遺傳歧異度對小米地方品系滲透逆境耐受性的影響
陳昶霖
1、張庭禎
2、張祐菱
2、林維怡
2*
1國立中興大學園藝學系 2國立臺灣大學農藝學系摘要
水份逆境對作物產量影響甚鉅,提高耐 受性已是重要育種目標。小米 (Setaria italica L.) 為小型穀類作物,屬於水份利用效率較高 的C4 植物,但遺傳歧異度對小米逆境耐受性 的影響目前了解有限。本研究以 158 個來自 臺灣及印度種原庫的小米地方品系為材料, 以25 % PEG 6000 的水耕系統模擬滲透逆境, 檢測這些品系逆境耐受性的差異。結果顯示, 所有品系在逆境處理 7 天後,株高和鮮重都 顯著低於控制組,從中挑選耐受性較高、中等 和較低的品系各10 個,測試逆境回復能力, 以及表型的關聯性,發現地上部和地下部的 生長勢與逆境耐受性呈負相關,進一步檢測 耐受性較高和較低的品系的氧化逆境指數, 與其他測試品系相比,品系 110 的抗氧化酵 素活性並未隨著逆境時間延長,甚至降低,葉 表面溫度也沒有明顯升高,顯示該品系的耐 受性較高。本研究揭露了遺傳歧異度對小米 滲透逆境耐受性的影響,也鑑定出耐受性較 高的品系,可作為未來育種和遺傳背景相關 研究之參考。 關鍵詞︰小米、滲透逆境、遺傳歧異度、逆境 耐受性、抗氧化酵素。The Impacts of Genetic Variation on
the Osmotic Stress Tolerance of
Foxtail Millet Landraces
Chang-Lin Chen1, Ting-Chen Chang2, Yu-Ling Chang2, Wei-Yi Lin2*
1 Department of Horticulture, National Chung
Hsing University, Taichung 402, Taiwan, ROC
2 Department of Agronomy, National Taiwan
University, Taipei 106, Taiwan, ROC
ABSTRACT
The crop production was threatened by various kinds of stress, including water stress. Enhancing the stress tolerance is one of main breeding objectives. Foxtail millet (Setaria italica L.), a C4 crop species, has relatively high water use efficiency. But little is known about the impacts of genetic variation on millet stress tolerance. In this study, the hydroponic culture system containing 25 % polyethylene glycol (PEG 6000) was used to mimic the osmotic stress conditions. We used 158 millet landraces derived from Taiwan and the seed stock center in India as materials to study the variation of stress tolerance under osmotic stress conditions. The results showed that after 7-day treatment the plant height and fresh weight were significantly lower than those under normal conditions. Thirty landraces with high, medium or low stress tolerance were selected to test their ability of stress recovery. The correlation analysis suggested that the fresh weight and root length is negatively correlated with stress tolerance. Furthermore, we analyzed the activity of antioxidative enzymes in the landraces with high or low stress tolerance and found that the enzyme activity in landrace 110 was maintained at similar level or even decrease during the treatment. Its leaf surface temperature was also comparable to plants grown under normal conditions. It suggests that
*通信作者, [email protected] 投稿日期:2020 年 7 月 17 日 接受日期:2020 年 9 月 10 日
作物、環境與生物資訊 16:209-220 (2019) Crop, Environment & Bioinformatics 16:209-220 (2019) DOI: 10.30061/CEB.201912_16(4).0003
this line has higher stress tolerance. Our study revealed the effects of genetic variation on the osmotic stress tolerance of millets. We also identified the landraces with high tolerance. It provides useful information for breeding and further studies about the association of genetic background with millet stress tolerance.
Keywords: Foxtail millet, Osmotic stress, Genetic
variation, Stress tolerance, Antioxidative enzyme.
