* 通信作者:孫智雯(Chih-Wen Sun);FAX:886-2-29312904;E-mail:[email protected]
阿拉伯芥
atToc159 轉運蛋白基因的細胞專一性表現與領先內插
子對基因表現的影響
劉玉山
孫智雯*
國立臺灣師範大學生命科學系(收稿日期:2013.4.12,接受日期:2013.5.9) 摘 要 根據蛋白質胺基酸序列的相似度與運輸蛋白質種類的不同,可將阿拉伯芥四種 Toc159 的同源蛋白質,分成atToc159/atToc90 和 atToc132/atToc120 兩大群。其中 atToc159/atToc90 被認為與辨識和運輸光合作用蛋白質有關,atToc132/atToc120 則可能負責辨識和運輸非光合 作用蛋白。在阿拉伯芥生長發育的過程中,這些轉運蛋白的基因必須被適當的調控,否則 色質體的發育與生理將因缺乏必需的蛋白質的輸入而被抑制。為了解atToc159 基因家族的 表現機制,轉殖分別攜帶不同長度的基因上游調控序列的重組質體進入野生型阿拉伯芥, 以獲得穩定轉殖的植株。分析這些轉殖植株中營養器官的GUS 酵素活性及 mRNA 表現量, 發現atToc159、atToc132/90 和 atToc120 基因在七天齡的幼苗中分別有最高,次之及最低的
表現量。此外,位於atToc120 和 atToc90 的 5′ UTR 內的領先內插子序列會明顯增加其基因
表現量。進一步探討光對內插子在基因表現上的影響,比較生長在光照和黑暗條件下的 120PUI 和 120P 轉殖株的 GUS 表現量,我們發現黑暗會提高內插子效應。除了進行不同組 織的基因表現之比較分析外,我們也進行了這些轉殖株中基因的細胞專一性表現的研究。 令人驚訝的,在與atToc159 和 atToc132 的比較下,atToc90 和 atToc120 在子葉保衛細胞中
基因的表現量明顯地要比葉肉細胞多,這顯示atToc90 和 atToc120 在保衛細胞運輸蛋白質進 入葉綠體上可能扮演一定的角色。這些研究結果支持葉綠體轉運蛋白atToc159 基因家族之 基因的表現量在不同的組織和發育階段的差異,可能是受到不同的發育和環境的因子刺激 所調節。 關鍵詞:阿拉伯芥、atToc159 基因表現、領先內插子、GUS 報導基因、保衛細胞
緒 言
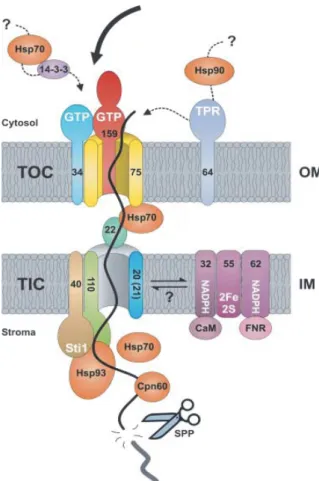
色質體(plastids)是植物細胞內最重要的 胞器之一,主要包括白色體(leucoplasts)、雜 色體(chromoplasts)和葉綠體(chloroplasts)等三 種。這些色質體均是由原色質體(proplastids) 衍生而來,只要給予適當的訊號,他們可以進 行彼此之間的轉換。根據演化的內共生理論 (endosymbiont hypothesis),色質體存在於植物 細 胞 中 起 源 於 遠 古 時 期 , 將 藍 綠 菌 (cyanobacteria) 經 由 胞 吞 作 用 (endocytosis) 移 至植物細胞質中。然而高等植物的演化過程中, 超過百分之九十的藍綠菌基因,卻被轉送到植 物寄主的細胞核內(Martin et al., 2002)。現今 植物在進行色質體的的生物發生(biogenesis) 時,必須將這百分之九十的基因產物準確地送 回色質體中,否則色質體內的脂質生合成(lipid synthesis)、色素合成(pigment synthesis)、 色質體基因表現(殘存在色質體的基因)、膜系 組裝(membrane assembly)及光訊號接收(light perception)皆會受到嚴重的抑制(Rochaix et al., 2004)。 色 質 體 內 大 多 數 的 細 胞 核 解 碼 蛋 白 (nucleus-encoded proteins)要運輸回到色質體 中,需要在細胞核內的基因轉錄及在細胞質內 轉譯並修飾後,再傳送回色質體內。大多數的 色質體內的蛋白質,多含有一段特別的胜肽構 造位於其胺基端延伸(N-terminal extension)部 分,稱之為運輸胜肽(transit peptide)。這些含 有 運 輸 胜 肽 的 色 質 體 前 軀 蛋 白( precursor proteins),會被位於色質體膜上的轉運蛋白機 組(translocons)所辨識,進而將其運輸至色質 體內。前人在豌豆的葉綠體中發現數種轉運蛋 白機組成員(圖一;Jarvis, 2008)。位於葉綠體
圖一、色質體TOC/TIC 轉運蛋白機組成員。
Figure 1. The translocon components of TOC/TIC
complex (modified from Jarvis, 2008).
