................................................................................91.
(11) 第一章 緒論 第一節. 研究背景與研究動機. 嚴重急性呼吸道症候群(Severe acute respiratory syndrome, SARS) 在2002年11月中國廣東省出現後,引發全球SARS風暴,傳到大約30個國 家,統計自2002年11月起至2003年7月為止,共計8096件通報世界衛生組 織之確定病例並造成超過900個死亡個案 (World Health Organization, WHO),而臺灣也包含其中。SARS其傳染途徑是藉由飛沫或體液傳染, 故若與病患有較密切接觸時,會透過吸入或黏膜接觸到病患飛沫或體液 而受到感染。感染後會造成嚴重之呼吸道症狀,如乾咳、發燒、呼吸困 難等,接著免疫能力下降、瀰漫性肺炎、肺部纖維化等等,最後造成死 亡(Lee et al., 2003; Tsang et al., 2003; Hsueh et al., 2004)。而SARS至今仍未 找到或開發出有效之疫苗及特效藥,目前其主要治療方法所造成之許多 副作用與感染後之後遺症卻仍然影響著癒後生活。 目前從文獻中得知,SARS PLpro之蛋白水解能力能將病毒蛋白處理 成為具活性之狀態,使病毒能成功複製、增生(Thiel, V. et al., 2003; Harcourt, B. H. et al., 2004),另外,SARS PLpro具有deubiquitining之活性 (Lindner et al., 2005),且能水解ubiquitin調節蛋白(ubiquitin-like modifier)干 擾 素 (IFN) 引 起 的 15-kDa 蛋 白 質 ( interferon-induced 15 kDa protein, ISG15) (Barretto et al., 2005),因此被推論出可能調控了宿主細胞之. 1.
(12) ubiquitination機制。故本研究針對SARS對患者造成之嚴重免疫傷害與 SARS PLpro調控宿主細胞之Interferon訊號傳遞路徑及ubiquitination之相 關聯性做研究。. 2.
(13) 第二節. 研究的重要性 Ubiquitin-proteasome是將細胞內多餘蛋白質降解之主要途徑,. 在細胞內扮演許多重要功能,包括:抗原處理、細胞週期調節、細胞 凋 亡 、 訊 號 傳 遞 、 轉 錄 調 節 、 以 及 DNA 修 復 等 (Glickman and Ciechanover 2002)。細胞內蛋白質降解主要藉由兩個連續性的途徑, (1) 泛素化作用 (Ubiquitination):泛素共價加掛到目標蛋白上。 (2)proteasome-mediated degradation:被泛素加掛的蛋白會被26S蛋白 酶體分解,並釋放泛素。 Ubiquitin之主要功能是一個蛋白質降解的訊號。在Ubiquitination 過程中有三個酵素反應參與 (reviewed by Gao and Luo, 2006)。首先 Ubiquitin會透過E1(ubiquitin-activating enzyme)的協助及ATP的參與而 活化,其C端之glycine殘基會構成thiol ester bond,之後活化的Ubiquitin 會經由thiol ester linkage轉移到E2(ubiquitin-conjugating enzyme)上。最 後由E3(ubiquitin-protein ligase)在Ubiquitin與目標蛋白的lysine殘基之 間構成共價的isopeptide bond,將Ubiquitin加掛到目標蛋白上。當 Ubiquitination循環數次後,會有多個Ubiquitin分子被加掛到目標蛋白 上,每個polyubiquitin鏈至少含有四個Ubiquitin分子。被標定的目標 蛋白會在短時間內被辨識且由26S蛋白酶體降解。 干擾素(interferon, INF) 是一種多功能的細胞素,具有抗病毒感. 3.
(14) 染 (anti-viral infection) 、 抗 增 殖 (anti-proliferative) 及 免 疫 調 節 (immuno-modulatory) 等功能 (Samuel, 2001; Levy and Garcia-sastre, 2001; Grander et al., 1997; Biron, 2001)。干擾素依其基因序列比對分析 可分為第一型及第二型干擾素。第一型干擾素包括二十種以上的 INFα、只一種的INF-β、六種的INF-ω、及INF-τ,而第二型干擾素只 有一種INF-γ(Allen and Diaz, 1994)。 病毒感染早期藉由非特異性免疫,包括干擾素、自然殺手細胞 (Natural killer cell,NK cell)來消滅病毒。當細胞被病毒感染後,會 誘發產生IFN-α、IFN-β,促使附近細胞抗病毒的機轉活化來抵抗病毒 的擴散。當病毒感染進入後期後,特異性免疫作用則被活化,IFN-γ 可以刺激MHC-I、MHC-II分子的表現增加來提高特異性免疫反應的 能力,並活化NK細胞的功用。 在訊息傳遞路徑方面,第一型干擾素受器可與非受器蛋白激脢 (protein kinase) JAK1(janus kinase 1)及TYK2 (tyrosine kinase 2)結合, 而 第 二 型 干 擾 素 受 器 則 與 JAK1 及 JAK2 結 合 (O’shea et al., 2002; Aaronson and Horvath, 2002),並吸引細胞質內之轉錄因子STAT(signal transducer and activatorof transcription)結合,將其酪胺酸殘基磷酸化 後,再與STAT形成雙體聚合的形式由細胞質轉移至細胞核,然後與 特異的啟動子區域結合,活化干擾素刺激基因群(interferon-stimulated. 4.
(15) genes, ISGs),傳達干擾素的生物效應 (Heim,1999)。 而干擾素刺激基因群的啟動子區域有兩段保留的核酸序列。第 一段保留的核酸序列是ISRE (interferon-stimulated response element), 由12到15個核酸(AGTTTCNNTTTCNC/T)所組成。第二段保留的核酸 序 列 為 GAS (gamma-interferon activation site) , 由 9 個 核 酸 (TTNCNNNAA)所組成(Yeh and Pellegrini, 1999; Meraz et al., 1996)。 干 擾 素 可 藉 由 活 化 ( 如 RNA-specific adenosine deaminase 、 2’, 5’ oligoadenylate. synthetase. [OAS] 、. dsRNA dependent. protein. kinase[PKR]、C56 protein、midolamine dioxygenase、人類白血球抗原 及腫瘤壞死因子受器),與抑制生長因子 (如basic fibroblast growth factor、platelet derived growth factor, IL-1 及腫瘤壞死因子) 之表現, 達到抗病毒、抗增殖及免疫調節等作用(Samuel, 2001; Biron and Sen, 2001; de Veer et al., 2001; Der et al., 1998)。 干擾素刺激基因群中,由 IFN-α/β 所誘導出來的 ISG15 蛋白,為一個 類泛素蛋白(ubiquitin-like protein) (Schwartz et al., 2003)。經第一型干擾素 處理後, ISG15 會藉由與細胞蛋白質共價結合,類似 ubiquitin 修飾 (ubiquitination 或 ubiquitylation)之過程 ,使細胞蛋白質被分解(Pickart, 2001)。ISG15 其接合酶 E1- (Ube1L)及 E2- (UbcH8)能促使 ISG15 與細胞 內蛋白質之接合作用(conjugation) (Kim et al., 2004)。. 5.
(16) 先前已有許多病毒被發現演化出不同策略,藉由 ubiquitin-proteasome 途徑,逃避先天性免疫中 IFN-α/β 之抗病毒作用。如:人類 B 型流感病 毒所產生之 NS1B 蛋白會抑制 ISG15 與目標蛋白之接合作用,使病毒逃 避細胞之免疫反應。由此可知,在 IFN-α/β 所誘導之抗病毒反應中,ISG15 接合作用為一關鍵抗病毒機制(Yuan and Krug, 2001)。Mumps virus 與 simian virus V protein 會活化 ubiquitin-proteasome 途徑,分解細胞因子 STAT protein,以抑制 JAK/STAT signaling,達到逃避 interferon 所引發 之 抗 病 毒 反 應 (Gotoh et al. 2002)。 而 SARS PLpro 已 被 推 論 具 有 De-ubiquitining 之活性,且能水解 ISG15,故可推測 SARS PLpro 對病毒 之抑制細胞抗病毒有其相關性。. 6.
(17) 第三節. 研究目的. SARS PLpro 蛋白在 SARS 病毒之複製與重組扮演著重要角色,且已 有文獻指出 SARS PLpro 與細胞免疫之傷害有相關聯性,故本研究欲藉由 基因工程技術表現 SARS PLpro 重組蛋白,分析 SARS PLpro 表現對細胞 之干擾素反應,深入探討 SARS PLpro 與 Ubiquitin 路徑之訊息傳送,以 期了解 SARS PLpro 之致病機制,開發出有效之抗 SARS 病毒策略。. 7.
(18) 第四節. 研究策略. 標的一:SARS PLpro 重組蛋白之表現與其活性分析 構築表現質體並以大腸桿菌表現出 SARS PLpro 蛋白,且以對 HRP 切割確定其是否具有活性。. 標的二:構築 SARS PLpro 表現細胞 構築細胞內表現質體並送入 HL-CZ 細胞中表現,且以 Intracellular fluorescence stain 與 Western blot 確定 SARS PLpro 蛋白於細胞內之表現。. 標的三:SARS PLpro 表現細胞經干擾素刺激前後之訊息分析 由 cis-actin plasmid reporter gene、real time RT-PCR 與 ELISA 分析並 探討 SARS PLpro 表現細胞對 Type I Interferon 訊號傳遞路徑之影響。. 標的四:以二維電泳分析 SARS PLpro 表現細胞經干擾素刺激前後之蛋 白質表現 以二維電泳技術分析 SARS PLpro 之表現與否對細胞內蛋白表現之 影響及改變,並分析 SARS PLpro 表現細胞對 Type I Interferon 訊號傳遞 路徑之影響。. 8.
(19) 標的五:以質譜儀技術分析二維電泳之蛋白質點,並探討其影響之訊息 及蛋白網絡架構 以質譜儀分析二維電泳膠體上差異之蛋白質點為何,再以西方墨點 法檢測其細胞內表現量之差異是否相同。. 9.
