以事件關聯電位(ERP)探索睡眠對於配對學習的促進效果 - 政大學術集成
89
0
0
全文
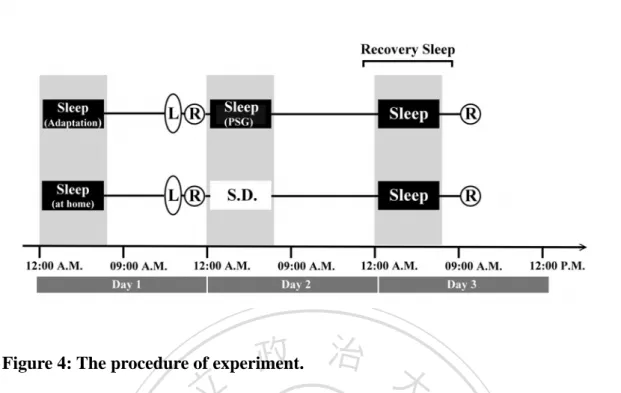
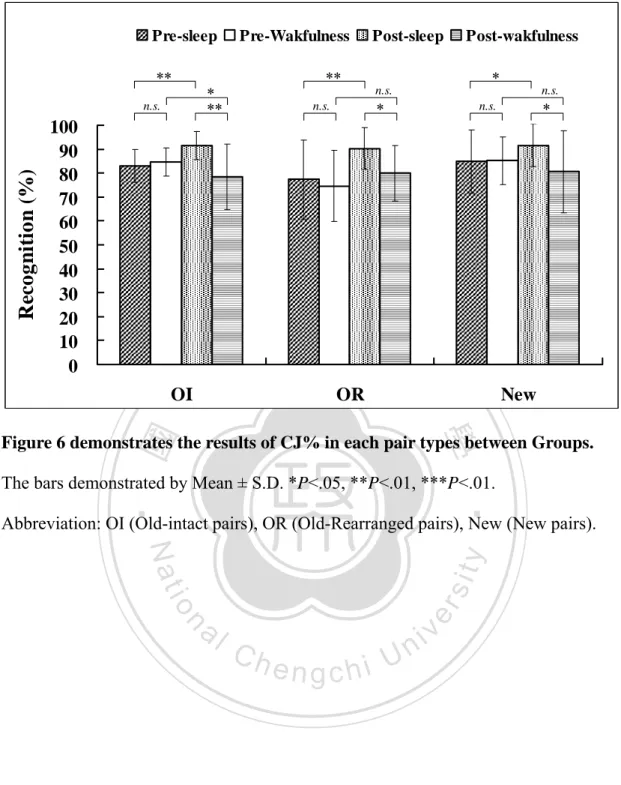
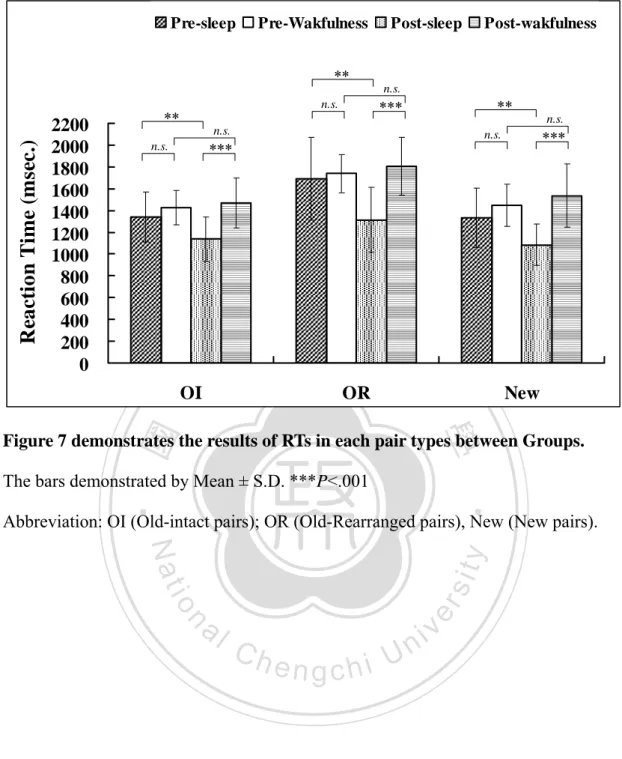
(2) ENGLISH ABSTRACT. The effect of sleep on declarative memory remains contradictory. Prior studies show that sleep benefits the learning of related word pairs consistently, while the learning of unrelated word pairs, however, show mixed results. It is possible that the behavioral measures used in previous studies are not sensitive enough to reveal subtle effects of sleep on new associations. N400, an event-related potential (ERP) component reflecting relatedness among words in semantic memory, was used in the present study to investigate the effect of sleep on the physiological process underlying new associations of unrelated word pairs. Participants were randomly assigned to either a Sleep group or a Wakefulness group. In the learning phase, participants were asked to memorize 80 visually presented unrelated word-pairs, followed by a pre-test phase with a recognition task. The participants then underwent either a night of nocturnal sleep (Sleep group) or sleep deprivation (Wakefulness group). A post-test was conducted after subjects had one night of recovery sleep. During both pre-test and post-test sessions, prime and target words were presented successively for the subjects to judge whether they were among the original pairs or new pairs. ERPs were recorded during both test phases. The behavioral data show that differences in improvement of recognition and decreases in reaction time from pre-test to post-test are significant between Sleep and Wakefulness groups. N400 peak amplitude attenuated significantly after sleep but not after wakefulness. The improvement of recognition negatively correlates with slow wave sleep (SWS). The number of word-pairs acquired in the learning phase, however, correlates positively with SWS. According to the two-stage memory consolidation theory of sleep, SWS involves in redistribution of memory (systematic consolidation). Therefore, that the participants with high performance showed more SWS may reflect the process of systematic consolidation. These results suggest that the sleep has an enhancing effect on the formation of novel association, and SWS may be involved in the process of redistributing memory representations.. 立. 政 治 大. ‧. ‧ 國. 學. n. er. io. sit. y. Nat. al. Ch. engchi. i Un. v. Keywords: sleep, slow wave sleep, sleep deprivation, learning, memory, consolidation, declarative memory, new association, recognition memory, paired-associates learning, event-related potential, ERP, N400. i.
(3) CHINESE ABSTRACT 中文摘要. 睡眠是否能鞏固陳述性記憶目前尚無定論。過去研究一致較支持睡眠能增進 相關字詞配對的學習,但睡眠是否能增進無關字詞配對的學習,目前仍存在不一 致的發現。造成該差異的原因可能是:過去研究多採用的行為測量指標,或許無 法充分反映出睡眠促進記憶新聯結(new association)產生的效果。事件關聯電 位(Event-related potential, ERP)的 N400 反映出語意記憶系統內每個字詞彼此 的相關程度,因此本研究使用 N400 來探討睡眠強化無關字詞配對形成新聯結的 電生理歷程。30 名健康受試者(15 位男性與 15 位女性,平均年齡為 20.7 歲) 隨 機分派至睡眠組或清醒組,第一晚在學習 80 組無關字詞配對後,接受第一次再 認記憶測驗,同時進行 ERP 的記錄。隨後睡眠組接受睡眠記錄(PSG) ,清醒組 則接受整晚的睡眠剝奪,兩組受試者皆在第二晚給予 8 小時的躺床時間,使他們 有機會充足睡眠以恢復精神,於第三天早上接受第二次再認記憶測驗及 ERP 記 錄。在記憶測驗時,無關字詞配對分別組成促發字(prime)與目標字(target) 先後出現,受試者需判斷先後出現的字詞是否為先前學過的完整配對,在測試階 段同時記錄腦電波訊號。行為測量結果顯示睡眠過後,再認表現的正確率顯著提 高且反應時間明顯縮短,但在睡眠剝奪後則顯示相反的結果。電生理測量發現睡 眠組的 N400 振幅在睡眠過後較清醒組明顯降低。另外,睡眠組較清醒組有較高 的正確率與較短的反應時間。睡眠組再認測驗的進步量與慢波睡眠呈現負相關, 而慢波睡眠與第一次再認測驗的正確率呈現正相關,根據二階段睡眠記憶鞏固理 論,慢波睡眠涉及重新組織記憶的歷程(系統性固化),因此學習表現較佳的受 試者出現較多的深度睡眠,可能反應其經歷系統性固化。本研究結果顯示睡眠對 於產生新聯結有明顯的增強效果,而且慢波睡眠可能參與了記憶表徵重新分配的 歷程。. 立. 政 治 大. ‧. ‧ 國. 學. n. er. io. sit. y. Nat. al. Ch. engchi. i Un. v. 關鍵字:睡眠、慢波睡眠、睡眠剝奪、學習、記憶、記憶固化、陳述性記憶、記 憶再認、配對學習、新聯結、事件關聯電位、N400. ii.
