Spatial and Temporal Variation of Coral Recruitment
in Taiwan
Keryea Soong1, Ming-hui Chen1, 2, Chao-lun Chen3, Chang-feng Dai4, Tung-yung Fan2, Jan-jung Li2, Honmin Fan1, Kun-ming Kuo5, Hernyi Hsieh6
1: Institute of Marine Biology, National Sun Yat-sen University, Kaohsiung, Taiwan 2: National Museum of Marine Biology & Aquarium, Pingtung, Taiwan
3: Institute of Zoology, Academic Sinica, Taipei, Taiwan
4: Institute of Oceanography, National Taiwan University, Taipei, Taiwan 5: Kenting National Park Authority, Pingtung, Taiwan
6: Penghu Aquarium, Taiwanese Fisheries Research Institute, Penghu, Taiwan
Corresponding author: Keryea Soong Fax no.: … -886-7-5255100,
E-mail: [email protected]
Abstract
Coral settlement off Taiwan’s southern coast between 1997 and 2000 was investigated
using artificial plates. A hierarchical experimental design was used, exploiting a total of
723 plastic plates. Scales of spatial variation were analyzed for the above nest-design for
4 consecutive years. In 1999 and 2000 a similar design was expanded to 5 other isolated
coral reefs as well as to 2 non-reef regions around the whole of Taiwan, with a total area
of about 400 km x 200 km, and exploiting an additional 902 plates.
Most coral recruits (~90%) belonged to brooding species, i.e., pocilloporids, with
some acroporids and a few unidentified ones. Densities of coral recruits varied by more
than 3 orders of magnitude between years at certain sites. Some reef sites had none or
very few recruits for the 4 consecutive years of this research.
Significant spatial variation occurred more frequently at small scales than at large
scales during the 4 years of analysis. For example, variations at the “Region” level, with
a scale of 50-200 km, can all be explained by that occurring at smaller scales. The two
non-reef regions recorded no coral recruits in both 1999 and 2000. No combinations of
significant levels repeated in any of the years of this study.
The pattern of recruitment variation found in this study suggests: 1. Self-seeding at
the scale of 10 m is a likely explanation for aggregation of pocilloporid recruits.
Moreover, most coral recruitment of a 10 km reef off Southern Taiwan for example, may
be limited to, or concentrated at, a particular site for several years.
2. Great spatial variation in recruitment may persist for several years for brooding
pocilloporids in Taiwan. The dominating factors, which influence coral recruitment in
Introduction
The effect of recruitment on the structure and dynamics of marine populations and
communities has received much attention in recent years (Caley et al. 1996, Gains and
Roughgarden 1985, Grigg 1988, Roughgarden et al. 1988, Sutherland 1990). Even for
coral reefs, which are often dominated by long-lived corals, recruitment plays a critical
role in the distribution and abundance of the species (Bak and Engel 1979, Connell et al.
1997, Harriott and Banks 1995, Miller et al. 2000, Rylaarsdam 1983).
The history of coral recruitment studies can be traced back a century (Duerden
1904, Harriott and Banks 1995, Stephenson 1931). However, most research on the
subject has been conducted in the last 3 decades (e.g., Birkeland 1977, Birkeland et al.
1981, Goreau et al. 1981, Grigg 1988, Hughes et al. 1999). Research has now shown that
coral larvae are sensitive to microhabitats, e.g., substrate composition (Benayahu and
Loya 1984, Harriott and Fisk 1987), neighbors (Maida et al. 1995), light conditions and
orientation of substrates (Babcock and Mundy 1996, Rogers et al. 1984). Densities and
composition of coral recruits also differ among seasons and between years, often by
several orders of magnitude (e.g., Harriott 1985, Hughes 1985, Rogers et al. 1984,
Wallace 1985, Yoshioda 1996).
Different reefs may have different patterns of recruitment, dominated by either
brooding or broadcasting species (Fitzhardinge 1986, Harriott 1992, Harriott and Fisk
1988, Kojis and Quinn 2001). The patterns of recruitment are known to change with
different spatial scales (Dunstan and Johnson 1998, Fisk and Harriott 1990, Fitzhardinge
the results so far seem to suggest that coral recruitment pattern is a characteristic
particular to a reef, and there is still a lot to learn before a reliable prediction can be made.
The scale of spatial distribution of coral recruitment has been explored mostly on
the GBR and its neighboring reefs (e.g., Hughes et al. 1999). It has been suggested that
the pattern found in this 2000-Km stretch differs from reefs that are relatively isolated
from one another (Dunstan and Johnson 1998, Harriott and Banks 1995). For example, a
larva in the GBR has a good chance, after drifting for 2 weeks and for 200 Km, to settle
on the GBR (Harriott and Fisk 1988). This scale of self-seeding is unlikely for many
other reefs that are relatively isolated and small (e.g., see Kojis and Quinn 2001). Thus, a
different recruitment pattern may exist, and the underlying mechanisms may be revealed
by first investigating the spatial patterns (Levin 1992, Underwood and Chapman 1996)
Information of coral recruitment is important when the reef is degrading due to
natural or anthropogenic factors. A damaged reef may or may not have the potential to
recover depending on the recruitment rate (Edwards and Clark 1998, Hughes and Tanner
2000). Thus recruitment patterns can potentially determine the best approach to conserve
or restore a natural coral reef, once the damaging agents have been eliminated
In this study, we first surveyed the coral recruitment pattern in Southern Taiwan
for two years, then the study sites were expanded to the whole of Taiwan for an additional
2 years. The purpose was to find out the scale of spatial variation that could reveal the
source of recruits, and especially if the above pattern was consistent throughout the years.