(20) 第二章 文獻查證 第一節. 流行病學. 全 球 首 例 確 定 之 嚴 重 急 性 呼 吸 道 症 候 群(Severe acute respiratory syndrome, SARS)病例在 2002 年 11 月由大陸廣東省發現後,臺灣在 2003 年 3 月發現第一起由香港境外移入之 SARS 病例,並在 4 月份向 WHO 通報為確定病例後,確定成為 SARS 感染地之一。 東南亞國家,如:大陸、香港、越南、馬來西亞、菲律賓、新加坡、 泰國等地,以及北韓、加拿大、美國等世界各地都出現感染 SARS 之確 定病患,其中以東南亞國家最為嚴重(World Health Organization, WHO.)。 直至2003年11月底臺灣共計出現346件確定病例,135件不確定病 例,造成138人死亡(其中37人為確定病例,101人為不確定病例),而大陸 出現349人、香港299人、新加坡33人、加拿大43人死亡,不到一年的時 間內總計造成全球約900多人死亡(Poutanen et al., 2003; Peiris et al., 2003; Drosten et al., 2003, WHO)。 因SARS其傳染途徑是藉由飛沫或體液傳染,故與病患有較密切接觸 時,會透過吸入或黏膜接觸到病患飛沫或體液而受到感染。且經許多病 例研究發現,SARS病毒在離開人體狀態下仍能在痰液與糞便中存活達30 多天,故在爆發期間出現許多連續性的感染,因而造成恐慌(WHO. Wang et al., 2004)。. 10.
(21) 第二節 臨床症狀 嚴 重 急 性 呼 吸 道 症 候 群 (SARS) 在 臨 床 症 狀 上 發 現 有 發 燒 ( 高 於 38℃)、寒顫、肌肉酸痛、頭痛、咳嗽(大多數是乾咳)等症狀,嚴重者 會出現瀰漫性肺炎與呼吸衰竭等症狀。經由肺病理學檢查顯示,發現 SARS 會 造 成 次 支 氣 管 上 皮 細 胞 剝 落 、 纖 毛 損 傷 、 多 核 融 合 細 胞 (multinucleated syncytial cell)、鱗狀細胞再生不良、肺泡及肺泡間質出現 纖維細胞增生與纖維化等肺組織病變產生(Nicholls et al., 2003; Lang et al., 2003)。 血 液 檢 查 則 會 有 白 血 球 正 常 或 下 降 (leukopenia) 、 淋 巴 球 數 低 下 (lymphopenia)、血小板低下(thrombocytopenia)、肝功能異常等現象,以及 C-reactive. protein(CRP) 、 creatine. phophokinase(CPK) 、 lactate. dehydrogenase(LDH) 、IL-1 (白細胞介素-1,Interleukin-1)、IL-6、IL-8、 IL-12 與干擾素(IFN)-γ 上升等情況產生(Wang et al., 2004; Huang et al., 2005)。另外,經由驗證證實 SARS 病患之 TH1 細胞會發生活化與發炎等 反應(Wong et al., 2004; Huang et al., 2005) 。. 目前 SARS 之主要治療方法為合併使用 Ribavirin 與類固醇,但在治 療後會造成許多副作用,如 Ribavirin 會造成紅血球破裂、貧血、致胎兒 死亡等,類固醇則會造成胃潰瘍、水腫、心臟負荷、骨質疏鬆等副作用,. 11.
(22) 並且治癒後仍可能會有 SARS 造成之肺部纖維化等症狀造成肺功能衰弱 等後遺症(WHO)。. 12.
(23) 第三節 病毒之遺傳物質與結構 SARS 由 SARS coronavirus(SARS CoV)感染所造成,而 SARS CoV 為一正股單股 RNA 套膜病毒,由三種膜蛋白,如刺突蛋白(Spike)、套膜 蛋白(Envelope)、膜蛋白(Membrane)組成套膜圍繞著病毒核(nucleocapsid core) (Siddell, 1995; Lai and Holmes, 2001)。 SARS CoV 其病毒基因約 29.7kb,包含有 14 個可能的開放閱讀框架 (open reading frames, ORFs) (Marra et al., 2003; Thiel et al., 2003)(圖 2-1), 其結構蛋白由基因之 5’端 Cap 至 3’端 poly A 可分別轉譯成:複製酶 (Replicase)-刺突蛋白(Spike)-套膜蛋白(Enevlope)-膜蛋白(Membrane)-核殼 蛋白(Nucleocapsid)。其基因能轉譯成兩個大片段、非結構性的多聚蛋白 (polyprotein),分別是 pp1a 與 pp1ab。在 pp1a 上帶有兩種蛋白酵素 3C-like protease(3CLpro)及 papain-like protease(PLpro)用以切割處理多聚蛋白 pp1a 與 pp1ab,產生多種參與病毒基因組複製及轉錄酵素,如 Helicase、 RNA polymerase、endoRNAase 等等。. 13.
(24) (Adapted from Tan et al., 2005). 圖 2-1 ORF1a 與 1b 占了整個 SARS 基因(29.7kb)前段 2/3,並能轉譯出 兩個多聚蛋白 pp1a(486kDa)與 pp1ab(790kDa),由病毒之半胱氨酸蛋白酶 (nsp3:PLpro,ns5:3CLpro 或 Mpro)將這兩個多聚蛋白修飾水解成為 16 個非結構性蛋白。. 14.
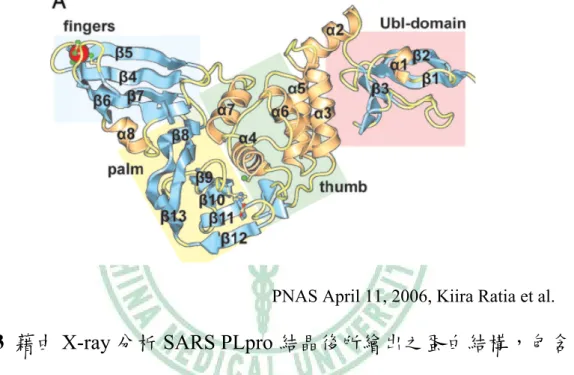
(25) 第四節 類木瓜酵素 Papain-like protease 類木瓜酵素(Papain-like protease,PLpro)為一種蛋白酶,位在 SARS 轉譯出的多聚蛋白 pp1a 上(圖 2-2),會特異性的辨識 LNGG 蛋白序列並 切割於 LNGG 的後方(Thiel et al., 2003; Lindner et al., 2005; Barretto et al., 2005),可以將 SARS 病毒之多聚蛋白 pp1a 切割成 NSP1、NSP2、NSP3(Kiira R et al.,2006) 。 不 同 於 其 他 的 冠 狀 病 毒 可 轉 譯 出 兩 個 papain-like protease(PLP1、PLP2),SARS CoV 只帶有一個 papain-like protease(Kiira R et al.,2006)。SARS PLpro 分子量大約 50kDa,在 pH 中性時具有較佳的活 性,可辨識序列 LNGG 並切割,但在酸性及鹼性溶液中活性則大大降低 (Yu-San Han et al.,2005)。. Kiira Ratia et al., PNAS April 11, 2006. Holger A. et al., JOURNAL OF VIROLOGY, Dec. 2005. 圖 2-2 PLpro 之基因序列位在 ORF1a 上,為 non-structure protein 3(nsp3)。. 15.
(26) 結構方面,在2006年首先由Kiira Ratia等人,成功的得到大小約1.85-Å 的SARS Papain-like protease結晶,並藉由X-ray繞射分析結晶,發現SARS PLpro 包含三個部份的結構,分別是zinc-binding domain、N-terminal ubiquitin-like domain , 以 及 一 個 整 體 相 似 於 USP14 及 HAUSP 的 已 知 deubiquitining之結構(DUB)( Kiira Ratia et al., 2006)(圖2-3)。. PNAS April 11, 2006, Kiira Ratia et al.. 圖 2-3 藉由 X-ray 分析 SARS PLpro 結晶後所繪出之蛋白結構,包含 zinc-binding domain、N-terminal ubiquitin-like domain 及整體相 似於 USP14 及 HAUSP 之已知 deubiquitining 結構。. 在生化特性方面,SARS PLpro 已於先前研究中發現,利用 N-terminal 及 C-terminal deleation 試驗確認 SARS CoV PLpro 完整活性序列在 pp1a/pp1ab 第 1541 至 1858 氨基酸的 35 kDa 蛋白(Lindner et al., 2005;. 16.
(27) Barretto et al., 2005) 。進而利用定點突變分析發現 SARS CoV 蛋白酶 PLpro. 活 性 需 要 含 有 鋅 結 合 區 (zinc-binding. site) 及 如. Cys1651-His1812-Asp1826 之蛋白酶活性催化作用區域(Barretto et al., 2005)。. 17.
(28) 第五節 Ubiquitining and de-ubiquitining 由於 SARS CoV 蛋白酶 PLpro 之高度保留性的切割序列為 LNGG (Thiel et al., 2003; Lindner et al., 2005; Barretto et al., 2005) , 與 deubiquitinating 辨認之序列相同,因此 SARS CoV 蛋白酶 PLpro 被認為 具有 deubiquitinating 活性。藉由水解 deubiquitinating 酵素受質能力 (deubiquitinating enzyme substrate),如 ubiquitin-7-amino-4-methylcoumarin (Ub-AMC)及 Z-LRGG-AMC,或同時加入 deubiquitinating enzyme inhibitor 之水解能力分析,驗證發現 SARS CoV 蛋白酶 PLpro 具有 deubiquitinating 活性(Lindner et al., 2005)。 另外,透過其細胞內共同表現試驗發現 SARS CoV 蛋白酶 PLpro 可 水 解 ubiquitin 調 節 蛋 白 (ubiquitin-like modifier)- 干 擾 素 (IFN) 引 起 的 15-kDa 蛋白質( interferon-induced 15 kDa protein, ISG15) (Barretto et al., 2005),因此,被推論調控宿主細胞的 ubiquitination 機制,逃避宿主細胞 之免疫反應,以助於 SARS CoV 病毒複製。. 綜合以上文獻發現,SARS 感染後會造成嚴重性的免疫傷害,使得 SARS 患者之白血球可能會有下降(leukopenia)之現象,淋巴球數低下 (lymphopenia)、血漿中細胞素(cytokine),如在干擾素(IFN)-γ,IL-1,IL-6 和 IL-12 會明顯升高等等之症狀,並產生肺部之不可逆傷害。而其中 SARS. 18.
(29) PLpro 證實具有 deubiquitining 之活性,被推論出可能調控了宿主細胞之 ubiquitination 機制,且能水解 Interferon 所引起之蛋白之特性,被認為能 藉以逃避宿主細胞之免疫反應,達到病毒複製之目的等,皆與細胞之免 疫反應有相關。 而在其他文獻中,流感病毒 NS1B 蛋白質被發現能與 ISG15 結合, 並抑制 ISG15 的 ubiquitinating 活動,也抑制了 Interferon 所引起的先天性 免疫反應(Yuan et al., 2001)。African swine fever virus 蛋白酶同樣也具有 deubiquitinating 活性,能明顯地使受感染之細胞無法產成 Interferon α/β(La Rocca et al., 2005)。另外,有文獻指出被 SARS CoV 感染之吞噬細胞 (macrophage) 可 釋 放 出 CXCL10/IFN-gamma-inducible protein 10 及 CCL2/monocyte chemotactic protein 1 , 但 卻 無 法 誘 發 產 生 Interferon β(Cheung et al., 2005)。 故本研究將探討 SARS PLpro 對第一型干擾素訊號傳遞路徑之影 響,藉由分析 SARS PLpro 對細胞之訊息傳遞改變與蛋白生成表現之影 響,從中了解其對細胞之致病機制,以期能找到良好之治療方針。. 19.