外膜的轉運蛋白稱之為Toc (translocon at the outer envelope membrane of chloroplasts),位於 葉綠體內膜的則稱為 Tic (translocon at the inner envelope membrane of chloroplasts)。豌豆 Toc 複合體主要由 Toc75 和 Toc159 和 Toc34 三種蛋白質所構成,其中Toc75 是一個擁有 β-筒狀(β-barrel)構造的蛋白質,是前軀蛋白質運 輸 時 主 要 穿 越 外 膜 的 通 道 蛋 白(protein- translocating channel)。Toc34 和 Toc159 屬於 GTP 水解酶(GTPase),主要功能是辨識前軀蛋 白,當作運輸胜肽最初的受器。Toc159 包含 三個不同的蛋白質區域,一個是羧基端嵌於外 膜內的M 區域(membrane anchor domain)、位 於中央部分具有GTP 水解酶功能的 G 區域以 及胺基端的A 區域(acidic domain) (Ivanova et
al., 2004)。另外,Toc34 結構中包含二個區域, 為具有GTP 水解酶功能的 G 區域與羧基端嵌 於外膜上的 M 區域。Toc34 的功能雖未完全 明瞭,但是許多證據顯示其可與前軀蛋白有直 接的交互作用,被認為可能是運輸胜肽的受器, 且與協助Toc159 與 Toc 複合體結合上扮演重 要的角色(Jarvis and Robinson, 2004)。
阿拉伯芥atToc159 具有兩種不同的型式, 一種是細胞游離型(soluble cytoplasmic form) 的atToc159,一種則是嵌於葉綠體外膜上的膜 結合型(membrane-bound form)的 atToc159。可 能游離型的atToc159 以其 G 區域與前軀蛋白 的運輸胜肽結合後,再藉由atToc33 的協助, 將攜帶前軀蛋白的游離型 atToc159 順利地嵌 入葉綠體外套膜上atToc75 通道蛋白旁,形成 膜結合型的atToc159,如此便可不斷將前軀蛋 白 導 入 atToc75 通 道 蛋 白 中 (Wallas et al., 2003)。整個嵌入過程最關鍵的部分,涉及 GTP 的水解活化atToc159 與 atToc33 的 G 區域, 也說明了 Toc 複合體運輸前軀蛋白進入葉綠 體需要GTP 參與 (Smith et al., 2002)。 阿拉伯芥含有四個豌豆psToc159的同源 基因,包括atToc159、atToc132、atToc120和 atToc90。AtToc159基因家族的蛋白質產物構造 上的主要差異在於胺基端的A區域的長度。各 種atToc159基因家族的人造蛋白質(chimeric protein)之試管(in virto)和活體(in vivo)研究結 果,證實A區域可能與選擇運輸何種蛋白質進 入葉綠體有關(Inoue et al., 2010)。另外,阿拉 伯 芥 中 也 含 有 二 個psToc34 的 同 源 基 因 , atToc33及atToc34。越來越多的研究顯示,阿 拉伯芥的atToc159和atToc34基因家族成員的 蛋白質產物可能與不同種類前軀蛋白的辨識 和運輸有關,一般認為atToc159可與atToc33 合作辨識並運輸光合性蛋白質(photosynthetic protein) , 而 atToc120 或 atToc132 則 負 責 與 atToc34 合 作 辨 識 並 運 輸 非 光 合 性 蛋 白 質 (Invanova et al., 2004;Smith et al., 2004)。 AtToc90的功能性研究顯示它似乎與運輸光合 作用蛋白有關,其辨識和運輸的機制可能與 atToc159 相 似 , 但 重 要 性 遠 不 及 atToc159 (Hiltbrunner et al., 2004)。
阿 拉 伯 芥 突 變 株 ppi2 (plastid protein import 2,atToc159 突變株)的幼苗出現嚴重白 化的現象,無法在土壤中存活。且葉綠體的發 育受到抑制,體內的光合作用蛋白含量明顯的 下降,而非光合作用蛋白的含量則與野生型沒 有明顯差異。顯示 atToc159 可能與光合作用 相關蛋白最初的辨識和運輸有關(Bauer et al., 2000)。此外,ppi2 突變株的葉子在發育後期 卻發生嚴重的構造上的缺失,葉肉細胞明顯的 變少,細胞間隙變多,海綿組織與柵狀組織無 法區分,葉綠體內 DNA 複製則在發育早期 即受到嚴重的抑制(Asano et al., 2004)。另一個 關於阿拉伯芥突變株 ppi2 的研究,卻發現突
35 變株 ppi2 植株子葉的保衛細胞的葉綠體內仍 可以看見葉綠素的累積和正常的囊狀膜的堆 疊,並有光合作用產物澱粉的生成,顯示突變 株 ppi2 的保衛細胞內的葉綠體仍具有運輸光 合作用蛋白的能力,亦即在保衛細胞的葉綠體 的生成過程中,atToc159 並非是必要的(Yu and Li, 2001)。另一個以 ppi2 突變株為基因背景下 過度表現atToc90 的實驗中,則發現大量表現 的atToc90 可以部分恢復 ppi2 突變株累積光合 作用蛋白的能力(Sibylle et al., 2011)。與野生 型植株比較,attoc132 與 attoc120 等單突變株 沒有外表型的瑕疵,但是attoc132/attoc120 的 雙突變株(double mutant)卻出現白化的現象, 顯見atToc132 和 atToc120 具有功能上的重疊 性(Ivanova et al., 2004;Kubis et al., 2004)。
植物生活史中不同的發育階段中,不同的 組織含有不同形式的色質體,色質體的發育必 須健全,植物細胞才能擁有的正常的生理功能。 所以這些轉運蛋白的基因必須被適當的調控, 否則色質體的發育與生理將因缺乏必須的蛋 白質的輸入而被抑制,進而影響植物的生長和 發育。因此,探討有關atToc34 和 atToc159 基 因家族的基因在不同的組織或發育階段是如 何被啟動或調節,不同的轉錄因子與基因調節 序列在基因表現上的影響以及mRNA 的穩定 度對基因表現的影響等,將可以進一步說明葉 綠體蛋白質運輸的可能調節機制,並提供色質 體正常發育的重要訊息。 基於以上的理由,我們曾將atToc34 基因 家族(atToc33 和 atToc34)不同上游調控序列連 接GUS 報導基因後,進行穩定轉殖植株分析。 結果顯示atToc33 及 atToc34 主要分別表現在 葉及根中。這兩個基因位於 5 端非轉譯區(5′ untranslated region, 5′ UTR) 的 領 先 內 插 子 (leader intron) 序 列 , 可 以 提 高 成 熟 植 株 的
atToc33 和 atToc34 基因的表現量。因為兩基
因的表現明顯受到不同的機制調節,atToc33 和atToc34 在葉綠體的功能及發育中可能扮演 不同角色(Chen and Sun, 2010)。
本研究將接續以上的實驗,嘗試進行