Since the way plates are set up influences the densities of coral settlement (see references
in Mundy 2000), several preliminary data sets were obtained before the experiment was
expanded to encompass all of Taiwan.
1. Plate orientation: Stacks of 11 plates with 1-cm space between them were fixed on
either horizontal or vertical orientations. This comparison was made at Hobihu, at 5
m depth. Coral spats that settled on the upper/lower surfaces of horizontal plates, and
those that had settled on surfaces of vertical plates, were compared 1.5 months later.
2. Plate material and density: We compared ceramic tiles and PVC plates in 1997, as
well as 2-plates versus 11-plates per rack in horizontal orientation in 1997 and in
vertical orientation in 1998.
3. Depths: We compared 4 depths, i.e., 1, 5, 10 and 15 m at 2 sites, i.e., Shiju (site 1)
and Tiaoshipi (site 7, Figure 1) in Southern Taiwan, April 1997.
All the above comparisons were made with other factors fixed.
Scale of spatial variation
The data used to analyze scales of spatial patterns were based on coral recruits on
horizontal ceramic tiles in 1997. Then vertical PVC plates were adopted in 1998 and
2000 for higher densities of coral recruits (see Results). The vertical carbonic plates used
in 1999 were no less suitable for coral spats, but they were less resistant to abrasion than
PVC plates. Although there were modifications in procedures between the years (Table
1), the orientation, number of plates per rack and the material of plates were kept constant
between sites in each year.
The plates were placed at 5-10 m depth in the Hengchun Penninsula area, on
(several km apart) were designated, and in each "Area" there were two "Sites", about
1-km apart from each other. Two racks, about 10 m apart, were placed in each of the 8 sites,
and this level (racks) was designated “Locations”. In addition to Southern Taiwan, the
experiment was expanded to 5 other "Regions" covering all the major reefs, i.e.,
Hsiaoliuchiu, Penghu, Northeast, Green Island and Orchid Island around Taiwan, and 2
non-reef “Regions”, i.e., Shitzwan and Yungan, in 1999 and 2000 (See Figure 1). In this
Taiwan–wide study, the whole of the Hengchun Peninsula in Southern Taiwan was
treated as a “region” with 8 sites, i.e., combining “Area” and “Site” into the “Area/Site”
level, with a scale of 1-5 Km between each other. All sites in the reef regions were
fringing reefs (Randall and Cheng 1977, Randall and Cheng 1979). The two non-reef
sites, Shitzwan and Yungan were located along the sandy West coast of Taiwan. A few
species of corals were present on hard or artificial substrates, but there were no reefs at
these two localities. Plates were set at a depth of about 5 m at all the sites.
Spawning of many broadcasting species occurs in late spring (around May) in
Southern Taiwan (Dai et al. 1992), so the plates were placed 1-2 weeks before the
spawning dates and retrieved 4-8 weeks later. In 1998, most late-Spring plates were lost;
although data of another period in January-February were available. Very few coral
recruits occurred in 2000, and retrieval of plates was delayed until September for Orchid
Island.
After collection, the plates were air-dried, and brought back to the laboratory for
analysis using a magnifying glass and a dissecting microscope. Identification of coral spat
was based on backtracking corals 1-2 years old in a separate study (Kuo 2001), as well as
Results
Preliminary experiments
A panel has an area of 225 cm2, including the holes (1.5 cm diameter) we drilled in the
plates. Thus 1 recruit per panel is equivalent to a recruitment rate of 44/m2. Coral
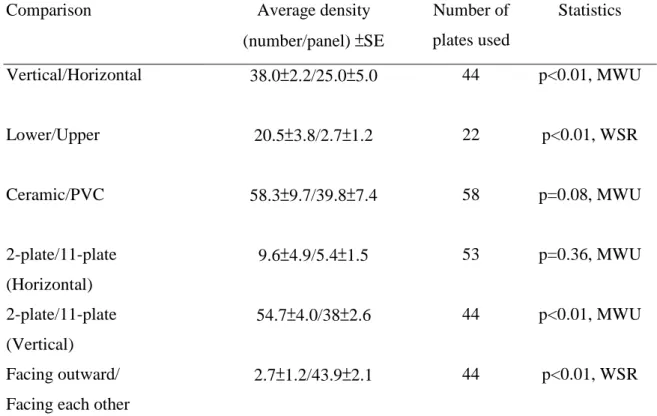
recruitment densities were almost twice as great on vertical as on horizontal plates (38.0
vs. 20.5/panel, p<0.01, Mann-Whitney U test). The above results were obtained after
ignoring the 2 outward facing surfaces of the 11-plate assemblages, since they are
obviously under different environmental conditions. These ignored surfaces often had
high amount of sediments and/or algae, and relatively low number of coral recruits.