前言
由於全球暖化的效應,使得各地降雨型 態改變,導致乾旱或淹水逆境發生頻率和強 度增加,著實威脅人類的糧食安全,其中乾旱 已 是 主 要 限 制 作 物 生 產 的 因 子 (Zhao and Running 2010, Lobell et al. 2011)。受到乾旱 逆境時,植物無法吸取足夠水份,氣孔因而關 閉以減少水份散失,但同時光合作用受到抑 制,無法接收游離電子,導致活性氧族 (ROS) 的產生而造成氧化逆境,破壞細胞結構與生 理反應 (Shanker et al. 2014);依乾旱嚴重情 形及逆境持續的時間對作物生長及產量造成 程度不一的影響,統計 1980 至 2015 年乾旱 相關田間研究成果發現,小麥和玉米產量分 別下降21%和 40% (Daryanto et al. 2016)。以 二 氧 化 碳 的 代 表 濃 度 路 徑 (representative concentration pathways) 為 8.5 來預估水 稻、大豆、小麥和玉米等產量,會下降約 8-24% (Elliott et al. 2014),同時人口壓力持續 增加,預估至2050 年全球作物生產需增加 60-110%才足以應付糧食需求 (FAO 2009, Ray et al. 2013),如何在乾旱逆境挑戰下維持甚至 提升產量為重要的研究目標之一。 生長於乾旱地區的植物已演化出許多應 付逆境的機制,例如:地上和地下部的形態改 變以減少水份散失及增加水份吸收效率,增 加滲透物質的累積以提高細胞滲透勢,提升 抗 氧 化 能 力 以 降 低 氧 化 逆 境 之 傷 害 等(Kar 2011, Shanker et al. 2014) 。 小 米 (Setariaitalica L.;foxtail millet) 起源於中國北方,
屬於禾本科中的黍亞科,與玉米和高粱同為
C4 植物,由於小米具有相對較厚的細胞壁和 較小的葉表面積,且水份利用效率高於玉米 和 高 粱 (Shantz and Piemeisel 1927, Koch 2002),因此可生長於乾旱及半乾旱地區。此 外,小米為2 倍體作物,基因體約 490 Mb, 比高粱和玉米等同亞科的旱作作物小,且已 完 成 基 因 體 定 序(Bennetzen et al. 2012, Zhang et al. 2012),因此被視為研究乾旱逆境 的新興模式作物。乾旱逆境反應的分子調控 機制已在許多植物中進行深入探討,透過同 源基因的檢測和分析也鑑定出小米基因體中 逆境相關的基因,如:SiNAC110、SiDREB2、 SiWD40、SiPDLα1 等藉由參與 ABA 相關或 非 ABA 相關的途徑以調控小米在乾旱和其
他逆境的耐受性 (Peng et al. 2010, Lata et al. 2011, Mishra et al. 2014, Xie et al. 2017)。而 受惠於基因體定序之完成,後續利用轉錄體 和蛋白質體分析更全面地揭露調控小米乾旱 反 應 及 逆 境 適 應 性 的 分 子 網 絡(Tang et al. 2017, Pan et al. 2018),對於耐旱育種提供相 當寶貴的資訊。然而上述之研究大多以少數 品系為材料,容易忽略基因多型性對生理性 狀的影響力;Lata 等人 (2011) 藉由檢測 45 個核心品系在乾旱逆境中脂質氧化和相對水 份含量的變化與SiDREB2 基因單一核苷酸多
型性 (single nucleotide polymorphism) 的 關聯性,顯示遺傳背景歧異度對生理性狀的 影響力。 由於臺灣原住民在小米栽培及種原保留 扮演重要角色,根據各族的生活環境、栽培管 理技術及文化傳統之差異,長期篩選適應當 地環境且符合需求之小米,因此造就多樣化 的小米地方品系;以DNA 分子標誌分析 324 個臺灣小米地方品系的33 個農藝性狀,可發 現這些地方品系的遺傳歧異度高,且可依地 理分布將之分為三大群(Lin et al. 2012),是為 研究小米生理性狀及育種的極佳材料。 為彌補過去對於遺傳歧異度與小米耐旱 性 相 關 知 識 的 不 足 , 本 研 究 利 用 聚 乙 二 醇 6000 (polyethylene glycol 6000;PEG 6000)
模擬滲透逆境,調查 158 個來自臺灣各地及 印度小米種原中心的地方品系於非逆境及逆 境環境下的地上部性狀,從中挑選耐受性高、 中及低的品系進行更進一步的探究,以了解 各品系的逆境適應能力,作為未來育種發展 之參考。
材料與方法
一、 小米水耕栽培與管理 本研究使用之品系中有47 個來自印度種 原中心的品系、108 個來自臺灣各地的地方品 系,以及由臺東區農業改良場選育之臺東選2 號、臺東選 6 號和臺東 8 號,共 158 種。臺 灣地方品系是從台東縣、屏東縣、高雄市、宜 蘭縣及嘉義縣等地分離純化而來,在第一次 試驗中挑選逆境耐性低、中及高的品系各 10 個進行第二次重複試驗,檢測逆境回復能力, 第三次試驗則從中挑選逆境耐性低及高品系 各5 個進行性狀調查及氧化逆境的檢測。 本研究以水耕系統栽培小米;首先將種 子以45℃處理 30 分鐘,再以 2%次氯酸鈉進 行表面消毒30 秒,最後以無菌水淋洗數次即 可進行發芽;萌芽及生長環境溫度為日溫 28 ℃/夜溫 22℃,光照時數及強度分別為 12 小 時/ 150-200 µmol photon m-2s-1,萌芽後3 天 (3 DAG) 定植於水耕泡棉,於 7 DAG 每個 品系移植 6 株幼苗至水耕系統,依序以 0.5 和1 1/2 Kimura 水耕液培養 3 天,最後改 以1.5的水耕液培養至材料收穫。 