atToc159 基因家族,包括 atToc159、atToc132、 atToc120 和 atToc90 等基因的不同上游調控序 列,連接GUS 報導基因的穩定轉殖植株研究。 除了找到細胞核中 atToc159 基因家族可能的 調節機制,也希望幫助我們更進一步暸解不同 組織與發育階段色質體蛋白質運輸的可能調 節方式,以及 atToc159 基因家族的蛋白質產 物在色質體功能和發育上所扮演的角色。
材料與方法
阿拉伯芥的栽培 將 消 毒 好 的 野 生 型 阿 拉 伯 芥 (ecotype Columbia; WT)或轉殖植株 T3同型合子的種子, 置於4 oC 冰箱中春化 2 天。再將種子均勻塗 佈於MS 培養基(Murashige and Skoog, 1962) 或種植於土壤中。置於生長箱中,日夜恆溫 22 oC,以 16 小時光照 8 小時黑暗的光照週期 (正常光照週期)或連續黑暗環境下培養。 Toc159 基因家族成員上游的調控序列的取得 阿拉伯芥 Toc159 基因家族(atToc159、 atToc132、atToc120、atToc90)的上游的調控序 列,可在阿拉伯芥基因線上資料庫TAIR (TheArabidopsis Information Resource, Lamesch et al., 2012)中,依據基因編號(gene accession
number)下載。包含該基因啟動子(promoter) 的和5′ UTR 區域。本論文所提及的基因編號 包 括 AT4G02510 (atToc159) 、 AT2G16640 (atToc132) 、 AT3G16620 (atToc120) 和 AT5G20300 (atToc90)。
引子的設計
利用上述方法所取得的基因序列,設計能 複製各基因上游調控序列之專一性引子。其中 前置引子(forward primer)的 5′加上含有限制 酶 Hind III 切點的序列,而反置引子(reverse primer)的 5′則加上含有限制酶Bam HI 切點的 序列。各基因引子之序列如下: Toc159 re-F1 5′ - gccaagcttggatttgtgttatgtttctcgc -3′ Toc159 re-R1 5′ - cggggatccccgctttgctactgagactc -3′ Toc132 re-F1 5′ - gccaagctttagctgcaccagcttattgag -3′ Toc132 re-R1 5′ - cggggatcctctagatcaccaccgctacg -3′ Toc132 re-R2 5′ - cggggatcccagaagttagagatagagagag -3′ Toc120 re-F1 5′ - gccaagcttagtgtggtgttgtttaagtgtg -3′ Toc120 re-R1 5′ - cggggatccctaggatcacccaaaatcacg -3′ Toc120 re-R2 5′ - cggggatccctggttagagaaggcaaaagtc -3′ Toc90 re-F1 5′ - gccaagcttagacgaagatgtcgtcattgg -3′ Toc90 re-R1 5′ - cggggatccaactatctgcccaacagcaag -3′
Toc90 re-R2 5′ - cggggatccttgtgttggcgagagaaagag -3′ 構築轉殖植株載體與植物轉殖 將上述七個 atToc159 基因家族的基因片 段 經 聚 合 酶 連 鎖 反 應(polymerase chain reaction, PCR)放大後,與含有 β-glucuronidase (GUS)基因的 binary vector pCAMBIA1391Z 黏 合 後 , 將 所 得 的 重 組 質 體 , 藉 由 農 桿 菌 GV3101 為媒介,並利用花序沾黏法的方式轉 殖進入野生型的阿拉伯芥植株中(Clough and Bent, 1998)。T1 種子培養於洋菜培養基(含 hygromycin 30 μg/mL)篩選出成功轉殖的植株, 並以上述特定專一性引子進行PCR 確定。這 些轉殖株分別命名為1391Z、159PUI、132PUI、 132P、120PUI、120P、90PUI 和 90P。除 1391Z 為空的pCAMBIA1391Z 質體的穩定轉植株, 當作本研究的控制組外,其餘轉植株則分別攜 帶不同長度的 atToc159 基因家族(atToc159、 atToc132、atToc120、atToc90)的基因上游的調
控序列的重組質體。這些轉殖株經過至少兩代 的 篩 選 , 最 後 獲 得 第 三 代 是 同 型 合 子(T3 homozygote)的個體,作為本研究使用的材 料。 GUS 活性分析 本實驗分別進行 GUS 酵素的螢光定量和 活體組織的GUS 活性定位染色(Jefferson et al., 1987),藉以分析不同轉植株的 GUS 基因表現 程度與位置。在GUS 酵素螢光的定量分析中, 細胞抽出物以螢光光譜儀(SpectraMax Gemini XPS, Molecular Device)偵測並定量;而活體組 織的GUS 活性定位染色,則是分別將植株幼 苗浸泡於GUS 活性染色液中,37 oC 環境下反 應 3 小時、8 小時、18 或 24 小時後,並置於 固定液中(內含 0.1 M sodium phosphate, pH 7.2, 0.1% formaldehyde, 0.1% triton X-100, 0.1% β-mercaptoethanol)保存。 即時定量反轉錄PCR (real-time quantitative reverse-transcription PCR, qRT-PCR)
以 TRIzol 試劑(Invitrogen, Chomczynski et al., 1987),抽取不同處理之阿拉伯芥轉殖株 的total RNA。取 5 g total RNA,利用寡核苷 酸 引 子 (oligo-dT primer) 以 及 反 轉 錄 酶 (M-MLV reverse transcriptase, Promega)反轉錄 成cDNA (complementary DNA)。以這些 cDNA 作為qRT-PCR 的模板,加上各 atToc159 家族 基因之專一性引子(UBQ10 作為內部控制組)
以及2X SYBR FAST qPCR kit 的試劑(KAPA Biosystems)進行實驗,其條件設定為 95 oC 3 分鐘,而後95 oC 3 秒、60 oC 30 秒為一個循環, 進行 40 個循環(StepOne Plus thermal cycler, Applied Biosystems),並分析結果。 GUS 活性定位染色的觀察 圖三是使用Leica M3Z 解剖顯微鏡觀察, Nikon Coolpix P5100 相機照像記錄。圖五 A 是以可見光為光源,使用Leitz Laborlux S 光 學 顯 微 鏡 觀 察 。 照 像 記 錄 系 統 則 是 使 用 PointGrey 公司生產的 CCD 影像裝置(型號 Grasshopper GRAS-20S4C),影像處理軟體為 Fly Capture 2.0。圖五 B、C、D 則是以可見 光為光源,使用Leica TCS SP2 共軛焦顯微鏡 (Leica TCS SP2 - Laser Scanning Spectral Confocal Microscope)觀察,CCD 影像裝置則 是使用Canon EOS 600D 系統與專屬影像處理 軟體。