Among vertical plates, for example, the surfaces facing each other had an average of 43.9
recruits/panel, higher than the external surface (2.7/panel, p<0.01, Kruskal-Wallis test).
Among horizontal plates, lower surfaces had higher densities of coral recruits than upper
surfaces (20.5 vs. 2.7/panel, p<0.01, Wilcoxon signed rank test).
The difference in settlement on ceramic tiles (58.3±9.7/panel) and PVC plates
(39.8±7.4/panel) was not significant (p=0.08, Mann-Whitney U test). The number of
plates per rack made a difference in coral recruit densities among vertically orientated
racks. Higher densities were recorded in 2-plate racks than in 11-plate racks (54.7 vs.
38.0/panel, p<0.01, Mann-Whitney U test). However, no such significant difference was
found among horizontal plates (9.6 vs. 5.4/panel, p=0.36, Mann-Whitney U test, Table 2).
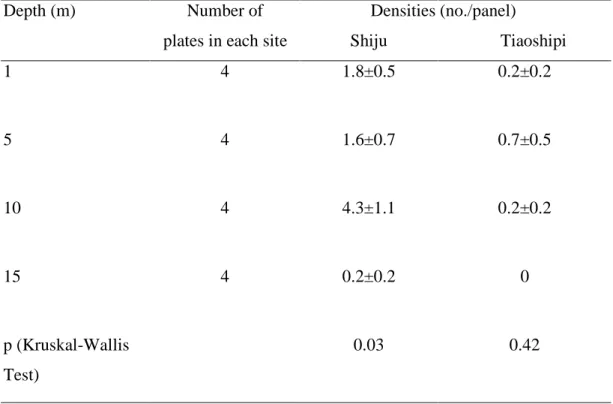
At both of the sites used to test the effect of depth on coral recruitment, 15 m was
found to have the lowest densities. The difference, however, was significant at only one
of the two sites. The other 3 depths, i.e., 1, 5 and 10 m did not differ in coral recruit
densities (Table 3).
In 1997, pocilloporids were the dominant recruits (95%), while acroporids
constituted the rest (5%). In 1998, the only year when data outside the broadcasting
season was used, only pocilloporids were found. In 1999 and 2000, pocilloporids were
still the dominant group (90 and 95%, respectively); in addition acroporids were found in
Penghu, a region in the Taiwan Straits. The total number of coral recruits ranged from
217 in 1999 to 468 in 1997.
In 4 years, only 2 recruits belonging to groups other than pocilloporids or
acroporids were discovered. Densities of recruits were low except at the Hobihu site
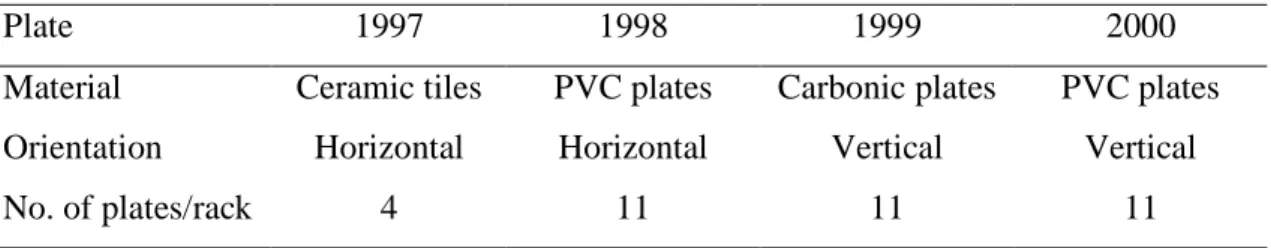
(Site 5 in Hengchun Region, Figure 1) in 1997 and 1998, where more than 10 coral spat
were recorded per panel. Otherwise, the highest averages at a site were all below 3/panel
in 1999 and 2000. The Hobihu site recorded the highest recruitment in the Southern
region of Taiwan (with at least 8 sites each year) for 3 consecutive years. This is
especially pronounced in 1997 and 1998 (Figure 1).
In Southern Taiwan, significant variation occurred at the "Site" level (1 km apart)
in both 1997 and 1998, and at the "Location" level (10 m apart) from 1998 to 2000 (Table
4). “Area” level spatial variation was significant only in 2000.
