科技部補助專題研究計畫成果報告
期末報告
工作記憶在知覺意識所扮演的角色
計 畫 類 別 : 個別型計畫 計 畫 編 號 : NSC 101-2410-H-004-088- 執 行 期 間 : 101 年 08 月 01 日至 103 年 07 月 31 日 執 行 單 位 : 國立政治大學心智、大腦與學習研究中心 計 畫 主 持 人 : 徐慎謀 計畫參與人員: 學士級-專任助理人員:陳彥親 報 告 附 件 : 出席國際會議研究心得報告及發表論文 處 理 方 式 : 1.公開資訊:本計畫可公開查詢 2.「本研究」是否已有嚴重損及公共利益之發現:否 3.「本報告」是否建議提供政府單位施政參考:否中 華 民 國 103 年 10 月 31 日
中 文 摘 要 : 現今意識的研究通常只比較有意識及無意識的狀態以便尋找 負責意識的神經機制。但以辨認短暫出現的人臉的作業為 例,視覺意識至少可區分為只探測到人臉但不知其身份以及 可完全辨認其身份兩種表徵層次。利用磁波儀,此研究發現能 意識到人臉的存在以及能意識到人臉的身份會激發在空間及 時間上相異的神經活動,顯示視覺意識不是經由單一的神經 模式調控。相反地,刺激物的表徵層次會導致不同的意識機 制運轉。此外,我們的結果還發現要能從探測到人臉的層次 躍升到可完全辨認人臉身份的層次與特定的神經律動息息相 關。 中文關鍵詞: 意識、人臉、腦磁波儀 英 文 摘 要 : 英文關鍵詞:
Consciousness is both the most obvious and the most mysterious feature of our minds. One important issue in the empirical studies of consciousness is how an external piece of information gains access to conscious processing. Previous approach to this issue has often capitalized on investigating how brain activity differentially responds to supraliminally- and subliminally-presented stimuli (Dehaene, et al., 2001; Melloni, et al., 2007). However, a few lingering issues remain. First, it is still a matter of debate how to clearly define the distinction between aware and unaware states (Clifford, Arabzadeh, & Harris, 2008; Seth, Dienes, Cleeremans, Overgaard, & Pessoa, 2008; Wiens, 2006). More importantly, investigating the nature of conscious perception based on the concept of aware/unaware dichotomy implies that stimuli are processed in a discrete, all-or-none manner (Campana & Tallon-Baudry, 2013; Kouider, de Gardelle, Sackur, & Dupoux, 2010). From this perspective, one question arises regarding whether the brain responds to stimuli in a constant fashion once the consciousness network is triggered.
According to hierarchical view of stimulus representations (Craik & Lockhart, 1972), a face, for instance, has at least two levels of representations, ranging from a low-level representation of face features to a high-level representation of the face identity (Grill-Spector, Knouf, & Kanwisher, 2004; Liu, Harris, & Kanwisher, 2002). When viewing a briefly-presented face from trial to trial, participants might be able to access high-level representational contents (face conscious identification) in some trials, or only be able to access low-level representational contents (face conscious detection) in other trials. As such, by capitalizing on the hierarchical contents of face stimuli, this study investigated the neural mechanisms underlying low-level conscious perception (only successful detection but no recognition of a briefly-presented and masked face) as well as high-level conscious perception (successful detection and recognition of a briefly-presented and masked face).
We hypothesize that if the access to conscious perception depends on the levels of representations of stimuli, conscious identification and detection of faces should elicit a certain distinct neural signatures. In addition, we may also examine whether there is a common neural correlate associated with both conscious identification and detection. In turn, this correlate may be regarded as the process directly corresponding to conscious experience, irrespective of conscious contents.
MATERIALS AND METHODS Participants
Fourteen right-handed participants without past neurological or psychiatric history participated in the present study (8 males, mean age ± STD = 26.43 ± 3.11 years, range = 21 – 31). All participants had normal or corrected-to-normal vision and provided written informed consent.
Stimuli
Stimuli comprised of 56 different faces of the same target celebrity (Andy Liu) and 24 faces of different non-target celebrities. All faces are collected from the internet and are depicted in various viewing conditions and backgrounds to reduce the possibility that participants would use a small set of low-level features to perform the task. Additionally, 34 texture patterns were generated by scrambling randomly-selected face pictures into squares with a size of 1 x 1 pixels. Forward and backward masks were created in a similar way by scrambling the faces in between into squares with a size of8 x 8 pixels. The face images subtended a horizontal visual angle of 2.4° and a vertical angle of 2.7° around the center of the screen. The stimulus presentation was controlled using Psychtoolbox.