結 果
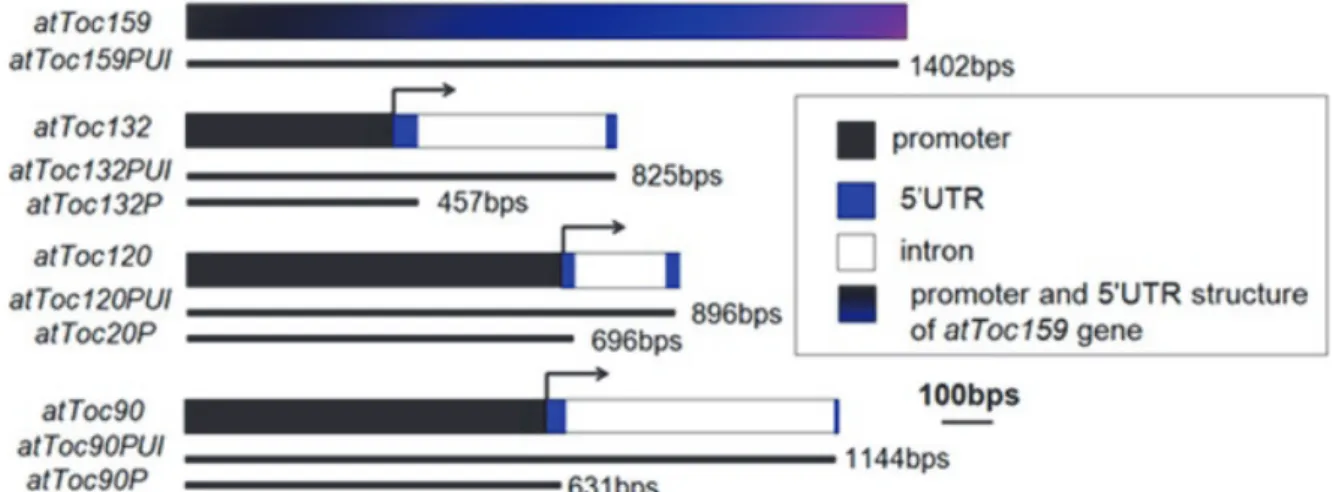
阿拉伯芥穩定轉殖植株的獲得 為了探討光照及內插子序列對於 Toc159 基因家族基因表現量的影響,我們選擇了七條 不同長度的Toc159 基因家族的基因上游調節 序列片段(圖二),並黏合至 GUS 報導基因的 重組載體。再將此重組載體以花絮沾黏法方式 轉殖進入野生型的阿拉伯芥中,以獲得穩定轉 殖植株。這七種轉殖株分成兩類,第一類含有 各基因的啟動子(P)、5′ UTR (U)及內插子(I) 序列,故命名為 PUI 植株,包含 159PUI、 132PUI、120PUI 及 90PUI。第二類只含各基 因的啟動子及部分5′ UTR 序列,故命名為 P 植株,包含132P、120P 及 90P。AtToc159 基 因轉錄起始點仍未知,因此沒有 159P 植株。 最後我們得到了21 個含有 atToc159 基因片段 的穩定轉殖株品系(159PUI-1~21);9 個含有 atToc132PUI 基 因 片 段 的 穩 定 轉 殖 株 品 系 (132PUI-1~9);23 個含有 atToc132P 基因片段 的 穩 定 轉 殖 株 品 系(132P-1~23);9 個 含 有 atToc120PUI 基 因 片 段 的 穩 定 轉 殖 株 品 系 (120PUI-1~9);50 個含有 atToc120P 基因片段 的穩定轉殖株品系(120P-1~50);14 個含有 atToc90PUI 基 因 片 段 的 穩 定 轉 殖 株 品 系 (90PUI-1~14);以及 11 個含有 atToc90P 基因 片段的穩定轉殖株品系(90P-1~11)。每一個轉 殖 株 各 選 三 個 同 型 合 子 的 T3 子 代( 分 為 159PUI-1、159PUI-3、159PUI-8; 132PUI-6、37
圖二、atToc159 家族成員基因的上游調控序列結構圖。箭頭所指的位置為轉錄起始點,然而 atToc159 基
因的轉錄起始點的位置仍未知。
Figure 2. Promoter and 5' UTR structure of four atToc159 gene members. The arrows indicate the transcription
start sites. However, the location of transcription start site of atToc159 is unclear. 132PUI-7 、 132PUI-8; 132P-1 、 132P-13 、
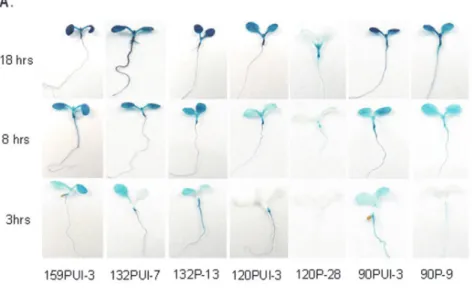
132P-19; 120PUI-1 、 120PUI-3 、 120PUI-9; 120P-3、120P-12、120P-28; 90PUI-1、90PUI-3、 90PUI-9; 90P-1、90P-3、90P-6 等),以作為本 研究的實驗材料。 幼苗發育初期光照因子對於 atToc159 基因家 族的基因表現量的影響 首先,我們以正常的光照週期(16 小時光 照和 8 小時黑暗)和連續黑暗下培養的七天齡 轉殖植株幼苗為材料,進行活體組織GUS 活 性定位染色。生長在正常光照週期之轉殖植株 幼苗浸泡於GUS 活性染色液中,分別在 37 oC 的下反應3 小時、8 小時和 18 小時後,比較 染色的結果。染色3 小時後,159PUI、132PUI 與 90PUI 轉殖植株幼苗全株即可清楚看到藍 色染色結果(圖三 A);在 18 小時染色後,120 PUI 及 90P 轉殖株則顯現全株藍色的結果,但 120P 轉殖株除根及上胚軸呈現淺藍色外,其 餘組織仍為白色。此結果顯示七天齡轉殖植株 中,atToc120 及 atToc90 的上游調節序列啟動 報導基因的能力較atToc159 及 atToc132 差, 這個結果也從 GUS 酵素的活性定量分析及 GUS mRNA 表現量獲得佐證(圖四 A 及 C)。 類似的結果也在七天齡的白化苗中呈現(圖三 B,圖四 B 及 D)。這些研究顯示 atToc159、 atToc132/90 和 atToc120 基因在七天齡的幼苗 中分別有最高,次之及最低的表現量。 內插子序列在 atToc159 基因家族基因表現的 影響 為了瞭解領先內插子序列在 atToc159 基 因家族基因表現上所扮演的角色,我們同樣以 轉殖植株幼苗的活體組織 GUS 定位染色、 GUS 酵素螢光定量和 qPT-PCR 實驗,分別比 較各基因內插子序列對基因表現的影響。檢驗 GUS 基因或 GUS 活性時,很明顯地觀察到 atToc120 基因在缺少內插子序列時,不論正常 光照週期或連續黑暗生長下的七天齡植株,其 表現量皆明顯地下降(圖三及圖四)。光照下, 120PUI 植株的 GUS 基因或 GUS 活性比較 120P 植株高出兩倍左右。而黑暗條件下, 120PUI 植株的 GUS 基因或 GUS 活性比較 120P 植株則提高到三倍左右。這表示 atToc120 的內插子序列能增加基因表現量,且黑暗生長 條件有較明顯的促進基因表現的效果。而 atToc132 和 atToc90 的內插子序列,似乎也有 類似促進基因表現功能,但沒有 atToc120 明 顯(圖三~圖四)。 十天齡轉殖植株之子葉的 atToc159 基因家族 基因表現量差異 在顯微鏡下觀察十天齡轉殖植株之子葉 的活體組織GUS 活性定位染色實驗中,我們 比較不同層面之細胞的染色結果發現,若僅觀 察子葉的下表皮表面,所有轉殖株子葉的保衛 細胞均可偵測到GUS 基因的表現量,尤其以 159PUI、132PUI 和 90PUI 的活性染色程度較 高。然而,利用偏光顯微鏡比較子葉的不同層 面之細胞後,可以發現159PUI、132PUI、132P 等轉殖植株的保衛細胞的染色程度均與葉肉 細胞相似,但90PUI 與 120PUI 等轉殖植株的
圖三、光照與黑暗培養條件下atToc159 基因家族成員的基因表現量差異。A. 正常照光週期(16 小時光照
與8 小時黑暗)生長之七天齡的轉殖植株幼苗的活體組織 GUS 活性定位染色。GUS 染色時間分別為 3、8 及18 小時。B. 連續黑暗生長下之七天齡的轉殖植株幼苗的活體組織 GUS 活性定位染色。染色時間為 3、 8 及 18 小時。
Figure 3. Expression of atToc159 gene members under light and dark conditions. A. In situ staining of 1-w-old
transgenic seedlings grown under a 16h light/8h dark cycle. Samples were incubated in GUS solution for 3, 8 and 18 hours, respectively. B. In situ staining of 1-w-old seedlings grown in continuous dark.