(30) 第三章 研究方法 第一節. 研究材料. 1. 全長SARS CoV基因組 cDNA – 感謝台灣大學醫學院陳培哲教授提供 予本實驗室。. 2. HL-CZ : 人 類 單 核 球 白 血 病 細 胞 ( human promonocytic leukemia cells) ,培養於 90%的 RPMI-1640,含有 5 % FBS (fetal bovine serum)。. 3. 大腸桿菌菌株(Escherichia coli) Top 10:基因行為 F-,mcrA ,△(mrr-hsdRMS-mcrBC),φ80lacZ,△M1, △lacX74,recA1,araD139,galK,△(ara-leu)7697,rpsL (StrR) ,endA1, nunpG,用於質體轉形(transformation)、質體備製。 BL21(DE3):基因型為 F-,ompT,hsdSa(rB- mB-),ga1,cdm(DE3)(camR), 用於蛋白表現(protein induction)。. 4. 質體(Vector) pET24a(Novagen):含有 5310 個鹼基配對,可在大腸桿菌 BL21 品係,經 IPTG 誘導後表現蛋白質,N’端帶有 T7-tag。 pcDNA3.1/His C(Invitrogen):含有 5513 個鹼基配對,可在哺乳類動物細 胞(mammalian cell)中,經由 human cytomegalovirus(CMV) immediate-early 20.
(31) promotor 表現蛋白,並在 N’端帶有 Anti-Xpress epitope tag、His-tag。 pEGFP-N1(BD Biosciences Clontech):含有 4733 個鹼基配對,可以在哺 乳 動 物 細 胞 (mammalian cell) 中 , 經 由 human cytomegalovirus(CMV) immediate-early promotor 表現 Enhanced green fluorescent protein(EGFP)蛋 白,在顯微鏡下產生螢光。. 5. Arrest-In transfection reagent:Open BIOSYSTEM.. 6. 單株抗體: a. Mouse anti Histidine tag :Purified IgG, Serotac. b. Mouse anti HSV tag:Purified IgG1, Novagen. c. Mouse anti β-actin:Purified IgG1, SIGMA. d. Goat anti mouse conjugate -AP :Purified IgG, ZYMED. e. Goat anti mouse conjugate-Rhodamin:Purified IgG, CHEMICON. f. Mouse anti Interferon-α antibody: abcam.. 7. Ubiquitin-AMC :Human recombinant, Boston Biochem.. 8. Dual-Luciferase Reporter Assay System:Promega.. 21.
(32) 9. Micro-to-Midi Total RNA Purification System:Invitrogen.. 10. PlusOne TM Silver Staining Kit :Amersham Biosciences.. 11. 引子(primer): SARS PLpro 4507-F(Kpn I): 5’-gC AAg gTA CCC AAC TCT CTA AAT gAg CCg-3’ SARS PLpro 5095-R: 5’-ACT CgA AAg CTT CAC TAC gTA gTg-3’ SARS PLpro-4951-F(Bam HI): 5’ -gTg gAT CCA TgT CTA TgA CAT ATg gAC AgC AgT TTg gTC CAA-3’ SARS PLpro-5840-R(Xho I): 5’- TgC TCg AgT ACg ACA CAg gCT TgA Tgg TTg - 3’. SARS PLpro-F(Eco RI): 5′- CTC CGA ATT CAA CTC TCT AAA TGA GCC GCT TGT C -3′ SARS PLpro-HSV-R: 5′- GAG GCT CGA GAT CCT CTG GGT CTT CAG GAG CGA GTT CTG GCT GTA CGA CAC AGG CTT GAT GGT TGT AGT G - 3′. 22.
(33) 第二節. 實驗方法. 標的一:SARS PLpro 重組蛋白之表現與其活性分析. Flowchart I:. Cloning:SARS-PLpro/pET 24a. Transformate to BL-21. Protein induction and purification. PLP activity. 23.
(34) 1. SARS CoV PLpro 基因選殖. 4507-509. 4951-584 Hind. S4:3772-5125. T7 tag PLpr. S5:4982-6358. His tag. pET-24a(+ SARS PLpro:1332bp. 欲構築之 SARS PLpro 核酸序列為 4507-5840,故先將 cDNA Fragment 5 (核酸 4982-6358)用引子 SARS PLpro-4951-F 以及 SARS PLpro-5840-R 以聚合酵素連鎖反應夾出核酸序列 4951-5840,再構築核酸 4951-5840 於 pET24a 中。 接著再將核酸序列 4507-5095 由 cDNA Fragment 4 (核酸 3772-5125) 中用引子 SARS PLpro 4507-F 以及 SARS PLpro 5095-R 以聚合酵素連鎖 反應夾出,並利用核酸序列 5090-5095 為 Hind III 切位,將 4951-5840-pET24a 及 4507-5095 片段以 Hind III 水解切割,再重新接合至 pET24a 中。. 24.
(35) 將此構築好的表現載體利用 Heat-shock 的方式轉形(transform)到 DH5α 大腸桿菌中並做篩選。經限制酶水解釋驗確定切割載體之片段大小 正確後,將載體送定序。. 2. 聚合酵素連鎖反應(polymerase chain reaction,PCR) 在 0.5ml 微量離心管中,取 cDNA 2ul 作為模版(template),依序加入 ddH2O 38.5ul,10x pfx amplication buffer 5ul, dNTP 1ul, MgSO4 1ul, primer F 與 primer R(10mM)各 1ul,最後加入 pfx 0.5ul,總體積 50ul。混 合均勻後加入約 20ul 礦物油再置於聚合酵素連鎖反應器(Thermohybrid) 的反應槽中。先以 94℃作用十分鐘,再以每個週期,94℃作用一分鐘的 DNA 變性反應,58℃作用一分鐘的 DNA 鏈合反應,72℃作用一分鐘二 十秒的 DNA 合成反應。進行 40 個反應週期,再用 68℃作用十五分鐘將 DNA 片段補齊。最後設定 4℃保存。將 PCR DNA 產物跑 2% 瓊酯凝膠 (1% agarose gel)進行電泳分析。確定 PCR 產物片段大小正確,以 Spin PCR Clean-Up system 試劑套組(Viogene)將 DNA 純化。. 3. 質體純化(plasmid purification) 含有質體的單一菌落,以 3ml LB 在 37℃培養箱搖隔夜後取用出,以 10000 G 離心五分鐘後,倒掉上清液,用 Gene-SpinTM Miniprep Purification 25.
(36) Kit 試劑組(Protech technology)來純化質體。 以 Solution I 溶液 200ul resuspend pellet,加入 Solution II 溶液 200ul 上 下倒置數次後,再加入 Solution III 溶液 200Uul 上下倒置數次,以 12,000rpm 離心 10 分鐘。將 Spin column 插入ㄧ收集管(collection tube)中, 離心後的上清液移至 spin column 中,離心一分鐘,移除下層濾液後,加 入 700ul Washing solution,離心一分鐘,再移除下層濾液,離心一分鐘。 再將收集管以 1.5ml 微量離心管置換,加入 20ul ddH2O 回收質體。. 4. 限制酵素水解試驗 PCR 得到的產物,以 Spin PCR Clean-Up system 試劑套組(Viogene) 將 DNA 純化後,以 40ul ddH2O 將 DNA 溶出,加入 10X Reaction buffer(Fermentas) 5ul 及切位的限制酵素 KpnI 與 XhoI 各 2.5ul,使總體積 為 50ul,以 37℃作用 3-5 小時。相對應的質體載體也以同樣的方式進行 限制酵素的水解,來構築實驗所需的表現載體。 將作用 3-5 小時的 DNA 產物跑 1% agarose gel,染 EtBr 十分鐘,在 UV 燈下將正確位置的 DNA 切下,以 Gel Extraction System 試劑套組 (Viogene)將 DNA 純化。. 26.
(37) 5. 質體構築:DNA 接合(DNA ligation) 質體載體 1ul,加上欲接合的 DNA 7ul,10x ligation buffer 1ul,T4 DNA ligase (Fermentas) 1ul,以 16℃作用至隔夜。. 6. 適應性細胞(competent cell)製備: 培養隔夜的 E.coli cell(Top 10、BL21) 2ml,倒入 200ml LB 中,以 37℃ 培養箱震盪培養 2-3 小時達到 Log-phase,取出後置於冰上 10 分鐘,再以 3000rpm、4℃離心 15 分鐘,倒掉上清液,以 0.1M CaCl2 resuspending pellet。於冰上靜置 30 分鐘後,再以 700 G、4℃離心 15 分鐘,小心倒掉 上清液,再以 0.1M CaCl2 1.6ml,glycerol 0.4ml (20%)將 pellet resuspend, 然後靜置於冰上 2 小時後,分裝置 1.5ml 離心管中,每管約 200ul 左右。. 7. 大腸桿菌之轉形(transformation) 將構築好的質體加入適應性細胞中,混合後置於冰上 30 分鐘。再經 42℃反應 90 秒後,迅速置於冰上 3 分鐘。再加入 800ul 不含抗生素之 LB 培養液,在 37℃培養箱培養 1 小時後,離心去除上清液,將 pellet 均 勻塗抹在含抗生素的 LB 培養基上。. 27.