二、 滲透逆境處理及性狀調查 於21 DAG 開始每個品系選取 3 棵植株 處理25% PEG 6000,其餘 3 株則維持在原水 耕液生長,是為控制組,處理 7 天後量測性 狀,第一次試驗檢測地上部株高和鮮重,處理 期間每二天記錄萎凋狀況,萎凋指數判斷是 參 考 國 際 水 稻 研 究 所 (International Rice Research Institute;IRRI) 的評估系統 (IRRI 2013);第二次 PEG 6000 處理方法如前述, 每個品系的植株數量在處理和控制組都增加 至6 株,處理 7 天後換回 1.5水耕液以檢測 植株復原能力,水勢回復 4 天後收穫,除了 上述之性狀,另量測根長及根鮮重;第三次試 驗則是收穫處理1 和 4 天的植株,處理和控 制組在每個時間點各收穫 4 株,本次試驗除 了上述之性狀,另量測葉溫,並收穫葉片供氧 化逆境指標之量測。 三、 葉表面溫度量測 第三次試驗以 InfReC 熱感相機分別拍 攝小米地上部的部分,每個品系在處理和控 制組各拍攝 4 株,影像以 InfReC Analyzer NS9500 軟體分析葉表面溫度,每株圈選所有 葉片進行分析並計算平均值,作為該植株的 平均葉表面溫度。 四、 抗氧化酵素活性量測 本試驗共檢測三種抗氧化酵素活性,包 括過氧化氫酶 (catalase;CAT)、抗壞血酸過 氧化酶 (ascorbate peroxidase;APX) 及穀 胱甘肽還原酶 (glutathione reductase;GR); 簡單說明測量方式如下:先將約100 mg 樣品 以 液 態 氮 研 磨 成 細 粉 , 加 入 2 ml 50 mM sodium phosphate buffer (pH 7) 進行萃取,於4℃以 12000 g 離心 20 分鐘,吸取上清液 以量測酵素活性。CAT 活性檢測參考 Kato 和 Shimizu 的方法 (1987),取 0.2 mL 萃取液, 加 入 2.7 mL 100 mM sodium phosphate buffer (pH 6.8) 和 0.1 mL 1 M H2O2,混勻 後以 OD240 量測 1 分鐘;APX 活性檢測參 考Nakano 和 Asada 的方法 (1981),取 0.1 mL 萃取液,加入 1 mL 150 mM potassium phosphate buffer (pH 7)、1 mL ascorbate、 0.4 mL 0.75 mM EDTA 和 0.5 mL 6 mM H2O2,混勻後以OD290 量測 1 分鐘;GR 活 性 檢 測 方 法 參 考 Foster 和 Hess 的 方 法 (1980),取 0.2 mL 萃取液,加入 1 mL 150 mM Tris-HCl (pH 7.5) 、 0.3 mL 30 mM MgCl2、0.5 mL 3 mM glutathione disulfide 和1 mL 0.45 mM NADPH,混勻後以 OD340
量測 1 分鐘。另檢測樣品蛋白質濃度作為酵 素活性標準化之依據,蛋白質濃度量測採用 Bradford assay (Bradford 1976),取 10 μL 萃
取液,加入240 μL 染劑,混勻後靜置 10 分
鐘,以OD595 量測樣品及標準品吸光值。
五、 丙 二 醛 (malondialdehyde ; MDA) 含量測定
MDA 的量測根據 Heath 和 Packer 的方
法 (1968),簡述如下:將約 100 mg 樣品以液
態氮研磨成細粉,以2 mL 5% trichloroacetic acid (TCA) 進行萃取,再於 20℃以 10000 g 離心5 分鐘,取 1 mL 上清液,加入 4 mL 0.5 % thiobarbituric acid (in 20% TCA),於 95
℃水浴30 分鐘後,將反應溶液立即置於冰上 以終止反應,以OD532 和 OD600 量測樣品 吸光值。 六、 統計分析方法 使用R 程式 (v3.6.1) 進行統計分析,常 態分布檢驗以Shapiro-Wilk 檢定進行,以相 關矩陣 (correlation matrix) 分析性狀間的 關聯性,各品系性狀之差異以ANOVA 分析, 再作Scheffe 事後檢定。
結果
一、 158 個小米地方品系於滲透逆境之 表現 為了解小米地方品系於滲透逆境下的表 現差異,我們以水耕系統栽培 158 個來自台 灣各地及印度種原中心的地方品系,於發芽3 週後將一半的植株移到含25 % PEG 6000 水 耕液處理1 週,其餘則維持 1.5水耕液培養, 作為控制組,調查二組樣品的株高、地上部鮮 重和萎凋指數,以Shapiro-Wilk 檢定分析族 群性狀,二組樣品的株高皆為常態分布,而且 處 理 組 的 株 高 分 布 波 峰 明 顯 小 於 控 制 組 (Fig. 1),顯示 PEG 6000 處理造成之逆境影 響小米的生長,而地上部鮮重的數值雖然不 是常態分布,但仍可見整體族群的鮮重分布 在PEG 6000 處理後有下降的趨勢,另計算處 理組和控制組的株高及鮮重比例作為評估耐 旱性的指標,族群中多數品系在逆境和正常 狀況下的株高比值為 0.8-0.9 者為最多 (Fig. 1C),顯示多數小米品系的株高生長對逆境反 應較小,而地上部鮮重的比值則集中在 0.5-0.7 (Fig. 2C),顯示鮮重對滲透逆境較為敏感。 根據各品系株高和鮮重在逆境下的變化以及 處理 1 週後葉片的萎凋指數,選取對滲透逆 境敏感程度較高、中等及較低的品系各10 個 進行第二次試驗。 