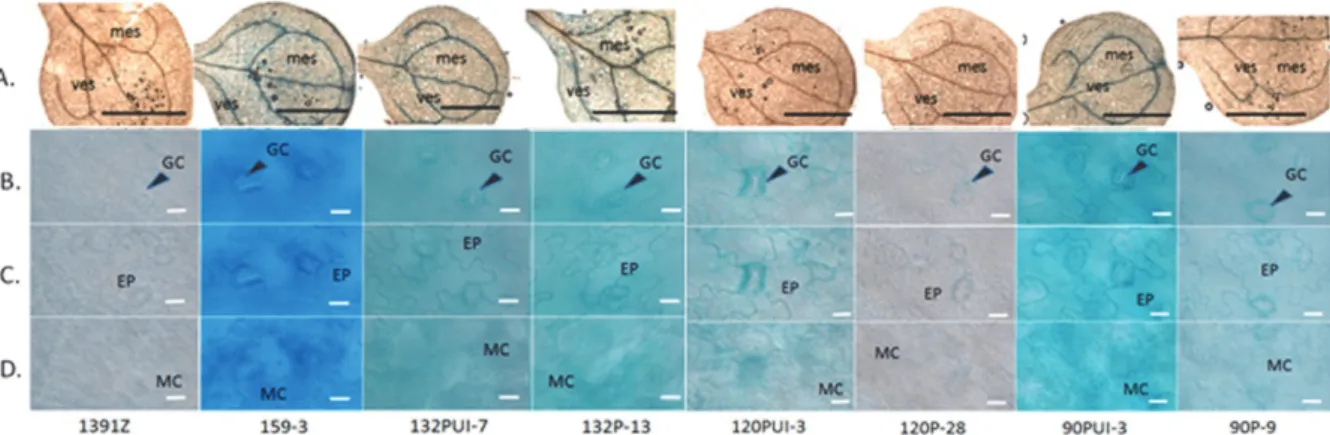
保衛細胞的染色程度均較葉肉細胞明顯。相同 的情形也出現在90P 與 120P 轉殖株中(圖五)。 顯 示 在 子 葉 的 不 同 組 織 中 ,atToc120 和 atToc90 傾向在保衛細胞中增加表現量,而 atToc159 和 atToc132 是葉肉細胞中主要的表 現基因。
討 論
經由穩定轉殖植株的GUS 定性及定量分 析,我們發現在七天齡的幼苗中,atToc159 家族成員呈現基因表現差異:atToc159 有最高量表現,atToc132 和 atToc90 次之,而 atToc120
最低。此外,位於5′ UTR 的領先內插子,似 乎可以提高atToc120 及 atToc90 的基因表現量。 此 種 內 插 子 媒 介 的 基 因 表 現 加 強 作 用 (intron-mediated enhancement,IME),在不同 的環境因子和發育階段對基因表現的影響程 度可能不同。我們的研究顯示,黑暗生長條件 似乎可以提高內插子序列對 atToc120 基因表 現上的影響。最後,atToc159 家族成員在子葉 的不同組織中,顯現細胞專一表現性。因此我 們認為 atToc159 家族成員的基因會受到光照 因子和組織位置的影響,藉由特定的機制調節
39
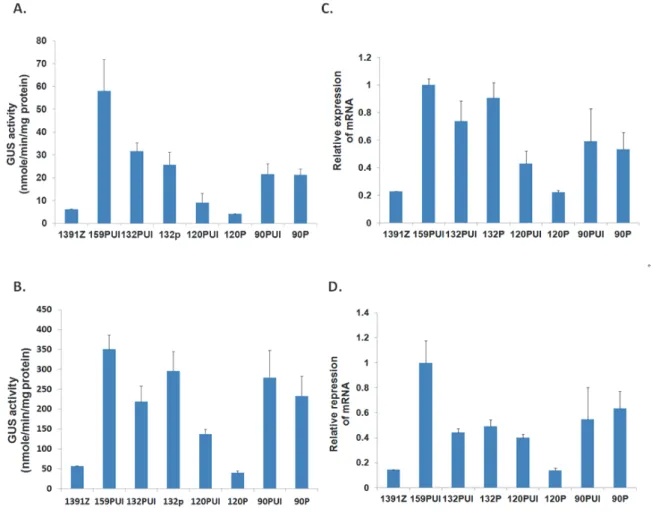
圖四、光照與黑暗條件下內插子效應對基因表現量的影響。A 及 B. 生長於正常照光週期(A)與連續黑暗 條件下(B)之七天齡的轉殖植株幼苗的 GUS 酵素螢光定量分析。GUS 活性的數值是三個不同代表性轉殖 株之平均值。B 及 D. 以 qRT-PCR 分析生長於正常照光週期(C)與連續黑暗條件下(D)之七天齡的轉殖植 株幼苗的GUS mRNA 相對表現量。Y 軸為 atToc159 基因家族成員基因對 UBQ10 基因的相對表現量。GUS
mRNA 量是三個不同代表性轉殖株之平均值。
Figure 4. Expression of atToc159 gene members under light and dark conditions. A and B. GUS enzyme
activity from 1-w-old transgenic seedlings grown under a 16h light/8h dark cycle (A) or continuous dark (B). C and D. GUS mRNA level from 1-w-old transgenic seedlings grown under a 16h light/8h dark cycle (C) and continuous dark (D). The relative expression level of atToc159 gene members to UBQ10 gene is indicated in Y axis. Data are means ± SD of three repeat experiments from three individual transgenic lines.