(38) 8. 重組 SARS PLpro 蛋白之表現與純化 將已選殖到的 SARS PLpro 基因架接到 pET-24a(+)上的 KpnI 及 XhoI 限制酶切位上。再將此構築好的表現載體利用 Heat-shock 的方式轉型 (transform) 到 BL21(DE3)pLysS 大 腸 桿 菌 並 做 篩 選 。 將 篩 選 到 含 有 pET-PLpro 載體的大腸桿菌,培養在 10ml 含 Ampicillin 之 Luria-Bertani broth(LB broth)中經 37℃,250rpm 震盪培養隔夜後,以 1:50 的比例接 種到 500ml 含 Ampicillin 之 LB broth 中,至於 37℃,250rpm 震盪 2 小時 後 , 加 入 IPTG(isopropyl-beta-D-thiogalactopyranoside) , 使 最 終 達 到 0.1mM,以誘導 T7 RNA 聚合酶表現,促使 pET 載體上基因表現。再經 過 16℃,250rpm 震盪培養 5 小時後,將菌液於 4℃、8000rpm 離心 5 分 鐘,以收集細菌。之後加入 20ml 的 10mM imidazole,再以超音波擊碎機 (sonicator)將細胞打碎。 利 用 LC 純 化 蛋 白 , 在 column 中 裝 填 10ml 的 Chelating SepharoseTM(Amersham Biosciences),將已打碎的細菌溶液離心,上清液 流過鎳離子親和性層析凝膠,先用沖洗緩衝液(100 mM imidazole)沖洗, 再用析出緩衝液(400mM imidazole)將蛋白質析出。再以 12% SDS-PAGE 檢測蛋白質純度。. 28.
(39) 9. 蛋白質濃度測定 純化後的蛋白質濃度用 Bio-Rad protein assay 染劑(取 1ml,以二次水 稀 釋 到 5ml) 進 行 測 試 。 用 牛 血 清 白 蛋 白 (bovine serum albumin, BSA)1mg/ml 為標準樣品,分別配成濃度為 0、0.2、0.4、0.6、0.8、1.0mg/ml 各 20ul,各別加入 1ml 染劑中,算出線性迴歸公式,作出標準曲線。再 取蛋白質樣本 20ul 加入 1ml 染劑,測 OD595nm 的吸光值,並分析蛋白質 濃度。. 10. 蛋白質之電泳分析(SDS-PAGE): 實驗所使用的電泳裝置為 Bio-Rad 蛋白質電泳槽。待鑄膠裝置組合 後,先加入分離膠體溶液(Separating gel),再加入二次水將膠體水平壓 平。待凝固後倒掉二次水並擦拭乾後,加入集膠溶液(Stacking gel),插上 齒模,膠體凝固後放置於電泳槽中,注入電泳緩衝液(Running buffer)。 將蛋白質樣本與 2x Sample loading buffer 以 1:1 比例混合均勻,以 100℃加熱五分鐘後,置於冰上冷卻。待冷卻後取 15ul 注入樣本槽,並注 入蛋白值標準溶液以對照蛋白質分子大小。先以 80V 進行電泳,待藍色 染劑跑到分離膠體時,調整電壓至 110V 繼續電泳,直到藍色染劑跑到膠 體底部,關閉電源取出膠體,以 Commassie brilliant blue 染色五至十分鐘 後,再用脫色液(Destain solution)脫色至蛋白質色帶清楚。 29.
(40) 11. 西方墨點法(Western blotting): 實驗所使用的電泳裝置為 Bio-Rad 半乾式電泳轉漬槽。將 SDS-PAGE 切除集膠體的部份後,將膠片浸泡在轉漬緩衝液(Transfer buffer)中,取轉 印紙(硝化纖維紙,nitrocellulose paper)切成膠片大小後浸在轉漬緩衝液 中,先在 Bio-Rad 半乾式電泳轉漬槽上鋪上三張以轉漬緩衝液浸濕的 3M 濾紙後,依序鋪上溼潤的轉印紙、濕潤的膠片後,再鋪上三張以轉漬緩 衝液浸濕的 3M 濾紙(過程中需趕走每層間的氣泡),以 200mA 轉印一小 時。. 轉印後,將轉印紙放入含有 5% 脫脂牛奶的 1x TBST 中,室溫搖晃 一小時進行 Blocking,以填平轉印紙上沒有蛋白質轉印的空間。倒掉 Blocking 的牛奶後,加入第一次抗體 anti-T7 tag(1:1000 稀釋),置於室 溫搖盪反應一至二小時或於 4℃反應至隔夜。倒掉或回收第一次抗體重複 使用,以 1x TBST 清洗七分鐘三次後,加入第二次抗體 anti-mouse Co-AP(1:1000 稀釋),置於室溫搖盪反應一至二小時。倒掉或回收第二 次抗體後,以 1x TBST 清洗七分鐘三次,加入 NBT/TCIP 顯色劑,反應 至呈色(約十至二十分鐘),加入二次水洗去顯色劑以中止反應,晾乾後保 存。. 30.
(41) 12. SARS PLpro 活性分析 利用 SARS PLpro 會辨識並切割 LNGG 片段之特性,設計出兩個活 性分析試驗。. (1) 利用 HRP(Horseradish Peroxidase)為基質(substrate)測定吸光值: 因 HRP 序列中帶有 LAGG 與 LSGG,若 SARS PLpro 具有活性,則可 將 HRP 切割成不具活性之片段,以 HRP 之呈色系統反應則在 O.D405nm 時其吸光下降,可作為判讀 SARS PLpro 是否具有活性。. ¾ ELISA:HRP substrate + PLpro → Incubate at 37℃, 1hr → ABTS/H2O2 → O.D.405nm. (2) SARS PLpro 切割 HRP 活性試驗 利用 HRP(Horseradish Peroxidase)作為 substrate,取 5 μg 與不同濃度 之 PLP(0、0.02、0.04、0.06 μg)混合,於 37℃下反應 6 小時,將蛋白樣 本與不含 SDS 之 2X Sample loading dye 混合,取 5ul 注入 12% Native gel 樣本槽,並將電泳槽置於冰上。先以 80V 進行電泳,待藍色染劑跑到分 離膠體時,調整電壓至 110V 繼續電泳,直到藍色染劑跑到膠體底部,關. 31.
(42) 閉電源取出膠體,再加入 HRP 呈色劑 TMB membrane peroxidase substrate 產生藍色區域。 當藍色區域變小變淡,則表示具活性之 HRP 較少,即 被 SARS PLpro 切割,可判讀 SARS PLpro 是否具有活性。. ¾ Native gel:HRP substrate + PLpro →TMB menbrane peroxidase substrate. 32.
(43) 標的二:構築 SARS PLpro 表現細胞. Flowchart II:. Cloning:SARS-PLpro-HSV tag/pcDNA3.1 His C. Transfect to HL-CZ. Western blotting. Immuno-fluorescence staining. 33.
(44) 1. 構築 SARS PLpro 之細胞內表現載體 將 cDNA 與設計的引子 SARS PLpro-F 5′- CTC CGA ATT CAA CTC TCT AAA TGA GCC GCT TGT C -3′ 以及 SARS PLpro-R 5′- GAG GCT CGA GAT CCT CTG GGT CTT CAG GAG CGA GTT CTG GCT GTA CGA CAC AGG CTT GAT GGT TGT AGT G - 3′,利用聚合酶連 鎖反應(PCR)將SARS PLpro基因從cDNA選殖出來。其中Reverse之引子帶 有HSV-tag(畫底線處)。 將已選殖到的 SARS PLpro 基因架接到 pcDNA3.1 His C(+)上的 Eco RI 及 XhoI 限制酶切位上。再將此構築好的表現載體利用 Heat-shock 的方 式轉型(transform)到 DH5α 大腸桿菌中並做篩選。經限制酶水解釋驗確定 切割載體之片段大小正確後,將載體素定序。. 2. DNA 轉染(DNA transfection) 細胞的轉染是利用 lipofection 技術,Arrest-In transfection reagent 試 劑(Open BIOSYSTEM),利用 Direct Hydrophilic Conjugation(DHC)技術來 提高在細胞株達到最佳的轉殖效率。DHC 技術合成的多種修飾後的 polymer 所組成。可提供 DNA /Arrest-InTM complexes 做最有效的結合,也 可以降低 cytotoxicity,提高轉殖的結果。. 34.
(45) 懸浮細胞轉染實驗當天將 3ug SARS PLpro 細胞內表現質體與 pEGFP 以 9:1 比例混合,以 500ul 不含胎牛血清的培養液稀釋;將 15ul Transfection Reagent 以 500ul 不含胎牛血清的培養液稀釋,再將以上兩者 混合均勻靜置於室溫 20-30 分鐘,使得 DNA /Arrest InTM complexes 形成。 將要轉染的細胞移去舊的培養液,以 PBS 清洗,將 DNA /Arrest InTM complexes 加入,於 37℃,5% CO2 下培養 4-5 小時後,再更換含有胎牛 血清的培養液繼續培養 48-72 小時。. 3. 細胞內分子之螢光 IF (Intracelluar fluorescence staining) 收集HL-CZ細胞培養液以1500 rpm離心5分鐘,沉降之細胞以PBS (Phosphate buffer saline)溶液清洗後再離心,將細胞以10 % methanol 浸 泡並置於-20°C 固定l 小時,再以PBS 清洗離心,倒掉上清液,在PBS 中加入anti-His tag (1:5000)或anti-HSV tag(1:5000),在室溫下反應60分鐘 後重複清洗離心的步驟。再加入anti-Rhodamin (1:5000) 避光60分鐘,最 後經兩次以上的清洗去除多餘的染劑後加入少許PBS 使細胞懸浮其中, 將懸浮液倒在培養盤中於螢光顯微鏡下觀察、拍照。. 35.
(46) 4. 西方墨點法(Western blotting): 將 Mock cell(HL-CZ-pEGFP)與 SARS PLpro 表現細胞離心、收集下 來,以 PBS 清洗後,100℃加熱 5 分鐘後,以 SDS-PAGE 膠體電泳分離 但白後,再將 SDS-PAGE 轉印到轉印紙後,以 anti-HSV tag(1:1000 稀釋) 為第一次抗體,置於室溫搖晃反應一至二小時或於 4℃反應至隔夜,再以 anti-mouse Co-AP(1:1000 稀釋)為第二次抗體,置於室溫搖盪反應一至 二小時後,加入 NBT/TCIP 顯色劑,反應至呈色(約十至二十分鐘),加入 二次水洗去顯色劑以中止反應,晾乾後保存。. 36.
(47) 標的三:SARS PLpro 表現細胞經干擾素刺激前後之訊息分析. Flowchart III:. Signal pathway assay:cis-acting plasmid. Real-time PCR: PKR mRNA. ELISA:Interferon α expression. 37.
