以肝病相關轉錄因子活性建立藥物搜尋系統; Research and Development of Drug Disease System Based on Liver Disease Relative Transcription Factors
59
0
0
全文
(2) 中國醫藥學院醫學研究所 碩士候選人學位考試. 論文題目. 以肝病相關轉錄因子活性建立藥物搜尋系統 之研究. Research and Development of Drug Disease System Based on Liver Disease Relative Transcription Factors. 本文係鄭欣怡 於中國醫藥學院醫學研究所完成之碩士論文經考試委員審查及口試合格特此 證明. 考試委員. 所長:. 中華民國九十二年 2. 月. 日.
(3) 誌謝. 感謝指導教授吳世祿老師及共同指導教授項千芸老師在實驗上及生活上對我的教誨,讓我 不只在實驗上受益良多,瞭解到在面對實驗時所應秉持的態度和精神。並感謝口試委員黃蘭如 老師對本論文辛勞審查,惠予寶貴意見及指正,使此論文得以更為完善。 研究期間並蒙侯庭鏞老師、李妙蓉老師、張乃文老師、方世華老師、劉昭君老師、鍾景光 老師及所裡諸位老師之關懷指導,受益匪淺,在此由衷的感謝。 研究期間並受到賴羿如學姊、王家男同學、李昌諴醫師、彭彥鈞醫師、劉旭然學長、張仲 達醫師、趙德澂學長、嘉穗學妹、欣裕學弟、淑婷學妹、佳橙學弟、暖螢學妹、奕瑾學妹等在 實驗工作上的協助與生活上的關心。以及同窗好友林麗瑩同學、邱華浩同學、李佳陽同學、陳 宣嘉同學、林美蘭同學、以及葉靜華、林伶娟同學、及醫學所陳麗如學姊在這兩年的相互扶持 與砥礪,讓我成長許多,心中仍有不捨之情。 最後要感謝我的家人,由於他們的鼓勵與支持使得學業得以順利進行,願將一切成果獻與 他們。. 欣怡謹誌. 3.
(4) 中文摘要. 肝細胞在受到病毒、化學物質或氧化傷害時,細胞內的轉錄因子會開始活化並引起炎症作 用並造成肝細胞的轉形。目前已知在肝細胞轉形的過程中,轉錄因子 Activator protein-1(AP-1) 在細胞中控制著細胞的生長、分化和細胞凋亡; 也控制著許多和腫瘤生成或細胞轉形相關的基 因表現。至於 Nuclear factor-?B (NF-?B) 則可調控細胞素的表現,而調控炎症作用的發生。因 此我們利用 AP-1 和 NF-?B 為標的,建立抗肝細胞轉形及抗發炎作用的藥物搜尋系統。 在抗肝細胞轉形的藥物搜尋方面 ,12-O-tetradecanoylphorbol-13-acetate (TPA)是為人所熟 知的腫瘤促進劑,會使細胞中的 AP-1 活化,進而促使細胞的轉形。因此我們利用兩種癌化程 度不同的肝細胞株 Chang liver/AP-1 與 HepG2/AP-1 細胞,架構成能報導細胞中 AP-1 活性的重 組細胞株,並以 TPA 作為誘發劑利用 Reporter assay 研究肝細胞中的 AP-1 活化與肝細胞轉形 的相關性;進而利用此模式作為抗肝細胞轉形的高速藥物搜尋系統,並從其中分析植物分類, 傳統中醫學上對於藥物的分型,以及在兩種不同肝細胞株 Chang liver/AP-1 與 HepG2/AP-1 細 胞中具有抑制 TPA 誘發 AP-1 活性的藥物,以及這些藥物中可能發揮生物活性的物質和含有這 些藥物的複方在臨床上的應用。並發現搜尋系統中選出的抑制能力較佳的藥物,在植物分類學 上及臨床中藥複方的應用上具有相關性。 在炎症作用的模式方面,酒精的氧化傷害和肝炎病毒感染的情況下,會引起肝細胞的炎症 作用,此時細胞中的 NF-?B 會被高度的活化,並促使與發炎作用的細胞素相關基因大量表現。 在我們的研究中,前期利用細菌之內毒素 lipopolysaccharide (LPS) 充當抗肝細胞炎症作用藥物 的搜尋模式,由 543 種藥物中選出抑制 LPS 誘發肝細胞內 NF-?B 活性能力排名前 54 名的藥 物,再利用酒精的代謝物—乙醛作為酒精對肝細胞氧化傷害的搜尋模式觀察這 54 種藥物對於 乙醛誘發 HepG2 細胞中 NF-?B 活性的影響。 前述的肝病相關因子與肝病重要誘發物質代表特定疾病的搜尋模式中,毛茛科植物黃連與 芸香科植物黃柏的根莖、韌皮部所分離的生物鹼— 小檗鹼具有抑制在 HepG2 細胞中乙醛所誘 發的 NF--?B 活性之能力。探究其抑制轉錄因子的作用機轉主要原因為:小檗鹼會抑制 I-?B 的 磷酸化的訊息傳導路線。此外小檗鹼對於 HepG2 細胞中 NF-?B 所調控和炎症作用具重要相關 4.
(5) 性的細胞素— Tumor necrosis factor-α(TNF-a)與 Interleukin-1β(IL-1ß)的分泌具有抑制的 作用未來將進一步研究小檗鹼是否對細胞核內, NF-?B 與 DNA 的結合能力有所影響,並觀 察小檗鹼在體內試驗中所表現的生物活性。. 5.
(6) Abstract. Several transcription factors have been shown to be involved in the pathogenesis of hepatitis virus, chemicals, and oxidative stress- induced chronic liver disease. Activator protein-1(AP-1) is an important factor in carcinogenesis and cellular transformation, while nuclear factor- kappa B (NF-?B) is an inflammation–related factor that the expression stimulate inflammatory cytokine genes activation during inflammation. In this study, the induction of AP-1 and NF-?B in human hepatocyte Chang liver and HepG2 by TPA, LPS and acetaldehyde, respectively, represented the hepatocellular transformation and inflammation models. The methanol extracts of Chinese herbal were used to evaluate screen their therapeutic efficiency. Our data showed that 10 µM acetaldehyde induced AP-1 and NF-?B activities in HepG2 cells at 8 hours. Berberine, an alkaloid from Coptis chinensis, Huang lian, (Ranunculaceae) or Phellodendron amurense, (Rutaceae) roots and cortex, that exhibited a significant inhibitory effect in the acetaldehyde-induced NF-?B activities, its 50% of effective concentration is 18.0 µM , repectivety. Berberine can inhibit NF-?B activities by suppressing the phosphorylation of I-?B. Furthermore, acetaldehyde-induced inflammatory cytokines, Tumor necrosis factor-a(TNF-a) and Interleukin-1ß(IL-1ß), were inhibited by berberine. These results suggested that inhibit the alcoholic oxidative damage, resulting in the protective effect in hepatocytes.. 6.
(7) 目錄. 中文摘要… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ...4 英文摘要… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ...6 目錄… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ...7 圖目錄… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ...8 表目錄:… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … .… .9 第一章. 前言… … … … … … … … … … … … … … … … … … … … … ..… … … … … … … … ..10. 第二章. 文獻探討… … … … … … … … … … … … … … … … … … … … … … … … … … .… ...12. 第三章. 實驗架構與研究設計… … … … … … … … … … … … … … … … … … … … .… … ...17. 第四章. 研究材料與統計方法… … … … … … … … … … … … … … … … … … … .… … .......20. 第五章. 研究結果… … … … … … … … … … … … … … … … … … … … … … … … … … … ....23. 第六章. 討論… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … 29. 第七章. 結論與建議… … … … … … … … … … … … … … … … … … … … … … … … … … … 32. 參考文獻:… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … .47 著作權聲明… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ... 7.
(8) 圖目錄. 圖一、酒精性肝炎之發生機轉… … … … … … … … … … … … … … … … … ..… … … … … … .… .15 圖二、酒精代謝物乙醛對 AP-1 及 NF-?B 上游訊息傳導路線的影響… … … … .… … … … ...16 圖三、本論文的研究架構… … … … … … … … … … … … … … … … … … … … … … … … … .… ...19 圖四、本實驗使用的藥物庫名單… … … … … … … … … … … … … … … … … … … … … … … ....33 圖五、TPA 可以誘發 Chang liver/AP-1 和 HepG2/AP-1 細胞中 AP-1 的活性… … … … … ....34 圖六、藥物對 TPA 誘發 Chang liver/AP-1 細胞中 AP-1 活性的影響:… … … … .............… ....35 圖七、藥物對 TPA 誘發 HepG2/AP-1 細胞中 AP-1 活性的影響… … … … … … ....… … … .....36 圖八、植物科別對 Chang liver/AP-1 與 HepG2/AP-1 細胞中 AP-1 活性的影響… … ..… … ..37 圖九、LPS 可以誘發 HepG2/NF-?B 細胞中 NF-?B 的活性… … … … … … … … … … … … … .39 圖十、藥物對 LPS 誘發 HepG2/NF-?B 細胞中 NF-?B 活性的影響… … … … … … … … ........40 圖十一、乙醛可以誘發 HepG2 細胞中 NF-?B 活性的劑量和時間作用… … … … … … … .… 41 圖十二、54 種藥物對乙醛誘發 HepG2 細胞中 NF-?B 活性的影響.… ..… … … … … … … … .42 圖十三、小檗鹼可以抑制 HepG2 細胞中乙醛所誘發的 NF-?B 活性… … … … … .… … … ....43 圖十四、小檗鹼可以抑制 HepG2 細胞中乙醛誘發的 I-?B 磷酸化… … … … .… … … … … … 44 圖十五、小檗鹼可以抑制乙醛誘發 HepG2 細胞 TNF-a 的分泌… … … … … … … ................45 圖十六、小檗鹼可以抑制乙醛誘發 HepG2 細胞 IL-1ß 的分泌… … … … … … … … … … … ...46. 8.
(9) 表目錄. 表一、在 Chang liver/AP-1 及 HepG2/AP-1 細胞中皆能抑制 TPA 誘發 AP-1 活性的中藥… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ..38. 9.