各個基因的表現量,進而使不同種類的前驅蛋 白在正確的時間運輸進入色質體,以幫助色質 體的發育並協助其發揮正常的功能。 本研究中不管光照或黑暗下生長的植株, atToc159 基因均有一定的 GUS 表現量(圖三~ 五)。雖然一般認為 atToc159 主要負責辨識和 運輸與光合作用有關的蛋白質,atToc159 基因 主要表現幼嫩的發育早期的綠色組織,但也有 一些研究發現attoc159/attoc132 雙突變株會有 嚴重的胚胎死亡(embryo-lethal)的現象(kubis et al., 2004)。因此 atToc159 在所有前軀蛋白的 辨識和運輸中扮演的角色,應不僅僅只是幫助 光合作用有關的前軀蛋白運輸進入色質體 (kubis et al., 2004)。而 atToc90 也被歸類為與
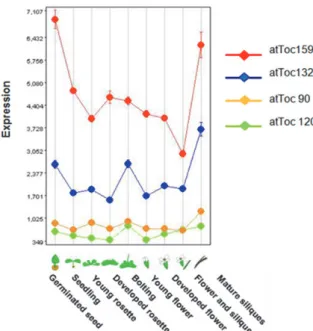
運輸光合作用前軀蛋白有關,但其功能遠不及 atToc159 (Hiltbrunner et al., 2004) 。 因 為 atToc90 可與 atToc33 和 atToc75 直接的進行交 互作用,協助辨識和運輸光合作用的前軀蛋白 (Hiltbrunner et al., 2004)。 比較本研究與 GENEVESTIGSTIGATOR 資料庫的微陣列晶片分析(microarray) 的實 驗數據(圖六,Zimmermann et al., 2004),個別 atToc159 成員的表現形式大致與我們的結果 吻合。atToc159 與 atToc132 的表現量在七天 齡植株中,atToc159 的表現量約為 atToc132 基因的表現量的一倍。但是Kubis 等人(2004) 的RNA 膠體墨漬法(RNA gel blot)實驗結果卻 顯示,十天齡的野生型阿拉伯芥之atToc159
圖五、十天齡轉殖植株之子葉的GUS 活性定位染色。子葉下表皮的細胞形態分別以光學顯微鏡觀察外部 構造(A)及以偏光顯微鏡觀察相同下表皮區域中不同層細胞的 GUS 活性(B、C 及 D)。B 代表下表皮保衛 細胞,C 代表下表皮細胞層,D 代表葉肉細胞層。染色時間為 1.5 小時。mes, 葉肉組織; vas, 維管束組織; GC, 保衛細胞; MC, 葉肉細胞; EC, 表皮細胞。Bars = 5 mm (A)或 20 μm (B、C 及 D)。
Figure 5. Histochemical localization of GUS activity in 10-day-old transgenic seedlings. The external and
internal structures of abaxial epidermal layer in cotyledon were observed by optical microscope (A) and polarized optical microscope with visible light source (B, C and D). B~D represents guard cells on abaxial epidermal surface, abaxial epidermal cells and mesophyll cells, respectively. Tissues were incubated in GUS solution for 1.5 hours. mes, mesophyll; vas, vascular tissue; GC, guard cell ; MC, mesophyll cell; EC, epidermis cell. Bars = 5 mm (A) or 20 μm (B, C and D).
表現量較atToc132 的表現量多 10 倍左右。Yu
等人(2001)以 RT-PCR 技術比較一個月齡的野 生型植 株之基因表 現,卻發 現 atToc159、 atToc132 及 atToc120 的 mRNA 表現量幾乎相
同。雖然不同研究的結論差異很大,但可以確 定的是 atToc159 基因的表現量會受到發育早 晚期的影響。另一方面,不同的研究室針對 atToc90 表現量的結論就有很大的差異。我們 認為atToc132 與 atToc90 在七天齡植株中的表 現 量 的 大 致 相 似 , 而 GENEVESTIGSTIGA TOR 資料庫卻出現 atToc132 表現量較 atToc90 表現量高兩倍左右,Kubis 等人(2004)認為 atToc90 表現量較 atToc132 表現量甚至高到三 倍左右。因為我們是以含有 atToc159 家族成 員的上游調節序列及報導基因重組質體之穩 定轉殖株,去比較各成員的基因表現量,這與 植物內生性基因表現的真正情況可能有差異。 但不同的研究方法和資料庫有關 atToc159 家 族基因表現量的高度差異,仍有待進一步的實 驗來加以釐清。 已有報告指出一些特殊位置的內插子序 列可能在基因調節上扮演重要的功能,例如領 先內插子序列會調節特定基因的表現形式和 表現量(Jeong et al., 2006;Rose et al., 2008), 雖然其詳細的作用機制仍不是很清楚。這種內 插子序列可以提高基因的表現量的現象,特稱 為 內 插 子 強 化 基 因 表 現 (IME) 的 作 用 (Mascarenhas et al., 1990)。許多研究顯示內插 子提升基因表現量的程度,可能與內插子所在 的位置、鄰近內插子的序列和轉殖質體所含的 報導基因種類有關(Luehrsen and Walbot, 1991; Rethmeier et al., 1998),也可能與所在的組織 或細胞種類有關(Gallie and Toung, 1994)。在 本研究中除了 atToc159 基因的起始轉錄點尚 未確認外,其他atToc159 家族基因的 5′ UTR 內均具有一個領先內插子序列。我們發現在不 同的植株發育階段與不同組織,atToc120 及 atToc90 各自的領先內插子序列,都可以提高 植株內其相對應基因的表現量。所以我們認為 在這些基因的領先內插子序列中,可能包含與 發育調節和組織細胞表現專一性有關的基因 強化子(enhancer),當然我們需要設計更多的 實驗,才能證實這樣的說法。 一般而言,植物體內葉肉細胞和保衛細胞 中均存在正常功能的葉綠體。其中,葉肉細胞 是植物主要進行光合作用製造醣類的場所。而 植物必須藉由保衛細胞控制氣孔的開閉,植物 才能具有蒸散作用和吸收CO2 的能力,這些生 理功能也都與保衛細胞葉綠體的光合作用有 關,所以維持保衛細胞內葉綠體的正常功能必 定對於植物的生長發育具有關鍵的影響力。令 人感興趣的是,我們有關十天齡的轉植株GUS 活性定位染色結果顯示, 120PUI與90PUI植 株的保衛細胞GUS酵素活性比葉肉細胞高(圖 五)。Yu等人(2001)研究發現ppi2 (atToc159 knockout mutant)突變株的子葉的碘液染色中
41 圖六、atToc159 基因家族成員在阿拉伯芥不同發育
階段的基因表現量。資料來自GENEVESTIGATOR database 網站(Zimmermann et al., 2004)。
Figure 6. Expression profile of atToc159 gene