(48) 1. 細胞內訊息傳遞之分析 利 用 Luciferase 報 告 基 因 質 體. pISRE(interferon-stimulated. response element)-Luc,pNF-κB-Luc等,接受訊號傳遞所活化的轉錄 因子結合後,即可表現Luciferase。依Luciferase活性,可快速檢測及 定量(圖3-1) 。各種Enhancers的活化可以顯示何訊息傳遞途徑與細胞 激素或細胞凋亡反應,如NF-κB enhancer可檢測經IKK/NF-κB途徑, 引發Stress及Cell cycle的反應;而ISRE可檢測經JAK/STAT途徑,引發 Inflammation 及 cell cycle 的 反 應 。 而 共 同 轉 染 所 送 入 細 胞 之 質 體 pRenilla-Luc則作為Internal control以比對出Luciferase之表現程度。 實驗當天先準備HL-CZ細胞,培養至六孔盤中,細胞約七分滿 後,測pISRE-Luc、pRenilla-Luc核酸濃度並以9:1之比例混合轉殖入 細胞內之質體量,使六孔盤每格3ug/ml。準備兩管15㏄兩管離心管, 取A離心管內加入ISRE-Luc、pRenilla-Luc、 RPMI-1640 serum free達 體積3ml,取B離心管內加入Arrest-In、RPMI-1640 serum free達體積 3ml。將A、B管液體混和均勻, 靜置20分鐘。將6孔盤中之HL-CZ吸 出,以PBS清洗後,各加入A、B管混和液1mL,再將細胞培養至六孔 盤中,放入培養箱,反應5小時後加入含10 % FBS的RPMI-1640 1mL/well,再放入培養箱中培養。. 38.
(49) 第三天將六孔盤中之細胞培養到二十四孔盤中,加入Interferon α、Interferon β(各3000U/μL)反應4小時後將細胞吸出,以PBS 清洗, 加入Lysis buffer 100μl/well,作用15分鐘後將細胞離心,取20μl上清 液,放入測Luciferase assay 專用96孔盤中,加入Lucferrase 試劑 100μl/well,測冷光量,紀錄。再加入Lucferrase-STOP 試劑100μl/well, 測STOP冷光量,紀錄並做統計。 In vivo signal transduction pathway cis-reporting system. 圖 3-1. 2. 細胞 RNA 抽取(purification of cell RNA) 7. 將 Mock cell(HL-CZ-pEGFP)與 HL-CZ-PLP cell 以 1×10 個為一組, 各取兩組。其中一組 Mock cell 與 HL-CZ-PLP cell 分別加入 Interferon α 3000U/ml,另一組則維持不變,放入培養箱培養四小時後抽取其細胞全 mRNA。細胞 RNA 的抽取是使用 PureLink 39. TM. Micro-to-Midi Kit (Total.
(50) RNA Purification System,Invitrogen),先將 HL-CZ 細胞準備好 1*107 個 /dish,加入 Interferon α 處理四小時後,將 HL-CZ 細胞直接以培養液 RPMI-1640 沖洗下來,離心 300 G、五分鐘去除培養液後,再以磷酸鹽溶 液 PBS(Phosphate buffer saline)清洗,離心 300 G、五分鐘去上清液, 換為 1.5 cc 微量離心管,離心 300 G、五分鐘去上清液去上清,各加入 0.6ml 的 RNA Lysis buffer (2-Mercaptoethonal:RNA Lysis solution =1:100),用 空針 syringing 數次後,離心 500 G、5 分鐘去上清液,加入等量 0.6ml 70% 酒精後,移到 spin column 中過濾,離心 10000 G、2 分鐘去除下方 wash tube 中之廢液,換 wash tube 後加入 wash buffer I 0.7 ml 離心 10000 G、3 分鐘 去除下方 wash tube 中之廢液,加入 wash buffer II 0.5 ml 離心 10000 G、3 分鐘去除下方 wash tube 中之廢液,再加入 wash buffer II 0.5 ml 離心 10000 G、3 分鐘去上清液。換上新的 1.5 ml 離心管,加入 RNase-free Water 50μl, 靜置 2 分鐘後,離心 10000 G、2 分鐘,將 RNA 溶出,溶出來的 RNA 先 轉錄成為 cDNA 後,將剩餘的 RNA 放置於-80 ℃冰箱保存。. 3. 反轉錄聚合酶連鎖反應(Reverse transcriptase PCR) 解凍保存於-80℃冰箱中的細胞 RNA,測量其 OD260 值,定量 RNA, 取細胞 RNA 1μg,加入 RNase-free Water 至體積至 11μl,在 PCR 反應器 上,以 55℃加熱 15 分鐘後,立即插入冰上冷卻。加入 10mM dNTP 混合 40.
(51) 液 1μl 與 0.5 μg/ul Oligo dT primer 1μl,加完後放置於 65℃中 5 分鐘,接 著在冰上依序加入 5 倍的第一股反應緩衝液 (20 mM Tris-HCL pH = 7.5 ,0.1mM EDTA,0.01%(V/V)NP-40,50%(V/V)glycerol 100mM MgCl2) 4μl,0.1mM Dithiothreitol (DTT) 2μl,加完後放置於 42℃中平衡溫度。2 分 鐘 後 加 入 濃 度 為 200U /μl 之 反 轉 錄 酵 素 Superscript III Reverse Transcriptase (Invitrogen) 1 μl,於 42℃反應 80 分鐘進行第一股互補 DNA (cDNA)的合成,再將溫度提高到 70℃,反應 15 分鐘,得到與第一股互 補 DNA(cDNA),以此互補第一股之 DNA 當作接著進行聚合酵素連鎖反 應 (PCR)的模板。. 4. 即時定量聚合酶連鎖反應(real-time polymerase chain reaction) 水解探針(Hydrolysis probe or TaqMan probe) 在 0.2 ml 微量離心管(Optical tubes)中,分別取 cDNA 2.5μl 作為 模版(template),依序加入 Master Mix with dUTP & ROX(Roche) 12.5 μl, ddH2O 8.25μl,再各自加入 OAS、 PKR 與 GAPDH 三對 primer F 與 primer R(1mM)各 0.25μl,與其各自的 probe 0.25μl,使總體積為 25μl。 再置於 ABI 7000 的反應槽中。先以 50℃作用 2 分鐘,後以 95℃作用十 分鐘,再以每個週期,95℃作用十五秒,58℃作用一分鐘,並偵測其螢 光數值,進行 40 個反應週期。最後計算其 Ct 值與複製倍數。 41.
(52) PKR Primer: 5’-CAACC AGCGG TTGAC TTTTT -3’ 5’-ATCCA GGAAG GCAAA CTGAA -3’ OAS. Primer :. 5’-GATGT GGTTA GGTTT ATAGCTG -3’ 5’-TTGGG GGTTA GGTTT CTGCCTTT -3’ GAPDH. Primer:. 5’-AGCCA CATCG CTCAG ACAC -3’ 5’-GCCCCA ATACG ACCAA ATCC -3’. 5. 酵素連結免疫吸附分析法(Enzyme-linked immunoassay;ELISA) 將固定數量之 HL-CZ 細胞培養至 96 孔盤中,以八格為一組,Mock 細胞一組加入 Interferon β 1000U/ml,另一組不加入 Interferon β 之對照 組,PLpro 表現細胞同樣一組加入 Interferon β 1000U/ml,另一組不加入 Interferon β 之對照組,放入培養箱中培養兩天。 兩天後將 96 孔盤每格加入 10ul 福馬林固定細胞,並將 96 孔盤放置 在抽風櫥至乾燥,再加入 PBS 輕輕清洗 3 次。96 孔盤每格加入 200ul 含 10% 丙酮之 PBS 室溫反應 2 小時後,以 PBS 輕輕清洗 3 次,加入 200ul 含 5% 脫脂牛奶的 1x TBST 中,室溫搖盪一小時進行 Blocking。倒掉 Blocking 的牛奶後,加入 anti-Interferon α 抗體(abcam,1:5000 稀釋),. 42.
(53) 置於室溫搖盪反應二小時後,倒掉抗體,以 1x TBST 清洗七分鐘三次後, 加入第二次抗體 anti-mouse Co AP(Zymed,1:5000 稀釋),置於室溫搖盪 反應二小時。倒掉第二次抗體後,以 1x TBST 清洗七分鐘三次,加入 p-Nitrophenyl phosphate liquid substrate(SIGMA)呈色,再以 O.D.(405nm) 測其吸光值。. 43.
(54) 標的四:以二維電泳分析 SARS PLpro 表現細胞經干擾素刺激前後之蛋 白質表現 1. 蛋白質二維電泳: HL-CZ-SARS-PLpro-HSV /pcDNA3.1+pEGFP. HL-CZ-pEGFP+pcDNA3.1. 1×107 cell. 1×107 cell. 1×107 cell. Treat IFN-α. 1×107 cell. Treat IFN-α. 37℃ 3 Days. 37℃ 3 Days. 2D gel electrophoresis. 2D gel electrophoresis. 7. 將 Mock cell(HL-CZ-pEGFP)與 HL-CZ-PLP cell 以 1×10 個為一組, 各取兩組。其中一組 Mock cell 與 HL-CZ-PLP cell 分別加入 Interferon α 3000U/ml,另一組則維持不變,放入培養箱培養三天。. 三天後,將細胞各別收到 15ml 離心管中,以 PBS 清洗三次,去除上 清液後,加入含有蛋白質抑制劑及 DTT 的 Lysis buffer 300 μL ,均勻混 合後,在 4℃反應 4 小時,再以超音波震盪機震盪 15 秒,以 10000 G,4 ℃離心 30 分鐘後,將上清液移到新的微量離心管中。. 44.
(55) 測定出蛋白質濃度後,取一定量蛋白質(120 μg)加入 Rehydration buffer 至體積 250 μL 後,混合均勻,加到 strip holder 中,再將 strip 正面 朝下,放置到 strip holder 中,並確定無產生氣泡後,加入 Cover fluid(450 μL)覆蓋在 strip 上,再蓋上 strip holder 蓋子。. 將 strip holder 移至 IPGphor II(GE, Amersham Biosciences)上,蓋上機 器上蓋,設定條件為:Rehydration 16 hours,IEF at 20℃,50 μA/strip , S1 step 500V 500VHr,S2 step 1000V 1000VHr,S3 step 2000V 2000VHr, S4 step 4000V 4000VHr, S5 step 6000V 6000VHr, S6 step 8000V 8000VHr, S7 step 300V 0:00VHr。設定完成後,按下開始鈕,即開始 各步驟。. 待完成 IEF 電泳後,將 strip 取出,放到含有 50 mg DTT,5 ml SDS equilibration buffer 之玻璃管中,進行 Equilibration 15 分鐘後,倒去含有 50 mg DTT , 5 ml SDS equilibration buffer , 再 加 入 含 有 125 mg iodoacetamide(IAA),5 ml SDS equilibration buffer,進行 Equilibration 15 分鐘。. 將 strip 取出,放入 10 % polyacrylamide gel 上方,並在 strip 正極旁 邊放入一小片沾有蛋白質分子大小標誌液之濾紙,加入 0.5% agarose 45.