Procedure
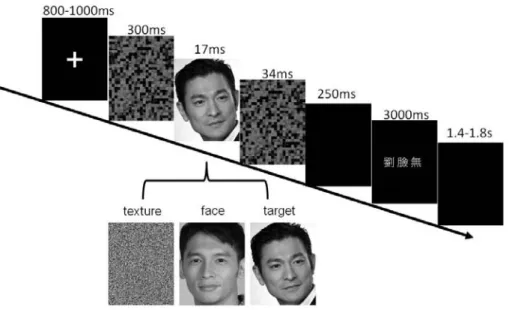
As shown in Figure 1, each trial started with the presentation of a fixation cross for 800-1000ms, followed by a forward mask for 300ms, a stimulus for 17ms, and a backward mask for 33ms. The stimulus was a pre-specified target face (Andy Liu), a face of another celebrity, or a texture. Next, after a blank of 250 ms, a response window was displayed with three choices. Participants had up to 3000ms to report that (1) they could identify the pre-specified target by selecting the option “Liu”, (2) they could detect the presence of a face but could not identify whether or not the face was the pre-specified target by selecting the option “Face”; (3) could not see a face by selecting the option “No”. The response choices were made with participants’ right index, middle and ring fingers via button press. The positions of the response choices were randomized across trials. There were 12-14 runs in the experiment. Each runs consisted of 28 pre-specified targets, 12 other celebrity faces, and 17 textures. Therefore, 30% of the total trials were catch trials for conscious detection of faces (17 texture-present trials out of all 57 trials), whereas 30% of the face-present trials were catch trials for conscious identification of faces (12 celebrity-face trials out of 40 face-present trials). Prior to the experiment, mask intensities were adjusted for individuals to obtain the luminance threshold of the masks, so that the face identification rates would be above 15%. The experimental paradigm was the same as described above and each individual was administered 1-3 runs. The false alarm rates of face detection and face identification were controlled under 10 % in both the calibration and experimental sessions.
Figure 1. Experimental paradigm.
MEG recording
The MEG data were recorded using a whole-head system comprising 157-axial-gradiometer channels (Yokogawa, Co., Tokyo). The signals were digitized at 1000 Hz and filtered with 0.3 Hz high-pass and 500 Hz low-pass cutoff and a 60 Hz notch. To minimize head movements between runs, the participants’ head positions relative to the MEG sensors were monitored using a set of head localization coils placed at the nasion and the left and right ear canals. FieldTrip and MATLAB (MathWorks, Natick, MA) software were used for data preprocessing, analysis and visualization.
Analysis
MEG signals were segmented into 2000-ms epochs starting from 1000-ms before stimulus onset. Trials contaminated with eye movements over 2.5 degrees, eye blinks, and muscular artifacts were rejected through visual inspection and semi-automatic functions implemented in Fieldtrip. The remaining trials were then submitted to the following analyses.
For event-related field (ERF) analysis, the data were first resampled at 250 Hz and digitally filtered with a 50 Hz low-pass filter. Next, the data were averaged across trials for each experimental condition and each participant. The averaged activity was baseline-corrected by subtracting the mean activity during the baseline period (300 to 100 ms preceding the onset of the forward mask) for each data point.
For spectral analysis, time-frequency representations of power were computed using Morlet’s wavelets (m = 7) on every senor-frequency (8-100 Hz, step: 2 Hz)-time data (-400 ms to 1000 ms, step: 10 ms) point in each trial. The obtained signal power was then averaged across trials for each experimental condition and each participant. To compensate for the 1/f decay, the averaged power activities were normalized to a
baseline from 400 to 200 ms preceding the onset of the forward mask. The normalization involved calculating the 10log transform of the power ratio of each data point to the mean baseline power on a frequency-by frequency basis.
The phase-locking of spectral signals was quantified by the inter-trial coherence (ITC). ITC was computed by normalizing the lengths of the complex Fourier coefficients (resulting from wavelet transformation) to one and then computing their average across trials. To control for biases due to difference in trial numbers, the ITC was transformed to Rayleigh Z values, Z = N x ITC2, where N is the trial number (Bonnefond & Jensen, 2012).