(10) 第一章. 前言. 第一節. 研究緣起. 癌症,歷年來一直處於國人十大死因的前一、二名(行政院衛生署),而肝癌又常居癌症十大 死因的前一、二名。肝病和肝硬化和其他的肝臟相關疾病,則位於十大死因的第六名(2001, 行政院衛生署)肝病與肝炎病毒的感染有關,在統計學上,B 型肝炎帶原者惡化成肝硬化和肝 癌的比例較非帶原者高 20 倍,近二十年來,由於 B 型肝炎疫苗預防注射的推廣,使得國人在 20 歲以下的年齡層的帶原率降低到 1.0 %,但是在壯年的人口中仍然有 10 %屬於 HBsAg 呈現 陽性作用,另外,由於輸血、性行為或垂直感染 C 型肝炎的人口在近十年來有增加的趨勢, 尤其是在台灣西部沿海地區 C 型肝炎陽性的比率達到 10 %(嘉義) ,在罹患肝炎的狀況下,酗 酒、藥物濫用會使得肝病更加嚴重,除了預防和治療外,從現代科學研究的層面去了解肝病的 致病機轉,和如何針對這些機轉去預防或減弱甚至阻斷肝病的發生,是一個值得去研究的課 題;而利用肝病的致病機轉來導引新藥的研發也是值得國人努力的方向。本研究利用重組的人 類肝癌細胞株,分析肝病重要的誘發因子引發疾病的分子機轉,建立肝病用藥的高速搜尋模 式;並利用與 GMP 中藥廠分讓和坊間購買的中藥材,以標準流程萃取的萃取液和臨床上常用 的藥物,與一些生藥的組成成分,搜尋以上的物質是否對肝細胞中特定轉錄因子的活性具有抑 制作用,並篩選出具有抑制特定轉錄因子活性的藥物,分析分子作用機轉與藥物對下游調控基 因活性的影響。希望能夠把基礎研究和實際的臨床應用結合,達到新藥開發的目的,並? 短藥 物的研發時間。. 10.
(11) 第二節. 研究目的. 藉由兩種不同癌化程度的重組人類肝細胞株 Chang liver/AP-1 與 HepG2/AP-1 或者是 HepG2/NF-?B 等細胞,Chang liver 細胞的性狀較接近於正常人的肝細胞,在實驗系統中用來 代表正常人的肝細胞;而觀察肝病之重要誘發因子,誘發細胞中肝病相關轉錄因子的活性,代 表特定疾病模式;觀察中藥或臨床所使用的藥物對特定誘發劑活化轉錄因子活性的影響。選擇 可以抑制特定誘發劑誘發轉錄因子活性的藥物,深入研究藥物抑制轉錄因子活性之分子機轉與 其對下游基因的影響,解釋藥物發揮藥效的原因,以達到新藥開發和舊藥新用途的目的。. 11.
(12) 第二章. 文獻探討. 第一節 肝病和肝細胞轉形的重要因子 Activator protein 1(AP-1)是一個控制細胞生長分化和死亡的重要轉錄因子,由 Jun 與 Fos family 的蛋白質所組成,與 DNA 結合發揮轉錄活性的部位是稱為 Leucine zipper 的忌水性 構造。. 在囓齒類動物胚胎發育的過程中,如果 Jun 或 Fos family 的基因被剔除,會使得胚胎. 的卵黃囊、心臟、腦的發育受阻導致死亡。而 Jun 或 Fos family 的基因在細胞或體內大量表現 時則易導致細胞轉形或癌症的發生。近年來的研究指出 AP-1 的活化,主要是經由稱為 Mitogen activated protein kinase(MAPK)的訊息傳導路線;分為 ERK、JNK 和 p38 等。近年的研究指 出,當 B 型肝炎的 X 蛋白質或 C 型肝炎的核心蛋白質(core protein)在基因轉殖鼠的體內表 現時,會活化 ERK 和 JNK 的路徑並造成肝細胞癌的發生(Tsutsumi et al., 2002;Ruchika et al., 2001),為了解 MAPK 中主要經由何種途徑導致肝細胞的轉形加入 MAPK 中對不同訊息傳導 路徑的抑制劑發現對於 ERK 上游蛋白質激?. MEK-1 專一性的抑制劑 PD98059,可以抑制肝. 癌的發生(Ruchika et al., 2001)。 TPA(12-O-Tetradecanoylphorbol 13-acetate)是一個為人所熟知的腫瘤促進劑,以 TPA 刺激囓齒類動物的皮膚後,會使皮膚產生炎症作用,並促使皮膚癌變產生(Tokuda et al., 1996)。另外於基因轉殖鼠中,以不同途徑分別投與 TPA 後,會造成各組織 AP-1 的活化,尤 其以塗抹 TPA 的方式為最為顯著(Zhong et al.,2001)。於本實驗室之前的研究中,以 TPA 刺 激重組人類肝細胞株 Chang liver/AP-1 與重組人類肝癌細胞株 HepG2/AP-1,在數小時後會使 細胞中的 AP-1 活化,在數週後則會促使細胞轉形的發生(賴, 2001) ;加入 PD98059 後則會呈 現抑制肝細胞中 TPA 所誘發的 AP-1 活性,並抑制肝細胞的轉形(賴, 2001)。 自 1999 年以來,以 DNA 晶片等技術用來觀測大規模的藥物對細胞作用,引發不同基因 波動來詮釋藥物-基因體之間的交互作用,並藉由分析單一藥物在多種細胞中對各個基因的影 響,以求藥物有更廣泛的應用性,並探究在癌化過程中特定基因所扮演的角色,希望以此用來. 12.
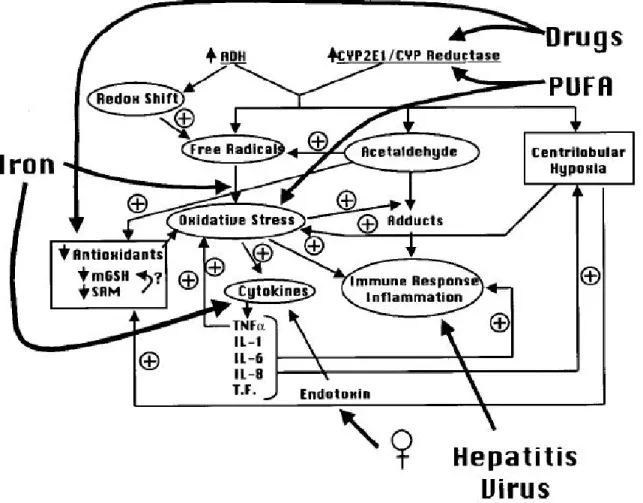
(13) 做藥物新用途的開發,或觀測各種疾病在基因上的關聯性,已成為了一個新趨勢(Pinkel, 2000;Ross et al., 2000;Scherf et al., 2000)。在此我們希望以能夠報導肝細胞中控制細胞轉 形,或炎症作用與氧化傷害等作用的系統,並利用特定誘發劑誘發特定轉錄因子活性,代表特 定疾病觀測傳統使用的中草藥,臨床常用之抗癌藥等,對於肝細胞中 AP-1 活性的影響,並從 其中搜尋出可能的用藥,以提供新藥開發、藥物新用途等功能,並提供更快速更靈敏的藥物搜 尋平台。. 第二節. NF-?B 和肝病的相關性. Nuclear Factor-kappa B(NF-?B)為一個控制細胞免疫相關因子表現的轉錄因子,許多細胞 素的啟動子區域,都有NF-?B的結合位置。另外在腫瘤生長或發生炎症作用的組織中,也有 NF--?B大量表現的情形,肝臟在長期受酒精刺激或是革蘭氏陰性菌細胞壁的脂多糖體 (Lipopolysaccharide, LPS)存在的情況下,會使得肝細胞內的NF-?B高度活化,並造成發炎性 的細胞素如TNF-a、CD14(LPS receptor) 、TGF-ß、IL-1及 IL-6等大量表現促進炎症作用產生, (Gutierrez et al., 1999;Michael and Ronald, 2002;Yin et al., 2001;Bode et al., 1993;Luster et al., 1994)。. 第三節. 酒精性肝病的重要誘發因子— 乙醛對肝細胞中 AP-1 和 NF-?B 的影響. 在酒精性肝炎發生的過程中;乙醛為重要的促進因子,乙醛在體內會與血液中的白蛋白形成 Malondialdehyde-acetaldehyde-protein adduct的複合物,以便於輸送(Willis et al., 2002)。乙醛 在代謝的過程中,會產生自由基對組織造成氧化傷害。在許多研究指出,乙醛在肝臟中會造成 肝實質細胞和儲脂細胞內AP-1和NF-?B的活化( Chen and Bernard . 2000;Fernandez-Checa, 2000;Michael et al., 2002; Juan et al., 2000. Youn et al., 2002 ),並促使一些下游基因啟動造 成TNF-a、IL-6等發炎性細胞素的表現,促進肝臟產生炎症作用(Gutierrez et al., 1999),(圖 一)。在肝的儲脂細胞中,乙醛會透過JNK磷酸化的訊息傳導路線活化,使AP-1活化,使得細 胞中常在的轉錄因子Basic transcription element binding protein(BTEB)大量表現,並結合到第. 13.
(14) 一型膠原蛋白基因的啟動子區域促使第一型膠原蛋白的大量表現,使儲脂細胞轉形成為纖維芽 母細胞(Myofirboblast)(Roman et al., 2000),造成肝的纖維化。在肝實質細胞的方面,在亞 洲人體內因為Acetaldehyde dehydrogenase II(ALDH 2)酵素的活性比高加索人種的活性低使 得對酒精的耐受性降低並產生心博加快、血中乙醛上升的副作用,進而對肝細胞造成氧化傷 害。而在人類肝癌細胞株Hep G2中也因為此種酵素的缺乏與具有B型肝炎病毒的基因體存在等 特性,所以可以用來作為研究在酒精性肝損害的的過程中,酒精代謝物傷害肝臟中肝實質細胞 的細胞模式,並用來觀察乙醛對肝細胞中一些因子的影響。. 第四節. 肝病的治療用藥. 由以上諸多因子對肝細胞的影響,除了可以用來作為了解肝病的發生機轉外,也可用以 上物質作為代表特定肝病的誘發劑,並以相關因子作為標的來作為藥物活性搜尋的平台。除了 可以為現代臨床上使用的藥物開發新用途外,並可以從傳統使用的生藥中,除了給予傳統醫 理、藥理的現代科學解釋外,並且達到由傳統藥物中,找出可以供給新藥開發的標的。在臨床 上醫治病毒性肝炎,除了注射 ?-干擾素,抑制病毒侵襲正常的肝細胞。然而一些植物所含的 物質,如從水飛薊所提煉的物質 Silymarin、Silibinin 具有保肝的功能(Varga et al.,1991; Somogyi et al., 1989),柴胡皂素 a 在人類肝癌細胞株中具有促進細胞生長抑制的活性 (Wu, 2003)。另外如 Berberine 具有抑制四氯化碳誘發之齧齒類動物的肝纖維化、保肝等作用(Hwang et al., 2002),而在酒精與 LPS 攻擊囓齒類動物造成肝損傷的模式,Dilinoleoylphosphatidylcholine 具有降低發炎性細胞素 TNF-a 的功能(Cao et al., 2002)。由以上前人的研究給予我們一個方向 -由傳統使用的中草藥方,及一些固有的西藥中,搜尋有用的材料,或是開發藥物新的用途, 是一個嶄新的方向。. 14.
(15) 圖 一、酒精性肝炎之發生機轉。 分為初級引發因子和二級之危險因子,初級因子包括細胞內經酵素代謝後之乙醛、自由 基等分子,使得細胞產生氧化傷害的作用; 並促使二? 因子的活化:如細胞素表現,造 成肝細胞的炎症作用。另外,肝炎病毒細菌內毒素的存在也會刺激免疫作用造成發炎現 象的產生,也是乙醛對肝臟造成損害的原因 。 (Hidekazu et al., 2001). 15.
