387
Ann. N.Y. Acad. Sci. 1042: 387–395 (2005). © 2005 New York Academy of Sciences. doi: 10.1196/annals.1338.059
Tumor Necrosis Factor
␣, Prostaglandin E2,
and Cyclooxygenase-2 Production in
Lipopolysaccharide-Activated Mouse
Macrophages
SHENG-TUNG HUANG,a CHIEN-TSU CHEN,a KUR-TA CHIENG,a SHIH-HAO HUANG,b BEEN-HUANG CHIANG,b LENG-FANG WANG,a HSIEN-SAW KUO,a AND CHUN-MAO LINa
aCollege of Medicine, Taipei Medical University, Taipei, Taiwan
bGraduate Institute of Food Science and Technology, National Taiwan University,
Taipei, Taiwan
ABSTRACT: Isovitexin, isolated from rice hull of Oryza sativa, has been charac-terized as a potent antioxidant. Its antioxidant activity, determined on the basis of inhibition of lipid peroxidation by the Fenton reaction, was comparable with that of ␣-tocopherol, a well-established antioxidant. Isovitexin was able to re-duce the amount of hydrogen peroxide production inre-duced by lipopolysaccha-ride (LPS) in mouse macrophage RAW264.7 cells. In this study, we assessed its effects on the production of tumor necrosis factor ␣ (TNF-␣),prostaglandin E2 (PGE2), and the expression of cyclooxygenase-2 (COX-2) in LPS-activated RAW
264.7macrophages. Isovitexin inhibited the release of TNF-␣, a proinflammato-ry cytokine, upon LPS activation with a 50% inhibitoproinflammato-ry concentration (IC50) of
78.6 M. Isovitexin markedly reduced LPS-stimulated PGE2 production in a
concentration-dependent manner, with an IC50 of 80.0 M. The expression of
COX-2 was also inhibited by isovitexin treatment. Our results suggest that sup-pression of ROS-mediated COX-2 exsup-pression by isovitexin is beneficial in re-ducing inflammation and carcinogenesis.
KEYWORDS: antioxidant; inflammation; isovitexin; COX-2; PGE2; tumor
necrosis factor
INTRODUCTION
Oxidative stress has been implicated in a variety of pathological processes, in-cluding aging, cancer, diabetes mellitus, atherosclerosis, neurological degeneration, and arthritis. Increased uptake of antioxidants may prevent organ injury associated
Address for correspondence: Chun-Mao Lin, Ph.D., College of Medicine, Taipei Medical Uni-versity, No. 250, Wu-Hsing Street, Taipei 110, Taiwan. Voice: +886-2-27361661 ext. 3152; fax: +886-2-27361661 ext. 3163.
with excessive generation of reactive oxygen species (ROS).1,2 ROS have been shown to initiate a wide range of toxic oxidative reactions.3 The effects of oxidants on signaling pathways are often characterized as resulting from heightened oxidative stress. Many antioxidants also exhibit anti-inflammatory effects. Theses observa-tions provide a significant molecular basis for understanding the mode of acobserva-tions of selected dietary ingredients in preventing diseases associated with heightened oxi-dative stress.
Tumor necrosis factor α (TNF-α), a proinflammatory cytokine, and prostaglan-din E2 (PGE2) are key mediators in inflammatory reaction. TNF-α generated in
in-flammation may induce tissue damage.4 Prostaglandins, especially PGE2, are involved in stimulating cellproliferation, tumor growth, and suppressing the im-mune responseto malignant cells. Overproduction of prostaglandins from upregula-tion of cyclooxygenase-2 (COX-2) in cells had been associated with malignant growth.5 COX-2 is rapidly induced by tumor promoters,growth factors, cytokines, and mitogens in various cell types.6 Many cell types associated with inflammation, suchas macrophages and endothelial cells and fibroblasts, may be induced to over-expressCOX-2.7 Treatment with TPA (12-O-tetradecanoyl-phobol-13-acetate) in mice led to edema and papilloma formation by enhancing COX-2 expression. Spe-cific COX-2 inhibitors could counteract these biological events. Suppression of COX-2 induction and or its activity may be an effective approach for the prevention of carcinogenesis in several organs. Selected inhibitors of COX-2 may also have a therapeutic role in certain cancers.