Statistics
To determine whether the power differences between experimental conditions were significant, cluster-based permutation tests were conducted. This statistical test does not require specific assumptions about the shape of the population distribution and allows for controlling multiple comparison problems. In the tests, the experimental differences were quantified by means of paired t-tests for every sample from -400 to 1000 ms. The samples were selected based on t values larger than the threshold (p < 0.05). The selected samples were then clustered in connected sets on the basis of spatial, temporal or frequency adjacency. The cluster with the maximum sum of the t values was used as test statistic. A distribution was then generated by randomly permuting the data across the conditions within each participant and recalculating the test statistic 1000 times using a Monte Carlo estimate. Lastly, the p values were determined by evaluating the proportion of the obtained distribution resulting in test statistic larger than the observed statistic.
RESULTS
All our analysis focused exclusively on pre-specified target faces (Andy Liu). This procedure may allow for examining how conscious perception alternated in the face of identical visual stimulation. The target trials, during which the stimuli were Andy Liu’s faces, were sorted into three experimental conditions: (1) FI: the trials in which participants could recognize the facial identity of the targets (mean number of trial ± SEM = 55 ± 6), (2) FD: the trials in which participants could not recognize the
identity of the targets, but could detect the presence of the target faces (193 ± 17), and (3) FN : the trials in which participants could not detect the presence of the target faces (96 ± 14).
Behavioral results
The hit rates and false alarms in each experimental condition were calculated based on the definition described in the table below (Grill-Spector, Knouf, & Kanwisher, 2004):
Both conscious identification and detection of faces was above the threshold (d' identification = 1.44 ± 0.11, paired t test against zero, t(13) = 13.04, p < 0.001; λ’ identification = 1.75 ± 0.07, t(13) = 27.04, p < 0.001; d' detection = 2.72 ± 0.13, t(13) = 22.08, p < 0.001; λ’detection = 1.16 ± 0.09, t(13) = 13.56, p < 0.001).
No significant RT difference was found among the FI (944.29 ± 50.91 ms from
response screen onset), FD (903.28 ± 63.68), and FN (944.29 ± 50.91) trials (one-way repeated measures ANOVA after sphericity correction, F(2, 26) = 0.68, p = 0.4).
Planar gradients of ERFs
A neural correlate that represented conscious identification was identified by contrasting the planar gradients between the FI and FN conditions. The planar gradients were stronger in the FI condition than the FN condition over posterior sites between 560 to 570 ms after the onset of the first mask (cluster-based permutation test: p = 0.05, Figure 2A). For conscious detection, a separate neural correlate was
observed (p = 0.047, Figure 2B) after contrasting the planar gradients between the FD and FN conditions. The planar gradients in this correlate were higher in the FN relative to the FD condition over the right posterior sites during the 300 to 380 ms period.
FI - FN FD - FN
Figure 2. The scalp topographies of the differences of the planar gradients of ERFs between
the FI and FN conditions (A) and the FD and FN conditions (B). The color scales in the scalp topographies are expressed in t statistic values. The back dots in the scalp topographies highlight the clusters of sensors showing significant differences.
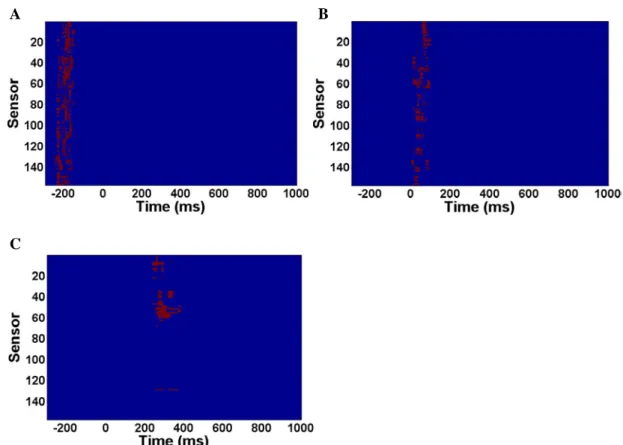
We proceeded to investigate the neural correlate reflecting the transition from conscious identification to conscious detection by contrasting the FI and FD
conditions. The results showed a rhythmic occurrence of the differences of the planar gradients (Figure 3). The stronger planar gradients in the FI relative to the FD
condition first occurred from -250 to -150 ms over most of the sensors (Figure 3A).
The difference of planar gradients were observed again between the 0 to 100 ms period over most of the sensors (p = 0.01, Figure 3B) and 240 to 380 ms period over the left middle sensors (p = 0.02, Figure 3C).
Figure 3. The sensor-time representations of the differences of the planar gradients between
the FI and FD conditions. Three significant clusters were identified in this contrast. Red dots in the figures indicate the sensor-time points which show significant differences. Zero denotes the onset of the first mask.