(16) 圖二、酒精代謝物乙醛對 AP-1 及 NF-?B 上游訊息傳導路線的影響。 乙醛會經由 PKC 的路徑,分別促進經 I-?B 的磷酸化和解離,造成 NF-?B 轉移到細胞 核中並啟動下游基因的表現。另外經由 MAPK 路徑 ERK 與 JNK 的磷酸化,造成細胞 核內 AP-1 的活化。 (Gutierrez et al., 1999;Juan et al., 2000;Won et al., 2001). 16.
(17) 研究架構與研究設計. 第一節. 研究設計. 基於想要研究藥物對於不同癌化程度的肝細胞之影響,使用兩株人類的肝細胞株,並 利用特定誘發劑誘發肝細胞中特定轉錄因子活性,代表特定肝病模式作為新藥開發的系 統。在此我們用形態性質接近於正常人類肝細胞的 Chang liver 細胞,代表正常的肝細胞。 另外因為 HepG2 細胞具有 HBV 的表面抗原表現和 a-fetoprotein 的大量表現。在此我們用 來代表感染 B 型肝炎病毒,並用來代表開始癌化的肝細胞。在藥物庫的蒐集和建立方面; 由順天堂公司分讓或由坊間購得之中藥藥材,以標準流程萃取,並執行品質管制;或是從 Sigma 等廠商購買藥物和植物所含的主要成分,共 543 種藥物。藥物與特定誘發劑的測試 方式為:加入藥物後,再加入特定誘發劑作用,觀察以上物質對細胞中特定轉錄因子活性 的影響。. 第二節. 研究架構. 首先利用實驗室架構的 Chang liver/AP-1、HepG2/AP-1(賴,2001)與 HepG2/NF-?B 的 重組細胞株(賴,2001) ,加入稀釋成不同濃度的 TPA、LPS 或是乙醛作為 AP-1 或 NF-?B 的誘發劑選擇誘發轉錄因子活性的最佳條件值,作為之後藥物搜尋模式的起點。在搜尋結 果中,除了做藥物對轉錄因子活性影響的分析外,並選出能抑制特定誘發劑誘發特定轉錄 因子活性的藥物,探究藥物能夠抑制轉錄因子活性的機轉,並研究藥物是否對特定轉錄因 子調控和疾病有高度相關性的基因具影響力。. 17.
(18) 第三節. 研究假說. 在實驗室的研究中,發現 TPA 可以活化肝細胞中 AP-1 的活性並促進肝細胞的轉形(賴, 2001)。LPS 可以誘發肝細胞中 NF-?B 的活性,並造成和肝細胞炎症作用相關的細胞素- TNF-a、IL-1ß 蛋白質的表現(Dong et al., 1998a)。而乙醛也具有誘發肝細胞中 NF-?B 活性 的能力(Fernandez-Checa, 2000)。由以上這些誘發劑誘發不同的肝細胞株中 AP-1 或 NF-?B 的活性,代表特定疾病模式,作為藥物搜尋的起點觀察中藥或一些? 物質,是否具有抑制這 些誘發劑活化轉錄因子活性的能力,並利用酵素免疫吸附法等方式觀查這些誘發劑促進轉錄 因子的活化,是否可以促進一些受特定轉錄因子調控,與肝細胞的轉形或發炎作用相關基因 的表現。從其中選出能夠抑制特定誘發劑誘發特定轉錄因子活性的用藥,分析藥物抑制特定 轉錄因子活性是作用在何種路徑,並研究藥物是否也具有抑制受到特定轉錄因子調控,與疾 病發生有高度相關性的基因表現;例如發炎性的細胞素與一些成份性的蛋白質的過量表現, 會使肝組織產生炎症作用,或是促進肝細胞的轉形,進而阻止肝細胞轉形及氧化傷害作用的 發生。. 18.
(19) 肝病相關誘發 劑誘發重組人 類肝細胞株中 AP-1 與 NF--?B 活性代表特定 肝病模式的建 立. 藥物庫 的建立. 以 TPA 作為肝 細胞轉形模式 的誘發物 藥物對 特定肝 病模式 的高速 搜尋系 統. 以 LPS 作為肝細 胞炎症作用模 式的誘發物 以乙醛作為酒精代 謝物造成肝細胞氧 化傷害模式的誘發 物 中藥萃取液的 標準製作流程 及品質管制. 抑制特定肝病模 式中轉錄因子活 性用藥的篩選. 大量搜尋的資 料分析統計. ? 化合物 的收集. 藥物分子作用機轉的研究. 新藥開發. 圖三、本論文的研究架構。 (A) 以特定誘發物誘發重組人類肝細胞株中 AP-1 與 NF-?B 活性,代表特定肝病模式的 建立:分別以 TPA、LPS、乙醛代表不同肝病模式的誘發因子,並利用誘發劑誘發轉錄 因子活性。 (B) 藥物庫的建立:藥物的蒐集和建立方面-由順天堂公司分讓或由坊間購得之中藥藥 材,以標準流程萃取並執行品質管制,或是透過購買的方式取得純物質。 (C) 肝病可能用藥的研究:抑制特定誘發劑誘發轉錄因子活性的藥物分子機轉研究。 19.
(20) 第四章. 第一節. 研究材料與統計方法. 研究材料. 中藥萃取 由順天堂公司分讓或由坊間購得之中藥藥材,以 9 克的藥材對 10 毫升的甲醇(Merck)的比 例,密封在 50 毫升離心管中,以室溫 100 rpm,搖晃萃取 48 小時後,移至-70℃冷凍 12 小時 後,再將中藥萃取液做無菌檢測;確認有無細菌生長後,以 0.45 µm 的濾膜(PALL)過濾,去除 微生物並加以分裝,另外取 1 毫升以 LABCONCO 的真空乾燥機,低溫乾燥以便估算? 重, 中藥萃取液的濃度以 ug/ul 表示在後來的大量藥物搜尋中,中藥萃取液以 1/1000(v/v)的體積, 以一倍的 Dulbecco’s Modified Eagle Medium(GIBCO) DMEM 稀釋,作為搜尋模式中所使用的 濃度。. 第二節. 細胞培養與純化合物蒐集. 在本實驗中主要是利用 Chang liver/AP-1 和 HepG2/AP-1(賴,2001),來描述藥物對不同 癌化程度的人類肝細胞株中 AP-1 的影響;此外使用 HepG2/NF-?B 的重組細胞株(賴,2001), 測定藥物對肝細胞中 NF-?B 的影響,以含有 10 % 胎牛血清 (Hyclone)的 Dulbecco’s Modified Eagle Medium(GIBCO) DMEM,培養於 37℃的培養箱中,LPS (Lipopolysaccharide from E. coli) 和 TPA 與黃酮類、多酚類等藥物購自 Sigma。LPS 以純水溶解,TPA 和黃酮類等藥物以絕對 酒精或甲醇溶解。. 第三節. AP-1 與 NF-?B 的活性測定. 細胞在 75T flask (Grenier)中培養至全滿以 trypsin 消化,以含有 10 %胎牛血清的 DMEM 稀釋,把細胞均勻分散到 96 孔培養盤中;過了 24 小時後,加入以 DMEM 稀釋的藥物作用, 在固定時間後,先吸去上層之培養基後以冰冷的一倍 PBS (Phosphate Buffer Saline)清洗,再以 Lysis buffer (含 50 mM Tris-HCl, pH 7.8,1% Triton X-100, 1 mM DTT,)溶解細胞。最後取 20µl 20.
(21) 的細胞溶解液,以 1:1 (v/v)的比例加入 Luciferase reagent (含 33.3 mM DTT,270 µM Coenzyme A, 530 µM ATP, 470 µM Luciferin (Firefly)), LAR (含 20 mM Tricine, pH 7.8, 1.07 mM (MgCO3 )4 Mg(OH)2 , 2.67 mM MgSO4 , 0.1 mM EDTA)作用,以 FB15 Lumiometer 測定冷光強度; 單位以 RLU (Relative Light Unit)表示,代表細胞內加入藥物固定時間後轉錄因子的活性。. 第四節. 細胞活性測定 (MTT method). 把細胞培養在 96 孔盤,過了 24 小時後加入以 DMEM 稀釋的藥物作用在固定時間後,加 入 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),作用 4 小時,加入 0.1 % SDS-HCl (含 10 mM HCl, 20% SDS),24 小時後,以 570 nm 的波長測定吸光度的強弱,來代 表細胞的活性;吸光度越強代表細胞活性越佳,反之則否。藉此用來描述藥物對細胞活性的影 響。. 第五節. 西方點墨法 ( Western blot). 細胞在 25T flask 中,培養至全滿,移去上層之培養基後,加入以 DMEM 稀釋的藥物,作 用固定時間後把細胞放在冰上,? 止作用,並以冰冷的一倍 PBS 清洗;加入 SDS sample buffer (含 62.5 mM Tris-HCl, pH 6.8, 2% SDS, 10% Glycerol, 50 mM DTT, 0.1% Bromophenol blue), 溶解細胞,以超音波震盪器震盪 10 秒,擊碎細胞後,再以 4℃、12000 rpm、離心 5 分鐘後, 以 100℃煮沸 5 分鐘後,用含 10% polyacrylamide 的膠體做蛋白質電泳,並以硝酸纖維薄膜 ( Nitrocellulose paper, NC paper, Amersham® ) 轉印,再加入用 Blocking buffer (含 1 倍 Tris buffer saline (TBS), 0.1% Tween-20, 5 % Non- fat dry milk)稀釋的抗 I-?B,phospho-I-?B 的抗體 (New England Biolab) 做雜合作用後,加入抗兔子的 IgG 抗體,最後以 ECL (Amersham® )呈色最後 以 X 光底片顯影。. 21.
(22) 第六節. 細胞素的酵素連結免疫吸附法( Cytokine ELISA). 細胞以前述方式繼代和加藥後,過了固定時間吸取上層之培養基,以每一格 200 µl 的體 積加入作用盤中,接著使用 BD 的 OPtEIAT M human TNF-a 和 OPtEIATM human IL-1ß 套組, 以含有一倍 ABTS 的檸檬酸/磷酸緩衝液,作為受質,並以 405 nm 的波長測量吸光度;並以內 插法的方式計算出樣品的濃度,以 pg/mL 表示。. 第七節. 統計方法. 本研究中使用 Excel XP (Microsoft® )計算多次重複實驗中數據的平均值與標準差,並利用 Student’s t test 分析有無顯著差異存在。另外藥物對誘發劑活化轉錄因子活性的影響,則以誘 發劑誘發細胞中轉錄因子活性的平均倍率為基準(1 倍),以 Relative AP-1activity 或 Relative NF-?B activity 表示。. 22.