Rice is an important dietary staple in Asia, where the incidence of breast and co-lon cancer is markedly lower than that in the western world.8 It has been reported that rice constituents counteract chemical-induced mutagenicity, tumor promotion, and carcinogenicity.9 Constituents from rice bran have been found to be beneficial for cancer prevention by epidemiological survey. Rice bran contains several classes of chemopreventive agents (e.g., flavonoids and their glycosides, tocotrienols, and γ-oryzanol). Isovitexin and related flavonoids are constituents of the rice hull of
Ory-za sativa and have been shown to exhibit potent antioxidant activity,10 including in-hibition of xanthine oxidase, protection of DNA from oxidative damage, and prevention of heavy-metal–induced cell injury.11 Here, we further studied the anti-oxidative properties of isovitexin by examining its effects on LPS-induced PGE2 and
TNF-α production and COX-2 expression in the murine macrophage-like cell line RAW264.7.
MATERIALS AND METHODS Materials
Isovitexin, a glycosylflavonoid, was isolated from rice hull as described previous-ly.12 The final product showed a major peak on capillary chromatography (P/ACE 5000; 75 µm × 37 cm [Beckman, Fullerton, CA]; borax buffer, 20 mM, pH = 10.0; UV absorbance at 214 nm; 25°C; UV absorbance at 214 nm, 25°C, 10K voltage; in-jection pressure, 80 psi; migration time: 6.64 min.). LPS (Escherichia coli O127:B8) and chemicals were purchased from Sigma (St. Louis, MO) unless specified.
Cell Culture
The mouse monocyte–macrophage cell line RAW 264.7 (ATCC TIB-71) was cul-tured in Dulbecco’s modified Eagle’s medium (Gibco, Grand Island, NY) supple-mented with 10% heat-inactivated fetal bovine serum (Gibco). Cells were plated in 24-well plates or petri dishes before activation by LPS (100 ng/mL). Isovitexin dis-solved in dimethyl sulfoxide (DMSO) with the final DMSO concentration of less than 0.2% (vol/vol) was administered with LPS. Control samples contained the same concentration of DMSO.
Lipid Peroxidation
Ethyllinoleate was oxidized by the Fenton reaction (Fe2+/H2O2). In selected ex-periments, equal concentration (30 µM) of apigenin, isovitexin, and α-tocopherol were added to 0.5 mL of an aqueous solution containing 1.5 mg/L ethyl linoleate, 0.25 mM Trizma–HCl/0.75 mM KCl buffer (pH 7.4), 0.2% N-lauroyl sarcosine, 1µM ferrous chloride, and 0.5 µM hydrogen peroxide in a 2-mL microtube. The mixture was incubated for 16 h at 37°C. The quantity of oxidation was measured by the thiobarbituric acid (TBA) assay.11 The antioxidative activity of the samples was calculated according to the following formula: antioxidative activity (%) = 1 – (ab-sorbance of sample/ab(ab-sorbance of control) × 100. The level of TBA-reactive sub-stance (TBARS) by autooxidation of ethyllinoleate was calculated as the amount of malondialdehyde.
Determination of TNF-␣ Release
Murine macrophages were seeded in 24-well plates at a density of 5.0 × 105 cells/
well the day before the experiment. Cells were treated with or without isovitexin (10–100 µM) and/or LPS (100 ng/mL) in 500 µL of medium containing 10% fetal bovine serum for 1 h at 37°C. TNF-α levels in the media were then determined by a quantitative sandwich enzyme-linked immunosorbent assay using the commercially available mouse TNF-α immunoassay kit (Amersham, Buckinghamshire, UK) ac-cording to the manufacturer’s instructions. All experiments were done in triplicate.13
PGE2 Assay
Cells were plated at a density of 1.0 × 106 cells/mL in 24-well culture plate and stimulated with LPS (50 ng/mL) in the presence or absence of various concentrations of isovitexin for 18 h. The culture medium of control and treated cells was collected and centrifuged. Thelevel of PGE2 released into culture medium was determined
us-ingan enzyme immunoassay according to the manufacturer’sinstructions (Amer-sham, Buckinghamshire, UK).4
Western Blotting
The cellular protein fraction was prepared using lysis buffer containing 10% glyc-erol, 1% Triton X-100, 1 mM sodium orthovanadate, 1 mM EGTA, 5 mM EDTA, 10 mM NaF, 1 mM sodium pyrophosphate, 20 mM Tris–HCl (pH 7.9), 100 µM β-glycerophosphate, 137 mM NaCl, 1 mM phenylmethylsulfonyl fluoride (PMSF), 10 µg/mL aprotinin, and 10 µg/mL leupeptin. Proteins (50 µg) were separated on
so-dium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and elec-trotransferred to a polyvinylidene difluoride (PVDF) membrane (Immobilon-P; Millipore, Bedford, MA). The membrane was incubated with an anti–COX-2 mon-oclonal antibody (Oncogene Science, Cambridge, UK) containing 1% bovine serum albumin and 0.2% NaN3 overnight at 4°C. After incubation with horseradish
perox-idase–conjugated anti–mouse immunoglobulin G antibody (Oncogene Science, Cambridge, UK), the immunoreactive bands were visualized with enhanced chemi-luminescent reagents (ECL; Amersham, Buckinghamshire, UK). Relative protein content in each band was assessed using a densitometer (Alpha Innotech IS-1000; Digital Imaging System, San Leandro, CA).