二、 地上和地下部性狀與滲透逆境耐受 性之關係 植物的逆境耐受性除了是指在逆境發生 期間生長和生理功能的維持,也包括逆境結 束後的回復能力,為了解這些地方品系於滲 透逆境的回復能力,選用30 個耐受性不同的 地方品系進行試驗,在PEG 6000 水耕液處理 1 週後將植株移回正常滲透壓的 1.5水耕液 培養4 天,除了量測逆境處理 7 天和滲透壓 回復 4 天後的植株株高和萎凋指數,並於回 復 4 天時收穫植株以量測地上部鮮重、根長 及根鮮重。在植株鮮重部分,可發現逆境耐受 性較高的品系如:110、144、19、33、152 等 在逆境及復水處理後的鮮重與無處理的控制 組相比僅減少10-20% (Fig. 3A and B);反之, 逆境耐受性較低的品系如:100、209 和 219 在處理後鮮重則是較控制組低 40-50 % (Fig. 3A and B);在萎凋指數部分,耐受性較高的 品系在滲透逆境處理 7 天後及復水 4 天後萎 凋指數皆低於耐受性較低者,比較二個時間 點的萎凋指數以了解逆境回復能力,耐受性 較高者也展現較高的回復力 (Fig. 3C);進一 步分析性狀間的相關性,結果發現處理組和 控制組的鮮重差異比值 (osmotic/CK SFW & osmotic/CK RFW) 與 萎 凋 指 數 (osmotic_withering) 呈 顯 著 負 相 關 (Fig. 4),說明鮮重差異比值確實能反應逆境耐受 性,有趣的是,控制組的株高 (CK_height)Fig. 1. The distribution of plant height among 158 millet landraces under (A) normal and (B) osmotic stress conditions. (C) The ratio of plant height under osmotic and normal condition was calculated as the indicator of stress tolerance. Three seedlings were harvested for phenotype investigation. The Shapiro-Wilk test for normality was performed and the p-value was shown.
Fig. 2. The distribution of shoot fresh weight among 158 millet landraces under (A) normal and (B) osmotic stress conditions. (C) The ratio of shoot fresh weight under osmotic and normal condition was calculated as the indicator of stress tolerance. Three seedlings were harvested for phenotype investigation. The Shapiro-Wilk test for normality was performed and the p-value was shown.
Fig. 3. (A) The shoot and (B) root fresh weight of 30 millet landraces under normal (CK) and osmotic conditions. The ratio of fresh weight under normal conditions to osmotic conditions was calculated. (C) The wilting score of plants was evaluated at 168 hours (hr) after osmotic treatment and at 96 hr after rewatering and the ratio was calculated. Six plants were harvested and data was presented as mean ± SD.
和鮮重 (CK_SFW 和 CK_RFW) 與萎凋指
數呈正相關 (Fig. 4),顯示逆境耐受性較高者
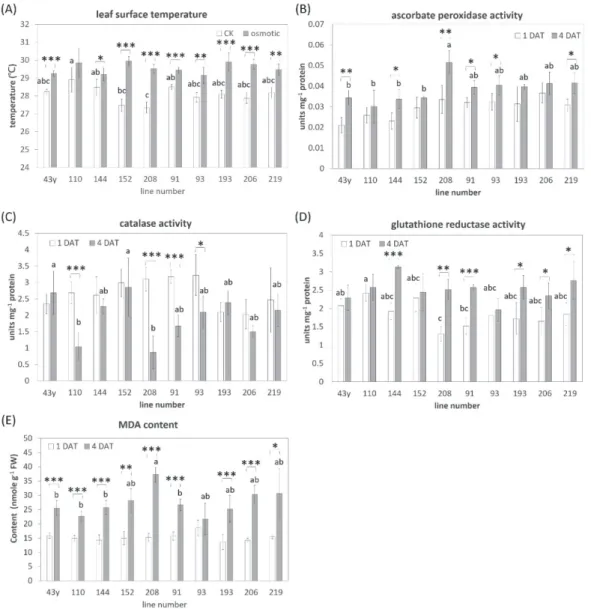
在正常狀況下生長勢較耐受性低者弱,而處 理 組 和 控 制 組 的 根 長 (CK_root length & osmotic_root length) 與萎凋指數呈正相關, 但與鮮重差異比值呈負相關 (Fig. 4),暗示著 根長度影響滲透逆境耐受性。 三、滲透逆境下抗氧化指數之變化 為進一步探究影響滲透逆境耐受性的原 因,挑選耐受性較佳的品系:43y、110、144、 152 和 208,以及較差的品系:91、93、193、 206 和 219,共 10 個進行試驗,將 21 天大的 植株以25 % PEG 6000 處理 4 天,以熱影像 儀量測控制組和處理組的葉表面溫度,另外 於處理後第 1 天和第 4 天收穫葉片以量測抗 氧化酵素活性及 MDA 含量。在控制組中品 系 208 葉表面溫度最低而品系 110 為最高, 然 而 在 滲 透 逆 境 下 各 品 系 間 則 無 明 顯 差 異 (Fig. 5A);比較各品系在控制組和處理組的 差異,品系110 沒有顯著差異,而品系 144 有 些微差異,推測 4 天的滲透逆境處理對這二 個品系的氣孔開闔影響較其他品系輕微。
Fig. 4. The correlation matrix of phenotype derived from 30 millet landraces under normal (CK) and osmotic stress conditions.