(23) 第五章. 研究結果. 第一節. TPA 可以誘發 Chang liver/AP-1 與 HepG2/AP-1 細胞中 AP-1 的活性. 細胞在繼代到 96 孔盤後,待其生長到全滿;把生長環境中的血清去除,加入含 0.1 % 胎牛血清的一倍 DMEM,培養超過 24 小時稱為 starvation,目的是為了使細胞的細胞週期停 止在 G1 phase。另外,AP-1 為一個受血清調控的因子,一旦有血清加入,AP-1 就會呈現持續 活化的狀態。加入以一倍 DMEM 稀釋不同濃度的 TPA 溶液,作用 16 小時後,觀察 TPA 對細 胞中 AP-1 的作用。在 Chang liver/AP-1 細胞中濃度為 1 ng/mL 的 TPA 即可活化 AP-1 的活性 約為 2 倍(圖五,A),而高於此濃度後,仍然有繼續上升的趨勢,到了濃度為 100 ng/mL 時約 可活化 6.5 倍。而 HepG2/AP-1 細胞則不易受到 TPA 的活化,TPA 的濃度大於 50 ng/mL 之後, 才有較明顯的活化的作用到了濃度為 100 ng/mL 時約可活化 2 倍(圖五,B)。. 第二節. 藥物對 TPA 誘發 Chang liver/AP-1 細胞 AP-1 活性的影響. 細胞以前述方式繼代和去除血清培養以後,加入以一倍 DMEM 稀釋的藥物包含中藥的 甲醇萃取液,黃酮類、多酚類等和常用的抗癌藥物等。甲醇萃取液以千分之一的濃度,黃酮類 等植物的主要成份之最終濃度為 25 µM ,抗癌藥物其成份分為五大類(1)DNA 的烷化劑(2) DNA 雙股的插入劑(3)Topoisomerase I 的抑制劑(4)Topoisomerase II 的抑制劑(5)鹼基的 類似物等,則以一倍的 DMEM 稀釋成 1 µM 的濃度,再加入濃度為 20 ng/mL 的 TPA,培養 16 小時後,以材料與方法所敘述的方式溶解細胞,測定冷光活性得到以下結果(圖六)。以 TPA 在細胞中活化的平均倍率為 1(紅色),藥物如果有抑制 TPA 所誘發 AP-1 的活性,會呈 現作用向冷色調(右方)移動的圖譜,反之則會向暖色調(左方)移動,20 ng/mL 的 TPA 約 可活化細胞中的 AP-1 活性達 14.5 倍,至於對抑制 Chang liver/AP-1 細胞 TPA 所誘發 AP-1 活 性的藥物,其植物分類對 AP-1 活性的影響和傳統藥性的區別,對 AP-1 活性的影響,在 508 種藥物對 TPA 所誘發 HepG2/AP-1 細胞 AP-1 活性的大量搜尋會把這兩種細胞做分析比較。. 23.
(24) 第三節. 藥物對 TPA 所誘發 HepG2/AP-1 細胞活性的影響. 細胞以前述的方式繼代和加藥作用 16 小時後,收取細胞的溶解液,測定冷光活性。以 TPA 在 HepG2/AP-1 細胞中活化的平均倍率為基準,觀察藥物對 TPA 誘發 AP-1 活性的影響,如果 藥物對 HepG2/AP-1 細胞中 TPA誘發的 AP-1 活性呈現抑制者,偏向冷色系;藥物對 HepG2/AP-1 細胞中 TPA 誘發的 AP-1 活性呈現活化者,偏向暖色系(圖七)。在第四節開始把藥物對 TPA 誘發 Chang liver/AP-1 及 HepG2/AP-1 細胞 AP-1 活性的作用,做植物分類學上,各個科別對兩 種不同肝細胞株中 TPA 誘發 AP-1 活性影響的比較;或是在不同的肝細胞株中皆有抑制 TPA 誘發 AP-1 活性能力的藥物,做以下的分析比較。. 第四節. 植物科別對兩種不同肝細胞株中 TPA 誘發 AP-1 活性的影響. 在藥物庫中一共有 102 種不同科別的植物,除此之外尚有西藥、中藥複方、動物和礦物藥 等。因為動物藥有包含節肢、軟體、環節和哺乳動物們等動物組織入藥,牽涉較為複雜並且組 個數太少,在此部份沒有進一步去探討。分析方式以計算同一科別不同植物對 AP-1 的作用為 一個體,計算同一科別不同物種對 AP-1 影響的平均並計算標準差。在抗癌藥物的部份;有幾 種藥物在 Chang liver/AP-1 細胞中對 AP-1 沒有明顯的影響,但是在 HepG2/AP-1 細胞中,卻 會促使 TPA 誘發的 AP-1 活性高度活化,可能是因為 HepG2 細胞為開始癌化的細胞株,所以 會有不同的作用(圖八)。 藥物庫中屬於被子植物門,雙子葉植物,科別內有較多種植物的有:豆科、菊科、薔薇科、 芸香科、繖形科等。而單子葉植物的部份;在藥物庫中收集最多的為禾本科的植物,另外也有 真菌類入藥。在兩種肝細胞中對 TPA 誘發的 AP-1 活性皆呈現抑制作用的有:毛茛科 (Ranunculaceae) 、小檗科(Berberidaceae) 、蓼科(Polygalaceae) 、桑寄生科(Loranthaceae)、 防巳科(Menispermaceae) 、菫菜科(Violaceae)等。在較接近於正常的肝細胞的 Chang liver/AP-1 細胞呈現抑制,但是在 HepG2/AP-1 中對於 TPA 誘發的 AP-1 活性沒有抑制,呈現持平的有棕 櫚科(Palmae)、木蘭科(Magnoliaceae)、木犀科(Oleaceae)、桑科(Moraceae)、五加科 (Araliaceae)等在 HepG2 細胞中對 AP-1 呈現抑制作用,在 Chang liver/AP-1 細胞中對 AP-1. 24.
(25) 活性沒有影響的有錦葵科(Malvaceae)、天南星科(Arecaceae)、旋花科(Convolvulaceae)、 桔梗科(Campanulaceae)等在兩種肝細胞中皆呈現活化 AP-1 作用的有鳶尾科(Iridaceae)、 報春花科(Primulaceae)等植物,但是因為有一些科別的植物在藥物庫中的個數不多,所以 還要在收集更多的植物才能下定論。另外在一些較多種植物樣本的科別如豆科、芸香科等則呈 現很大的岐異度;同一科別的不同種植物,有的對 TPA 誘發的 AP-1 呈現抑制作用,有的則沒 有差異或呈現活化的作用。有鑑於此,稍後會敘述在兩種不同的肝細胞株中,抑制 TPA 誘發 AP-1 活性的前十四名藥物的分析以及臨床之應用。. 第五節. 在 Chang liver/AP-1 和 HepG2/AP-1 細胞中皆能抑制 TPA 所誘發 AP-1 活性的前 十四名藥物的分析. 由以上大量搜尋的結果中,選出抑制在兩種肝細胞中具有抑制 TPA 誘發 AP-1 活性的前 50 名藥物(結果在此不顯示),再選出兩種細胞中皆能抑制 AP-1 活性的藥物;使用以下的方 式分析藥物可能抑制 AP-1 活性的原因。分為科別、指標性成分、抑制 50 % AP-1 活性所需的 濃度(表一所顯示的為藥物在 HepG2/AP-1 細胞中,抑制一半 TPA 誘發 AP-1 活性所需的濃度) (EC50 )和臨床上的應用(複方)以求證搜尋系統的有效性(表一)。在科別的部份;十四種 藥物中有三種繖形科的植物(當歸、羌活,柴胡)芸香科的植物有兩種(黃柏、吳茱萸)和兩 種龍膽科的植物(龍膽、秦? ) 。 因為藥物庫中也有一部份的植物指標成分,也可做為雙重確 認。例如:黃連和黃柏都具有抑制 TPA 誘發 AP-1 活性的功能,而其中所含的有效物質:小檗 鹼( Berberine)也有相同的藥效。從此處也可以觀察到,有一些複方會同時使用以上幾種藥物, 除了證實搜尋系統的有效性,也可以指出一個未來的方向,包括複方的運用方式或是藥物的併 和使用會具有藥效加成的作用;另外,也可以使一些藥物開發新的使用途徑。. 第六節. LPS 可以誘發 HepG2/NF--?B 細胞中 NF-?B 的活性. 細胞繼代到 96 孔盤生長到全滿,LPS ( 5µg/µL),以胎牛血清以 1:4 的比例稀釋,並以 37℃ 水浴加溫,作用 1 小時 30 分使 LPS 活化,以一倍 DMEM 稀釋,濃度為 0.1、1、5、10、25、 50、100 ng/mL 的 LPS 溶液加入細胞中,於 37℃培養箱,作用 16 小時後測量冷光的活性。LPS 25.
(26) 在濃度大於 1 ng/mL 後,開始活化 HepG2 細胞的 NF-?B 轉錄活性;到了濃度為 10 ng/mL 時, 約可活化細胞中的 NF-?B 活性達 2 倍(圖九),在更高的濃度仍然可以持續活化細胞中的 NF--?B 活性;LPS 濃度為 100 ng/mL 時,NF-?B 約可活化 2.5 倍。以上的實驗經三次重複後, 決定以濃度 20 ng/mL 的 LPS,作為 HepG2 細胞中,抗 LPS 誘發 NF-?B 活性藥物的搜尋起點。. 第七節. 543 種藥物對 HepG2/NF-?B 細胞中 LPS 誘發的 NF-?B 活性之影響. 細胞繼代到 96 孔盤經過 24 小時後,加入以一倍 DMEM 稀釋的藥物與 LPS,使其最終濃 度為含有千分之一稀釋的藥物和含有 20 ng/mL 的 LPS,經 16 小時後,收取細胞溶解液測定冷 光活性,並以 LPS 所活化的平均倍率作為基準(1 倍) ,表示藥物對 LPS 所活化的 NF-?B 活性 影響(圖十) ,並以能夠抑制 LPS 誘發 HepG2/NF-?B 細胞中 NF-?B 的活性,排名前 54 名的藥 物(結果在此不顯示)作為以下抗乙醛誘發 HepG2/NF-?B 細胞中 NF-?B 的活性藥物搜尋的起 點。. 第八節. 乙醛可以誘發 HepG2 細胞中 NF-?B 的活性. 在較早期的前驅實驗中,選定以一倍 DMEM 稀釋成濃度為 0、50、100、200 µM 的乙醛與 HepG2/NF-?B 細胞作用觀察乙醛在 1、4、8 小時對細胞中 NF-?B 活性的影響發現以 8 小時, 50 µ? 的狀態下對 HepG2 細胞中 NF-?B 的活化效應最佳(結果在此不顯示);在後續的實驗 中,HepG2/NF-?B 細胞以濃度為 50 µM 的乙醛刺激後,在 1、4、8、16、24 小時收集細胞的 溶解液,以 Reporter assay 的方法測量冷光的強度,觀察乙醛對 HepG2 細胞中 NF-?B 的活化 效應。在 HepG2/NF-?B 細胞中 NF-?B 的活性活化作用;乙醛在 4 小時,100 µM 的濃度下可 以活化 NF-?B 達到 1.9 倍;到了八小時,50 µM 時活化程度達到最高(圖十一,B)。乙醛活 化細胞中 NF-?B 的活性可以維持到 48 小時(圖十一,B) 。為了進一步了解在八小時,何種濃 度的乙醛可以活化細胞中 AP-1 和 NF-?B 的活性達到最高,於是選用濃度為 1、5、10、25、 50、100 、200 µ? 的乙醛作用;在 NF-?B 的部分:以 10 µM 的乙醛效果較佳(圖十一,A)。. 26.