(56) sealing solution 至滿,待 agarose sealing solution 凝固後,將 polyacrylamide gel 移到電泳槽(SE 600 Ruby, GE, Amersham Biosciences)中,放入電泳緩 衝液(Running buffer),以 300V,15mA 進行電泳,約 18 小時可跑完。. 2. 銀染色法: 將 polyacrylamide gel 取 下 , 依 PlusOne. TM. Silver Staining. Kit(Amersham Biosciences)步驟染色,加入固定液固定 30 分鐘,倒去固定 液,加入致敏感液反應 30 分鐘使膠體增加敏感度後,倒去致敏感液,以 二次水清洗三次,每次五分鐘。再加入銀染劑染色 20 分鐘,倒去銀染劑, 以二次水清洗兩次,每次一分鐘,加入顯色液反應 2-5 分鐘後,倒去顯色 液再加入中止反應液反應 10 分鐘,以二次水清洗三次,每次五分鐘,最 後加入保存液後保存膠體。. 3. 二維電泳膠體分析: 將膠體以 ImageScanner(GE, Amersham Biosciences)做掃描影像保 存,再用 ImageMasterTM 2D Platinum Software(GE, Amersham Biosciences) 做影像分析,判別出蛋白質點表現與否及表現量之差異。. 46.
(57) 標的五:以質譜儀技術分析二維電泳之蛋白質點,並探討其影響之訊息 及蛋白網絡架構。 1. In gel digest 將蛋白質點挖下,放入微量離心管中,加入100 μl含50% ACN之100 mM ammonium bicarbonate buffer (pH 8.0),置於室溫10分鐘後,吸去上清 液,加入100 μl ACN放置於室溫5分鐘,直到膠體收縮,移去上清之ACN, 放置於室溫30分鐘至膠體乾燥後,再加入含10% trypsin之50 mM ammonium bicarbonate buffer (pH 8.0),放置於37℃、16小時,隔天加入 elution solution (50% ACN in 5.0% TFA)使胜肽片段釋出至上清中。. 2. LC-MS 使用Ultimate capillary LC system (LC Packings, Amsterdam, The Netherlands) 連接至 QSTARXL quadrupole-time of flight (Q-TOF) mass spectrometer (Applied Biosystem/MDS Sciex, Foster City, CA, USA)以分析胜肽片段,並將分析到之 Protein ID 數據比對 NCBI 及 MASCOT search program( http://www.matrixscience.com)資料庫比對。. 47.
(58) 第四章 研究結果 標的一:SARS PLpro 重組蛋白之表現與其活性分析 1. SARS PLpro 之質體構築 將 SARS PLpro 之序列片段以聚合酶連鎖反應夾出,得到大小約 1.3kb 之 SARS PLpro 產物(圖 4-1-A )。最後以限制酵素水解試驗確定所構築之 pET24a 載體中所帶之片段大小為 1.3kb(圖 4-1-B )後,再送定序確定為 SARS PLpro 之序列。. 2.SARS PLpro 之表現及純化 將帶有 SARS PLpro 片段之 pET 載體再轉型到大腸桿菌 BL21(DE3) 中,經過不同濃度 IPTG 誘導後,將細菌粗萃取物利用 SDS-PAGE 分析, 得到表現在不可溶性蛋白部份的 SARS PLpro 蛋白,分子量約 55kDa(圖 4-2-A.B.)。 再將 SARS PLpro 蛋白大量表現後,利用 LC 純化並重摺疊蛋白,使 SARS PLpro 恢復到具活性的構型,並將純化到的蛋白以 SDS-PAGE 及 Western blotting 分析(圖 4-3-A.B.),可以在分子量約 55kDa 的位置得到單 一的色帶。. 48.
(59) 3. SARS PLpro 活性分析 利用 SARS PLpro 會辨識並切割 LNGG 片段之特性,設計出兩個活 性分析試驗。 (1) 利用 HRP(Horseradish Peroxidase)做為基質(substrate): 因 HRP 序列中帶有 LAGG 與 LSGG,若 SARS PLpro 具有活性,則可 將 HRP 切割成不具活性之片段,以 HRP 之呈色系統反應則在 O.D405nm 時其吸光下降,可作為判讀 SARS PLpro 是否具有活性。發現在有 SARS PLpro 蛋白存在下,隨著 SARS PLpro 濃度上升,HRP 在 O.D405nm 吸光 值亦隨之下降,可得知 SARS PLpro 具有活性(圖 4-4)。. (2) SARS PLpro 切割 HRP 活性試驗 利用 HRP(Horseradish Peroxidase)作為 substrate,取 5 μg 與不同濃度 之 PLP(0、2、4、6 μg/ml)混合,於 37℃下反應 6 小時,以 12% Native gel 將蛋白展開後,再加入 HRP 呈色劑 TMB membrane peroxidase substrate 產生藍色區域。當 SARS PLpro 濃度上升,藍色區域則隨之變小變淡,表 示具活性之 HRP 較少,即被 SARS PLpro 切割,可得知 SARS PLpro 具 有活性。(圖 4-5). 49.
(60) 標的二:構築 SARS PLpro 表現細胞 1. SARS PLpro 之細胞表現質體構築 以設計帶有 HSV tag 之反股引子將 SARS PLpro 以 PCR 方式夾出, 得到大小約 1.3kb 之片段(圖 4-6-A),最後再以限制酵素水解試驗確定所 構築之 pcDNA3.1 載體中所帶之片段大小為 1.3kb,(圖 4-6-B ),再送定序 確定為 SARS PLpro 之序列。. 2.以免疫螢光染色做細胞內蛋白表現之確認 (1) 利用 Anti-His tag 作為質體是否送入細胞內之確認: 將細胞以福馬林固定、甲醇打洞後,以 anti-His tag 作為一抗,在室溫 反應一小時後,加入 anti-mouse Rhodamin 作二抗,室溫避光反應一小時 後 , 以 螢 光 顯 微 鏡 觀 察 ( 圖 4-7) , 可 發 現 到 不 論 是 Mock 細 胞 (HL-CZ-pEGFP+pcDNA3.1)或是 SARS PLpro 表現細胞都可以看到紅色螢 光之 Rhodamin,表示質體 pcDNA3.1 有送入細胞中,另外,可以看到 pEGFP 所呈現之綠光,可知 pEGFP 亦有在細胞中表現。. (2) 利用 Anti-HSV tag 作為質體是否送入細胞內之確認: 將細胞以福馬林固定、甲醇打洞後,以 anti-HSV tag 作為一抗,在室 溫反應一小時後,,加入 anti-mouse Rhodamin 作二抗,室溫避光反應一 小時後,以共軛焦顯微鏡觀察(圖 4-8),發現只有 SARS PLpro 表現細胞 50.
(61) 可以看到紅色螢光之 Rhodamin,表示 SARS PLpro 之 HSV tag 被染色, 即 SARS PLpro 有送入細胞內表現。. 3. 西方墨點法確認細胞內表現 PLpro 將構築之 SARS PLpro 細胞株收下部份細胞,以 12% SDS-PAGE 將 蛋白展開。再將 SDS-PAGE 轉印到 NC PAPER 上後,以 anti-HSV tag 做 一抗室溫反應 2 小時後,加入二抗 anti-mouse Co-AP 室溫反應 2 小時, 加入 NBT/TCIP 顯色劑,反應至呈色(約十至二十分鐘),加入二次水洗去 顯色劑以中止反應,晾乾後保存。 可以發現 SARS PLpro 表現細胞在 52kDa 位置有表現,而 Mock 細胞 (HL-CZ-pEGFP+pcDNA3.1)則無(圖 4-9-A ),可以得知 SARS PLpro 細胞 確實有表現 SARS PLpro 蛋白。 同樣以西方墨點法確認細胞是否表現 SARS PLpro 蛋白,將帶有 Anti-His tag 之奈米金抗體與 NC PAPER 結合反應,可以在 SARS PLpro 表現細胞之 52kDa 位置有 SARS PLpro 蛋白色帶可觀察到,而 Mock 細胞 (HL-CZ-pEGFP+pcDNA3.1)則無(圖 4-9-B )。 另外以 β-actin 為一抗反應,以 anti-mouse Co-AP 為二抗呈色,作為 西方墨點法之 Internal control。. 51.
(62) 標的三:SARS PLpro 表現細胞經干擾素刺激前後之訊息分析 1. 細胞內訊息傳遞之分析 將 Mock 細胞(HL-CZ-pEGFP+pcDNA3.1)與 SARS PLpro 表現細胞分 別再以 9:1 Co-transfection 帶有 reporter gene 之質體(如:ISRE、NFκB) 與帶有 Renilla 之質體。 (1) 分析 Interferon α 反應前後之 ISRE 表現程度 將已 Co-transfection ISRE 之 Mock 細胞與 SARS PLpro 表現細胞分別 分為兩組,一組加入 interferon α,另一組則維持不變,反應四小時後, 測其 Luciferase 表現量。 結果發現 Mock 細胞在加入 interferon α 前後其 ISRE 表現量上升 66 倍,SARS PLpro 表現細胞在加入 interferon α 前後其 ISRE 表現量上升 31 倍,mock cell 與 HL-CZ-SARS-PLpro 在經過 interferon α treat 後, HL-CZ-SARS-PLpro 只有 mock cell 的 39 %,即 SARS PLpro 有抑制 interferon α 刺激 ISRE 之表現(圖 4-10)。. (2) 分析 LPS 反應前後之 NFκB 表現程度 將已 Co-transfection NFκB 之 Mock 細胞與 SARS PLpro 表現細胞分 別分為兩組,一組加入 LPS,另一組則維持不變,反映四小時後,測其 Luciferase 表現量。. 52.
(63) 結果發現 Mock cell 在 treat LPS 前後 NFκB 訊號有約 1.3 倍的上升, HL-CZ-SARS-PLpro 在 treat LPS 前後 NFκB 訊號約下降成為 0.8 倍,而 mock cell 與 HL-CZ-SARS-PLpro 在經過 LPS treat 後,HL-CZ-SARS-PLpro 只有 mock cell 的 33 %,即 SARS PLpro 有抑制 LPS 刺激 NFκB 之表現(圖 4-11)。. 2. 即時定量聚合酶連鎖反應(real-time polymerase chain reaction) 由數據分析發現,Mock cell 經過 Interferon α 反應四小時後之 PKR 訊息較未以 Interferon α 反應之數值上升約 3500 倍,但 SARS PLpro 表現 細胞在經過 Interferon α 反應後僅較未加入 Interferon α 反應之數值上升約 500 倍,即 SARS PLpro 表現細胞能較 Moce cell 抑制 Interferon α 誘導之 下游基因 PKR 約 7 倍之表現程度(圖 4-12)。. 3. 酵素連結免疫吸附分析法分析 interferon α 表現程度 將等量之 Mock 細胞與 SARS PLpro 細胞培養至 96 孔盤上,各別加 入 interferon β 反應兩天後,將 96 孔盤之細胞固定並打洞,以 anti-interferin α 作為一抗,再以 anti-mouse Co-AP 做為二抗,最後以 p-Nitrophenyl phosphate liquid substrate 呈色,測其 O.D405nm 之吸光值。 Mock cell 在加入 interferon β 反應後,其 interferon α 表現量都有逐步 上升,而 SARS PLpro 表現細胞在處理 interferon β 前之 interferon α 表現 53.