(4) ACKNOWLEDGMENTS 致謝 論文在經過五十幾個值班的夜晚,以及許多重要他人的期盼下終於完成,同 時也意謂著漫長的碩士班學習歷程將告一段落,期間曾一度不知如何繼續下去? 放棄的聲音不斷地在耳邊呢喃,但總有人會告訴我再撐一下,謝謝這些曾經鼓勵 我的朋友。同時也感謝我的父母,給我很大的空間作我想做的事情,並給予我最 多的愛與支持,謝謝您們。 論文能夠順利完成,首先要謝謝我的指導教授 楊建銘老師,我曾經ㄧ度無 法適應碩士班生活,各方面表現幾乎能用荒腔走板來形容,所幸老師從未放棄過 學生,並深信學生的能力,給予學生一次又一次的機會,老師常用關心與鼓勵來 支持我每個挫折時候,並適時地協助學生無論是研究上或生活上的困難,讓我的 研究生涯有個很大的蛻變。老師除教導我許多學術上的知識外,同時也在生活生 上提供許多的經驗和意見,讓我受益良多,建銘老師:謝謝您!. 立. 政 治 大. ‧ 國. 學. ‧. 我也要謝謝 蔡玲玲老師和 鄭仕坤老師,十分感謝兩位老師願意百忙中抽空 擔任學生的口試委員。感謝蔡老師提供學生執行睡眠研究時的建議,並且詳細地 給予論文書寫上的建議。也感謝鄭老師提醒學生研究記憶議題時需要注意的事 項,並且提供不同的觀點讓我能夠從不同的角度去思考記憶研究的本質。兩位老 師豐富且寶貴的研究經驗,讓學生在學習作研究時,能夠更加深入且廣泛。. io. sit. y. Nat. n. al. er. 我特別要謝謝大學時候的一位教授 蔡欣志老師,蔡老師是帶領我進入學習 與記憶研究的啟蒙老師,在我研究所就讀期間,老師因病過世,很遺憾沒機會與 您分享我的研究成果,但您對於學生的教導與提攜,卻是我ㄧ輩子將謹記在心的 恩惠,特別將此論文獻給在天上的老師,謝謝您。. Ch. engchi. i Un. v. 論文的完成,我要謝謝我親愛的女朋友 Pinky,在論文書寫過程中,謝謝妳 給予我的鼓勵、支持與督促,讓我有明確的目標與方向,也謝謝妳的陪伴,無論 是開心或悲傷,從妳身上感受到許多的溫暖,並且有動力去實現未來的夢想。也 要謝謝好朋友宗翰,在生活煩悶時,能夠彼此鼓勵和一起瘋狂,豐富了研究所枯 燥的生活。Mina 是一位失聯的朋友,謝謝妳曾在我低潮時激勵我,讓我的論文 計畫書能夠快速完成。另外,也特別謝謝我在長庚睡眠中心的實習督導家碩,謝 謝你讓我發現正向心理學的力量,讓我有信心去完成許多事情。謝謝在美國的朋 友 Stella 和 Dan 協助英文論文的修改與校正。最後當然要謝謝政大睡眠實驗室提 供我幫助與指教的每位夥伴。因為有你們使我能順利完成碩士班的學習生涯。. iii.
(5) TABLE OF CONTENTS. ENGLISH ABSTRACT ……………………………………………………………… i CHINESE ABSTRACT ……………..………………………………………………. ii ACKNOWLEDGMENTS ………..………………………………………………….iii TABLE OF CONTENTS ………..……………………………………..…………… iv. CHAPTER ONE: GENERAL INTRODUCTION ...................................................1 1. STAGES OF SLEEP ....................................................................................................1 2. CATEGORIES OF MEMORY .......................................................................................3 3. SLEEP AND NON-DECLARATIVE MEMORY ...............................................................6 3.1 Effects of Total or Selective Sleep Deprivation on Memory ............................7 3.2 Changes in Recorded Sleep Parameters After Learning .................................9 3.3 Reactivation for Memory Processing During Sleep ...................................... 11 3.4 Facilitation of Memory With Cues During Sleep...........................................12 3.5 Interim Summary............................................................................................13 4. SLEEP AND DECLARATIVE MEMORY ......................................................................14. 立. 政 治 大. ‧ 國. 學. ‧. 4.1 Studies of Total or Selective Sleep Deprivation .............................................14 4.2 Post-learning Modifications in Sleep Architecture........................................18 4.3 Reactivation for Memory Processing During Sleep ......................................20 4.4 Influencing Memory Processing During Sleep by Cuing...............................21 4.5 Interim Summary............................................................................................22 5. NETWORK OF HUMAN MEMORY SYSTEM ..............................................................23 5.1 Representation of Memory Network ..............................................................23 5.2 Priming Paradigm to Measure the Associations Between Words ..................23 6. ERP STUDIES IN SLEEP-DEPENDENT MEMORY ......................................................25 6.1 Introduction to N400 of Event-related Potentials (ERPs) .............................25 6.2 Factors That Affect N400 Amplitude..............................................................26 6.3 Theories and/or Mechanisms of the N400 .....................................................27 6.4 ERP Research in Sleep and Declarative Memory..........................................29. n. er. io. sit. y. Nat. al. Ch. engchi. i Un. v. CHAPTER TWO: PRESENT STUDY ....................................................................31 CHAPTER THREE: MATERIALS AND METHODS..........................................32 1. MATERIALS AND METHODS ...................................................................................32 1.1 Subjects ..........................................................................................................32 1.2 Experimental Procedure ................................................................................32 1.3 Paired-associates Learning List ....................................................................33 iv.
(6) 1.4 Stimuli Presentation .......................................................................................34 2. ELECTROPHYSIOLOGICAL RECORDING ..................................................................36 2.1 Polysomnograhy Recording...........................................................................36 2.2 ERP Recording and Signal Extraction...........................................................36 3. STATISTICAL ANALYSIS .........................................................................................37 3.1 Behavioral Data.............................................................................................37 3.2 ERP Data .......................................................................................................38 3.3 Correlations ...................................................................................................38 CHAPTER FOUR: RESUTS....................................................................................39 1. BEHAVIORAL DATA................................................................................................39 1.1 Percentage of Correct Judgment (CJ%) ........................................................39 1.2 Reaction Time (RT) ........................................................................................39 2. ERP DATA .............................................................................................................40 2.1 N1-P2 Components ........................................................................................40 2.2 N400 Peak Induced by OI Pairs at Fz, Cz and Pz Sites ................................41 2.3 N400 Latency Induced by OI Pairs at Fz, Cz and Pz Sites............................41 2.4 The Peak Amplitude of N400 Evoked by Different Pairs Types at Cz site .....42 2.5 The Peak Latency of N400 Induced by Different Word Pairs at Cz site ........43 3. CORRELATIONS BETWEEN SLEEP STAGES AND PARAMETERS OF MEMORY. 立. 政 治 大. ‧. ‧ 國. 學. sit. y. Nat. MEASURES ................................................................................................................44 4. SUBJECTIVE RATING: STANFORD SLEEPINESS SCALE ............................................44. n. al. er. io. CHAPTER FIVE: DISCUSSION AND CONCLUSIONS.....................................45. LIST OF TABLES. Ch. engchi. i Un. v. Table 1. The Role of Sleep on Unrelated Paired-associates Learning (Total and/or Partial Sleep Deprivation) .......................................................................52 Table 2. The Role of Sleep on Related Paired-associates Learning (Total and/or Partial Sleep Deprivation)...................................................................................53 Table3. Demographic and Subjective Variable....................................................54 Table4. nPSG: Sleep Statistics of Sleep Group ....................................................55 Table5. Correlation Between Memory Measures and Distinct Sleep Stages .......56 LIST OF FIGURES Figure 1: The human sleep cycle .........................................................................57 Figure 2: Memory systems...................................................................................58 Figure 3: The Model of Semantic Memory and Purpose of Present Study. .........59 v.
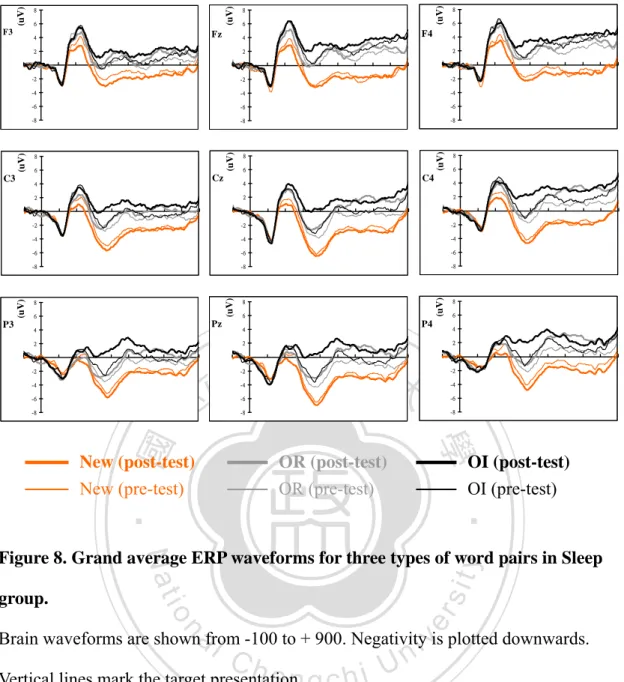
(7) Figure 4: The procedure of experiment. ..............................................................60 Figure 5: The presentation of experiment stimuli................................................61 Figure 6 demonstrates the results of CJ% in each pair types between Groups...62 Figure 7 demonstrates the results of RTs in each pair types between Groups.....63 Figure 8. Grand average ERP waveforms for three types of word pairs in Sleep group. ...................................................................................................................64 Figure 9. Grand average ERP waveforms for three types of word pairs in Wakefulness group. ..............................................................................................65 Figure 10. Grand average ERP waveforms for OI pairs in Sleep group are compared with Wakefulness group.......................................................................66 Figure 11. Grand average ERP evoked by OI pairs. ...........................................67 Figure 12. The N400 induced by different word pairs at Cz site. ........................68. 政 治 大. REFERENCES...........................................................................................................69. 立. SUPLEMENT 1..........................................................................................................80. ‧ 國. 學. APPENDIX I: STANFORD SLEEPINESS SCALE...............................................81 APPENDIX II: LISTS OF PAIRED-ASSOCIATES ..............................................82. ‧. n. er. io. sit. y. Nat. al. Ch. engchi. vi. i Un. v.