(27) 第九節. 54 種藥物對乙醛誘發 HepG2 細胞中 NF-?B 活性的影響. 由(圖十一)的結果,得知乙醛可以誘發 HepG2 細胞中 NF-?B 的活性,於是從 543 種藥 物(圖四)對 LPS 所誘發 HepG2 細胞中 NF-?B 活化的影響,在以上抑制的 LPS 誘發 HepG2/NF-?B 細胞中 NF-?B 的活性,排名前 54 名的藥物;包含中藥的甲醇萃取液、各種不同 成份之生藥主成分(含黃酮類、生物鹼、皂素類、多酚類等成份)以及抗癌藥物(結果在此不 顯示) 。藥物的作用濃度,分為 1/1000 (v/v),25 µ? 及 1 µ? 。乙醛濃度則為 50 µM,加入細胞 中培養 8 小時後,以 Reporter assay 測定藥物對細胞中 NF-?B 活性的影響(圖十二) ,在以上 54 種藥物中,黃連、黃柏、和小檗鹼都能抑制肝細胞中乙醛誘發的 NF-?B 活性,尤其以小檗鹼 的效果最好,相對 EC50 濃度約為 18.0±1.8 µM(結果在此不顯示);由於黃連、黃柏等植物內 主要活性成分為小檗鹼(4~7 %),所以決定選出小檗鹼做以下的研究。. 第十節. 小檗鹼可以抑制 HepG2 細胞中被乙醛誘發的 NF-?B 的活性. 細胞培養於 25T flask 中,分別加入一倍 DMEM 稀釋,濃度為 1、5、25、50、100 µM 的 小檗鹼 30 分鐘後,加入濃度為 5 µ? 的乙醛,經過八小時後以 Reporter assay 測定細胞中轉錄 因子的活性,小檗鹼在濃度為 1 µM 時即能開始抑制細胞中 NF-?B 的活性;其抑制細胞中 50 % NF-?B 活性的濃度(EC50 )為 18.0±1.8 µM(圖十三) ,由實驗可知小檗鹼對乙醛誘發肝細胞中 NF-?B 的活化具有抑制作用。. 第十一節. 小檗鹼可以抑制 HepG2 細胞中乙醛誘發之 I-?B 磷酸化. 乙醛在 HepG2 細胞中活化 NF-?B 上游之訊息傳導路線,主要是經由 I-?B 的磷酸化,並 造成 NF-?B 分離,使 NF-?B 由細胞質進入細胞核發揮活性(Juan et al., 2000)。由以上 reporter assay的結果,得知小檗鹼可以抑制細胞中乙醛所誘發的 NF-?B 活化。為了了解小檗鹼對 NF-?B 上游之訊息傳導路線的影響,分析細胞中 I-?B,I-?B 磷酸化及 ERK 等蛋白質的變化,在預先 加入濃度為 1、5、25、50、100 µM 的小檗鹼 30 分鐘後,加入濃度為 5 µ? 的乙醛刺激並以. 27.
(28) western blot 偵測細胞中訊息傳導分子的變化。乙醛可以促進細胞中 I-?B 磷酸化蛋白質量上 升,並促使 I-?B 的蛋白質量下降(圖十四),而小檗鹼從 5 µM 開始即可明顯抑制乙醛所誘發 的 I-?B 磷酸化,但對 I-?B 的蛋白質量沒有很明顯的影響(圖十四)。. 第十二節. 小檗鹼可以抑制 HepG2 細胞中乙醛所誘發的 IL-1ß 及 TNF--a 的表現. 細胞在加入以一倍 DMEM 稀釋,濃度為 1、5、25、50、100 µM 的小檗鹼 30 分鐘後,加 入濃度為 10 µM 的乙醛培養 48 小時,取上層之培養基以 BD 的 OPtEIAT M human TNF-a 和 OPtEIAT M human IL-1ß 套組,測量細胞分泌 IL-1ß 及 TNF-a 蛋白質到細胞外的量,乙醛可以 誘發細胞分泌 IL-1ß 及 TNF-a(圖十五、圖十六)。小檗鹼可以明顯抑制乙醛誘發的 IL-1ß 及 TNF--a 蛋白質的的表現(圖十五、圖十六) 。計算方式為:使用內插法運算,將樣品呈現的吸 光度和標準品在 405 nm 下呈現的吸光度做比較加以換算,轉換成濃度單位(pg/mL)。. 28.
(29) 第六章 第一節. 討論 結果討論. 近年來細胞的轉形的過程與轉錄因子之間的關係已經漸漸為人所了解,轉錄因子AP-1在細 胞生長分裂的過程中扮演重要的角色;在小鼠的上皮細胞株(JB6)中TPA或是Okadaic Acid等腫 瘤促進劑或EGF等生長因子,可以透過 (RTK)/Ras/Raf/PKC/MEK/MAPK的訊息傳導路線活化 AP-1 (Guangming et al., 2001;Ruchika et al., 2001; Liu et al., 2002; Eric et al., 2002;Liu et al., 2001)。此外為了了解小鼠上皮細胞在受到Okadaic acid、TPA等腫瘤促進劑刺激,導致細胞轉 形,主要透過MAPK中何種路徑活化;送入Dominant negative c-jun (TAM 67)到JB6細胞中,可 以抑制細胞的轉形(Eric et al., 2002),並且促使一些受AP-1調控的基因被抑制。另外,當小鼠 體內缺乏c-Jun N-terminal Kinase-1(JNK-1)時,會使得TPA促進的皮膚癌的發生受到抑制 (Nanyue et al., 2002)。在實驗室之前的研究中發現:在不同癌化程度的肝細胞中,TPA主要會 透過ERK磷酸化的訊息傳導路線,促使細胞內AP-1的活性上升促進細胞內AP-1和DNA 的結合 能力提高最後導致Soft agar上細胞的轉形(賴,2001) 。而B型肝炎的X蛋白質(X protein) ,或 C型肝炎的核心蛋白質(core protein)在基因轉殖鼠的體內表現時,會活化ERK和JNK的路徑 並造成和肝細胞增生相關的細胞素如:TNF-a、 IL-1ß、IL-6等基因表現,並伴隨肝實質癌症 的發生(Tsutsumi et al., 2002; Arsura et al., 2003)。為了了解肝癌的發生主要透過的訊息傳導 路線,加入對ERK上游蛋白質激? MEK-1專一性的抑制劑:PD98059則可抑制肝癌的發生 ( Ruchika et al., 2001)與TPA在促進肝細胞轉形的機轉是相近的。這也說明了使用TPA作為肝細 胞轉形的促進劑,作為相關藥物搜尋與新藥開發的可行性。在抗肝細胞轉形的藥物搜尋系統 中,可以觀察到某一些科別的植物具有抑制TPA誘發AP-1活性的趨勢,可能和這些植物所含有 的物質有關;例如:毛茛科(Ranunculaceae)、小檗科(Berberidaceae)的植物具有近源性, 其韌皮部含有以小檗鹼為核心的生物鹼類似物,可能是抑制肝細胞中AP-1活性的原因。另外 繖型科的植物:柴胡、當歸、羌活等也可以抑制TPA所誘發的AP-1活性,原因之一可能和此科 植物內富含皂素類成份的化合物,此類化合物在細胞中可以透過和類固醇荷爾蒙的接受器結. 29.
(30) 合,進入細胞核內和AP-1複合體產生相互拮抗的作用(Wu, 2003) 。另外在不同科別的植物中, 含有同樣的活性物質,也是抑制AP-1活性的原因之一。例如毛茛科的黃連和芸香科的黃柏可 以抑制TPA所誘發的AP-1活性,而這兩種植物所共同含有的生物鹼-小檗鹼也有同樣的效果, 進一步說明了此搜尋系統的對於偵測微量物質的靈敏度。 轉錄因子NF-?B,與許多和發炎作用相關的細胞素基因的啟動有關,如:TNF-a、IL-6、 IL-1ß、IL-8、TGF-ß的啟動子區域,具有NF-?B的結合序列。一些物質;如細菌性內毒素LPS、 ceramide或是TNF-a等則會經由結合特定的接受器並經由PKC/Rac/NIK/IKK的訊息傳導路線, 促進I-?B的磷酸化和分離,並促使由細胞質進入到細胞核中。NF-?B的活化與肝病在體內的試 驗所呈現的相關性;報告中指出在臨床上已確定罹患肝癌的病人其發生病變的組織中, NF-?B、AP-1與STAT等蛋白質與DNA結合能力比正常的肝組織高許多(Liu et a l., 2002)。另外, 酒精代謝物乙醛對HepG2細胞造成氧化傷害的作用中,會透過PKC的訊息傳導途徑,造成細胞 中AP-1和NF--?B的活化並使得這兩個轉錄因子與DNA的結合能力上升(Juan et al., 2000),並且 會和HBV的X蛋白質與HCV的核心蛋白質產生協同作用,促進TNF--a的大量表現加重肝細胞 的炎症作用(Won et al., 2001)。在發炎性的細胞素中;TGF-ß會透過結合細胞膜表面TGF-ß的接 受器,啟動一連串訊息傳導路線,並使肝儲脂細胞分泌大量第一型膠原蛋白誘發肝纖維化的產 生(Chen et al., 2000;Border and Noble, 1994;Bedossa and Paradis, 1995)。此外TGF-ß的活化, 也會經TAK1/IKK 的訊息傳導路線,促進肝細胞中NF-?B的活化,並抑制經AP-1/SMAD活化 誘導的細胞自殺作用(Arsura et al., 2003)。 TNF-a 除了會導致細胞中NF-?B的活化,並經由與 TNF-a接受器結合,啟動JNK的訊息傳導路線,導致細胞凋亡作用的發生,也可能是肝臟產生 發炎的原因之一。酒精及其代謝物乙醛和細菌性內毒素LPS等,對肝細胞造成炎症作用,原因 為這些物質會促進細胞生產生長因子與細胞素 TNF-a 及IL-1ß 的產生(Lotz et al., 1989; Friedman et al., 2000; Pennington et al., 1997)。在酒精性肝炎的組織和血清中也有類似的作 用。另外,金屬離子或H2O2等物質對肝細胞造成的氧化傷害作用中;也會觀測到TNF-a、IL-8 等細胞素的表現(Dong et al., 1998a;Dong et al., 1998b;Cao et al., 2002)。進一步說明了在肝病 的發生過程中,氧化傷害的刺激,會造成發炎性細胞素的表現之間的相關性。本研究中,首先. 30.