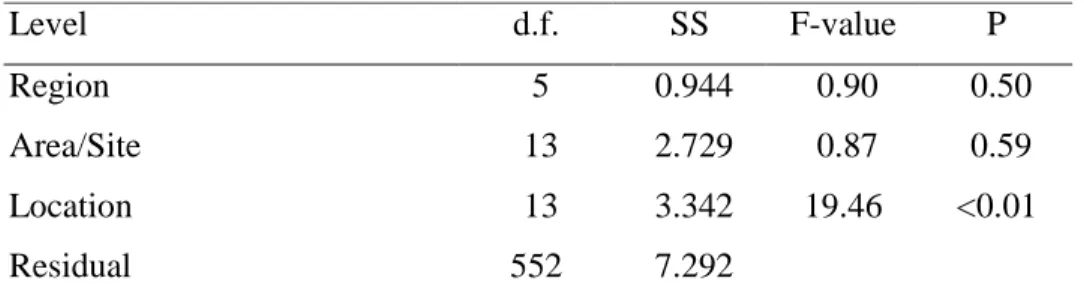
As to a comparison for the whole of Taiwan, significant spatial variation occurred
at the "Location" level in both 1999 and 2000, and at the “Area/Site” level (1-5 km) only
in 2000. No significant difference was found at the “Region” (100 km) level in either
year (Table 5). No coral recruits occurred at Yungan and Shitzwan, the 2 non-reef sites
of the study in 1999 and 2000.
Our preliminary results confirmed that microhabitats make a great difference in the coral
recruitment densities on artificial substrates (Table 2). Most of the factors have been
discussed in great details by earlier investigators (see citations in Introduction). Little
attention has been paid to the number of plates per rack, although it also made a
difference in some situations (when they were oriented vertically in our experiment). We
believe the mechanism may be related to an intensification effect (Pineda and Caswell
1997) in which the abundance of suitable substrate inversely affected the recruit densities.
To exclude the factors of microhabitats, our comparison of spatial patterns has to
be based on plates with same materials, sizes, orientation, depth as well as plate numbers
in the rack. This is accomplished within each year, but variation in set-up existed among
years (Table 1). The inconsistency among years is due to our effort to increase overall
recruit densities on the plates, so that possible patterns were more likely to be detected.
Densities of coral recruits were low in Taiwan from 1999 to 2000. This was
especially pronounced in the broadcasting species and when compared to results in the
GBR (up to 700/panel, see quotes in Harriott 1992). Whereas mass spawnings were
observed by both scientists and amateur divers in the past 10 year period, in Southern
Taiwan (Fan et al. 2001), successful recruitment of broadcasting species seems extremely
limited. Similar phenomena was also observed around the high latitude (30oS) islands
south of the Great Barrier Reefs (e.g., Dunstan and Johnson 1998, Harriott and Banks
1995).
High rates of recruitment may occur several months after mass spawning for
broadcasting species in Bowden Reef, GBR (Babcock 1988). Most planktonic larvae
might have traveled hundreds to thousands of Km away from the reefs producing them
of recruitment for these species (Harriott and Fisk 1988). The results of an independent
study with new plates placed every two months in Southern Taiwan did reveal that new
settlement of spawning corals might occur several months after the mass spawning time
in April-May, but the densities were nevertheless low (Kuo 2001).
The rates of coral recruitment is known to vary greatly among years (e.g.,
Fitzhardinge 1985, Hughes 1985, Hughes et al. 2000, Wallace 1985), thus one cannot rule
out the possibility that a low rate of recruitment is natural in most years for isolated reefs
like those around Taiwan. Occasional recruitment success of broadcasting species may
be enough to maintain the long-lived populations (Veron and Don 1979, Yoshioda 1996);
although an extended period of recruitment failure may adversely affect the coral
communities under threats from various sources (Hughes et al. 2000).
Coral reefs at the southern portion as well as at other regions of Taiwan are under
threats from both natural and anthropogenic sources (Dai et al. 1998). Eutrophication and
high sediment load are known to adversely affect coral recruitment (Babcock and Davies
1991, Harrison and Ward 2001, Hodgson 1990, Nzali et al. 1998, Wittenberg and Hunte
1992). These factors, however, cannot explain the low rates of coral recruitment in all
our study sites, since offshore islands, e.g., Green Island and Orchid Island, are apparently
neither affected by eutrophication nor by excessive sediment loads (personal observation).
Taiwanese coral reefs are, on average, 150 Km from one another (our estimation), thus
isolation may be invoked to explain the domination of recruitment by brooding corals
(see Dunstan and Johnson 1998, Harriott 1992, Harriott and Banks 1995, Kojis and
Quinn 2001). Despite their very low recruitment densities, broadcasting corals dominated
The consistently higher recruits at Hobihu among 8 sites in Southern Taiwan can
be explained 2 ways. The first hypothesis involves physical, or special current conditions
at Hobihu. A model, using surface current data and topographical features of the area,
predicts that there is a cyclonic (anticlockwise) eddy in the west side of the bay (where
Hobihu is) during ebb (Lee et al. 1999). Thus, if coral recruits settle during ebb tide, we
expect to find more recruits on the West than on the east side of the Southern Taiwan
Region. However, the resolution of the model is not fine enough to predict a difference
between the Western and the Northern sites (site 4 and 5 in Figure 1) in this study. A
similar mechanism, i.e., invoking local currents, has been used to explain aggregation
patterns of coral recruits in other studies (Fisk and Harriott 1990, Sammarco and
Andrews 1988). If the above mechanism indeed applies to Southern Taiwan, we would
expect the same aggregation pattern to apply to species of other groups, e.g., barnacles,
which remains to be investigated.