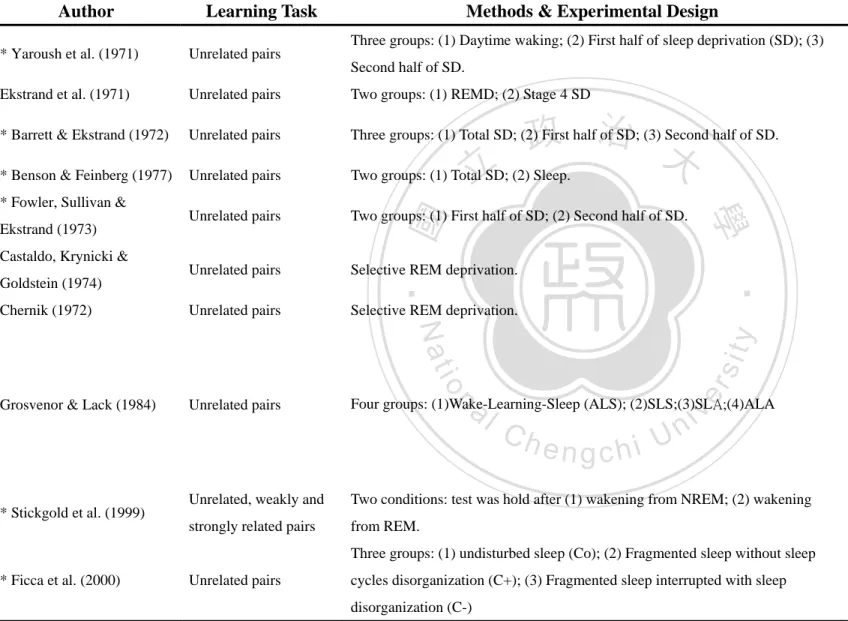
(8) CHAPTER ONE: GENERAL INTRODUCTION. Sleep has been recognized as a universal and essential human need. Sleep occupies one third of our life, and the role of sleep is crucial for one’s physiological and psychological functions during awake. One function of sleep that has been proposed and investigated in many studies is to facilitate memory consolidation. The structure and utilization of memory are central to our knowledge of the past, interpretation of the present, and prediction of the future. We can say that the process. 政 治 大. of our life is the trace of memory and learning. Memory stores information about. 立. everything in our life, and it constructs our precious story. Every thought, every action,. ‧ 國. 學. our very feeling of affection....is based on it. Without memory, all experience could be meaningless. The present thesis is attempted to explore the relationship between these. ‧. two crucial aspects of our life.. y. Nat. sit. This chapter includes four parts: First, an overview of sleep, including its. n. al. er. io. characteristic stages and associated neurobiology. Second, a description of the. i Un. v. categories of memory and its processes. Third, a systematic review of research on the. Ch. engchi. relation between sleep and memory processing, as well as between declarative memory and non-declarative memory, respectively. Finally, studies using event-related potential (ERP) to explore the relationship between sleep and memory are reviewed.. 1. Stages of Sleep The architecture and timing of the sleep stages and cycles can be studied objectively by using polysomnography (PSG). PSG recordings involve the simultaneous monitoring of electrical activity including brain waves (also termed 1.
(9) electroencephalogram, EEG), muscle contraction, eye movement, heart rate, respiratory patterns and blood oxygen levels. The data from PSG have been classified into two distinct types of sleep: non-rapid-eye-movement (NREM) and rapid-eye-movement (REM) sleep. NREM sleep includes four stages, increasing in depth of sleep from 1 to 4, with the final two stages collectively referred to as slow-wave sleep (SWS). In humans, NREM and REM sleep cycles alternate overnight every 90 min in a phenomenon known as an “ultradian cycle”. Although the NREM to REM cycle length generally remains stable throughout the night, the ratio dynamically changes within each 90 min cycle. Consequently, stages 3 and 4 of. 治 政 NREM dominate in the early night, while stage 2 NREM 大and REM sleep reign in the 立 latter half of the night (see figure 1). ‧ 國. 學. When the sleep stages are altered between REM and NREM at night, the brain. ‧. also undergoes significant alternations in neurochemistry. In NREM sleep, SWS is. sit. y. Nat. characterized by high amplitude, synchronous delta EEG, hippocampal sharp vertex. io. er. waves, and a less active cholinergic system in the brainstem and forebrain (Hobson, McCarley, & Wyzinski, 1975; Lydic & Baghdoyan, 1998). Moreover, the firing rate. al. n. iv n C of serotonergic raphe neurons and noradrenergic h e n g c hlocus i Uceruleus neurons are also. reduced relative to waking states (Aston-Jones & Bloom, 1981; Shima, Nakahama, & Yamamoto, 1986). During REM sleep it is characterized by a desynchronized saw-toothed theta rhythm in EEG, suppressed monoaminergic neurons, and elevated cholinergic systems (Kametani & Kawamura, 1990; Marrosu et al., 1995). Dramatic changes in brain electrophysiology, neurochemistry and functional anatomy across these sleep stages make them biologically different from the awake brain (Pace-Schott & Hobson, 2002). This summary describes the neuronal processes of sleep that are affected by the brain transit through wake-sleep states, which means that sleep itself cannot be taken 2.
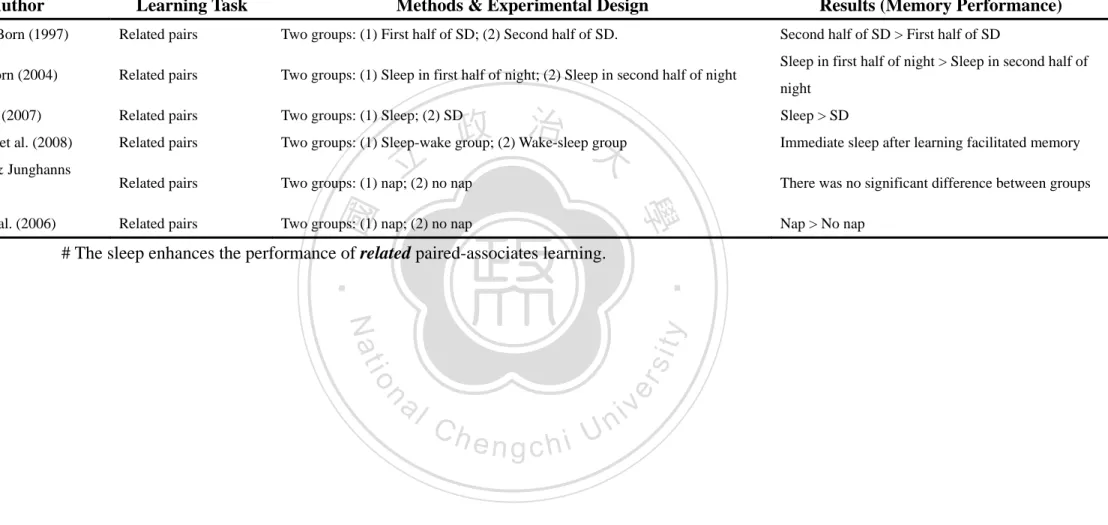
(10) as a homogeneous state. Despite much research, the function of sleep remains unknown. A considerable number of hypothetical functions of sleep have been proposed including homeostatic restoration, thermoregulation, tissue repair, immune control and memory processing. One of the most intriguing possibilities is that sleep contributes greatly to processes of memory and brain plasticity (Diekelmann & Born, 2010; Rasch & Born, 2007; Rauchs, Desgranges, Foret, & Eustache, 2005; Walker & Stickgold, 2006). The term “memory,” like “sleep,” is not a single entity. Human memory is sorted into different classifications.. 2. Categories of Memory. 立. 政 治 大. In recent years, a number of memory researchers have attempted to divide. ‧ 國. 學. learning and memory into multiple memory systems. These include the distinctions. ‧. between implicit and explicit, episodic and semantic memory, procedural and. sit. y. Nat. declarative memory, or data-driven and conceptually-driven processing. One of the. io. er. most accepted categorizations was to categorize memory into declarative and non-declarative memory. Declarative memory, also called explicit long-term memory,. al. n. iv n C is that in which a person can call tohmind, while non-declarative memory, also called engchi U implicit long-term memory, is that which is normally used without conscious. recollection (see figure 2). Declarative memory includes consciously accessible memories of fact-based information (i.e. knowing ‘what’), and contains several subcategories, including episodic memory that is memory for events in one’s past and semantic memory which is memory for general knowledge. By contrast, non-declarative memory includes all non-conscious memories. It can be further divided into several subcategories, such as classical conditioning, non-associative learning (i.e. habituation and sensitization), implicit memory and procedural memory (i.e. knowing ‘how’) (Squire, 1986; Squire & Zola, 1996; Tulving, 1972; Zola, 1998). 3.