(31) 利用LPS誘發HepG2細胞中NF--?B的活性,作為抗肝細胞炎症作用的藥物大量搜尋模式,觀察 了500多種藥物的作用圖譜。從以上的大量搜尋得到的結果中發現一些藥物會同時抑制肝細胞 中AP-1與NF-?B的活性,其可能與植物中所含的物質,會同時干擾AP-1與NF-?B上游的的訊息 傳導路線有關。另外一些植物或純物質會抑制這兩個轉錄因子之一的活性,但對另一個沒有影 響,可能也與藥物中所含的活性物質的成份有關。 在抑制乙醛誘發 HepG2 細胞中 NF-?B 的活性的藥物搜尋之中,觀測到黃連、黃柏及小檗鹼具 有抑制乙醛誘發的 NF-?B 的活性,小檗鹼為毛茛科植物黃連或芸香科植物黃柏的根莖、或韌 皮部所分離的生物鹼。具有抑制癌細胞生長、抑制炎症作用、抗金黃色葡萄球菌抑制 TPA 誘 發的小鼠皮膚的發炎作用以及抗下痢的功能(Kuo et al., 1995;Yasukawa et al., 1991;Anis et al., 2001)對於四氯化碳所引起的齧齒類動物的肝纖維化(Janbaz and Gilani , 2000),和肝功能損 害,皆有治療的功能。另外在大鼠的巨噬細胞中,小檗鹼具有抑制革蘭氏陰性菌的脂多醣體 (Lipopolysaccharide, LPS)所誘發的 NF-?B 活化的功能;並且抑制 LPS 所誘發的 TNF-a 的表 現(Kang et al., 2002) 。而在人類血癌細胞株中,小檗鹼具有促進細胞產生細胞凋亡的能力(Kuo et al., 1995)。 在此我們希望藉由相關的研究,了解小檗鹼在抑制酒精性肝炎發生的細胞模式中詳細的分子機 轉;並且為以後相關的新藥開發,提供一個更快速、高靈敏度的搜尋平台和療效指標。 由於小檗鹼為黃連等植物中之指標成份;在接續的實驗中,也發現小檗鹼對乙醛誘發 NF--?B的活性具有抑制的作用。對於炎症作用相關細胞素:TNF-a 及 IL-1ß的表現具有抑制的 功能。此外;小檗鹼會干擾細胞內I-?B磷酸化蛋白質的量,進一步抑制相關轉錄因子及下游基 因的表現。未來希望能夠了解小檗鹼是否會對轉錄因子與DNA的結合能力造成影響,並進一 步研究小檗鹼在體內是否也發揮同樣的生物活性。. 31.
(32) 第七章. 第一節. 結論與建議. 結論. 在本研究中,使用不同癌化程度的重組人類肝細胞株,偵測不同誘發劑誘發細胞中轉錄 因子活性,代表特定肝病模式。觀察藥物對AP-1與NF-?B的影響,並選擇具有抑制轉錄因子活 性,具新藥開發潛力的植物萃取物或化合物作為研究的目標。在諸多搜尋之中,小檗鹼具有抑 制乙醛誘發之NF-?B的活性,原因為其能干擾上游訊息傳導路線的活化,並抑制轉錄因子控制 下游基因的表現,對細胞生長造成抑制。希望藉由此模式作為未來研究的起點,進一步推展到 體內試驗,希望小檗鹼也可以發揮類似的功能. 第二節. 建議. 未來將進一步研究小檗鹼和乙醛在分子生物學上對NF-?B的作用,了解NF-?B在此扮演的 角色,以及對被這兩個轉錄因子所調控的基因做研究,釐清作用發生的原因與結果,並推動相 關的動物實驗,觀察小檗鹼在生物體內產生的作用是否也能和體外的模式一樣具有類似的功 能,希望小檗鹼可以對肝病的預防和治療有所貢獻並且為未來新藥開發的過程提供一個良好的 範本。. 32.
(33) 麥門冬、大棗、製半夏、茯神、枸杞子、香附子、燈心草、酸棗仁、淡豆豉、五味子、生薑皮、石膏、沙苑子、鵝不食草、荷葉、皂刺、女貞子、筆仔草、. A. 金銀花、藿香、蓮鬚、板藍根、茺蔚子、炮乾薑、山楂、當歸、黃精、郁李仁、滑石、木瓜、魚腥草、熟地黃、赤茯苓、雞血藤、菊花、枳殼、大腹皮、川 牛膝、五靈脂、車前子、黑薑、艾葉、薄荷葉、當歸尾、韭菜子、枳具子、雞香藤、冬瓜子、谷精、漏蘆、海藻、青蒿、大薊、地龍、忍冬藤、石決明、鳥 不宿、秦皮、旋覆花、菟絲子、炙黃耆、補骨脂、蘇子、桔梗、夜交藤、烏梅、神麴、射干、石菖蒲、澤蘭、槲寄生、淫羊藿、知母、蒲公英、豬苓、甘草、 白朮、麻黃、黃芩、赤芍、栝樓仁、栝樓根、枇杷葉、石斛、升麻、綿茵陳、炒梔子、山梔子、水丁香、鱉甲、萬點金、車前草、茯苓皮、桑螵蛸、地榆、 馬蹄金、丁香、白荳蔻、小金櫻、劉寄奴、萬年松、瞿麥、藕節、故紙花、檀香、化石草、荊芥、小茴香、北耆、羌活、益智仁、縮砂仁、紅花、萹蓄、百 合、桂枝、細辛、蘆根、五加皮、土茯苓、炙甘草、柴胡、北茵陳、訶子、黨參、前胡、杏仁、厚朴、昆布、白芍、蒿本、生地黃、製附子、澤瀉、薑黃、 地骨皮、薏苡仁、決明子、川丹皮、杜仲、山椒、牡蠣、玉竹、天麻、烏梅、栝樓實、炒火麻仁、合歡皮、山豆根、半夏麴、白頭翁、獨活、肉蓯蓉、鬱金、 蒲黃、白茅根、降真香、金錢草、九層塔、荔枝核、牽牛子、懷牛膝、松節、紅參鬚、柏子仁、三稜、夏枯草、扁柏、冬葵子(冏麻子)、豬簽草、益母草、 巴戟天、半枝連、木通、龍膽草、伏苓、山梔子、狗脊、鉤藤、沒藥、紫蘇葉、牛蒡子、何首烏、天門冬、蒼朮、當歸、黃水茄、荊芥穗、川楝子、炒槐花、 枳實、沙參、炙甘草、連翹、遠志、秦艽、白鮮皮、乾生薑、苦參根、紅參、川芎、白芨、阿膠、海蛤粉、麻黃根、銀杏葉、地榆炭、蒲黃炭、槐花炭、荊 芥炭、白雞冠花、倒地蜈蚣、敗醬草(菥蓂)、馬鞭草、紫蘇梗、紫花地丁、鎖陽、茜草、木賊草、三七、谷芽、紅骨蛇、 海螵蛸、蔓荊子、威靈仙、蒺藜、 白扁豆、葛根、浮小麥、龜板、蟬蛻、新木香、烏藥、桑白皮、延胡索、製南星、乾薑、辛夷、雞內金、粉防己、貝母、馬兜玲、旱蓮草、蛇床子、乳香、 萊服子、路路通、竹茹、 青皮、丹參、水梔子、 桑葉、紫苑、葶藶子、紫草、 龍骨、 蒼耳子、葛花、骨碎補、橘紅、一條根、 白蘞、赤小豆、貫眾、 王不留行、佛手、商陸、密蒙花、絲瓜絡、通草、馬勃、莪朮、桑枝、陳皮、桃仁、檳榔、 芡實、甜菊葉、厚朴、麻黃、檳榔、大黃、苦參、皂莢、甘草、 知母、吳茱萸、防己、 麻黃、柴胡、 木通、 豬苓、秦艽、白芷、防己、乾薑、吳茱萸、石膏、知母、山梔子、桑白皮、玄參、百合、 杏仁、貝母、天花 粉、紫苑、厚朴、枳實、赤芍藥、桃仁、澤蘭、當歸、川芎、白殭蠶、山茱萸、萆薢、蒿本、蒼耳子、白茅根、紫草、款冬花、薤白、白芍藥、淫羊藿、狗 脊、烏梅、白鮮皮、龍眼肉、合歡皮、海蛤、蟬蛻、射干、桔梗、半夏、天南星、旋覆花、萹蓄、刨附子、白芨、皂莢、薄荷、土茯苓、板藍根、蛇舌草、 半枝蓮、蒲公英、海藻、大青葉、龍膽草、大黃、白頭翁、連翹、青蒿、苦參、敗醬草、魚腥草、金銀花、荊芥、升麻、葛根、檳榔、牡丹皮、黃蓮、黃柏、 黃芩、莪朮、三陵、蛇床子、秦皮、苦楝子、五味子、夏枯草、槐花、甘草、陳皮、茵陳蒿、茯苓、延胡索、黃耆、人參、枸杞、 澤瀉、 麥門冬、 燈心 草、丹參、 逍遙散、 加味逍遙散、四逆散、 加味四逆散、 薄荷、 炙甘草、 炒白芍、 茯苓、 丹參、 虎杖、 枳實、 紫草、 柴胡、 當歸、 炒白朮、 火畏生薑、 白茅根、敗醬草(菥蓂)、白皮耆、蕤仁、柿蒂、蓮子芯、覆盆子、清葙子、肉荳蔻、槐花、刺五加、土茯苓、大青葉、玄參(元蔘)、薤白、白 芥子、地膚子、石榴皮、大金櫻、千里光、代赭石、麥芽、常山、仙鶴草、淡竹葉、續斷、白前、黃柏、 桂皮、天仙茅、百部、厚朴、大黃、絞股藍、雞 骨草、佩蘭、龍眼花、故紙花、胖大海、番泄葉、千年健、全蠍、蘇木、款冬花、香薷、吳茱萸、蓮子、栝樓皮、 白薇、 鳳尾草、 水蛭、 紫河車、 虻 蟲、 酒大黃、 黃連、 萆薢、 椿根白皮、 血竭、 藤瘤、 川椒、 白果、 丁豎朽、 虎杖、 五倍子、 草荳蔻、 海帶、 定經草、 白花蛇舌草、 山藥、 黃金桂、 大丁黃、 橘皮、 薏仁、 山藥、 靈芝、 槲寄生、 乾薑、 貝母、. B. Quercetin、 Rutin、 Hyperin、 Kaempferol、 Morin、 Galangin、 Silibinin、 3-Hydroxyflavone、 Baicalein、 Baicalin、 Chrysin、 Luteolin、 Apigenin、 Flavone、 Hesperetin、 Hesperidin、 Eriocitrin、 Naringenin、 Naringin、 (+)-Catechin、 (-)-Epicatechin、 Gentisin、 Puerarin、 Formononetin、 glycyrrhizin、 18-a-Glycyrrhetinic acid、 18-b-Glycyrrhetinic acid、 Stigmasterol、 Ginsenoside Rb1、 Ginsenoside Rb2、 Ginsenoside Rc、 Ginsenoside Rd、 Ginsenoside Re、 Ginsenoside Rg1、 Silymarin、 berberrin、 Berberine-Cl、 Berberine-SO4、 Emodin、 Rhein、 Paeoniflorin、 Paeonol、 Evodiamine. C. 5-Azacytidine、 5-Fluorouracil、 Aminopterin、 Camptothecin、 Cis-platinum(II) diamine dichloride、 Colchicine、 Cystine β-D-arabinofuranoside、hydrochloride、 Daunorubcin-hydrochloride、 Etoposide、 Mitomycin C、 Paciltaxel、 Vinblastine sulfate salt、 Vinblastine sulfate salt、 Sandoststin、 Forane、 Diprivan、 Ultane. 圖四、本實驗使用的藥物庫名單分為三類。 (A) 常用之中藥的甲醇萃取物,(B) 植物主要活性物質,(C) 常用之抗癌藥物和麻醉劑等。 33.