Six seedlings were harvested for phenotype investigation. The value in the circle is the correlation coefficient (p-value < 0.01). Osmotic_withering and Osmotic_height168 stand for the wilting score and plant height investigated after 7-day PEG 6000 treatment, respectively. Osmotic_height_RW is the plant height investigated after 4-day recovery. SFW, RFW and root length are the shoot and root fresh weight and root length investigated after 4-day recovery, respectively. Osmotic/CK is the ratio of seedling fresh weight under normal condition to those after 4-day recovery from osmotic stress.
Fig. 5. (A)The surface temperature, (B-D) antioxidant enzyme activity and (E) MDA content in the leaves of selected millet landraces. The data was presented as mean ± SD (n=4). DAT, days after treatment. Different characters were shown the significant difference among lines by ANOVA at p < 0.05. Student’s t test was performed to show the difference of samples under different treatments (A) or at different time points (B-E). *, P < 0.05; **, P < 0.01; ***, P < 0.001. 抗氧化酵素方面,我們量測APX、CAT 和 GR 活性以比較品系及處理時間長短的影 響;在APX 活性方面,除了品系 206,酵素 活性在逆境處理1 天後品系間沒有顯著差異, 但在處理 4 天後有 6 個品系的酵素活性顯著 提升,以品系208 的活性最高 (Fig. 5B),說 明隨著逆境處理時間增長需提高APX 活性以 維持細胞功能;在CAT 活性上,在逆境處理 1 天時品系間亦無顯著差異,但逆境處理 4 天 後,品系110、208、91 和 93 皆顯著下降 (Fig. 5C),其餘品系則維持相似程度的活性,顯示 這 4 個品系在處理 4 天後可能不需要較高的 CAT 活性來減低逆境傷害;在 GR 活性上, 在處理 1 天時已觀察到品系間的差異,以品
系 208 為最低,品系 110 為最高,但在處理 4 天時品系 144、208、91、193、206 和 219 的酵素活性皆顯著較處理 1 天時高,其中以 品系144 的活性為最高 (Fig. 5D)。 MDA 方面,在逆境處理一天後 MDA 含 量在各品系間並無顯著差異,而處理4 天後, 除了品系 93,其餘品系的含量都顯著增加 (Fig. 5E),說明氧化逆境造成之傷害隨時間而 增加,其中以品系 208 為最高,可能該品系 所受到之傷害較多,雖然該品系的CAT 活性 在處理4 天後下降,但 APX 和 GR 活性皆顯 著上升,尤其APX 活性遠高於其他品系,顯 示該品系確實需要較高的抗氧化酵素活性, 以減輕逆境帶來之傷害。
討論
小米為小型穀粒作物,雖不是主要糧食 作物,但由於具有相對較高的逆境耐受性,在 氣候變遷導致逆境頻度和強度加劇的時代, 小米生理和分子層面的研究逐漸受到重視, 然這些研究往往只選用一或二個栽培種來探 討小米的生理機制,忽略了遺傳歧異度的影 響力。本研究以 25% PEG 6000 處理來模擬 滲透逆境,選用 158 個來自臺灣及印度種原 中心的品系進行逆境耐受性的檢測,藉此了 解遺傳歧異度對耐受性的效應,以及各種生 理性狀或抗氧化能力與耐受性的關聯性。 遺傳歧異度決定物種的外表型和各種生 理特性,受惠於分子標記的開發及次世代定 序等技術的發展,物種內遺傳歧異度對表型 和逆境耐受性的相關研究蓬勃發展,例如:在 20% PEG 6000 處理下 645 個小麥地方品系的 株高、鮮重、根長和含水量等表型皆呈現不同 程 度 的 抑 制 , 後 續 應 用 genome-wide association study 鑑定出 57 個相關的數量性 狀基因座(Lin et al. 2019),顯示遺傳背景的差 異在逆境耐受性上扮演決定性的角色,透過 相關研究不僅有助於了解基因的功能,也提 供重要資訊作為未來育種之參考;本研究探 討158 個小米地方品系在 25 % PEG 6000 逆 境處理和非逆境環境下的株高、地上部鮮重 和萎凋指數,發現 7 天的逆境處理下整個族 群中的株高和鮮重都降低,降幅分別集中在 10-30 %和 30-50 % (Fig. 1 and 2),雖然這 個族群數量僅有 158,且約 3/4 地方品系皆 來自臺灣,但遺傳背景的差異仍使我們觀察 到不同程度的生長抑制,此外,族群表型分布 亦屬於常態分布,顯示本研究所使用之族群 的遺傳歧異程度應足以用於鑑定數量性狀基 因座和品種選育。 