The second hypothesis assumes that most larvae of the brooding pocilloporids do
not travel far after planulation; thus high local recruitment may simply reflects high local
abundance of fecund colonies (Harrison and Wallace 1990). This hypothesis is supported
by the results of a survey in 1998 of existing colonies of Seriatopora hystrix, the
dominant pocilloporid in the region. Hobihu site had the highest number of colonies,
largest mean colony diameters as well as the highest estimated coverage (3 times that of
the second highest site, and more than 10 times that of the rest) of S. hystrix along belt
transects of 20 m2, among the 8 sites investigated in the region (unpublished data). Under
this hypothesis, the dispersal of S. histrix may be limited even within the scale of a
The high variation in coral recruit densities between the years, e.g., that at Hobihu
(Figure 1), was also evident in other studies (Fitzhardinge 1985, Hughes 1985). Besides
the explanation of the yearly variation hypothesis mentioned previously, severe bleaching
occurred in all coral reefs around Taiwan in the summer of 1998 (personal observations).
This may be one of the causes of the reduction in coral recruitment in the following years
(see Michalek-Wagner and Willis 2001).
Significant spatial variation occurred at the “Site” level in 1997 and 1998, and at
the smallest, “Location” level in 3 out of 4 years of the surveys in the Southern Taiwan
region. It seems to suggest that the source of coral recruits is very local. This is
compatible with the hypothesis that most larvae of brooding pocilloporids do not disperse
far upon settlement. Transplantation of fecund colonies to sites with low abundance of
corals may effectively increase local densities of brooding species
Significant variations in various spatial scales are inconsistent over the years in
this study. This differs from that of the GBR in which both brooders and spawners had a
consistent pattern over 2 years (Hughes et al. 1999). In a 4-year study at Heron Island
(GBR), however, spatial pattern of coral recruits were not consistent between years, and
this prompted Dustan and Johnson (1998) to suggest that coral recruitment patterns are
determined by mechanisms that manifest over a large range of spatial scales. Wallace’s
(1985) discovery of reversed rank order of recruitment at compared sites from one year to
the next is compatible with the hypothesis that the dominating factors affecting
recruitment may shift over the years. Since coral recruits in Taiwan are dominated by a
few brooding taxa, spatial variation may be affected by very local events. For example,
dispersal of short-ranged brooding larvae may be highly variable from year to year. The
same mechanism may make little difference on larvae of broadcasting species, since they
have a long planktonic stage. Any small-scale variation tends to even out after a long
period.
Acknowledgements
We would like to thank the numerous volunteer divers who participated in this research.
Two reviewers, who wish to remain anonymous, contributed significantly to the
improvement of the manuscript. The project was sponsored by grants to K. Soong from
the National Science Council of Taiwan, ROC (NSC90-2611-B-110-001) and from the
Taiwan Power Company
References
Babcock R, Davies P (1991) Effects of sedimentation on settlement of Acropora millepora. Coral Reefs 9: 205-508
Babcock R, Mundy C (1996) Coral recruitment: consequences of settlement choice for early growth and survivorship in two scleractinians. J. Exp. Mar. Biol. Ecol. 206: 179-201
Babcock RC (1988) Fine-scale spatial and temporal patterns in coral settlement. Proc. 6th Int. Coral Reef Symp., Australia 2: 635-639
Baird AH, Babcock RC (2000) Morphological differences among three species of newly settled pocilloporid coral recruits. Coral Reefs 19: 179-183
Bak RPM, Engel MS (1979) Distribution, abundance and survival of juvenile hermatypic corals (Scleractinia) and the importance of life history strategies in the parent coral community. Mar. Biol. 54: 341-352
Benayahu Y, Loya Y (1984) Substratum preferences and planulae settling of two Red Sea alcyonaceans: Xenia macrospiculata Gohar and Parerythropodium fulvum fulvum
(Forskal). J. Exp. Mar. Biol. Ecol. 83: 249-261
Birkeland C (1977) The importance of rate of biomass accumulation in early successonal stages of benthic communities to the survival of coral recruits. Proc. 3rd Int. Coral Reef Symp. 1: 15-21
Birkeland C, Rowley D, Randall RH (1981) Coral recruitment patterns at Guam. Proc. 4th Int. Coral Reef Symp., Manila 2: 339-344
Caley MJ, Carr MH, Hixon MA, Hughes TP, Jones GP, Menge BA (1996) Recruitment and the local dynamics of open marine populations. Annu. Rev. Ecol. Syst. 27: 477-500
Connell JH, Hughes TP, Wallace CC (1997) A 30-year study of coral abundance,
recruitment, and disturbance at several scales in space and time. Ecol. Mono. 67: 461-488
Dai CF (1991) Reef environment and coral fauna of Southern Taiwan. Atoll Res. Bull. 436: 1-21
Dai CF, Chen YT, Kuo KM, Chuang CH (1998) Changes of coral communities in Nanwan Bay, Kenting National Park:1987-1997. J. Nat. Park (Taiwan, ROC) 8: 79-99 (in Chinese with English abstract)
Dai CF, Soong K, Fan TY (1992) Sexual reproduction of corals in northern and southern Taiwan. Proc. 7th Coral Reef Symp., Guam 1: 448-455
Duerden JE (1904) The coral Siderastrea radians and its postlarval development.