(34) 8. 0.7. 7. 0.6. 6 5. 0.4. 4 0.3. 3. 0.2. 2. 25. 50. 100 200. 200. 10. 100. 5 5. 50. 1. 25. 0.5. conc(ng/mL). TPA (ng/mL). 3. 0.7. 2.5. 0.6 0.5. 2. 0.4 1.5 0.3 1. OD 570 nm. Relative AP-1 activity fold. 0.2. 10. 0 1. 0 0.5. 0.1 0.2. 1. Mock. B. OD570nm. 0.5. MOCK. Relative AP-1 activation fold. A. 0.2. 0.5. 0.1. 0. 0 Mock. 0.2. 1. 5. 25. 50. 100. concentration (ng/mL) TPA (ng/mL). 圖五、TPA 可以誘發 Chang liver/AP-1 細胞和 HepG2/AP-1 細胞中 AP-1 的活性。 。 (A) Chang liver/AP-1 細胞(B)HepG2/AP-1 細胞在去除血清的培養基培養,經過 24 小時後,以濃度為 0.2、0.5、1、5、10、25、50、100 ng/mL 的 TPA 刺激細 胞,經過 16 小時後在 Reporter assay 中呈現的 AP-1 活化劑量作用;直條圖為 冷光的相對活性,折線圖則為 MTT assay 中細胞的活性。以上實驗為兩次實 驗的平均值±標準差。 34.
(35) 35.
(36) 36.
(37) 37.
(38) 表一、在 Chang liver/AP-1 及 HepG2/AP-1 細胞中皆能抑制 TPA 誘發 AP-1 活性的中藥。 1. 名稱. 2. 柴胡. 繖形. 科別 3 EC50 18.77. 4. 指標成分. Saikosaponin a, b, c, d. 5. 應用複方. 當歸龍薈丸, 茵陳大黃湯,舒經活血湯, 小柴胡湯,補中益氣湯,加味消遙散. 羌活. 繖形. Bergatene,. 8.7. isoimeratorin. 當歸龍薈丸、舒經活血湯、龍膽瀉肝湯、蘇. 當歸. 繖形. 11.1. 吳茱萸. 芸香. 11.4. Butylidenephthalide Evodiamine, Rutaecarpine, deoxyevodiamine. 黃柏. 龍膽. 芸香. 龍膽. 驅風解毒湯、九味羌活散. Berberine(EC 50 =24.4μM),. 35.9. 秦艽. 龍膽. 46.1. 黃連. 毛茛. 35.8. 檳榔. 棕?. 17.6. 厚朴. 木蘭. 28.47. 湯、補中益氣湯、廣茂潰堅湯、紫根牡犡湯 雞鳴散加茯苓、吳茱萸湯、當歸四逆加吳茱 萸生薑湯、廣茂潰堅湯 三黃瀉心湯、黃連解毒湯、當歸龍薈丸. Palmatine, Coptisine Gentipicrin, Amarogentin,. 57.8. 子降氣湯、秦艽飲、當歸四逆加吳茱萸生薑. 當歸龍薈丸、舒經活血湯、龍膽瀉肝湯. Loganin, Gentioside. Gentipicrin. 秦艽飲. Berberine(EC 50=24.4μM),. 三 黃瀉心湯,黃連解毒湯,當歸龍薈. Palmatine, Coptisine. 丸,廣茂潰堅湯. Guvacine, Isoguvacine,. 椒梅湯 ,雞鳴散加茯苓. Arecoline, Arecaidine 平胃散,胃苓湯 ,半夏厚朴湯,廣茂 β-Emdesmol, Magnolol,. 潰堅湯 ,蘇子降氣湯,大承氣湯. Honokiol. 大黃. 紫草. 蓼. 紫草. 73.27. 31.06. Emodin(EC50=30μM),. 茵陳大黃湯 ,三黃瀉心湯,黃連解. Aloe-emodin,(-)-Epicatechin,. 毒湯、當歸龍薈丸、大承氣湯、紫. (+)-Catechin, Rhein. 根牡犡湯. (EC50=34.3μM), Sennoside A, B Alkannin, Shikonin, (-)-Bomesitol, Allantonin. 丹參 槐花. Tanshinone,. 唇形. 21.09. 豆. 37.09. Salvianic acid Rutin(EC 50=44.9μM),. 紫根牡犡湯,加味消遙散 丹參飲,加味四逆散. 槐花散. Quercetin(EC 50=47.44μM). (1)名稱, (2)植物分類, (3)在 HepG2 細胞中到達 EC50 的濃度,中藥的甲醇萃取物的 38 濃度單位以(µg/mL)表示,(4)指標成分,藥物庫中擁有的成分其濃度單位為 (µM),(5)臨 床上應用的中藥複方。.
(39) 7. 0.6. 6. 0.5. 5. 0.4. 4 0.3 3. OD 570 nm. Relative NF-?B activity. Relative NF-kB activity. Luc MTT. 0.2. 2 1. 0.1. 0. 0 Mock mock. 0.1. 0.5. 1. 5. 10. 25. 50. 100. LPS concentration LPS (ng/ml)(ng/mL). 圖九、LPS 可以誘發 HepG2/NF-?B 細胞中 NF-?B 的活性。 細胞在繼代到 96 孔盤經 24 小時後,以一倍 DMEM 稀釋成濃度為 0.1、0.5、1、5、 10、25、50、100 ng/mL 的溶液作用 16 小時後,以 Reporter assay 測量 NF-?B 的活性, 並以未加藥的組別的平均值作為基準,觀察不同濃度的 LPS 造成的劑量作用。以上實 驗為三次實驗的平均值±標準差。. 39.
(40) 40.
(41) A. 3A. Relative NF-kB activity. Relative NF-?B activity. 2.5 2 1.5 1 0.5 0 mock. 1. 5. 10. 25. 50. 100. 200. conc (uM) B Relative NF-?B activity. Relative NF-kB activity. 4. 3. 2. 1. 0 1. 4. 8 Time (hour). 16. 24. 圖十一、乙醛可以誘發 HepG2 細胞中 NF-?B 活性的時間和劑量作用。 (A)乙醛在八小時誘發 NF-?B 的劑量作用,乙醛以一倍 DMEM 稀釋成 1、5、10、25、 50、100、200 µM 的濃度,在 8 小時所呈現的作用圖譜。(B) 50 µM 乙醛在 1、4、8、 16、48 小時對 NF-?B 的活化作用。(B)的結果為兩次實驗的平均值±標準差。(A) 的結果為三次實驗的平均值±標準差。 41.
(42) 42.
(43) 1 Relative NFkB activity. Relative NF-?B activity. 0.5. 0 mock Acet Ace5uM Mock 50 µM. 11 55 conc(uM) Berberine (µM). 25 25. 50 50. 100 100. 圖十三、小檗鹼可以抑制 HepG2 細胞中乙醛所誘發的 NF-?B 活性。 (A)小檗鹼可以抑制 HepG2 細胞中乙醛所誘發的 NF-?B 活性,以 0.1、1、5、25、 50、100 µM 的小檗鹼與 5 µM 的乙醛作用,在 8 小時呈現的作用圖譜。以乙醛 在 HepG2 細胞中誘發的 NF-?B 活化倍率作為基準。以上實驗值為三次實驗的平 均值±標準差。. 43.
(44) Acetaldehyde Berberine. mock. 5μM. -. + -. 11. + +. 5. 25. 50. + +. + +. + +. 100. Concentration(μM). + + I-?B. phospho-I-?B. ERK. 圖十四、小檗鹼可以抑制 HepG2 細胞中乙醛誘發的 I-?B 磷酸化。 細胞在預先以濃度為 1、5、25、50、100 µM 的小檗鹼處理 30 分鐘後,再加 入濃度為 5 µM 的乙醛作用 30 分鐘;觀察乙醛和小檗鹼對細胞的影響,ERK 蛋白質在此作為細胞中蛋白質總量的控制組。以上為一次實驗呈現的結果。. 44.
(45) TNF-α +1. A. κ 1 -587 -577. κ 2. κ 3. -210 -202. -98 -89 +89. -600. OD 405 nm. B. OD405 nm. 3’ -CCCGACAGGG-5’. 5’ -GGGGTATCCT-3’ 5’ -GGGTTTCTCC-3’. 3 2 1 0 0. 1. 2. 3.9. 7.8. 15.6. 31.2. 62.5. 125. 250. 500. TNF-a concentration (pg/mL) TNF-α (pg/mL). C 25. (pg/mL). TNF-a conc (pg/mL). α TNF(. 20 15 10 5. Berberine conc (uM). Berberine. 50. 100. 25. 50. 5. 25. 1. 5. 0.1. 1. Acet 10μM. 0.1. Mock. Ace 10uM. mock. 0 100. (μ M). 圖十五、小檗鹼在 48 小時可以抑制乙醛誘發 HepG2 細胞分泌 TNF-a (A)人類 TNF-a 基因啟動子區域中 NF-?B 的結合序列(B)TNF-a 的標準品 在 Cytokine ELISA 中呈現的標準曲線;橫軸為濃度(pg/mL) ,縱軸為在 450 nm 下呈現的吸光度(C)以濃度為 0.1、1、5、25、50、100 µM 的小檗鹼與 10 µM 的乙醛,作用 48 小時後,呈現以下的作用圖譜。以上實驗值為兩次實驗的平 均值±標準差。 45.