根是負責吸收水份和養份的主要器官, 當環境中發生輕度到中度乾旱時,根的形態 會重新調整以增加水份吸收效率:增加側根 數、根毛密度和長度以提高吸收表面積,根也 會延長至土壤深處吸取水份,耐旱品種的馬 鈴 薯 根 較 不 耐 旱 品 種 長 (Boguszewska-Mankowska et al. 2020),耐旱能力較佳的冬 小麥根長約為春小麥 1.5 倍(Djanaguiraman et al. 2019),深根系統也對於水稻等其他植物 擷取土壤深處的水份極為重要(Ho et al. 2005, Hund et al. 2009, Henry et al. 2012),而小米 可在乾旱地區栽培,本身已具備耐旱特性,包 括較厚的細胞壁、高密度的根系和較小葉面 積,在乾旱逆境時,根毛的密度和長度在土壤 深度30-35 公分處達最高,而在土表為最低, 相較於土表,30-35 公分深的土壤水含量達最 高(Liu et al. 2019),說明小米的根在乾旱時也 傾向往土壤深處擷取水源,一旦在特定深度 獲取相當水份,其餘區域根毛的發展程度相 對緩慢,顯示在乾旱逆境下,植物會將能量投 注在有利於生存的部分,如:區域性的根毛發 展以增加水分吸收效率;本研究檢測30 個耐 旱程度不一的品系在控制組和滲透逆境處理 組的表型,並計算表型間的相關性,結果發現 根長分別與萎凋指數和鮮重差異比值呈正相 關和負相關 (Fig. 4),顯示這個小米族群中根 愈長者滲透逆境的耐受性愈差,與前述其他 作物的適應策略相反,不同於土壤栽培,在土 壞的垂直面上會建立水份梯度,本試驗採用 水耕系統,PEG 6000 滲透逆境處理下每個區域的根皆接受到相似的逆境刺激,根系較長 者的水份吸收效率並不會較高,且可能需要 較高能量以維持較長根系的細胞活性,因此 推測根系較短者可將能量保留於其他更有利 於生存的策略,相似的推論可衍伸至小米生 長勢與逆境耐受性,控制組的植株株高和鮮 重皆與萎凋指數呈正相關,與處理組/控制 組鮮重比值呈負相關 (Fig. 4),顯示植株生長 勢愈佳者在遭遇滲透逆境後耐受性愈差,可 能因為這些植株需要相對較多能量以維持逆 境下的生長,但在逆境下無法獲取足夠能量 而展現逆境不適應表癥,呼應「平衡生長」 (the balanced growth) 理 論 (Bloom et al. 1985),在滲透逆境下,如何將能量有效率地 分配以維持生長是決定耐受性的關鍵。 在缺水逆境下植物發展出許多生理和生 化反應,在地上部由於水份吸收受阻,氣孔導 度下降以減少蒸散,葉表面溫度常因此而上 升,在本研究中品系 110 和 142 的葉表面溫 度在逆境處理 4 天後幾乎沒有增加,而其他 品系則有顯著提升 (Fig. 5A),暗示這二個品 系的氣孔開闔尚未受到逆境影響,光合作用 效率可能得以維持,但仍需進一步檢測加以 確認;葉片氣孔導度下降除了降低光合作用 效率,也會使游離電子缺乏接收者,進而造成 ROS 累積,誘發氧化逆境,ROS 如:超氧化 物 和 H2O2 等 會 造 成 脂 質 過 氧 化 而 產 生 MDA,因此 MDA 的累積常作為細胞膜系統 過氧化指標 (Laxa et al. 2019),本研究亦檢測 這10 個品系的 MDA 含量,除了品系 93,其 餘品系的 MDA 累積皆隨著逆境時間延長而 增加 (Fig. 5E),而這些品系處理組的葉表面 溫度大多都較控制組高,顯示多數品系都因 PEG 6000 處理受到氧化逆境傷害。 為有效移除 ROS,植物會活化抗氧化酵
素和生成抗氧化物質,APX 和 CAT 在 ROS
的 清 除 扮 演 重 要 角 色 , 其 中 APX 在
ascorbate-glutathione (ASC-GSH) cycle 中 負責將 H2O2還原成H2O 而達到降低毒性, 因此多數植物遭遇氧化逆境時常見APX 活性 的提升,除了 APX,GR 和其他 ASC-GSH cycle 的酵素也常在乾旱逆境中誘導表現,而 CAT 則是在過氧化體負責催化 H2O2轉變成 H2O 的反應,當植物遭受嚴重乾旱逆境時, CAT 活性為降低氧化逆境害的關鍵(Laxa et al. 2019);在本研究的抗氧化酵素活性分析結 果可見,10 個品系中有 6 個品系 APX 和 GR 活性隨著逆境處理時間延長而提高 (Fig. 5B and D),品系 144、208、91 和 219 的二種酵 素活性皆在處理 4 天後顯著上升,說明這 4 個 品 系 面 臨 滲 透 逆 境 時 會 活 化 ASC-GSH cycle,而品系 110 和 152 的 APX 和 GR 活性 並未隨時間改變,其中品系 110 的葉表面溫 度亦沒有因逆境而有變化,雖然 MDA 累積 有增加,推測該品系遭受逆境壓力相對較低, 因此抗氧化酵素活性尚未明顯提升,而品系 152 在處理組的葉表面溫度顯著高於控制組, 但APX 和 GR 活性沒有變化,可能酵素活性 足以應付逆境壓力,或是由其他抗氧化途徑 減輕逆境傷害,在CAT 活性方面,品系 110、 208、91 和 93 皆在處理 4 天後下降,其餘品 系則是維持相似活性 (Fig. 5C),說明這 4 個 品系可能在逆境處理4 天後較不需要 CAT 來 清除ROS,而其他品系仍需維持 CAT 活性。 綜觀這10 個品系的滲透逆境反應,可發現遺 傳背景的差異對逆境耐性有顯著的影響,值 得注意的是,品系 110 的葉表面溫度和抗氧 化酵素活性變化顯示該品系可能耐受性相對 較高,未來可透過田間試驗進行確認,作為抗 旱育種之參考。 本研究建立水耕系統以快速檢測小米地 方品系的滲透逆境反應,並探討品系差異對 逆境反應及抗氧化能力的影響,未來可透過 基因型的鑑定和逆境性狀調查結果,來了解 遺傳背景和逆境耐性的關聯性,並找出關鍵 基因座,將有助於加速耐旱小米的育種效率。