Carnegie Institute, Washington (1-130)
Dunstan PK, Johnson CR (1998) Spatio-temporal variation in coral recruitment at different scales on Heron Reef, southern Great Barrier Reef. Coral Reefs 17: 71-81
Edwards AJ, Clark S (1998) Coral transplantation: a useful management tool or misguided meddling? Mar. Poll. Bull. 37: 474-487
English S, Wilkinson C, Baker V (1997) Survey manual for tropical marine resources. Australian Institute of Marine Science, Townsville
Fan TY, Guo KM, Guan CS, Tsai YC, Dai CF (2001) Coral spawning in Kenting, Southern Taiwan, a 10-year review. Newsletter of Taiwanese Coral Reef Society (December, 2001), Taipei (1-6 (in Chinese))
Fisk DA, Harriott VJ (1990) Spatial and temporal variation in coral recruitment on the Great Barrier Reef: implications for dispersal hypotheses. Mar. Biol. 107: 485-490 Fitzhardinge R (1985) Spatial and temporal variability in coral recruitment in Kaneohe
Bay (Oahu, Hawaii). Proc. 5th Int. Coral Reef Cong. 4: 373-378
Fitzhardinge R (1986) Spatial variability in the recruitment of corals and other organisms in Kaneohe Bay, Oahu. In: Jokiel PJ, Richmond RH, Rogers CL (eds.) Coral reef population biology. Hawaiian Institute of Marine Science, Report #37 (pp 263-285)
Gains SD, Roughgarden J (1985) Larval settlement rates: a leading determinant of structure in an ecological community of the marine intertidal zone. Proc. Natl. Acad. Sci., USA 82: 3707-3711
Goreau NI, Goreau TJ, Hayes RL (1981) Settling, survivorship and spatial aggregation in planulae and juveniles of the coral Porites porites (Pallas). Bull. Mar. Sci. 31:
424-435
Grigg RW (1988) Recruitment limitation of a deep benthic hard-bottom octocoral population in the Hawaiian Islands. Mar. Ecol. Prog. Ser. 45: 121-126
Harriott VJ (1985) Recruitment patterns of scleractinian corals at Lizard Island, Great Barrier Reef. Proc. 5th Int. Coral Reef Symp., Tahiti 4: 367-372
Harriott VJ (1992) Recruitment patterns of scleractinian corals in an isolated sub-tropical reef system. Coral Reefs 11: 215-219
Harriott VJ, Banks SA (1995) Recruitment of scleractinian corals in the Solitary Islands Marine Reserve, a high latitude coral-dominated community in eastern Australia. Mar. Ecol. Prog. Ser. 123: 155-161
Harriott VJ, Fisk DA (1987) A comparison of settlement plate types for experiments on the recruitment of scleractinian corals. Mar. Ecol. Prog. Ser. 37: 201-208
Harriott VJ, Fisk DA (1988) Recruitment patterns of scleractinian corals: a study of three reefs. Aust. J. Mar. Freshwater Res. 39: 409-416
Harrison PL, Wallace CC (1990) Reproduction, dispersal and recruitment of scleractinian corals. In: Dubinsky Z (ed.) Ecosystem of the world 25, Coral Reefs. Elsevier, New York (pp 133-207)
Harrison PL, Ward S (2001) Elevated levels of nitrogen and phosphorus reduce
fertilization success of gametes from scleractinian reef corals. Coral Reefs 139: 1057-1068
Hodgson G (1990) Sediment and the settlement of larvae of the reef coral Pocillopora damicornis. Coral Reefs 9: 41-43
Hughes TP (1985) Life histories and population dynamics of early successional corals. Proc. 5th Int. Coral Reef Cong., Tahiti 4: 101-106
Hughes TP, Baird AH, Dinsdale EA, Moltschaniwskyj N, Pratchett MS, Tanner JE, Willis B (1999) Patterns of recruitment and abundance of corals along the Great Barrier Reef. Nature 397: 59-63
Hughes TP, Baird AH, Dinsdale EA, Moltschaniwskyj NA, Pratchett MS, Tanner JE, Willis BL (2000) Supply-side ecology works both ways: the link between benthic adults, fecundity, and larval recruits. Ecology 81: 2241-2249
Hughes TP, Tanner JE (2000) Recruitment failure, life histories, and long-term decline of Caribbean corals. Ecology 81: 2250-2263
Kojis BL, Quinn NJ (2001) The importance of regional differences in hard coral
recruitment rates for determining the need for coral restoration. Bull. Mar. Sci. 69: 967-974