(11) Neuroanatomically, declarative memory involves the hippocampus, along with adjacent entorhinal, parahippocampal, and perirhinal cortices (Eichenbaum, 2000; Squire & Zola, 1996), which are thought to form a temporally ordered retrieval code for neocortically stored information. The task used to be involved in declarative memory is word paired-associates learning. Mary Whiton Calkins, who was president of the American Psychological Association in 1905, invented this experiment procedure for studying memory. The paired associates method requires subjects to learn pairs of items by forming associations between them. There are two forms of paired associates tasks: one is “related” word pairs such as “doctor–nurse” which is. 治 政 semantically or associatively related (Backhaus & Junghanns, 大 2006; Gais et al., 2007; 立 Plihal & Born, 1997; Tucker et al., 2006). The other form of tasks are “unrelated” ‧ 國. 學. word pairs such as “doctor–leaf” which are self evident (Barrett & Ekstrand, 1972;. ‧. Ekstrand, Sullivan, Parker, & West, 1971; Ficca, Lombardo, Rossi, & Salzarulo, 2000;. sit. y. Nat. Gais, Molle, Helms, & Born, 2002; Grosvenor & Lack, 1984; Mazzoni et al., 1999;. io. er. Schmidt et al., 2006; Stickgold, Scott, Rittenhouse, & Hobson, 1999; Yaroush, Sullivan, & Ekstrand, 1971). Learning unrelated word pairs requires the forming of. al. n. iv n C completely novel associations, which hippocampus-dependent. Learning related h eis n gchi U word pairs, on the other hand, strengthens well-formed associations and is hippocampus-independent (Stickgold, 2004).. In addition to word pair learning, there are various tasks that have been used to explore the relationship between sleep and declarative memory including (1) word list learning (Tamminen, Payne, Stickgold, Wamsley, & Gaskell, 2010), (2) face-name association (Backhaus & Junghanns, 2006; Clemens, Fabo, & Halasz, 2005), (3) foreign language or vocabulary learning (De Koninck, Lorrain, Christ, Proulx, & Coulombe, 1989; Gais, Lucas, & Born, 2006; Meienberg, 1977), (4) story retention (Tilley & Empson, 1978), (5) visual-spatial learning (Rasch, Buchel, Gais, & Born, 4.
(12) 2007; Talamini, Nieuwenhuis, Takashima, & Jensen, 2008), (6) virtual maze- learning (Wamsley, Tucker, Payne, Benavides, & Stickgold, 2010), (7) semantic priming (Brualla, Romero, Serrano, & Valdizan, 1998; Plihal & Born, 1999) and (8) rule learning, in which participants learn a set of rules that they then apply to recombine sets of letters into logical sequences (Smith & Weeden, 1990). On the other hand, non-declarative memory is composed of many different cognitive forms. These various processes are related to distinct neuroanatomical structures, involving both cortical and subcortical networks (Doyon, Penhune, & Ungerleider, 2003). In the sleep-dependent non-declarative memory consolidation,. 治 政 three categories of learning were administrated in past 大 studies: (1) motor learning 立 (Fischer, Hallschmid, Elsner, & Born, 2002), (2) visual-perceptual learning (Gais, ‧ 國. 學. Plihal, Wagner, & Born, 2000; Karni, Tanne, Rubenstein, Askenasy, & Sagi, 1994),. ‧. and (3) auditory learning (Atienza, Cantero, & Dominguez-Marin, 2002; Atienza,. sit. y. Nat. Cantero, & Stickgold, 2004). The tasks involved the components of motor skills. io. er. broadly classified into two forms – “motor adaptation” (e.g. finger adaptation task) and “motor sequence” (e.g. sequential finger tapping task) (Doyon, et al., 2003).. al. n. iv n C Motor adaption tests the capacity tohcompensate for environmental changes, whereas engchi U motor sequence measures the incremental acquisition of movements into a. well-executed behavior. In daily life, learning how to use a computer mouse involves motor adaptation, whereas learning how to play a musical instrument like the piano is related to motor sequence. The finger adaptation task, for example, requires subjects to move a cursor with a computer mouse to reach a randomly moving target on a screen. Moreover, pursuit rotor task is also used in measuring motor adaptation. The pursuit rotor test includes a turntable with a metal dot on the platter. Subjects have to keep a flexible metal wand on the dot with motor coordination. The other task associated with motor function is the mirror-tracing task (Born, 1997; Tucker, nap 5.
(13) study). Additionally, a widely accepted mechanism of memory formation is brain plasticity. Brain plasticity refers to the ability of the brain to persistently change its structure and function according to the acquisition of new information. According to Hebb’s theory (1949), memory consolidation is functional changes at the synapses which increase the efficacy of synaptic connection and occurs when pre-synaptic neurons repeat and persistently take part in exciting the post-synaptic neurons. Hence, strengthening connections between neurons leads to synaptic plasticity to enhance memory. As a process, memory refers to the dynamic mechanisms associated with. 治 政 retaining and retrieving information as to past experience. 大 Memory processing can be 立 categorized into three stages: acquisition, consolidation, and retrieval. Acquisition is ‧ 國. 學. the time when the individual requires the information to be remembered, or. ‧. transforms data into a form of mental representation. Consolidation is the process by. sit. y. Nat. which recent memory is stabilized and enhanced into long-term memory. Retrieval is. io. er. the process of recalling a memory.. n. al. ni Ch 3. Sleep and Non-Declarative Memory U engchi. v. The idea that sleep periods are related to learning and memory is experimentally. testable. Overall, three main phases of memory processing may be manipulated during testing: exposure to the new stimulus (acquisition), processing of memory traces (consolidation), and performance at retest (retrieval). The conventional view is that sleep processes participate in the consolidation of the memory traces. Generally, there have been four main paradigms to study the relationship between sleep and memory in humans (Rauchs, et al., 2005; Smith, 2001). Literature is reviewed according to the methodological approaches and distinct memory systems.. 6.
(14) 3.1 Effects of Total or Selective Sleep Deprivation on Memory First approach is to deprive total sleep, or a specific stage of sleep, after learning, in order to induce possible memory impairment. The performance of the task is then examined again at a later time after the test and associated with the type of sleep loss. The rationale for this method is that memory will be impaired after deprivation of a certain sleep stage if that particular stage is critical to the memory process. There are numerous studies attesting to the effects of total or selective sleep deprivation on memory for non-declarative materials. In research by Smith and MacNeill (1994), subjects learned the pursuit rotor task and were then randomly. 治 政 assigned to one of several sleep deprivation conditions:大 (1) total sleep deprivation for 立 the whole night, (2) sleep deprivation for the last half of the night (REM sleep ‧ 國. 學. dominant), (3) selective deprivation of REM only, and (4) non-REM awakenings. ‧. control group. All participants were trained and tested on the task and were retested. sit. y. Nat. one week later. The results showed that subjects in the total sleep deprivation group. io. er. performed significantly worse than did subjects in the controlled non-REM awakening condition and REM deprivation groups. By contrast, subjects who were. al. n. iv n C sleep-deprived during the second half performed significantly worse than h eofnthegnight chi U the controlled groups, while the participants of the sole REM deprivation group. showed a slight, but not significant deficit. Considering that the second half of the night is mainly composed of stage 2 sleep and REM sleep, the authors conclude that stage 2 sleep, rather than REM sleep is the important and crucial determinant of successful motor memory enhancement (Smith & MacNeill, 1994). Walker and his colleagues provided similar results, whereby they let subjects learn a sequential finger tapping task which involves motor sequence learning. The participants who had a whole night’s sleep experienced an increase in motor speed without loss of accuracy, while participants undergoing an equivalent period of 7.
(15) wakefulness showed no significant benefit. In addition, the overnight improvement in performance was correlated to the amount of stage 2 sleep, particularly late in the night (Walker, Brakefield, Morgan, Hobson, & Stickgold, 2002). However, in the two studies mentioned above by Plihal and Born (1997, 1999), evidence was given in favor of a beneficial role of REM sleep on non-declarative memory. They found that the recall of mirror-tracing skills or a wordstem priming task improved more after REM sleep than after SWS (Plihal & Born, 1997, 1999). These results suggest that REM sleep may also be involved in non-declarative memory. Evidence suggests the involvement of both SWS and REM sleep from Stickgold et. 治 政 al (2000a) which demonstrates that participants in a visual 大 texture discrimination task 立 show improvement only after post training sleep, and is positively correlated with ‧ 國. 學. early-night SWS and late-night REM sleep. Compared to a sleep deprivation study. ‧. conducted by Gais et al. (2000), selective deprivation of either early sleep or. sit. y. Nat. late-night sleep impairs overnight consolidation. This evidence may suggest that. io. er. consolidation appears to require not only training, but subsequent events occurring during SWS early on in the following night and REM later that same night.. al. n. iv n C In another interesting study, Kuriyam his colleagues (2004) explored the h e nand gchi U. motor sequence performance profile after a night of sleep, hypothesizing that sleep selectively enhances the weak performance at first learning. Differences in transition speed between each of the separate key-press movements in motor sequence tasks, before and after sleep, were analyzed. When individual subjects’ transition-speed profiles were analyzed before sleep, the speed of individual pre-sleep transition within the sequence demonstrated considerable variability, with certain transitions being particularly problematic (slow), whereas others appeared to be relatively easy (fast). Surprisingly, after a night of sleep, the problematic slow transitions were selectively improved, whereas transitions that had already been effectively mastered prior to 8.