(64) 量約為 Mock 細胞之 57%,在處理 interferon β 1000U/ml 後,SARS PLpro 表現細胞之 interferon α 表現量約為 Mock 細胞之 50%,在處理 interferon β 3000U/ml 後,SARS PLpro 表現細胞之 interferon α 表現量約為 Mock 細 胞之 45%(圖 4-13),表示 SARS PLpro 表現細胞其受到 interferon β 反應後 不如 Mock 細胞會有 interferon α 表現增加的現象,即 SARS PLpro 細胞 有抑制 type I interferon 之刺激、生成之現象。. 54.
(65) PART IV:2D electrophoresis profile 1.二維電泳分析 以二維電泳分析之差異 (圖 4-14-1) ,結果發現,在 Mock cell 共分離 出約 364 個蛋白質點,在經過 Interferon α 刺激過後約有 366 個蛋白質點, 兩者間蛋白質點位置表現一致之比例達 99.17%,在 SARS PLpro 表現細 胞則分離出 405 個蛋白質點,經過 Interferon α 刺激過後約有 398 個蛋白 質點,兩者間蛋白質點位置一致之比例達 96.39%。 而兩兩比對 Mock cell 與 SARS PLpro 表現細胞之蛋白質點,則有約 80.36%蛋白質點位置一致,經過 Interferon α 刺激過後約 79.32%蛋白質點 位置一致(表 4-1-1)。並以 Mock cell+IFN 為 Reference Gel,將表現量有差 異之蛋白質點列出圖(圖 4-14-2),其餘尚在分析中。 發現同種細胞在加入 Interferon 前後之蛋白質點表現位置之相同比例 皆達到 90%以上,即其同種細胞之電泳再現性存在。而在加入 Interferon 後會造成其蛋白質表現量之上升或下降之差異。. 55.
(66) PART V:Identified MASS profile 將以 Mock cell+IFN 為 Reference Gel 比對,表現量有差異之蛋白質 點以質譜儀分析,並以其數值比對出可能之蛋白質點為何種蛋白(表 4-2)。 其餘差異之蛋白質點尚在分析中。. 56.
(67) 第五章 討論 在本研究中,重組 SARS PLpro 蛋白藉由 SARS PLpro 活性試驗分析 此重摺疊之重組蛋白,發現此片段之 SARS PLpro 可抑制帶有 LAGG、 LSGG 蛋白序列的 HRP 在 O.D405nm 之吸光值,且在以 Native gel 分析 時,可觀察到 HRP 之表現量隨著 SARS PLpro 濃度上升而下降,與已知 文獻所表示 SARS PLpro 具有特異性的辨識 LNGG 蛋白序列,並切割於 LNGG 的後方之蛋白酶活性相印證(Thiel et al., 2003; Lindner et al., 2005; Barretto et al., 2005),故可推論此重組 SARS PLpro 蛋白是具有活性、功 能性的。 而本研究利用 HRP 作為 SARS PLpro 蛋白酵素切割之受質,並以 HRP 直接呈色之 O.D405nm 吸光值強弱作為 SARS PLpro 活性之判讀,能更快 速、便利且便宜的測試 SARS PLpro 之活性,可用來作為篩選抑制 SARS PLpro 切割活性藥物之新分析平台,提供治療 SARS 病毒之新方向。 在文獻中發現,當 SARS 病毒感染於 STAT1 抑制之老鼠時,SARS 病毒會造成更長期的病毒血症、嚴重的肺部症狀及多種的器官擴散感 染,由此可知 Interferon 對 SARS 病毒具有抑制其病毒複製與致病之作用 (Hogan, R. J. et al., 2004) 。因此,在本研究中即針對 SARS PLpro 與 Interferon 之相關聯性,作為抗病毒機制之方向。 本研究發現,在 SARS PLpro 表現細胞之細胞內訊號傳遞途徑試驗. 57.
(68) (Reporter gene assay)中,SARS PLpro 能抑制 Interferon α 刺激細胞內 ISRE(interferon-stimulated response element)訊息之表現,使 ISRE 訊息表 現程度下降。並以 Real-time RT-PCR 測定 ISRE 其下游基因 PKR 之表現, 發現 SARS PLpro 表現細胞在經過第一型干擾素刺激後,其 PKR 之表現 較同樣經第一型干擾素刺激之未表現 SARS PLpro 細胞表現程度下降。由 此實驗可證實,SARS PLpro 之表現可抑制第一型干擾素之下游訊息及基 因之表現,使 ISRE 與下游基因 PKR 之表現程度皆下降。再利用 ELISA 實驗發現,當 Mock cell 經由 Interferon β 刺激後,其 Interferon α 會隨之 上升,但 SARS PLpro 表現細胞在同樣經過 Interferon β 刺激後其 Interferon α 之表現量僅有 Mock cell 之 50%,即 SARS PLpro 蛋白之表現使 Interferon β 促使之細胞 Interferon α 表現量受到抑制,故可證實 SARS PLpro 不但抑 制細胞中第一型干擾素誘導之下游訊息及基因之表現,也抑制第一型干 擾素誘導之蛋白表現,與先前文獻發現 SARS PLpro 能水解 Interferon 所 誘導之蛋白 ISG15(Barretto et al., 2005)對照之下,發現 SARS PLpro 與 Interferon 之間相關聯性越顯重要。 另外,當 SARS PLpro 表現細胞經過 LPS 刺激,以促使發炎反應過 後,其 NFκB 之訊息表現程度僅為 Mock cell 經 LPS 刺激後之 33%,即 SARS PLpro 能抑制細胞中發炎相關轉錄因子 NFκB 之訊息表現。 病毒藉由抑制 Interferon 以達到破壞細胞之免疫系統作用為常見之模. 58.
(69) 式,在其他文獻中發現,如:流行性感冒病毒(influenza virus)之 NS1 蛋 白會藉由與 double-stranded RNA 結合達到抑制細胞之干擾素誘導之免疫 反應(Bergmann, M. et al., 2000;Hatada, E. et al., 1992 Lu, Y. et al., 1995); 伊波拉病毒(Ebola virus)所轉譯出的蛋白 VP35 會藉由結合並隔離開 double-stranded RNA 以達到抑制 Interferon 之作用,而蛋白 VP24 則藉由 妨礙活化的 STAT1 進入細胞核以達到抑制 Interferon 之作用(Basler, C. F. et al., 2000 Reid, S. P. et al., 2006);尼帕病毒(Nipah virus)會轉譯出 3 種蛋 白去中和、抑制干擾素上游 STAT1 以達到逃避細胞免疫反應之作用(Park, M. S. et al., 2003)。而 SARS 之 ORF3b、ORF6 及核蛋白衣(nucleocapsid) 被發現具有能中和 Interferon 之功效(Sarah A. et al., 2007) ,因此我們推論 SARS PLpro 具有中和 Interferon 之功效,能使細胞之免疫能力下降,保 護 SARS 病毒不受感染細胞之免疫攻擊。 以二維電泳檢測細胞全面性的蛋白質表現,初步以 Mock cell 加入 Interferon α 為 Reference gel 比對其他膠體發現,蛋白質點 697、635、691 與 SARS PLpro 表現細胞之同位置蛋白質點有表現量之差異,經由 MASS 分析蛋白質點分別為:Heat shock 27 kDa protein、Chloride intracellular channel protein 1、Myosin-9 與 NADH dehydrogenase iron-sulfur protein 3, 其中 Heat shock 27 kDa protein 已知與 anti-apoptosis 有關,在受到第一型 干擾素刺激下會增加表現(M. Caraglia1 et al., 1999) ;Chloride intracellular. 59.
(70) channel 可調控細胞內外之電位平衡、液體傳送以及維持細胞體積之功能 (Michael Eisenhut et al., 2006) ;而 NADH dehydrogenase 為粒線體之前驅 物質,在受到第一型干擾素刺激下會增加其表現,但在本研究之二維電 泳分析結果卻發現其表現量為下降。其它二維電泳分析之 Interferon 相關 蛋白質點表現之改變尚在分析中。 在未來的分析中,希望藉由從二維電泳分析出差異性之蛋白質點為 何,並從中探討 SARS PLpro 所影響、改變之蛋白表現與訊息途徑,架構 出整體蛋白質表現網絡系統,以用於將來開發 SARS 診斷及治療之新方 向。. 60.
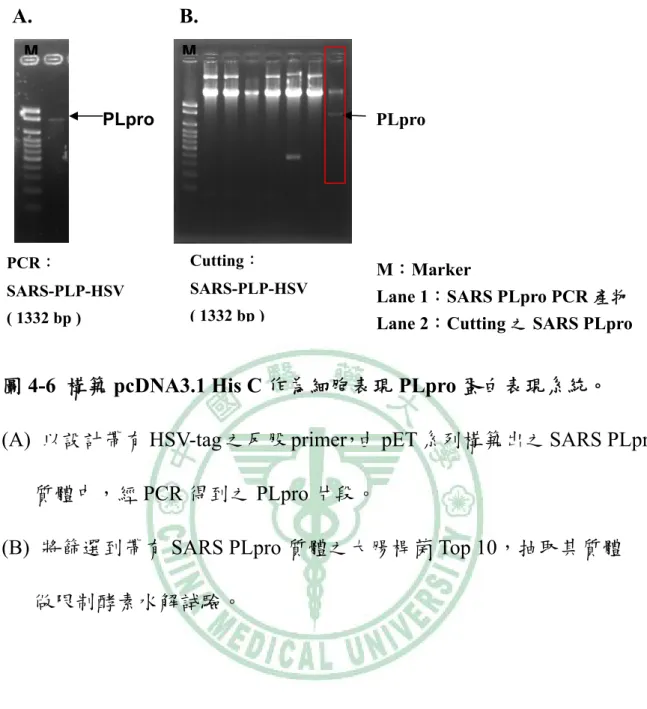
(71) A. M. B. C. M. M. 1500bp 1000bp 600bp 500bp. 1000bp 700bp. S4:589bp. S5:890bp. (4507-5095). (4951-5840). SARS-PLP(1332 bp ) 4507-5840. 圖 4-1 構築 pET 表現系統以作為細胞外表現 PLpro 蛋白。 (A) 為利用 PCR 得到的 SARS PLpro 4507-5095 之片段,以 2 %膠體做電 泳分析。 (B) 為利用 PCR 得到的 SARS PLpro 4951-5840 之片段,以 2 %膠體做電 泳分析。 (C) 將篩選到帶有 SARS PLpro 質體之大腸桿菌 Top 10,抽取其質體做限 制酵素水解試驗,以 1 %膠體做電泳分析。. 61.