FIGURE 1. Antioxidative activity of isovitexin. (A) Isovitexin (Itx) inhibited the
Fen-ton reaction–induced lipid peroxidation. Isovitexin (Itx), α−tocopherol, and apigenin (30 µM) were compared on the basis of their antioxidative activities. Quantity of oxidation was measured using the TBA assay. Antioxidative activity of the samples was measured accord-ing to levels of TBA-reactive substance (TBARS). (B) Isovitexin suppressed LPS-induced production of hydrogen peroxide. RAW 264.7 cells were treated with LPS (100 ng/mL) alone or LPS and isovitexin (20 and 40 µM) for 30 min. Cells were stained with DCFH-DA and subjected to flow cytometry. Peaks of FL1-H fluorescence intensity are indicated for each treatment.
Flow Cytometric Detection of Hydrogen Peroxide
RAW 264.7 cells were suspended in phenol red–free medium at a density of 105cells/mL. Cells were stained with 100 µM 2′,7′-dichlorofluorescin diacetate (DCFH-DA) in the dark for 30 min and then analyzed using a FACScan (Becton Dickinson, San Jose, CA). Oxidation of green DCH fluorescence by hydrogen per-oxide in living cells was detected using the FL1-H wavelength band. The fluores-cence signals of 10,000 cells were processed using a logarithmic amplifier as described previously.14
RESULTS AND DISCUSSION Antioxidative Activity of Isovitexin
Many antioxidants exert anti-inflammatory effects, and many polyphenolic com-pounds have been reported to be potent ROS scavengers. The antioxidative activity of isovitexin was evaluated by the TBA method. The inhibitory effect on ethylli-noleate oxidation by isovitexin (69.5% inhibition, TBARS value = 2.43 µM) was comparable to that of α-tocopherol (70.8% inhibition, TBARS value = 2.33 µM) and higher than that of apigenin, an aglycon flavonoid, (40.7% inhibition, TBARS value = 4.73 µM) (FIG. 1A). The results suggest that isovitexin is a potent antioxidant.
Isovitexin also lowered the LPS-induced increase in the cellular content of hydro-gen peroxide in RAW 264.7 macrophage cells in a dose-dependent manner (FIG. 1B).
Isovitexin Inhibition of TNF-␣ Production
To determine the effect of isovitexin on LPS-activated TNF-α production, we stimulated the cells with LPS in the presence or absence of isovitexin at
concentra-FIGURE 2. Inhibition of TNF-α production by isovitexin. RAW 264.7 cells were
treat-ed with LPS (100 ng/mL) in the presence or absence of isovitexin (Itx) (0, 10, 30, and 100 µM, respectively) for 1 h. Amount of TNF-α released into culture medium was determined. Data are presented as means ± standard error of the mean (n = 3).
tions ranging from 10 to 100 µM for 60 min. The production of TNF-α by the un-stimulated RAW 264.7 cells was 20 pg/mL (n = 3). Incubation of these cells with LPS (100 ng/mL) for 1 h caused a substantial increase in TNF-α production (318.9 pg/mL; n = 3). When RAW 264.7 cells were stimulated with LPS in the pres-ence of isovitexin (10–100 µM), a concentration-dependent inhibition of TNF-α production with a 50% inhibitory concentration (IC50) value of 78.6 µM was observed (FIG. 2).