members during Arabidopsis development. This expression profile was downloaded from GENEVESTIGATOR database (Zimmermann et al., 2004). 以保衛細胞最為明顯,並在保衛細胞中仍可發 現正常的葉綠囊膜的堆疊和澱粉的累積。相反 的,葉肉細胞的色質體發育卻出現嚴重的缺失。 顯然地,正常功能的atToc159是葉肉細胞色質 體發育所必需,但atToc159在保衛細胞色質體 的功能上扮演並非絕對的需要的角色。我們認 為喪失 atToc159正常功能的ppi2變株的子葉 中,葉肉細胞的葉綠體也失去正常的功能,因 為atToc159在葉肉細胞中仍扮演重要的葉綠 體的辨識和運輸機組蛋白角色。這樣的推論可 以由一些有關ppi2植株的葉肉組織發育的相
關研究得到佐證(Asano et al., 2004)。而ppi2植 株的子葉保衛細胞卻仍存在正常功能的葉綠 體,則可能是因為atToc159功能由atToc90或 atToc120等其他同源蛋白所取代。也就是說, 子葉保衛細胞中可能存在與atToc159無關的 運輸光合作用蛋白質的途徑。這樣的研究結果 似乎支持Bischof等人(2011)的蛋白質體學與 轉錄體學的研究,色質體前驅蛋白質可能分為 兩 大 類 , 亦 即 非Toc159 依 賴 型 (Toc159- independent) 和 Toc159 依 賴 型 (Toc159- dependent)兩種,且atToc159對於一些光合性 前驅蛋白的運輸並不具有專一性。 雖然最近以ppi2 突變株為基因背景下過 度表現atToc90 基因的研究中,已發現大量表 現atToc90 可以部分恢復 ppi2 突變株累積光合 作用蛋白的能力(Infanger et al., 2011),但是至 今仍缺乏相關於 atToc159 基因家族成員基因 在保衛細胞專一性表現的相關研究,只有一些 關於保衛細胞原生質體的微陣列晶片分析的 實驗數據(Zimmermann et al., 2004)。所以 atToc90 或 atToc120 在保衛細胞中葉綠體發育 上所扮演的角色,有待進一步的實驗加以證實。 此外,有關 ppi2 突變株的組織發育上研究也 發現,ppi2 突變株的子葉的大小、根形態、莖
頂分生組織(shoot apical meristem)的構造與發 育早期的葉原基組織(leaf primordia)與野生型 並沒有不同。然而,ppi2 突變株的葉子在發育 後期卻發生嚴重的構造上的缺失(Asano et al., 2004)。似乎指出 atToc159 在發育早期所扮演 的角色較不重要,或者色質體對幼苗早期發育 較不重要,或是 atToc159 的其他三種同源蛋 白部分取代了 atToc159 的功能。綜合以上的 結果可以發現,Toc159 主要在葉肉細胞中發 揮重要的功能,而其他不同的 atToc159 的同 源蛋白質的其他成員似乎作用在不同的組織 或細胞中發揮功能,不同的 atToc159 蛋白質 機組成員所組成 TOC 複合體可能具有細胞的 專一性。 綜合以上的結果,我們可以知道不同的 atToc159 家族基因成員的相關啟動子的序列 會影響基因的組織和細胞的專一性表現,而5′ UTR 內領先內插子序列則可增加基因的表現 量 。 這 些 研 究 結 果 支 持 葉 綠 體 轉 運 蛋 白 atToc159 基因家族之基因的表現在不同的組 織和發育階段的差異,可能是受到不同的發育 和環境的因子刺激所調節。希望這些研究,不 僅只是找到細胞核中 atToc159 基因家族能的 調節機制,也幫助我們更進一步暸解色質體可 能發育的過程。
誌 謝
本報告之完成要感謝行政院國家科學委 員 會 提 供 經 費(NSC98-2311-B-003-002 and NSC99-2311-B-002-MY3)。感謝師大生命科學 系張永達教授慷慨提供光學顯微鏡設備與師 大影像核心實驗室提供共軛焦顯微鏡設備,作 為子葉GUS 活性定位染色觀察使用。參考文獻
Asano T, Yoshioka Y and Machida Y. 2004. A defect in atToc159 of Arabidopsis thaliana
causes severe defects in leaf development. Genes Genet. Syst. 79: 207-12.
Bauer J, Chen K, Hiltbunner A, Wehrli E, Eugster M, Schnell D and Kessler F. 2000. The major protein import receptor of plastids is essential for chloroplast biogenesis. Nature 403: 203-207.
Bischof S, Baerenfaller K, Wildhaber T, Troesch R, Vidi PA, Roschitzki B, Hirsch- Hoffmann M, Hennig L, Kessler F, Gruissem W and Baginsky S. 2011. Plastid proteome assembly without Toc159: photosynthetic protein import and accumulation of N-acetylated plastid precursor proteins. Plant. Cell 23: 3911-28. Chen YJ and Sun CW. 2010.Transgenic study of
chloroplast translocon gene regulation in
Arabidopsis thaliana. Bot. Stud. 51:
147-153.
Chomczynski P and Sacchi N. 1987. Single Step Method of RNA Isolation by acid Guanidinium Thiocyanate-Phenol- Chloroform Extraction. Anal. Biochem.
162: 156-159.
Clough S and Bent A. 1998. Floral dip: a simplified. method for agrobacterium- mediated transformation of Arabidopsis
thaliana. Plant J. 16: 735-743.
Gallie DR and Young TE .1994. The Regulation of Gene Expression in Transformed Maize Aleurone and Endosperm Protoplasts' Analysis of Promoter Activity, Intron Enhancement, and mRNA Untranslated Regions on Expression. Plant Physiol. 106: 929-939.