(72) A.. M 1. 2. 3. 4. 5. 6. 7. 8 M. 55kDa. B.. M. PLpro. 1. 2. 3. 4. 5. 6. 7. M. 8. 55kDa. 圖 4-2 pET 系列表現 SARS PLpro 蛋白 將 確 定 帶 有 SARS PLpro 片 段 之 pET 載 體 再 轉 形 到 大 腸 桿 菌 BL21(DE3) 中 , 經 過 不 同 濃 度 IPTG 誘 導 後 , 將 細 菌 粗 萃 取 物 利 用 SDS-PAGE 分析。 (A) 經 IPTG 誘導表現之蛋白(pellet),在 55kDa 可看到 PLpro 之表現。 (B) 經 IPTG 誘導表現之蛋白(sup.)。 M:Marker. 1:non-induction. 2:IPTG 50μM. 3:IPTG 100μM. 4:IPTG 400μM. 5:IPTG 500μM. 6:IPTG 1000μM 7:IPTG 1500μM 8:IPTG 2500μM. 62.
(73) A.. B. M. 1. M. 2. 2. 170kDa 130kDa. 116.0kDa 97.4kDa. 95kDa. 66.2kDa. PLpro 37.6kDa. 1. 75kDa 55kDa. PLpro. 43kDa 34kDa. 28.5kDa. M:Marker 1:Non-purified PLpro 2:Purified PLpro. 圖 4-3 pET 系列重組 SARS PLpro 蛋白之表現與純化 將 SARS PLpro 蛋白大量表現後,利用 LC 純化並重摺疊蛋白,使 SARS PLpro 恢復到具活性的構型,再做電泳分析。 (A) 以 12% SDS-PAGE 電泳分析,再以 Commasian blue 染色。 (B) 以 12% SDS-PAGE 電泳後做 Western blotting 分析,用 Anti His-tag 為一抗,Anti mouse Co-AP 為二抗反應後,以 NBT/BCIT 呈色。. 63.
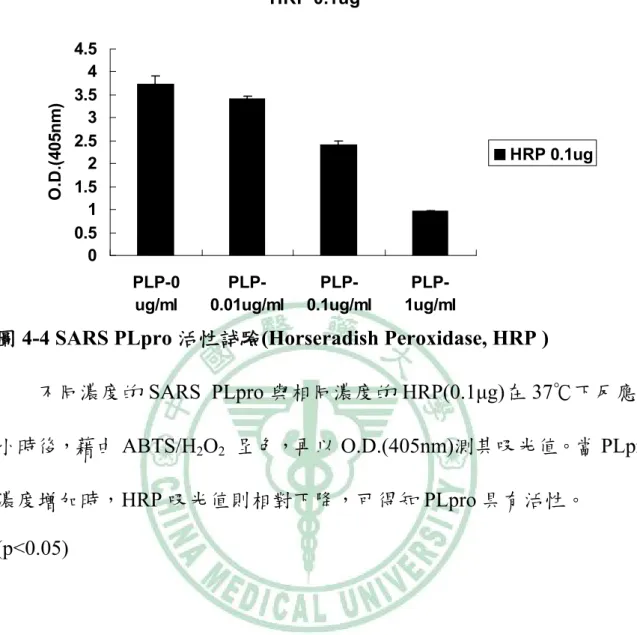
(74) O.D.(405nm). HRP 0.1ug 4.5 4 3.5 3 2.5 2 1.5 1 0.5 0. HRP 0.1ug. PLP-0 ug/ml. PLP0.01ug/ml. PLP0.1ug/ml. PLP1ug/ml. 圖 4-4 SARS PLpro 活性試驗(Horseradish Peroxidase, HRP ) 不同濃度的 SARS PLpro 與相同濃度的 HRP(0.1μg)在 37℃下反應 1 小時後,藉由 ABTS/H2O2 呈色,再以 O.D.(405nm)測其吸光值。當 PLpro 濃度增加時,HRP 吸光值則相對下降,可得知 PLpro 具有活性。 (p<0.05). 64.
(75) 圖 4-5 SARS PLpro 切割 HRP 活性試驗 利用 HRP(Horseradish Peroxidase)作為 substrate,取 5 μg 與不同濃度 之 PLP(0、2、4、6 μg/ml)混合,於 37℃下反應 6 小時,以 Native gel 將 蛋白展開後,加入 HRP 呈色劑 TMB membrane peroxidase substrate 產生 藍色區域,並以 Density meter 換算出藍色區域減少之百分比。. 65.
(76) A.. B. M. M. PLpro. PLpro. PCR:. Cutting:. SARS-PLP-HSV. SARS-PLP-HSV. ( 1332 bp ). ( 1332 bp ). M:Marker Lane 1:SARS PLpro PCR 產物 Lane 2:Cutting 之 SARS PLpro. 圖 4-6 構築 pcDNA3.1 His C 作為細胞表現 PLpro 蛋白表現系統。 (A) 以設計帶有 HSV-tag 之反股 primer,由 pET 系列構築出之 SARS PLpro 質體中,經 PCR 得到之 PLpro 片段。 (B) 將篩選到帶有 SARS PLpro 質體之大腸桿菌 Top 10,抽取其質體 做限制酵素水解試驗。. 66.
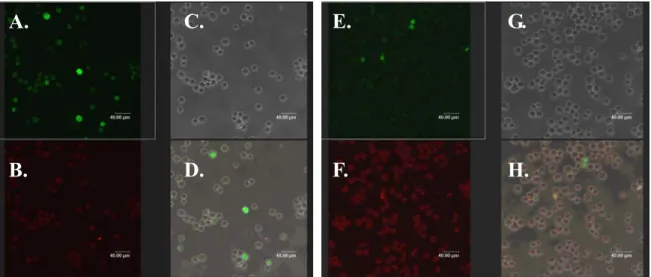
(77) A.. B.. C.. D.. E.. F.. 圖 4-7 以 Anti His-tag 為抗體做免疫螢光染色之細胞內蛋白表現確認 將細胞以福馬林固定、甲醇打洞後,以 Anti His-tag 為第一抗體,在 37℃下反應 1 小時,再以 Anti mouse Rhodamin 為第二抗體,37℃下反應 1 小時,最後以螢光顯微鏡觀察 A~C 圖為以 9:1 比例轉染入 pcDNA3.1 + pEGFP 之 HL-CZ 細胞。 D~F 圖為以 9:1 比例轉染入 SARS PLpro-pcDNA3.1 + pEGFP 之 HL-CZ 細胞。 A、D 圖為在可見光下觀察到之細胞型態,B、E 圖為在螢光激發下觀察 到 pEGFP 之綠光,C、F 圖為在螢光下觀察到載體 pcDNA3.1 因帶有 His-tag,經由染色所觀察到之紅色螢光。. 67.
(78) A.. C.. E.. G.. B.. D.. F.. H.. 圖 4-8 Anti HSV-tag 免疫螢光染色之細胞內 SARS PLpro 蛋白表現確認 以 Anti HSV-tag 為第一抗體,37℃下反應 1 小時,再以 Anti mouse Rhodamin 為第二抗體,37℃下反應 1 小時後,以共軛焦顯微鏡觀察。 A~D 圖為 9:1 比例轉染入 pcDNA3.1 + pEGFP 之 HL-CZ 細胞。 E~H 圖為 9:1 比例轉染入 SARS PLpro-pcDNA3.1 + pEGFP 之 HL-CZ 細 胞。 A、E 圖為在螢光激發下觀察到 pEGFP 之綠光,B、F 圖為在螢光下觀察 到 SARS PLpro 因帶有 HSV-tag,經由染色所觀察到之紅色螢光,C、G 圖為在可見光下觀察到之細胞型態,D、H 圖則為綠光、紅光與可見光之 重疊圖。. 68.
(79) A. B 1. 2. 1. 2. SARS PLpro. 1°:Anti HSV tag 2°:Anti mouse Co-AP. SARS PLpro. Anti HSV tag conjugate nano gold. C 1. 2. 1:Mock cell 2:HL-CZ –SARS-PLpro. β-actin. 1°:Anti β-actin 2°:Anti mouse Co-AP. 圖 4-9 西方墨點法確認細胞內表現 PLpro (A) 以 Anti HSV-tag 為第一抗體,37℃下反應 2 小時,再以 Anti-mouse Co-AP 為第二抗體,37℃下反應 2 小時後呈色。 (B) 以奈米金粒子-Anti His tag 抗體,結合 NC Paper 上蛋白。 (C) 以 Anti β-actin 為第一抗體,37℃下反應 2 小時,再以 Anti-mouse Co-AP 為第二抗體,37℃下反應 2 小時後呈色,作為 Internal control。. 69.
數據



+7



相關文件
SF15140A 楊勝舜 利用 JKB-122 評估對於經干擾素(長效 型或短效型)或干擾素和 Ribavirin 組合 治療沒有反應的 C 型肝炎病毒陽性患 者之肝臟功能 (丙胺酸轉胺酶
6 《中論·觀因緣品》,《佛藏要籍選刊》第 9 冊,上海古籍出版社 1994 年版,第 1
In this paper, we would like to characterize non-radiating volume and surface (faulting) sources for the elastic waves in anisotropic inhomogeneous media.. Each type of the source
Results for such increasing stability phenomena in the inverse source problems for the acoustic, electromagnetic, and elastic waves can be found in [ABF02, BLT10, BHKY18, BLZ20,
Owing to the pandemic of novel coronavirus pneumonia, Visitor Expenditure Survey was suspended in the 2 nd quarter of 2020, and thus no information is released for this
The novel coronavirus outbreak has drawn increased international attention as the virus quickly spreads across the world..
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
Then g is defined on [a, b], satifies (11), and is continuous on [a, b] by the Sequential Characterization of Limits.. Thus, f