(16) sleep did not change. There was no such improvement in the profile of either slow or fast transitions after waking intervals. These findings suggest that the sleep-dependent learning process selectively provides maximum benefit to motor-skill procedures that prove to be most difficult prior to sleep. Furthermore, it implies that the role of sleep in procedural materials is more subtle and complex than to simply increase the strength of existing non-declarative memory. Moving from the motor learning to visual perception learning, Karni et al. (1994) demonstrated that selective disruption of REM sleep but not NREM sleep results in a loss of performance in visual texture discrimination. These results indicate that a. 治 政 process of human memory consolidation, active during大 sleep, is strongly dependent 立 on REM sleep. Moreover, using the same task, Gais et al. (2000) compared the effects ‧ 國. 學. of early and late sleep. It was shown that performance of a basic visual discrimination. ‧. task significantly improved over early sleep, further improved over a normal night’s. sit. y. Nat. sleep, but did not improve after late sleep alone. Additionally, Stickgold et al. (2000b). io. er. also shows that there is no improvement in performance during a day without any sleep episode. Deprivation of the first night’s sleep after the training also strongly. al. n. iv n C impaired acquisition even after twohnights of recoveryUsleep. engchi. Taken together, these reports build a convincing argument in support of. sleep-dependent learning across several forms of procedural learning. All these studies indicate that a night of sleep triggers delayed learning, without the need for further training. In addition, different sleep stages may be involved in producing this form of overnight consolidation.. 3.2 Changes in Recorded Sleep Parameters After Learning A second method was to train subjects on tasks and then record the changes in sleep parameters after training. In this paradigm, the effect of learning could reveal 9.
(17) changes in sleep architecture if specific sleep stages are important for certain memory formation. That is, existence of sleep-dependent consolidation will lead to sleep modification on the following night. Following an acquisition of procedural tasks or tasks with a procedural component, the changes in sleep parameters were recorded in order to realize the specific sleep modifications on non-declarative memory processing. When examining post-learning modifications in sleep, the increases in the duration of REM sleep or increases in the number of REM sleep have been observed after acquisition of implicit visuo-motor adaptation (Mirror Trace Tasks) and. 治 政 cognition skills (Tower of Hanoi) in the later part of the 大night. Furthermore, an 立 increase in REM densities after training was significantly correlated with the amount ‧ 國. 學. of improvement on the tasks at retest one week later (Smith, Nixon, & Nader, 2004).. ‧. This result is consistent to sleep deprivation studies (Plihal & Born, 1997), in which. sit. y. Nat. motor tasks are associated with REM sleep in the last half of the night.. io. er. However, there is a study that demonstrates the number and density of stage 2 sleep spindles increasing obviously in subjects following acquisition of simple motor. al. n. iv n C tasks (Fogel & Smith, 2006). Spindles been proposed to occur in synaptic h ehave ngchi U. plasticity to consolidate human memory (Sejnowski & Destexhe, 2000). Recently, Peter et al. (2007) used pursuit rotor tasks to examine the effect of initial skill level in the relationship between sleep architecture and procedural memory consolidation. The results demonstrate an increase in stage 2 spindles in the high-skill subjects, and there is a significant correlation between spindle density that occurred after learning and pursuit rotor performance at retest one week later. In contrast, in the low-skill subjects, the variable associated with the performance during retest one week later was a change in REM density. The authors suggest that the changes in sleep modifications that take place after learning depend on the initial skill level. These results imply that 10.
(18) the REM sleep serves the purpose of strengthening weak skills, whereas Stage 2 sleep plays a role of enhancing the well learned skills. In the past, REM sleep has been hypothesized to be involved with the reprocessing of procedural tasks that require a new cognitive strategy (e.g. mirror trace tasks), while simple motor tasks (e.g. pursuit rotor tasks) involve Stage 2 sleep (Smith, 1995, 2001). This notion is supported by a sleep deprivation study (Aubrey, Smith, Tweed, & Nader, 1999), which shows that REM sleep deprivation impairs performance on the mirror tracing tasks, but not on the simple tracing tasks. Conversely, Stage 2 sleep disruption impairs performance on the simple tracing tasks. 治 政 but not the mirror tracing tasks. Therefore, the discrepancies 大 of sleep effects may 立 result from different components of procedural memory and/or the initial level of. sit. y. Nat. 3.3 Reactivation for Memory Processing During Sleep. ‧. ‧ 國. 學. learning.. io. er. A third way is to measure brain reactivation with EEG variety or other brain imaging techniques during post-training sleep. Common techniques for studying this. al. n. iv n C issue include event-related potentials (ERPs), positronUemission tomography (PET), he ngchi and functional magnetic resonance imaging (fMRI). This paradigm supposes that. physiological activities that correlate with effects of learning may be obtained during sleep. Memory formation depends on neuron plasticity, which lasts in structural and functional changes in neurons in response to a stimulus. If sleep is to be considered as a critical mediator of memory enhancement, then evidence of brain activities during sleep may reflect the off-line processing. In recent years, following advancements in techniques, the investigation has benefited from functional neuroimaging. Maquet et al. (2000) have conducted a PET study using a serial reaction time motor task. When retested the next morning, the 11.
(19) subjects’ performance had improved significantly relative to the evening training sessions. Furthermore, several brain areas that had been activated during training were significantly more active during REM sleep in subjects who had previously trained on the task than in untrained ones. It may interestingly suggest that procedural memory may be enhanced during REM sleep, and such sleep-dependent neuronal replay may allow for adaptation of synaptic strengths within specific networks, strengthening some synaptic connections while weakening others. Recent evidence through neuroimaging studies also confirms the beneficial role of REM sleep in consolidating procedural-based skill information (Maquet, 2001; Peigneux, Laureys, Delbeuck, & Maquet, 2001).. 立. 政 治 大. Maquet et al. (2003) have demonstrated sleep-dependent plasticity using a. ‧ 國. 學. procedural visuomotor pursuit task in combination with functional MRI (fMRI).. ‧. Subjects were trained on the task and subsequently retested 3 days later. In sleep. sit. y. Nat. group as compared with sleep-deprivation group, subjects' performance was improved,. io. er. and their brain activity was greater in the superior temporal sulcus.. n. al. ni Ch 3.4 Facilitation of Memory With Cues During Sleep U engchi. v. The last means was to give the subjects some cues during various sleep stages in. the night. These cues are related to the learning task and sometimes resulted in memory enhancement, which implies that presenting such a cue during sleep may positively influence the formation of memory. Whereas the above discussion demonstrates the presence of memory reactivation during sleep, their functional role for memory formation can only be derived from researches of memory retrieval after direct manipulation of such reactivations during prior sleep. So far, no human studies have been made to block memory reactivations during sleep. However, a promising alternative approach taken in some studies is to 12.
(20) induce memory reactivations by presenting such a cue. It implies the cue related to the learning task sometimes plays the role of a reminder, thereby enhancing memory formation during sleep. The method of giving an auditory cue during sleep is less utilized for humans because these auditory cues may affect the sleep cycle (Velluti, 1997) To my knowledge, there is only one study conducted to explore the relationship between sleep and procedural memory via this method. Su (2010) attempted to enhance participant’s procedural memory by auditory cues given during sleep. Subjects of two experiment groups learned a finger sequence tapping task with a. 治 政 presentation of clicking cues, and in the following night, 大the auditory cues were 立 presented to them during either SWS or REM sleep. The other control group learned ‧ 國. 學. the task without clicking cues during learning phase, but they received the same. ‧. auditory cues during REM. The results demonstrate that the subjects re-exposed to the. sit. y. Nat. auditory cues during subsequent REM distinctly improved later performance of. io. REM might consolidate procedural memory.. n. al. Ch. engchi. er. procedural tasks. These findings suggest that reactivation of task-related cues during. i Un. v. 3.5 Interim Summary Previous studies in the association of sleep and non-declarative memory indicate that sleep plays a critical role in posttraining memory consolidation. These results build a cohesive argument that sleep confers a robust learning benefit of non-declarative memory from the behavior studies. Moreover, from the electrophysiological studies, considering the strong evidence for post learning changes in spindles during non-REM stage 2 sleep and REM sleep, this sleep stage probably acts per se or cooperatively with SWS sleep to enhance consolidation. One possibility to explain this phenomenon is that learning a task does not lead to isolation 13.