Kuo KM (2001) Study on the recruitment, growth and survival of juvenile corals at Nanwan. Institute of Marine Biology. National Sun Yat-sen University, Kaohsiung, Taiwan
Lee HJ, Chao SY, Fan KL (1999) Tide-induced eddies and upwelling in a semienclosed basin: Nanwan. Estuar. Coast. Shelf Sci. 49: 775-787
Leu Y (2001) Adaptation mechanism of eclosion date dimorphism in the marine midge
Pontomyia oceana (Diptera: Chironomidae). Institute of Marine Biology. National
Sun Yat-sen University, Kaohsiung (51)
Maida M, Sammarco PW, Coll JC (1995) Effects of soft corals on scleractinian coral recruitment. I: directional allelopathy and inhibition of settlement. Mar. Ecol. Prog. Ser. 121: 191-202
Michalek-Wagner K, Willis BL (2001) Impacts of bleaching on the soft coral
Lobophytum compactum. I. Fecundity, fertilization and offspring viability. Coral
Reefs 19: 231-239
Miller MW, Weil E, Szmant AM (2000) Coral recruitment and juvenile mortality as structuring factors for reef benthic communities in Biscayne National Park, USA. Coral Reefs 19: 115-123
Mundy CN (2000) An appraisal of methods used in coral recruitment studies. Coral Reefs 19: 124-131
Nzali LM, Johnstone RW, Mgaya YD (1998) Factors affecting scleractinian coral recruitment on a nearshore reef in Tanzania. Ambio 27: 717-722
Pineda J, Caswell H (1997) Dependence of settlement rate on suitable substrate area. Mar. Biol. 129: 541-548
Randall RH, Cheng Y-M (1977) Recent corals of Taiwan. Part I. Description of reefs and coral environments. Acta Geol. Taiwanica 19: 79-102
Randall RH, Cheng Y-M (1979) Recent corals of Taiwan. Part II. Description of reefs and coral environments. Acta Geol. Taiwanica 22: 1-32
Rogers CS, Fitz HCI, Gilnack M, Beets J, Hardin J (1984) Scleractinian coral recruitment patterns at Salt River submarine canyon, St. Croix, U.S. Virgin Islands. Coral Reefs 3: 69-76
Roughgarden J, Gaines S, Possingham H (1988) Recruitment dynamics in complex life cycles. Science 241: 1460-1466
Rylaarsdam KW (1983) Life histories and abundance patterns of colonial corals on Jamaican reefs. Mar. Ecol. Prog. Ser. 13: 249-260
Sammarco PW (1985) The Great Barrier Reef vs. the Caribbean: comparisons of grazers, coral recruitment patterns and reef recovery. Proc. 5th Int. Coral Reef Cong. 4: 391-397
Sammarco PW, Andrews JC (1988) Localized dispersal and recruitment in Great Barrier Reef corals: the helix experiment. Science 239: 1422-1424
Scheltema RS (1986) On dispersal and planktonic larvae of benthic invertebrates: an eclectic overview and summary of problems. Bull. Mar. Sci. 39: 290-322 Stephenson TA (1931) Development and the formation of colonies in Pocillopora and
Porites. Part I. Sci. Rep. Great Barrier Reef Exped. 1928-29 3: 113-134
Sutherland JP (1990) Recruitment regulates demographic variation in a tropical intertidal barnacle. Ecology 71: 955-972
Underwood AJ, Chapman MG (1996) Scales of spatial patterns of distribution of intertidal invertebrates. Oecologia 107: 212-224
Veron JEN, Don TJ (1979) Corals and coral communities of Lord Howe Island. Aust. J. Mar. Freshwater Res. 30: 1-34
Wallace CC (1985) Seasonal peaks and annual fluctuations in recruitment of juvenile scleractinian corals. Mar. Ecol. Prog. Ser. 21: 289-298
Williams MD, Wolanski E, Andrews JC (1984) Transport mechanisms and the otential movement of planktonic larvae in the central region of the Great Barrier Reef. Coral Reefs 3: 229-236
Wittenberg M, Hunte W (1992) Effects of eutrophication and sedimentation on juvenile corals. I. Abundance, mortality and community structure. Mar. Biol. 112: 131-138 Yoshioda PM (1996) Variable recruitment and its effects on the population and
Figure Legend
Figure 1. Recruitment density (No./plate surface) of pocilloporids in Taiwan. The settlement plates were recovered at sites 1, 2, 4, 5, 6, 7, 9, 10 in 1997 and 1998; at sites 2, 3, 4, 5, 6, 7, 9, 11 in 1999; and at sites 1, 3, 4, 5, 6, 8, 9 in 2000. Site labels on the X-axis were deleted for those sites with no plates retrieved.