(46) A. 5’. NF-κB. 3’. GGGAAAAT B. -300. OD 405 nm. OD 405 nm. 3 2 1 0 0. 2. 3.9. 7.8 15.6 31.3 62.5 125 IL-1B concentration (pg/mL) IL-1β (pg/mL). 250. 500. 1000. 10. IL-1ß (pg/mL). IL-1B conc (pg/mL). C. 5. 5. 25. 50. 100. 1. 50. 5. 1. 0.1. 25. Mock Acet 10μM. 0.1. Ace 10uM. mock. 0 100. Berberine Berbeine conc(μM) (uM). 圖十六、小檗鹼在 48 小時可以抑制乙醛誘發 HepG2 細胞分泌 IL-1ß。 (A)人類 IL-1ß 基因啟動子區域中 NF-?B 的結合序列(B)IL-1ß 的標準品在 Cytokine ELISA 中呈現的標準曲線;橫軸為濃度(pg/mL),縱軸為在 405 nm 下呈現的吸光度(C)以濃度為 0.1、1、5、25、50、100 µM 的小檗鹼與 10 µM 的乙醛,作用 48 小時後,呈現以下的作用圖譜。小檗鹼在 48 小時可以抑制乙 醛誘發細胞分泌 IL-1ß。以上實驗值為兩次實驗的平均值±標準差。. 46.
(47) 參考文獻: 行政院衛生署。中華民國八十九年國人十大死因之調查。中華民國九十年。 行政院衛生署。中華民國八十八年國人十大死因之調查。中華民國八十九年。 賴羿如。肝細胞轉形之分子機制與分子療程之探討。中國醫藥學院醫學研究所碩士論文。中華民國九十年。 Abiru S, Nakao K, Ichikawa T, Migita K, Shigeno M, Sakamoto M, Ishikawa H. Acetaldehyde prevents nuclear factor-kappa B activation and hepatic inflammation in ethanol-fed rats. Lab Invest. 1999. 79: 799-806. Ahmad, N. and Mukhtar, H. Green tea polyphenols and cancer : biologic mechanisms and practical implications. Nutr Rev. 1999. 57: 78-83.. Anis KV, Rajeshkumar NV, Kuttan R. Inhibition of chemical carcinogenesis by berberine in rats and mice. J Pharm Pharmacol. 2001. 53: 763-768. Ann M. Bode, Wei-Ya Ma, Young-Joon Surh, and Zigang Dong.Inhibition of epidermal growth factor-a induced cell transformation and Activator Protein 1 activation by [6]-Gingerol. Canc Res 2001. 61: 850-853.. Anping Chen and Bernard H. Davis. The DNA Binding protein BTEB mediates acetaldehyde-induced, jun n-terminal kinase-dependent ai(i) collagen gene expression in rat hepatic stellate cells. Mol Cell Biol, 2000. 20: 2818-2826.. Anping Chen, Li Zhang, Jianye XU and Jun Tang. The antioxidant (-)-epigallocatechin-3-gallate inhibits activated hepaticstellate cell growth and suppresses acetaldehyde-induced gene expression. Biochem. J. 2002. 368: 695-704.. Arnott CH, Scott KA, Moore RJ, Hewer A, Phillips DH, Parker P, Balkwill FR,Owens DM. Tumour necrosis factor-alpha mediates tumour promotion via a PKC alpha- and AP-1-dependent pathway. Oncogene. 2002. 21: 4728-4738.. Arsura M, Panta GR, Bilyeu JD, Cavin LG, Sovak MA, Oliver AA, Factor V, Heuchel R, Mercurio F, Thorgeirsson SS, Sonenshein GE. Transient activation of NF-kappaB through a TAK1/IKK kinase pathway by TGF-beta 1 inhibits AP-1/SMAD signaling and apoptosis: implications in liver tumor formation. Oncogene. 2003. 22: 412-425. Bae, H. W., Geiser, A. G., Kim, D. H., Chung, M. T., Burmester, J. K., Sporn, M. B., Roberts, A. B. and Kim, S. J. Characterization of the promoter region of the human transforming growth factor-ß type II receptor gene. J Biol Chem. 1995. 270: 29460-29468. .Baraona, E., Matsuda, Y., Pikkarainen, P., Finkelman, F.,Lieber, C.S.,. Effects of ethanol on hepatic proteinsecretion and microtubules: possible mediation by acetaldehyde. Curr in Alco. 1981. 8: 421-434.. 47.
(48) Bedossa, P., Paradis, V.,. Transforming growth factor-b:a key-role in liver fibrogenesis. J Hepatol. 1995. 22: 37-42. Bird, G.L.A., Sheron, N., Goka, J., Alexandra, G., Williams,R.S. Increased plasma tumor necrosis factor insevere alcoholic hepatitis. Ann Intern Med. 1990. 112: 917-920. Bissell, D.M., Wang, S.S., Jarnagin, W.R., Roll, F.J. Cell-specific expression of TGF-ß1 in rat liver-evidence for autocrine regulation of hepatocytes proliferation. J Clin Invest. 1995. 33: 447-455.. Bode, C., Fukin, H., Bode, J.C.,. Hidden endotoxin in plasma of patients with alcoholic liver disease. Eur J Gastrol Hepatol. 1993. 5: 257-262. Border, W.A., Noble, N.A.,. Mechanisms of disease: transforming growth factor beta in tissue fibrosis. N Engl J Med. 1994. 331: 1286-1292. Cao Q, Mak KM, Lieber CS. Dilinoleoylphosphatidylcholine decreases acetaldehyde-induced TNF-alpha generation in Kupffer cells of ethanol-fed rats. Bio Biop Res Comm. 2002. 299: 459-464. Casini, A., Galli, G., Salzano, R., Rotella, C. M. and Surrenti, C. Acetaldehydeprotein adducts, but not lactate and pyruvate, stimulate gene transcription of collagen and Fibronectin in hepatic fat-storing cells. J. Hepatol. 1993. 19: 385-392. Chen A, Davis BH. Chi CW, Chang YF, Chao TW, Chiang SH, P'eng FK, Lui WY, Liu TY. Flowcytometric analysis of the effect of berberine on the expression of glucocorticoid receptors in human hepatoma HepG2 cells. Life Sci. 1994. 54: 2099-2107. Chen A, Zhang L, Xu J, Tang J. The antioxidant (-)-epigallocatechin-3-gallate inhibits activated hepatic stellate cell growth and suppresses acetaldehyde-induced gene expression. Biochem J. 2002. 368: 695-704. Chen J, Clemens DL, Cederbaum AI, Gao B. Ethanol inhibits the JAK-STAT signaling pathway in freshly isolated rat hepatocytes but not in cultured hepatocytes or HepG2 cells: evidence for a lack of involvement of ethanol metabolism. Clin Biochem. 2001. 34: 203-209.. Chen N, Ma WY, She QB, Wu E, Liu G, Bode AM, Dong Z. Transactivation of the epidermal growth factor receptor is involved in 2-O-tetradecanoylphorbol-13-acetate-induced signal transduction.J Biol Chem. 2001. 276: 46722-46728.. Chen N, Nomura M, She QB, Ma WY, Bode AM, Wang L, Flavell RA, Dong Z. Suppression of skin tumorigenesis in c-Jun NH(2)-terminal kinase-2-deficient ice.Canc Res. 2001. 61: 3908-3912.. 48.
(49) Chen, A. and Davis, B. H. The DNA binding protein BTEB mediates acetaldehyde-induced, jun N-terminal kinase-dependent aI(I) collagen gene expression in rat hepatic stellate cells. Mol Cell Biol. 2000. 20: 2818-2826. Chen, A. and Davis, B. H. UV irradiation activates JNK and increases aI(I)collagen gene expression in rat hepatic stellate cells. J Biol Chem. 1999. 274: 158-164. Chuanshu Huang, Yi Huang, Jingxia Li, Wenwei Hu, Robeena Aziz, Moon-shong Tang, Nanjun Sun, John Cassady, and Gary D. Stoner Inhibition of Benzo(a)pyrene Diol-Epoxide-induced Transactivation of Activated Protein-1 and Nuclear Factor kappa B by Black Raspberry Extracts. Canc Res. 2002. 62: 6857-6863. Chunfang, G., Gressner, G., Zoremba, M., Gressner, A.M., TGF expression in isolated and cultured rat hepatocytes. J Cell Phys. 1996. 167: 394-405. Chung YW, Jeong DW, Won JY, Choi EJ, Choi YH, Kim IY.H(2)O(2)-induced AP-1 activation and its effect on p21(WAF1/CIP1)-mediated G2/M arrest in a p53-deficient human lung cancer cell. Bio Biop Res Comm. 2002. 293: 1248-1253. Clemens DL, Forman A, Jerrells TR, Sorrell MF, Tuma DJ. Relationship between acetaldehyde levels and cell survival in ethanol-metabolizing hepatoma cells. Hepatol. 2002. 35: 1196-1204.. Colburn N. Transgenic mice demonstrate AP-1 (activator protein-1) transactivation is required for tumor promotion. Proc Natl Acad Sci U S A. 1999. 96: 9827-9832.. Dharwan. A, Young MR, Colburn NH. The role of AP-1, NF-kappaB and ROS/NOS in skin carcinogenesis: the JB6 model is predictive.Mol Cell Bio. 2002. 20: 185-193.. Dong Z, Cmarik JL, Wendel EJ, Colburn NH. Differential transformation efficiency but not AP-1 induction under anchorage-dependent and -independent conditions.Carcinog. 1994. 15: 1001-1004. Dong, W., Simeonova, P.P., Gallucci R. Cytokine expression in hepatocytes: role of oxidant stress. J. Interf Cyto Res. 1998a. 18: 629-638. Dong, W., Simeonova, P.P., Gallucci, R., et al.,, Toxic metals stimulate inflammatory cytokines in hepatocytes through oxidative stress mechanisms. Tox Appl Pharmacol. 1998b. 151: 359-366.. Eric J. Thompson, Jacalyn MacGowan, Matthew R. Young, Nancy Colburn, and G. Tim Bowden. A Dominant Negative c-jun Specifically Blocks Okadaic Acid-induced Skin Tumor Promotion. Canc Res. 2002. 62: 3044-3047. Felver, M.E., Mezey, E., McGire, M., Nathan, C. Plasma tumor necrosis factor alpha predicts long-termsurvival in severe alcoholic hepatitis. Alco Clin Exp Res.1990.114: 255-269. 49.
數據
相關文件
Xianggang zaji (miscellaneous notes on Hong Kong) was written by an English and translated into Chinese by a local Chinese literati.. Doubts can therefore be cast as to whether
identify different types of tourist attractions and examine the factors affecting the development of tourism in these places;.4. recognize factors affecting tourist flows and the
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
Calligraphy plays an integral role in the development of Buddhism, including the transcription of scriptures or the distribution of Buddhist words and phrases in writing,
(1) Skin melanoma cells show molecular alterations of the RAS-BRAF-MEK-ERK mitogen activated protein kinase (MAPK) signalling pathway, mediating uncontrolled proliferation of
194,195 In cases of medication- induced salivary gland hypofunction, the unstimulated whole saliva flow rate is usually significantly reduced, whereas the chewing- stimulated
Microphone and 600 ohm line conduits shall be mechanically and electrically connected to receptacle boxes and electrically grounded to the audio system ground point.. Lines in