(21) activation of a single memory network because memory is a dynamic process based on the interaction between the different systems. In summary, that non-declarative memory is sleep-dependent is relatively consistent in contrast to declarative memory.. 4. Sleep and Declarative Memory 4.1 Studies of Total or Selective Sleep Deprivation The classic study by Jenkins and Dallenbach (1924) demonstrates that subjects that had slept after learning recalled more than those who had remained awake. Since this pioneering report, a substantial number of studies have been carried out to explore. 治 政 the possibility that sleep is related to the cognitive processes 大 of learning and memory. 立 For example, Benson and Feinberg (1977) used unrelated paired-associates as the ‧ 國. 學. learning materials and found that participants who had a night of sleep after learning. ‧. showed superior memory to participants who stayed awake for an equal amount of. sit. y. Nat. time. There were no sleep parameters recorded in this study, so it is difficult to know. io. er. which state of sleep was most influential to memory consolidation. Ekstrand’s group (1972; 1971) introduced a protocol for comparing the effects of. al. n. iv n C specific stages of sleep on declarative They compared the performance of h ememory. ngchi U unrelated paired-associates learning following a sleep episode in the early part of. nocturnal sleep and a sleep episode placed in the latter part. It is well known that SWS dominates the first half of sleep while REM sleep is predominant in the second half of the night. They found that subjects slept in first half of night performed superiorly to those slept in second half of night; therefore, the authors concluded that while stage 4 sleep facilitates memory consolidation, REM sleep did not. However, there is a study from the same group showed opposite results. The investigator showed that the paired-associates task was not impaired by REM deprivation, and performance was unaffected by stage 4 sleep deprivation as well (Ekstrand, et al., 1971). 14.
(22) Nonetheless, Fowler et al. (1973) conducted a very similar study, in which investigators trained subjects to learn unrelated paired-associates tasks, then allowed them to sleep either the first half of night sleep or the second half of night sleep. The results demonstrate that memory over an interval with relatively high amounts of REM sleep is inferior to memory over an interval with relatively high amounts of stage 4 sleep. Consequently, the authors temporarily concluded that REM sleep does not facilitate memory consolidation and that SWS may be beneficial to memory. Furthermore, Castaldo et al. (1974) also reported no learning deficits of selective REM sleep deprivation following acquisition of unrelated paired-associates tasks.. 治 政 Similar results were obtained by other investigators. 大For example, Plihal and Born 立 used the same technique to study the effects of sleep on both declarative and ‧ 國. 學. non-declarative memory. They found that the recall of related paired-associates word. ‧. lists (Plihal & Born, 1997) and the performance of mental spatial rotation tasks with. sit. y. Nat. declarative properties (Plihal & Born, 1999) are facilitated by early sleep, which. io. er. implies that SWS promotes consolidation of declarative memory. On the other hand, procedural or non-declarative memory is improved by late sleep dominated by REM. al. n. iv n C sleep. The following study by Gaish and Born also shows e n g c h i U improvement in the related. paired-associates tasks, after early night sleep, is rich in SWS (Gais & Born, 2004). In this study, the authors show that an administration of the cholinesterase inhibitor physostigmine during SWS inhibits the consolidation of paired-associates learning but has no effect on motor learning. Therefore, Gais and Born suggeste that consolidation of declarative memory may require low acetylcholine level during SWS. The results of the previously mentioned studies imply that both related and unrelated word pairs are consolidated during SWS (Benson & Feinberg, 1977; Gais & Born, 2004; Plihal & Born, 1997, 1999). However, in one study, SWS on memory consolidation shows short-term but not long term effect. Grosvenor and Lack (1984) 15.
(23) manipulated sleep-wake states before and after unrelated paired-associates learning. There were four treatment groups in this experiment. Each group represented one of four possible combinations of two different conditions (sleep or awake) prior to learning and post-learning. They were designed as the Awake-Learning-Sleep (ALS), the Awake-Learning-Awake (ALA), the Sleep-Learning-Sleep (SLS), and the Sleep-Learning-Awake (SLA). The first two groups stayed awake during first half of night, and after learning, either went to sleep (ALS) or kept awake (ALA); another two group were allowed to have first half of sleep, and after learning, then they either slept again for the second half of night (SLS) or kept awake (SLA).All groups learned. 治 政 equally well during learning phase. The recall test was大 measured 4 hours and 6 days 立 later, respectively, after learning. At 4h recall test, the data showed that prior awake ‧ 國. 學. groups recalled better than prior sleep groups. There was also a significant effect of. ‧. post learning condition in which post sleep groups recalled better than post awake. sit. y. Nat. groups 4 hours later. However, the retest 6 days later showed no benefit of. io. er. post-training sleep on the task. The authors concluded that sleep following learning benefited recall performance only over short interval (4h), but not in long-term (6. al. n. iv n C days) recall or recognition. It seemshthat sleep has a shot-term effect on memory engchi U consolidation, but this facilitation is not seen in a long-term period.. Although the researches reviewed as above mentioned showed that SWS effect on memory consolidation, there are some evidences from previous studies showed that REM sleep may also involved in memory processing. For example, Stickgold et al. (1999) explored the influence of distinct sleep stages of different strengths of associative links between pairs using a lexical decision task. The task featured lists of prime-target pairs comprising unrelated, weakly related, strongly associated pairs, as well as word/non-word pairs, in which subjects were required to decide, as quickly as possible, if the second word of the pair was a word or a non-word. Priming effects 16.
(24) were used to quantify the relatedness between words, which reflected by shorter reaction times for target items that were preceded by a semantically linked prime, than for weakly related ones. Subjects waking up from NREM sleep (stage 2) displayed priming effects in strongly related pairs whereas subjects waking up from REM sleep presented this effect in weakly related pairs. These data suggest that REM sleep is associated and increases strength of weak associations in cognitive networks, whereas the related association is stabilized during NREM sleep. This notion is supported by the following study. Walker et al. (2002) used similar protocol as participants undergoing tests of anagram word puzzles (e.g. “OSEOG”=”GOOSE”) following. 治 政 REM and NREM wakings throughout the night, and waking 大 performances during the 立 day. The results show accuracy of waking from REM sleep is greater than that ‧ 國. 學. following waking from stage 2 NREM sleep. These findings suggest that. ‧. neurophysiological states of REM sleep are possible to activate weak associations. sit. y. Nat. compared to NREM sleep. Other studies using different tasks involved in declarative. io. er. memory also demonstrate the facilitating role of REM. The recall of words which are grammatically correct but composed of meaningless sentences and prose passages is. al. n. iv n C significantly impaired by selective h REM sleep deprivation e n g c h i U (Empson & Clarke, 1970).. Furthermore, Tilley and Empson (1978) found that the recall accuracy of short stories is significantly poorer when followed by REM deprivation than when followed by stage-4 sleep deprivation. These results seem to suggest that declarative memory is sensitive to REM sleep deprivation. But there are two studies that provide inconsistent results. Lewin and Glaubman (1975) found no negative effect of overnight REM deprivation on memory for serial verbal word lists or word clusters. Chernik (1972) also failed to find any significant impairment of adjective pair retention after REM sleep deprivation. These results suggest the role of REM sleep remains inconclusive. Recently, some investigators proposed that the role of the NREM-REM 17.
(25) alternation, rather than just losing either NREM or REM sleep per se, is critical for recall of verbal material (Ficca & Salzarulo, 2004). There are two studies which conclude that the NREM and REM sleep cycle length is related to declarative memory processing. In the first study, elderly subjects learned unrelated paired words. They were then administrated a test for cued recall as they slept. The results demonstrate that the level of recall positively correlates with the average duration of NREM-REM cycles (Mazzoni, et al., 1999). In the second experiment, the recall of unrelated pairs was impaired after fragmented sleep of disturbed sleep cycle activity, but not after fragmented sleep without sleep cycle disorganization (Ficca, et al., 2000).. 立. 政 治 大. From the literature reviewed, it indicates that the relationship between sleep stages. ‧ 國. 學. and declarative memory is obviously complex. Even in nap studies, the role of. ‧. napping sleep on declarative memory also remains contradictory. For example,. sit. y. Nat. Backhaus and Junghanns (2006) propose that naps do not improve the performance of. io. er. either related paired-associates or face-name association learning, whereas Tucker et al. (2006) provides an opposite finding that naps enhance the performance of related. n. al. paired-associates learning.. Ch. engchi. i Un. v. Hence, all results exploring the effect of sleep on declarative memory remain elusive. Some studies report a positive effect of sleep on memory formation, whereas others report negative results. For the positive role of sleep, the kind of sleep involved remains a question. In summary, consolidation in the declarative memory system may depend on SWS, REM or on a combination of SWS and REM sleep.. 4.2 Post-learning Modifications in Sleep Architecture Examining the changes in sleep physiology after training, there are some studies that demonstrate a significantly increased REM density following an intensive 18.
(26) learning period (De Koninck, et al., 1989; Smith & Lapp, 1991). These results support the idea of REM sleep being involved in long-term memory processing. However, this finding is not sustained by one investigation which both serially presents trigrams and unrelated paired associates used as declarative memory tasks. The results of which demonstrate that no post-acquisition changes in neither REM nor NREM sleep parameters (Castaldo, et al., 1974). Meienberg (1977) also proposes that no changes have been shown in the tonic component of REM sleep or in any other sleep stage following learning of a verbal memory task. On the other hand, impairment in REM sleep has apparent deficits in memory, if REM sleep is crucial for memory. Some. 治 政 authors, who believe that sleep has no effect, propose that 大patients with diminished 立 REM sleep continue to maintain productive and unaffected lives (Vertes, 2004; Vertes ‧ 國. 學. & Eastman, 2000; Vertes & Siegel, 2005). Therefore, the effects of REM sleep on. ‧. declarative memory remain unclear.. sit. y. Nat. Some studies indicate that sleep spindle density is higher after learning (Clemens,. io. er. et al., 2005; Gais, et al., 2002; Schmidt, et al., 2006), which suggests that spindles represent a reorganization mechanism of consolidating declarative memories. A. al. n. iv n C reported increase in spindle densityhearly in the night U e n g c h i following unrelated. paired-associates learning (Gais, et al., 2002) and increased spindle density after learning difficult lists of words (Schmidt, et al., 2006), demonstrates a positive role of sleep spindles in declarative memory processing. Similarly, it reveals a strong correlation between the number of sleep spindles and overnight memory performance (Clemens, et al., 2005). Therefore, spindle activity during non-REM sleep appears to be particularly sensitive to learning consolidation and, thus, successful recall of information associated with declarative memory. Through the studies using this method comparing the change of sleep structure, it shows that either REM (De Koninck, et al., 1989; Smith & Lapp, 1991) or Stage 2 19.