ITC analysis
For conscious identification (Figure 4A), the ITC values were significantly higher in the FD condition than the FI condition. The deactivation was observed in the beta range (16-24 Hz) over the left middle posterior sensors during the period of 400 to 520 ms (cluster-based permutation test, p = 0.048). For conscious detection (Figure 4B), there was increased ITC values in the FD compared with the FN condition over the left posterior sites (p = 0.027, 360 to 620 ms, 16 to 32 Hz).
FI - FN FD - FN
A B
C
Figure 4. The scalp topographies of the ITC differences between the FI and FN conditions (A)
and the FD and FN conditions (B). The color scales in the scalp topographies are expressed in t statistic values. The back dots in the scalp topographies highlight the clusters of sensors showing significant differences.
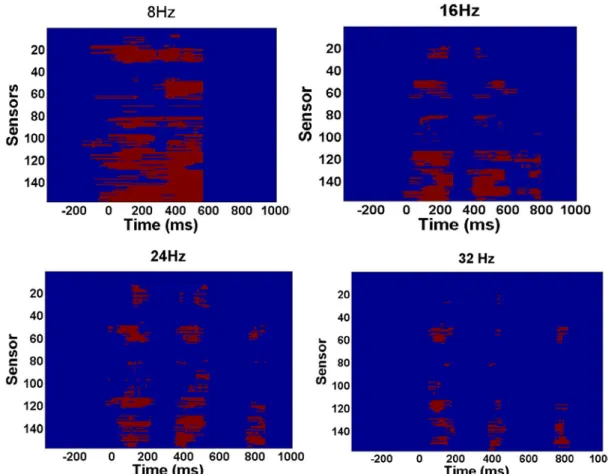
By contrasting the ITC values between the FI and FD conditions, a sustained ITC difference was observed over most of the sensors between 8-50 Hz during the -160 to 820 ms period (p < 0.001). A closer examination of this finding on a
frequency-by-frequency basis, a rhythmic modulation of ITC differences was found. Specifically, we examined the significant ITC differences between the FI and FD conditions at each frequency point along the sensor and time dimensions. Figure 5 shows the significant ITC differences in the sensor-time domain at four representative frequency points.
Figure 5. The time-frequency represenatations of the ITC differences between the FI and FD
conditions at 8Hz (A), 16 Hz (B), 24Hz (C) and 32 Hz (D). Red dots in the figures indicate the sensor-time points which show significant ITC differences as identified by the
cluster-based permutation tests. Zero denotes the onset of the first mask.
Discussion
Taken together, both the results of the planar gradients and ITCs showed distinct spatial-temporal neural correlates for conscious identification and conscious detection.
This suggests that conscious identification and detection rely on different processes and the access to consciousness varies according to the representational levels of the stimuli. Moreover, additional rhythmic activity is required to support the transition from conscious detection to conscious identification.
REFERENCES
Bonnefond, M., & Jensen, O. (2012). Alpha oscillations serve to protect working memory maintenance against anticipated distracters. Curr Biol, 22(20), 1969-1974.
Campana, F., & Tallon-Baudry, C. (2013). Anchoring visual subjective experience in a neural model: The coarse vividness hypothesis. Neuropsychologia, 51(6), 1050-1060.
Clifford, C. W. G., Arabzadeh, E., & Harris, J. A. (2008). Getting technical about awareness. Trends in Cognitive Sciences, 12(2), 54-58.
Craik, F. I. M., & Lockhart, R. S. (1972). Levels of Processing - Framework for Memory Research. Journal of Verbal Learning and Verbal Behavior, 11(6), 671-684.
Dehaene, S., Naccache, L., Cohen, L., Le Bihan, D., Mangin, J. F., Poline, J. B., et al. (2001). Cerebral mechanisms of word masking and unconscious repetition priming. Nature Neuroscience, 4(7), 752-758.
Grill-Spector, K., Knouf, N., & Kanwisher, N. (2004). The fusiform face area subserves face perception, not generic within-category identification. Nature Neuroscience, 7(5), 555-562.
Kouider, S., de Gardelle, V., Sackur, J., & Dupoux, E. (2010). How rich is consciousness? The partial awareness hypothesis. Trends in Cognitive Sciences, 14(7), 301-307.
Liu, J., Harris, A., & Kanwisher, N. (2002). Stages of processing in face perception: an MEG study. Nature Neuroscience, 5(9), 910-916.