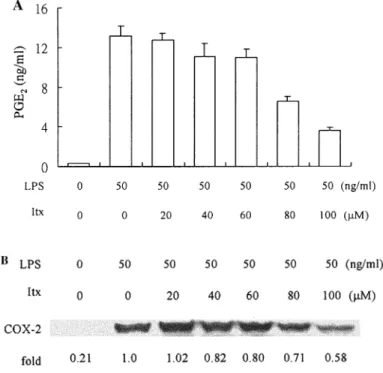
Isovitexin Inhibition of PGE2 Production and COX-2 Expression
The effect of isovitexin on PGE2production and COX-2 protein expression fol-lowing LPS stimulation was examined 18 h after treatment with LPS (50 ng/mL). PGE2 production was reduced in a concentration-dependent manner with an IC50 of 80.0 µM (FIG. 3A). The inhibition was not due to general cellular toxicity (data not shown). LPS-induced COX-2 protein expression was also suppressed by isovitexin in a concentration-dependent manner after treatment for 18 h. The relative protein levels of COX-2 in the presence of 20, 40, 60, 80, and 100 µM isovitexin were 1.02,
FIGURE 3. Effects of isovitexin on LPS-induced PGE2 generation and COX-2 expres-sion. (A) Cells were treated with LPS (50 ng/mL) in the presence or absence of isovitexin (Itx) for 18 h. Amount of PGE2 released into culture medium was determined. Data are pre-sented as means ± standard error of the mean (n = 3). (B) Values for expression of COX-2 protein upon various treatments for 18 h were quantities after resolving in 8.0% SDS- PAGE and Western blot analysis. Relative level of COX-2 expression observed with LPS alone was set at 1.0.
0.82, 0.80, 0.71, and 0.58, respectively, in reference to a value of 1.00 in samples treated with LPS alone (FIG. 3B).
APossible Mechanism for Isovitexin Inhibition of LPS Activity
Macrophages release various mediators in cellular inflammatory response. Among those released are ROS including H2O2, nitric oxide, superoxide, and hy-droxyl radical. The intracellular ROS production is associated with several cellular events including the activation of NAD(P)H oxidase, xanthine oxidase, and the cel-lular mitochondrial respiratory chain.15,16 ROS thus formed are potent activators of NF-κB, an essential transcription factor in the inflammatory signal transduction pathway.17 ROS plays an essential role in the initiation of the NIK/MAPK cascade to trigger a series of responses in which IκB is phosphorylated to free NF-κB from IκB inhibition.18 The active form of NF-κB is translocated from cytoplasm to the
nu-cleus to bind the cognate NF-κB binding site in the promoter regions of genes in-cluding COX-2.19,20 COX-2 overexpression mediated by activation of NF-κB is a possible molecular mechanism in cellular transformation to neoplasia. Another con-sequence of excessive ROS generation is oxidative DNA damage leading to gene
FIGURE 4. Possible mechanisms in which isovitexin (Itx) exerts its action against
mutations and cancer formation. The inflammation-signaling transduction pathway mediated by LPS on macrophage is depicted in FIGURE 4.
In FIGURE 4, the roles of isovitexin in inhibiting inflammation and carcinogenesis
are proposed. Isovitexin, with its potent antioxidant activity, is likely to suppress NF-κB activation. This contention is supported by the finding that the expression of TNF-α and COX-2, both transactivated by NF-κB, was suppressed by isovitexin. Re-duced PGE2 formation by isovitexin is likely due to its inhibition of COX-2
expression.
In summary, isovitexin is a potent antioxidant that inhibits TNF-α and COX-2 ex-pression caused by LPS in mouse macrophage RAW264.7 cells. This effect of iso-vitexin is likely due to its inhibition of NF-κB activation via its antioxidant activity. Isovitexin also inhibits PGE2 formation, likely secondary to its suppression of COX-2 expression. These antioxidant and anti-inflammatory effects of isovitexin may have implications in carcinogenesis. Isovitexin is a food phytochemical enriched in rice. Whether lower incidence of selected neoplasia such as breast and colon cancer in countries with rice as a major staple can be attributed to the antioxidant gradients in rice including isovitexin remains to be determined.
ACKNOWLEDGMENTS
This study was supported by grants from the National Science Council (NSC92-2113-M-038-005 and NSC92-2113-M-038-003).
REFERENCES
1. FITZPATRICK, F.A. 2001. Inflammation, carcinogenesis and cancer. Int. Immunophar-macol. 1: 1651–1667.
2. PAVLICK, K.P., F.S. LAROUX, J. FUSELER, et al. 2002. Role of reactive metabolites of oxygen and nitrogen in inflammatory bowel disease. Free Radic. Biol. Med. 33: 311–322.