誌謝
本研究由科技部計畫 (MOST 107-2321-B-002-060-)補助研究經費,特此致謝。引用文獻
Bennetzen JL, J Schmutz, H Wang, R Percifield, J Hawkins, AC Pontaroli, M Estep, L Feng, JN Vaughn, J Grimwood, J Jenkins, K Barry, E Lindquist, U Hellsten, S Deshpande, XW Wang, XM Wu, T Mitros, J Triplett, XH Yang, CY Ye, M Mauro-Herrera, L Wang, PH Li, M Sharma, R Sharma, PC Ronald, O Panaud, EA Kellogg, TP Brutnell, AN Doust, GA Tuskan, D Rokhsar, KM Devos (2012) Reference genome sequence of the model plant Setaria. Nat. Biotechnol. 30: 555-561.
Bloom AJ, FS Chapin, HA Mooney (1985) Resource limitation in plants–an economic analogy.
Annu. Rev. Ecol. Syst. 16: 363-392.
Boguszewska-Mankowska D, K Zarzynska, A Nosalewicz (2020) Drought differentially affects root system size and architecture of potato cultivars with differing drought tolerance. Am.
J. Potato Res. 97: 54-62.
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248-254.
Daryanto S, LX Wang, PA Jacinthe (2016) Global synthesis of drought effects on maize and wheat production. Plos One 11: e0156362.
Djanaguiraman M, PVV Prasad, J Kumari, Z Rengel (2019) Root length and root lipid composition contribute to drought tolerance of winter and spring wheat. Plant Soil 439: 57-73. Elliott J, D Deryng, C Mueller, K Frieler, M
Konzmann, D Gerten, M Glotter, M Florke, Y Wada, N Best, S Eisner, BM Fekete, C Folberth, I Foster, SN Gosling, I Haddeland, N Khabarov, F Ludwig, Y Masaki, S Olin, C Rosenzweig, AC Ruane, Y Satoh, E Schmid, T Stacke, QH Tang, D Wisser (2014) Constraints and potentials of future irrigation water availability on agricultural production under climate change.
Proc. Natl. Acad. Sci. USA 111: 3239-3244.
FAO. (2009) Global agriculture toward 2050 (Rome: FAO).
Foster JG, JL Hess (1980) Responses of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol. 66: 482-487. Heath RL, L Packer (1968) Photoperoxidation in
isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch.
Biochem. Biophys. 125: 189-198.
Henry A, AJ Cal, TC Batoto, RO Torres, R Serraj (2012) Root attributes affecting water uptake of rice (Oryza sativa) under drought. J. Exp. Bot. 63: 4751-4763.
Ho MD, JC Rosas, KM Brown, JP Lynch (2005) Root architectural tradeoffs for water and phosphorus acquisition. Func. Plant Biol. 32: 737-748.