Hiltbrunner A, Grunig K, Alvarez-Huerta M, Infanger S, Bauer J and Kessler F. 2004. AtToc90, a new GTP-binding component of the Arabidopsis chloroplast protein import machinery. Plant Mol. Biol. 54: 427- 440.
Infanger S, Bischof S, Hiltbrunner A, Agne B, Baginsky S and Kessler F. 2011. The chloroplast import receptor Toc90 partially restores the accumulation of Toc159 client proteins in the Arabidopsis thaliana ppi2 mutant. Mol. Plant 4: 252-263.
Inoue H, Rounds C and Schnell D. 2010. The molecular basis for distinct pathways for protein import into Arabidopsis chloroplasts. Plant Cell 22: 1947-1960.
Ivanova Y, Smith MD, Chen K and Schnell D. 2004. Members of the Toc159 import
receptor family represent distinct pathways for protein targeting to plastids. Mol. Biol. Cell 15: 3379-3392.
Jarvis P and Robinson C. 2004. Mechanisms of protein import and routing in chloroplasts. Curr. Biol. 14: 1064-1077.
Jarvis P. 2008. Targeting of nucleus-encoded proteins to chloroplasts in plants. New Phytol. 179: 257-285.
Jefferson RA. 1987. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mol. Biol. Rep. 5: 387-405.
Jeong YM, Mun JH, Lee I, Woo JC, Hong CB and Kim SG. 2006. Distinct roles of the first introns on the expression of Arabidopsis profile in gene family members. Plant Physiol. 140: 196-209. Kubis S, Patel R, Combe J, Bédard J,
Kovacheva S, Lilley K, Biehl A, Leister D, Ríos G, Koncz C and Jarvis P. 2004. Functional Specialization amongst the Arabidopsis Toc159 Family of Chloroplast Protein Import Receptors. Plant Cell 16: 2059-2077.
Lamesch P, Berardini TZ, Li D, Swarbreck D, Wilks C, Sasidharan R, Muller R, Dreher K, Alexander DL, Garcia-Hernandez M, Karthikeyan AS, Lee CH, Nelson WD, Ploetz L, Singh S, Wensel A and Huala E. 2012. The Arabidopsis Information Resource (TAIR): improved gene annotation and new tools. Nucleic Acids Res. 40: 1202-1210.
Luehrsen KR and Walbot V. 1991. Intron enhancement of gene expression and the splicing efficiency of introns in maize cells. Mol. Gen. Genet. 225: 83-93.
Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, Lins T, Leister D, Stoebe B, Hasegawa M and Penny D. 2002. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc. Natl. Acad. Sci. 99: 12246-12251. Mascarenhas D, Mettler IJ, Pierce DA and Lowe
HW. 1990. Intron-mediated enhancement of heterologous gene expression in maize. Plant Mol. Biol. 15: 913-20.
Murashige T and Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant 15: 473-497.
43 Rethmeier N, Kramer E, Va M, Montagu N and
Cornelissen M. 1998. Identification of cat sequences required for intron-dependent gene expression in maize cells. Plant J. 13: 831-835.
Rochaix JD. 2004. Genetics of the biogenesis and dynamics of the photosynthetic machinery in eukaryotes. Plant Cell 16: 1650-1660.
Rose AB, Elfersi T, Parra G and Korf I. 2008. Promoter proximal introns in Arabidopsis
thaliana are enriched in dispersed signals
that elevate gene expression. Plant Cell 19: 1898-1911.
Smith M, Hiltbrunner A, Kessler F and Schnell D. 2002. The targeting of the atToc159 preprotein receptor to the chloroplast outer membrane is mediated by its GTPase domain and is regulated by GTP. J. Cell Biol. 159: 833-843.
Smith M, Rounds C, Wang F, Chen K, Afitlhile M and Schnell D. 2004. The atToc159 is a selective transit peptide receptor for the import of nucleus-encoded chloroplast proteins. J. Cell Biol. 165: 323-334.
Wallas T, Smith M, Sanchez-Nieto S and Schnell D. 2003. The roles of Toc34 and Toc75 in targeting the Toc159 preprotein receptor to chloroplasts. J. Biol. Chem. 278: 44289-44297.
Yu TS and Li Hm. 2001. Chloroplast protein translocon components atToc159 and atToc33 are not essential for chloroplast biogenesis in guard cells and root cells. Plant Physiol. 127: 90-96.
Zimmermann P, Hirsch-Hoffmann M, Hennig L and Gruissem W. 2004. GENEVESTIGA TOR. Arabidopsis microarray database and analysis toolbox. Plant Physiol. 136: 2621-2632.
Cell-type Specific and Leader Intron-regulated Expression of Arabidopsis
atToc159 Translocon gene family
Yu-Shan Liu and Chih-Wen Sun
*Department of Life Science, National Taiwan Normal University Taipei, Taiwan
(Received: 12 April 2013, accepted: 9 May 2013)
ABSTRACT
In Arabidopsis, four Toc159 homologues are reported and could be classified to two subgroups, atToc159/atToc90 and atToc132/atToc120, based on their sequence homology and substrate specificity. Therefore, expression of these translocon genes have to be regulated properly, otherwise the development and maintenance of plastids would be prohibited due to lack of essential protein import. In order to reveal the regulatory mechanism of atToc159 gene family members, transgenes containing various lengths of the upstream sequences of these translocon genes and GUS coding sequence were transferred to wild-type
Arabidopsis. The steady accumulation of GUS expression from the vegetative tissues of these
transformants was examined first. In general, atToc159, atToc132/atToc90, and atToc120 have relatively high, moderate, and low expression levels in various tissues of 7-day-old seeldings. Furthermore, the expression of atToc120 and atToc90 could be up-regulated by their corresponding leader intron sequence within 5′ untranslated region (5′ UTR) region. To further understand the intron effect of atToc120 expression was regulated by light, the GUS activity of 120PUI and 120P plants grown under light/dark cycle and continuous dark conditions were compared. Our data suggests that the intron sequence of
atToc120 is more responsive to dark signal. In addition to tissue-specific expression, the cell-specific
expression of these transgenic plants was also determined. Surprisedly, atToc90 and atToc120 had higher GUS activity in the guard cells than mesophyll cells of cotyledon when compared to atToc159 and
atToc132. This indicates that atToc90 and atToc120 might play a more important role in regulating
chloroplast protein import of guard cells. These data support that the spatial and temporal expression of
atToc159 gene family members is regulated by different developmental and environmental stimuli.