1 2 4 5 6 7 9 1 0 0 1 0 2 0 3 0 4 0 5 0 6 0 7 0 8 0 9 0 100 1997 1 0 2 0 3 0 4 0 5 0 1998 2 3 4 5 6 7 9 1 1 0.00 0.25 0.50 0.75 1.00 1.25 1.50 2 3 4 5 6 7 9 1 1 0.00 0.25 0.50 0.75 1.00 1.25 1.50 1999 0.50 1.00 1.50 2.00 2.50 2000 1 2 0.00 0.25 0.50 0.75 1.00 1.25 1.50 1 2 0.00 0.25 0.50 0.75 1.00 1.25 1.50 1999 1 2 3 4 0.00 0.25 0.50 0.75 1.00 1.25 1.50 1999 1 2 0.00 0.25 0.50 0.75 1.00 1.25 1.50 1999 1 2 0.00 0.50 1.00 1.50 1999 1 2 0.00 0.25 0.50 0.75 1.00 1.25 1.50 1999 Hengchun 1 2 0.00 0.50 1.00 1.50 2.00 2.50 1 2 0.00 0.50 1.00 1.50 2.00 2.50 1 2 0.00 0.50 1.00 1.50 2.00 2.50 2000 Yungan Penghu Hsiaoliuchiu 1 2 0.00 0.50 1.00 1.50 2.00 2.50 2000 1 2 0.00 0.50 1.00 1.50 2.00 2.50 1 2 0.00 0.50 1.00 1.50 2.00 2.50 2000 Shitzwan 1 2 0.00 0.50 1.00 1.50 2.00 2.50 2000 1 2 0.00 0.50 1.00 1.50 2.00 2.50 2000 1 2 0.00 0.50 1.00 1.50 2.00 2.50 1 2 0.00 0.50 1.00 1.50 2.00 2.50 2000 Orchid Island Green Island Northeast 1 2 3 4 0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 2000 1 2 0.00 0.50 1.00 1.50 2.00 2.50 2000 2000 1 2 0.00 0.50 1.00 1.50 2.00 2.50 1 2 0.00 0.50 1.00 1.50 2.00 2.50 122 24 22 * Yungan * Shitzwan Green Island Orchid Island Hengchun Hsiaoliuchiu Northeast*
Table 1 The set up of plates and racks in each year.
Plate 1997 1998 1999 2000
Material Ceramic tiles PVC plates Carbonic plates PVC plates
Orientation Horizontal Horizontal Vertical Vertical
Table 2 Summary of preliminary comparisons of settlement densities of coral recruits
Comparison Average density
(number/panel) ±SE Number of plates used Statistics Vertical/Horizontal 38.0±2.2/25.0±5.0 44 p<0.01, MWU Lower/Upper 20.5±3.8/2.7±1.2 22 p<0.01, WSR Ceramic/PVC 58.3±9.7/39.8±7.4 58 p=0.08, MWU 2-plate/11-plate (Horizontal) 9.6±4.9/5.4±1.5 53 p=0.36, MWU 2-plate/11-plate (Vertical) 54.7±4.0/38±2.6 44 p<0.01, MWU Facing outward/ Facing each other
2.7±1.2/43.9±2.1 44 p<0.01, WSR
Table 3 Average settlement densities of pocilloporids on the lower surfaces of plates at different depths at two sites
Depth (m) Number of
plates in each site
Densities Shiju (no./panel) Tiaoshipi 1 4 1.8±0.5 0.2±0.2 5 4 1.6±0.7 0.7±0.5 10 4 4.3±1.1 0.2±0.2 15 4 0.2±0.2 0 p (Kruskal-Wallis Test) 0.03 0.42
Table 4. Nested-ANOVA table of pocilloporid recruitment in Hengchun Penninsula, Southern Taiwan. Scale of Area: 2-5 km, Site: 1 km, Location: 10 m. All the data was transformed by using log(n+1).
(a) 1997/4/10-1997/5/23 Levels d.f. SS F-value p Area 3 2.696 0.83 0.54 Site 4 4.294 9.67 <0.01 Location 8 0.889 1.82 0.10 Residual 49 2.996 (b) 1998/1/14-1998/2/28 Levels d.f. SS F-value p Area 3 10.187 0.97 0.49 Site 4 13.972 40.15 <0.01 Location 8 0.700 2.81 <0.01 Residual 160 4.974 (c) 1999/4/22-1999/6/27 Levels d.f. SS F-value p Area 3 0.131 0.76 0.57 Site 4 0.235 0.66 0.65 Location 5 0.436 8.30 <0.01 Residual 209 2.192 (d) 2000/4/2-2000/8/8 Level d.f. SS F-value p Area 3 7.217 63.32 <0.01 Site 4 0.154 0.43 0.78 Location 5 0.438 3.32 <0.01
Table 5. Nested-ANOVA table of pocilloporid recruitment in Taiwan. Scale of Region: about 100 km, Area/Site: 1-5 km, Location: 10 m
(1) 1999/4/22-1999/6/27 Level d.f. SS F-value P Region 5 0.944 0.90 0.50 Area/Site 13 2.729 0.87 0.59 Location 13 3.342 19.46 <0.01 Residual 552 7.292 (2) 2000/4/2-2000/8/8 Level d.f. SS F-value P Region 7 2.624 0.40 0.89 Area/Site 15 14.030 11.08 <0.01 Location 17 1.435 3.97 <0.01 Residual 760 16.135