(27) sleep (Clemens, et al., 2005; Gais, et al., 2002; Schmidt, et al., 2006) reveal an enhancement effect on declarative memory. Taking all the results using sleep deprivation into consideration, SWS sleep also demonstrates a positive role on consolidation of declarative memory (Plihal & Born, 1997, 1999). In addition, some investigators emphasize that NREM-REM sleep cycles rather than sleep stage per se are important (Ficca, et al., 2000). Therefore, there are still no evident conclusions about which sleep stage is important to declarative memory.. 4.3 Reactivation for Memory Processing During Sleep. 治 政 A recent PET imaging study investigated how consolidation 大 affects the neural 立 correlates of memory retrieval over short and long retention periods. Their results ‧ 國. 學. indicate that the pattern of brain activation is dramatically different at the different. ‧. retention intervals. Peigneux et al. (2004) report a neuron-imaging study using a. sit. y. Nat. virtual route-learning task, a kind of declarative spatial memory task. They discovered. io. er. that hippocampal regions that were activated while doing route retrieval prior to sleep were re-activated during subsequently SWS but not REM sleep. In addition, the. al. n. iv n C hippocampal activity expressed during was positively correlated with overnight h eSWS ngchi U performance improvement in route retrieval on the following day. These results. support the notion that enhanced hippocampal activity during post training SWS reflects the off-line processing of memory trace, which possibly led to the plastic changes underlying the overnight improvement. In the same vein, using functional MRI, Gais et al. (2007) discovered that sleep leads to a redistribution of memory traces from the hippocampus to neocortical sites for long-term storage. Subjects participated in both a sleep and sleep-deprivation condition, they then underwent (1) immediate recall test, (2) second recall test after 48 hours, or (3) third recall test after 6 months without announcement. The results show 20.
(28) that the hippocampal involvement in correct recall increased significantly only when subjects slept, but not after sleep deprivation. Activities of hippocampus during post-learning sleep may process initial consolidation of memory. However, the results of the recall test after six months demonstrate that memories activated the medial prefrontal cortex more significantly. This evidence suggests that the neural circuits supporting declarative memory undergo synaptic reorganization through sleep.. 4.4 Influencing Memory Processing During Sleep by Cuing Smith and Weeden (1990) report that when subjects are trained on a complex. 治 政 rule-learning task in the presence of a ticking alarm clock, 大 the re-exposure of the 立 clicks at time of rapid-eye movement during subsequent sleep enhanced later ‧ 國. 學. performance on the task. However, the auditory cues presented during sleep may. ‧. interfere with sleep structures (Velluti, 1997). In one study, subjects learned. sit. y. Nat. object-location associations while hearing object sounds (e.g. cat with sound of. io. er. “meow”). They had a 75 min nap after learning. During the entire period of the nap, unobtrusive white noise was given, whereas the sounds for half objects were. al. n. iv n C presented during NREM sleep. Individuals required to place each object in it’s h e n gwere chi U. correct location. The results show the accuracy of these locations for cues was better than that of other locations without reminder cues. The author suggests that reminder during sleep can reactivate and strengthen human memories (Rudoy, Voss, Westerberg, & Paller, 2009). In order to minimize the effect of sleep interference by the sound of clicks, Born. and his colleagues conducted (2007) a study using an olfactory stimulus to reactivate declarative memory. Participants learned a visual-spatial learning task under the presence of an odor. Re-exposure to the odor during subsequent SWS distinctly improved later retrieval of the task stimuli. Controlled experiments specify that the 21.
(29) memory enhancement after odor-exposure during sleep depends crucially on the presence of the odor prior to learning. Furthermore, odor re-exposure was effective only during SWS, but not during REM sleep or wakefulness. In an additional experiment using fMRI, hippocampal activity during reactivation in SWS was significantly stronger compared to odor re-exposure during wakefulness after learning, indicating an enhanced sensitivity of the hippocampus during SWS to stimuli capable of reactivating declarative memory. Together, these findings provide support for the notion that reactivations occurring spontaneously during sleep might serve in the same way to consolidate new memories.. 立. 4.5 Interim Summary. 政 治 大. ‧ 國. 學. Taken as a whole, these studies regarding declarative memory consolidation. ‧. present a degree of mixed results. The effect of sleep on unrelated-pairs learning. sit. y. Nat. showed inconsistent results (see table 1), whereas the role of sleep on related-pairs. io. er. learning demonstrated consistently facilitating effect (see table 2). As to specific sleep stage on memory, most of the results are in favor of the role of SWS in memory. al. n. iv n C consolidation, but still some studieshare in favor of REM e n g c h i U sleep. This discrepancy can be explained by the material used and the processes involved while performing the task. For example, learning related or unrelated word pairs may occur either in distinct brain regions or in different processes of memory consolidation. Moreover, because both sleep and memory are involved in distinct processes and states, consequently it is not easy to discover the clear one-to-one relationship. Each sleep stage processes a set of neuro-physiological and neuro-chemical mechanisms that may contribute uniquely to distinct tasks involving different aspects of memory formation. However, from the literature reviewed, although there are discrepant results, overall sleep has an effect on declarative memory. 22.
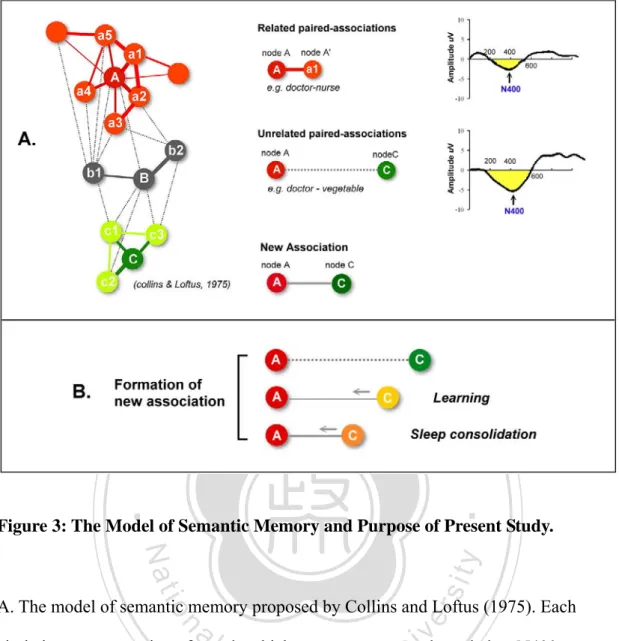
(30) 5. Network of Human Memory System 5.1 Representation of Memory Network Collins and Loftus (1975) propose that the structure of semantic memory is an associative network (Figure 3A). The association represents a relationship between two lines of information, and associations at different levels of mental representation are essential to models of declarative memory. Specific concepts, words, or perceptual features in the declarative memory system are represented as nodes, which are points located in space of semantic memory. Furthermore, concept nodes are bridged. 治 政 together by links. In such a structure, concepts are linked 大in a semantic network. The 立 length and strength of links represents the association between nodes. Therefore, ‧ 國. 學. related concepts are linked closely together in psychological and neuronal space, and. ‧. vice versa. Concepts closer together in the hierarchy require less time for retrieval and. sit. y. Nat. are more central to the meaning of the concepts. This is the semantic relatedness. io. er. effect describing the distance between concepts in memory systems. If a semantic concept is activated by memory access, related nodes should be retrieved easily. al. n. iv n C because the links between nodes arehstronger. Hence, U e n g c h i its reaction time and recognition will be facilitated (Ashcraft, 1994; Neely, 1991).. 5.2 Priming Paradigm to Measure the Associations Between Words Priming is an important technique for assessing association between nodes in semantic memory with behavioral or electrophysiological measures. The term of priming is used to refer to several different phenomena, including (1) item repetition priming, (2)semantic priming, and (3) new association priming (Wagner & Koutstaal, 2002). Semantic priming refers to a facilitation of responding that occurs as results of the presentation of a semantically related word, as when presentation of the word 23.