Hund A, N Ruta, M Liedgens (2009) Rooting depth and water use efficiency of tropical maize inbred lines, differing in drought tolerance.
Plant Soil 318: 311-325.
IRRI (2013) Standard evaluation system for rice. International Rice Testing Program, Manila. 36 pp.
Kar RK (2011) Plant responses to water stress: role of reactive oxygen species. Plant Signal Behav. 6: 1741-1745.
Kato M, S Shimizu (1987) Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in senescing tobacco leaves; phenolic-dependent peroxidative degradation. Can. J.
Bot. 65: 729-735.
Koch DW. (2002) Foxtail milet-management for supplemental and emergency forage. In: SMRR Info Source. K Griffin (eds.) University of Wyoming, Wyoming.
Lata C, S Bhutty, RP Bahadur, M Majee, M Prasad (2011) Association of an SNP in a novel DREB2-like gene SiDREB2 with stress tolerance in foxtail millet [Setaria italica (L.)]. J. Exp. Bot. 62: 3387-3401.
Laxa M, M Liebthal, W Telman, K Chibani, KJ Dietz (2019) The role of plant antioxidant system in drought tolerance. Antioxidants 8: 94. Lin H, C Chiang, S Chang, G Liao, C Kuoh (2012) Genetic diversity of foxtail millet (Setaria italica) germplasm as determined by agronomic traits and microsatellite markers. Aust. J. Crop Sci. 6: 342-349.
Lin Y, X Yi, S Tang, W Chen, FK Wu, XL Yang, XJ Jiang, HR Shi, J Ma, GD Chen, GY Chen, YL Zheng, YM Wei, YX Liu (2019) Dissection of phenotypic and genetic variation of drought-related traits in diverse chinese wheat landraces.
Plant Genome 12: 1-14.
Liu TY, N Ye, T Song, Y Cao, B Gao, D Zhang, F Zhu, M Chen, Y Zhang, W Xu, J Zhang (2019) Rhizosheath formation and involvement in
foxtail millet (Setaria italica) root growth under drought stress. J. Integr. Plant Biol. 61: 449-462. Lobell DB, W Schlenker, J Costa-Roberts (2011)
Climate trends and global crop production since 1980. Science 333: 616-620.
Mishra AK, M Muthamilarasan, Y Khan, SK Parida, M Prasad (2014) Genome-wide investigation and expression analyses of WD40 protein family in the model plant foxtail millet (Setaria italica L.). PLoS One 9: e86852.
Nakano Y, K Asada (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 22: 867-880.
Pan J, Z Li, Q Wang, AK Garrell, M Liu, Y Guan, W Zhou, W Liu (2018) Comparative proteomic investigation of drought responses in foxtail millet. BMC Plant Biol. 18: 315.
Peng YL, JP Zhang, GY Cao, YH Xie, XH Liu, MH Lu, GY Wang (2010) Overexpression of a PLDα1 gene from Setaria italica enhances the sensitivity of Arabidopsis to abscisic acid and improves its drought tolerance. Plant Cell Rep. 29: 793-802. Ray DK, ND Mueller, PC West, JA Foley (2013)
Yield trends are insufficient to double global crop production by 2050. PLoS One 8. e66428. Shanker AK, M Maheswari, SK Yadav, S Desai, D
Bhanu, NB Attal, B Venkateswarlu (2014) Drought stress responses in crops. Funct. Integr.
Genomic 14: 11-22.
Shantz HL, LN Piemeisel (1927) The water requirement of plants at Akron, Colo. J. Agri.
Res. 34: 1093-1190.
Tang S, L Li, Y Wang, Q Chen, W Zhang, G Jia, H Zhi, B Zhao, X Diao (2017) Genotype-specific physiological and transcriptomic responses to drought stress in Setaria italica (an emerging model for Panicoideae grasses). Sci. Rep. 7: 10009.
Xie L-n, M Chen, D-h Min, L Feng, Z-s Xu, Y-b Zhou, D-b Xu, L-c Li, Y-z Ma, X-h Zhang (2017) The NAC-like transcription factor SiNAC110 in foxtail millet (Setaria italica L.) confers tolerance to drought and high salt stress through an ABA independent signaling pathway. J. Integr. Agric. 16: 559-571.
Zhang GY, X Liu, ZW Quan, SF Cheng, X Xu, SK Pan, M Xie, P Zeng, Z Yue, WL Wang, Y Tao, C Bian, CL Han, QJ Xia, XH Peng, R Cao, XH Yang, DL Zhan, JC Hu, YX Zhang, HN Li, H Li, N Li, JY Wang, CC Wang, RY Wang, T Guo, YJ Cai, CZ Liu, HT Xiang, QX Shi, P Huang, QC Chen, YR Li, J Wang, ZH Zhao, J Wang (2012) Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential. Nat. Biotechnol. 30: 549-554. Zhao MS, SW Running (2010) Drought-induced
reduction in global terrestrial net primary production from 2000 through 2009. Science 329: 940-943.