Melloni, L., Molina, C., Pena, M., Torres, D., Singer, W., & Rodriguez, E. (2007). Synchronization of neural activity across cortical areas correlates with conscious perception. Journal of Neuroscience, 27(11), 2858-2865.
Seth, A. K., Dienes, Z., Cleeremans, A., Overgaard, M., & Pessoa, L. (2008). Measuring consciousness: relating behavioural and neurophysiological approaches. Trends Cogn Sci, 12(8), 314-321.
Wiens, S. (2006). Subliminal emotion perception in brain imaging: findings, issues, and recommendations. Prog Brain Res, 156, 105-121.
1
國科會補助專題研究計畫項下出席國際學術會議心得報告
日期:103年 10 月 30 日
一、參加會議經過
The Society for Cognitive Neuroscience Annual Meeting is the primary international organization dedicated to cognitive neuroscience research. This year the conference covers a wide range of topics, from emotion, decision-making and neuroscience and law. I presented our recent data in the poster session on April 14.
二、與會心得
It appears that most of research in this conference employed fMRI as a primary tool. Therefore, my study using MEG methodology was not well received. Nevertheless, I still benefit a lot from attending the keynotes speech, symposiums, and face-to-face interactions with other researches during the poster sessions. The most impressive speech to me is the one given by Joseph Ledoux. He argued that most of emotion research in the past focused on how the brain responds to the threats, which is the mechanism crucial to our survival. However, the capacity for the processing of threats is not the same as the capacity to consciously experience of fear when well-being is threatened. In other words, hard-wired survival circuits are often mistakenly described as emotion circuits. I totally agree with his viewpoint. However
計畫編號
NSC 101-2410-H-004-088
計畫名稱
工作記憶在知覺意識所扮演的角色
出國人員
姓名
徐慎謀
服務機構
及職稱
政治大學心智大腦與學習中心
助理研究員
會議時間
102 年 4 月 13 日
至
102 年 4 月 16 日
會議地點
美國舊金山
會議名稱
(中文)認知神經科學會議
(英文)Cognitive Neuroscience Society 2013 Annual Meeting
發表論文
題目
(中文)藉由阿爾法波瞭解連續決策的神經機制
(英文)The neural mechanisms of sequential decision-making as revealed
by alpha-band oscillations
2
the challenge lying ahead is how to measure consciously emotional experience. This new issue may open a new window for my future research.
三、考察參觀活動(無是項活動者略)
N/A四、建議
N/A五、攜回資料名稱及內容
The PDF version of the program and abstract book
六、其他
N/A科技部補助計畫衍生研發成果推廣資料表
日期:2014/10/30科技部補助計畫
計畫名稱: 工作記憶在知覺意識所扮演的角色 計畫主持人: 徐慎謀 計畫編號: 101-2410-H-004-088- 學門領域: 實驗及認知心理學無研發成果推廣資料
101 年度專題研究計畫研究成果彙整表
計畫主持人:徐慎謀 計畫編號: 101-2410-H-004-088-計畫名稱:工作記憶在知覺意識所扮演的角色 量化 成果項目 實際已達成 數(被接受 或已發表) 預期總達成 數(含實際已 達成數) 本計畫實 際貢獻百 分比 單位 備 註 ( 質 化 說 明:如 數 個 計 畫 共 同 成 果、成 果 列 為 該 期 刊 之 封 面 故 事 ... 等) 期刊論文 0 0 100% 研究報告/技術報告 0 0 100% 研討會論文 0 0 100% 篇 論文著作 專書 0 0 100% 申請中件數 0 0 100% 專利 已獲得件數 0 0 100% 件 件數 0 0 100% 件 技術移轉 權利金 0 0 100% 千元 碩士生 0 0 100% 博士生 0 0 100% 博士後研究員 0 0 100% 國內 參與計畫人力 (本國籍) 專任助理 0 0 100% 人次 期刊論文 0 1 100% 論文撰寫中 研究報告/技術報告 0 0 100% 研討會論文 1 1 100% 篇 部 分 結 果 已 發 表 於 2013 認知神經 科學會議 論文著作 專書 0 0 100% 章/本 申請中件數 0 0 100% 專利 已獲得件數 0 0 100% 件 件數 0 0 100% 件 技術移轉 權利金 0 0 100% 千元 碩士生 0 0 100% 博士生 0 0 100% 博士後研究員 0 0 100% 國外 參與計畫人力 (外國籍) 專任助理 0 0 100% 人次其他成果