(31) “nurse” facilitates access to the word “doctor”. Behavioral indices of semantic priming effects are shorter reaction time (RT) and more accuracy of responses. Neurophysiological priming is expressed by smaller deflection in N400 component of event related potential (ERP) (Bentin, McCarthy, & Wood, 1985; Kounios & Holcomb, 1992). New association priming, also known as associative priming, refers to priming of the relationship or association between unrelated words. Priming for new association is measured by presenting pairs of pre-experimentally unrelated stimuli, such as words, in a study phase (e.g. TABLE-SNAL, GRASS-POLICEMAN) and then. 治 政 presenting three types of pairs during a subsequent test大 phase: intact pairs which 立 words presented in their original pairing (TABLE-SNAL), rearranged pairs ‧ 國. 學. (GRASS-SNAL) and new pairs. Measures of new association priming are based on a. ‧. comparison of performance for intact pairs relative to rearranged pairs. Associative. sit. y. Nat. priming is a process of priming for new associations, information not pre-existing in. io. er. the memory system (Graf & Schacter, 1985; Schacter & Buckner, 1998). To build up novel word associations, subjects require integrating target into prime during learning,. al. n. iv n C by processing co-occurrence and/orhforming relationsU e n g c h i between them. (Goshen-Gottstein & Moscovitch, 1995a, 1995b; McKone & Slee, 1997). If intact pairs show greater facilitation than rearranged pairs, this can be attributed to the retention and use of associative information because the individual terms in both types of pairs were encountered previously- the only difference is their conjunctive or associative paring (Lin & Yang, 2010). In the same vein, item repetition priming refers to instances where the same stimulus repeats after an initial occurrence, which occurs when response to rearranged pairs is better than response to unstudied new pairs. Item repetition priming reflects the processes of perceptual representations that were functionally independent of the semantic or episodic memory system (Schacter 24.
(32) & Buckner, 1998; Tulving & Schacter, 1990).. 6. ERP Studies in Sleep-dependent Memory 6.1 Introduction to N400 of Event-related Potentials (ERPs) ERPs are voltage fluctuations recorded on the scalp. ERPs reflect changes in electric (or magnetic) field potentials, mainly resulting from transmembrance currents in neural dendrites during synaptic activity. In the ERPs techniques, brain waves are recorded in conjunction with specific stimuli, response, and/or cognitive events. ERPs can be analyzed as a function of the precise onset of the events. ERPs provide very. 治 政 good information of time-course of task-related brain activity. 大 立 Although current ERP methods do not provide precise neuroanatomical ‧ 國. 學. localization, the temporal resolution of ERPs allows critical events to be monitored. ‧. over a course of milliseconds. The resulting wave forms show characteristic spikes. sit. y. Nat. related to the timing of electrical activity. ERPs are analyzed in terms of positive- or. io. er. negative-going deflections relative to baseline in lock of time. For example, a negative deflection within the period of 300-500 msec. after the presentation of the. al. n. iv n C stimulus called “N400” of ERPs components, negative-going waveform centered h e n g ca h i U around the 400-msec time interval.. N400 component was first described by Kutas and Hillyard (1980). Kutas and Hillyard report that semantically inappropriate words (e.g. He spread the warm bread with socks) elicited a large N400, relative to the N400 induced by semantically appropriate words (e.g. It was his first day at work). These results showed that the N400 is sensitive to semantic processing, which is enlarged when semantic context is violated. Since then, many studies have been conducted to explore the nature of the N400 component. Therefore, some investigators suggest the N400 effect reflects the process of semantic integration of the critical word with the preceding context 25.
(33) (Friederici, 1997; Kutas & Hillyard, 1980). According to this integration view, N400 amplitude is greater in semantically anomalous context because integration is easier in congruent context. The integration view also explains the finding that less expected endings induce larger N400 amplitude than more expected endings since integration is suggested to be difficult when expectations are not met (Roehm, Bornkessel-Schlesewsky, Rosler, & Schlesewsky, 2007). While the initial discovery of the N400 was during sentence processing, it was soon discovered that sentences are not needed to elicit N400s but that single words are sufficient (Bentin, McCarthy, & Wood, 1985). Some studies using lexical decision or. 治 政 category judgment tasks demonstrate that semantically大 unrelated targets evoke a large 立 N400 than related targets (Bentin, et al., 1985; Chwilla & Kolk, 2005; P.J. Holcomb & ‧ 國. 學. Neville, 1990; Nunez-Pena & Honrubia-Serrano, 2005; Rugg, 1985). Studies showed. ‧. that the morphology, timing, and topography of the N400 semantic priming effect for. sit. y. Nat. target words following semantically related versus unrelated primes is not different. io. er. from those for the N400 effect to sentence final words of congruent versus anomalous sentence (Kutas & Federmeier, 2011). Therefore, semantic relationship effects have. al. n. iv n C been observed in N400s collected in of pair experiments using lexical hword e n list gchi U. decision, semantic categorization, semantic rating, or letter search task. Following discovery of N400, many investigators attempt to explore the variables that affect N400, because knowing these factors is essential to realize what underlying mechanisms reflected by N400.. 6.2 Factors That Affect N400 Amplitude Extensive research has been carried out to understand what kinds of experimental manipulations affect the N400. The variables that may affect the amplitude of N400 include: (1) semantic association: the N400 is sensitive to the relatedness between 26.
(34) words. The amplitude of N400 reduced when a target word is preceded by a word that is semantically, morphologically, or orthographically related to it (Kutas & Federmeier, 2000). (2) lexicality effect: in various studies using different materials, the N400 elicited by pseudoword has been shown to be larger than that elicited by real word (Kutas & Iragui, 1998). (3) word repetition: repeating words in a list or in text elicit smaller amplitude of N400 than the first presentation of the words (Besson, Kutas, & Van Petten, 1992; Rugg, 1985, 1990; Van Petten, Kutas, Kluender, Mitchiner, & Mclsaac, 1991). (4) word frequency: high-frequency words tend to elicit a smaller N400 response than low-frequency words (Rugg, 1990). Since frequency and. 治 政 repetition are the factors that may increase the accessibility 大 of lexicon, and the 立 amplitude of N400 is influenced by semantic association, frequency, and repetition. ‧ 國. 學. N400 is therefore considered to reflect a common mechanism associated with the. Nat. io. er. 6.3 Theories and/or Mechanisms of the N400. sit. y. ‧. interaction of all these three factors (Young & Rugg, 1992).. There were a growing number of the literatures attempted to develop theories. al. n. iv n C and/or mechanisms of the N400. Some pay attention on the neurobiological h eofnthese gchi U. level, seeking to delineate the brain networks responsible for the N400 (Lau, Phillips, & Poeppel, 2008) and linking the component to specific neural functions. Amplitude reductions of N400 might reflect smaller postsynaptic potentials in the same neurons in a population, and less temporal synchrony among the generating neurons. Others focus on the functional level, mapping the N400 onto particular cognitive operations, such as orthographic/phonological analysis (Deacon, Dynowska, Ritter, & Grose-Fifer, 2004) or semantic memory access (Kutas & Federmeier, 2000). However, there is still much debate as to exactly what kind of neural and comprehension processes the N400 indexes. One of debate is whether it reflects an 27.
(35) automatic process (spreading activation) (Deacon, Hewitt, Yang, & Nagata, 2000; Deacon, Uhm, Ritter, Hewitt, & Dynowska, 1999) or a controlled process (semantic integration) (Chwilla, Brown, & Hagoort, 1995; Neely, 1991). The spreading activation model (Collins and Loftus, 1975) proposes a network organization of semantic memory with related semantic concepts represented in neighboring nodes. If a semantic concept is activated by memory access, activation should spread automatically to related nodes through the links and increase their activation level. Hence, if a word denoting a related concept is presented, its recognition and reaction time will be facilitated. Spreading activation should only occur when the stimulus. 治 政 onset asynchrony (SOA) is short because the network activation 大 should decay with 立 time. On the other hand, controlled processes are thought to dominate at longer SOA. ‧ 國. 學. One mechanism of controlled process is based on Rugg and Doyle’s (1994). ‧. integration model: unrelated targets are more difficult to integrate than related targets.. sit. y. Nat. This additional requirement for semantic processing should prolong reaction times. io. er. and evoke a larger N400. Another mechanism of controlled process is based on expectancy effects. If a prime is presented, the subject might expect a target word that. al. n. iv n C is semantically related to the prime.hA related target word e n g c h i U will therefore be easier to process than an unrelated target (Roehm, et al., 2007).. Despite these controversies, modulation of the N400 (in one form or another) is seen in most semantic ERP paradigms. The robust nature of this pattern makes it a good candidate for tracking changes in word semantic knowledge. However, one thing should be especially mentioned that most N400 researches address the semantic associations that pre-exist in the memory network (Canas, 1990; Chwilla & Kolk, 2005; Hill, Ott, & Weisbrod, 2005; Hill, Strube, Roesch-Ely, & Weisbrod, 2002; P. J. Holcomb, 1988; Nunez-Pena & Honrubia-Serrano, 2005; Rhodes & Donaldson, 2008; Roehm, et al., 2007). It is unclear if N400 can reflect newly established association 28.
數據
+7

相關文件
(A) To explain why it takes longer for old people to sleep (B) To explain why people have difficulties in sleeping (C) To explain why people have variations in sleep duration (D)
To explore different e-learning resources and strategies that can be used to successfully develop the language skills of students with special educational needs in the
educational needs (SEN) of students that teachers in the mainstream English classroom need to address and the role of e-learning in helping to address these needs;.. O To
educational needs (SEN) of students that teachers in the mainstream English classroom need to address and the role of e-learning in helping to address these needs;.. O To
Explore different e-learning resources and strategies that can be used to successfully develop the language skills of students with special educational needs in the..
Narasimhan, Ram, and Anand Nair (2005), “The Antecedent Role of Quality, Information Sharing and Supply Chain Proximity on Strategic Alliance Formation and Performance,”
Another point to financial performance indicators used to measure the performance of the industry's performance, to explore market in domestic container shipping industry
Investigating the effect of learning method and motivation on learning performance in a business simulation system context: An experimental study. Four steps to