3. CUZZOCREA, S., D.P. RILEY, A.P. CAPUTI, et al. 2001. Antioxidant therapy: a new phar-macological approach in shock, inflammation, and ischemia/reperfusion injury. Pharmacol. Rev. 53: 135–159.
4. JU, H.K., S.H. BAEK, R.B. AN, et al. 2003. Inhibitory effects of nardostachin on nitric oxide, prostaglandin E2, and tumor necrosis factor-alpha production in lipopolysac-charide activated macrophages. Biol. Pharm. Bull. 26: 1375–1378.
5. CROFFORD, L.J., R.L. WILDER, A.P. RISTIMAKI, et al. 1994. Cyclooxygenase-1 and -2 expression in rheumatoid synovial tissue. Effects of interleukin-1β, phorbol ester, and corticosteroids. J. Clin. Invest. 93: 1095–1101.
6. INOUE, H., C. YOKOYAMA, S. HARA, et al. 1995. Transcriptional regulation of human prostaglandin-endoperoxide synthase-2 gene by lipopolysaccharide and phorbol ester in vascular endothelial cells. J. Biol. Chem. 270: 24965–24971.
7. HLA, T., A. RISTIMAKI, S. APPLEBY, et al. 1993. Cyclooxygenase gene expression in inflammation and angiogenesis. Ann. N.Y. Acad. Sci. 696: 197–204.
8. KOO, L.S., O.W. MANG, & J.H. HO. 1997. An ecological study of trends in cancer inci-dence and dietary changes in Hong Kong. Nutr. Cancer 28: 289–301.
9. TAKESHITA, M., S. NAKAMURA, F. MAKITA, et al. 1992. Antitumor effect of RBS (rice bran saccharide) on ENNG-induced carcinogenesis. Biotherapy 4: 139–145. 10. RAMARATHNAM, N., T. OSAWA, M. NAMIKI, et al. 1989. Chemical studies on novel rice
hull antioxidants. 2. Identification of isovitexin, a C-glycosyl flavonoid. J. Agric. Food Chem. 37: 316–319.
11. LIN, C.M., C.T. CHEN, H.H. LEE, et al. 2002. Prevention of cellular ROS damage by isovitexin and related flavonoids. Planta Medica 68: 363–365.
12. OSAWA, T., H. KATSUZAKI, Y. HAGIWARA, et al. 1992. A novel antioxidant isolated from young green barley leaves. J. Agric. Food Chem. 40: 1135–1138.
13. GAO, B. & M.F. TSAN. 2003. Endotoxin contamination in recombinant human heat shock protein 70 (Hsp70) preparation is responsible for the induction of tumor necrosis factor alpha release by murine macrophages. J. Biol. Chem. 278: 174–179. 14. UBEZIO P. & F. CIVOLI. 1994. Flow cytometric detection of hydrogen peroxide
produc-tion induced by doxorubicin in cancer cells. Free Radic. Biol. Med. 16: 509–516. 15. IRANI, K., Y. XIA, J.L. ZWEIER, et al. 1997. Mitogenic signaling mediated by oxidants
in RAS-transformed fibroblasts. Science 275: 1649–1652.
16. LEE, S.F., Y.T. HUANG, W.S. WU, et al. 1996. Induction of c-jun protooncogene expres-sion by hydrogen peroxide through hydroxyl radical generation and p60src tyrosine kinase activation. Free Radic. Biol. Med. 21: 437–448.
17. SCHRECK, R., P. RIEBER, & P.A. BAEUERLE. 1991. Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-kappa B transcription factor and HIV-1. EMBO J. 10: 2247–2258.
18. CHEN, C.C., & J.K. WANG. 1999. p38 but not p44/42 mitogen-activated protein kinase is required for nitric oxide synthase induction mediated by lipopolysaccharide in RAW 264.7 macrophages. Mol. Pharmacol. 55: 481–488.
19. SCHMIDT, K.N., P. AMSTAD, P. CERUTTI, et al. 1995. The roles of hydrogen peroxide and superoxide as messengers in the activation of transcription factor NF-kappa B. Chem. Biol. 2: 13–22.
20. CHUN, K.S., H.H. CHA, J.W. SHIN, et al. 2004. Nitric oxide induces expression of cyclooxygenase-2 in mouse skin through activation of NF-kappaB. Carcinogenesis