國
立
交
通
大
學
理學院 應用科技 學程
碩
士
論
文
探討
Sortase A 突變株 D165A、K196T 和
P 9 4 S / D 1 6 0 N / D 1 6 5 A / K 1 9 6 T 的 反 應 性
Enzymatic activity of Sortase A mutants-D65A, K196T and
P94S/D160N/D165A/K196T
研 究 生:李達人
指導教授:李耀坤 教授
摘要
轉肽酶 Sortase A 藉由特殊的胺基酸序列-N 端的 LPETG 序列與 C 端的
GGGGG 序列,可將兩端不同功能的蛋白質或胜肽段連接起來,同時保持正確的
構形,形成多功能嵌合蛋白。本研究藉由兩種不同的螢光受質 Abz-LPETG-Dnp
與GGGGGK-FITC,來描述 Sortase A 野生株與突變株的水解及接合之功能。發
現Sortase A 的 P94S、D160N、D165A、K196T 四點突變株比 Sortase A 的野生株
在水解初速率上大3.14 倍,在接合初速率上大 1.18 倍。同時應用 Sortase A 的作
用,將GGGGGK-FITC 的胜肽段接合在具有 LPETG 的 QCM 晶片載體上。未來
可擴大此應用在具有LPETG 的晶片載體上,接合不同的抗體作探針進行免疫檢
查。
Abstract
Sortase A is kind of transpeptidase which can connect two different function proteins or peptides together to form the multifunction chimeric protein without interrupt the structure and function by using the special amino acid sequences: the N terminal LPETG and the C terminal GGGGG. In this study, we compare the function of hydrolysis and connection between the wildtype and mutant line by using two different fluorescence substrates the Abz-LPETG-Dnp and GGGGGK-FITC. We discover the P94S, D160A, D165A, K196T 4 point mutant which is 3.14 times faster in hydrolysis and 1.18 times faster in connection initial rate than the wildtype sortase A. By sortase A, we can link the protein or peptide with GGGGGK-FITC sequence to the QCM chip with LPETG sequence; using this method, we can link the antibodies to a chip to do the immunoassays.
誌謝辭
從民國九十八年七月至一百零二年二月,從桃園龍潭至新竹市交通大學來 回約百公里每週三到五趟的路程終於在今日可以立下一個里程碑。 民國九十八年在家人的鼓勵下考取了交通大學理學院碩士專班應用科技 組,雖然大學是唸物理系,目前從事教育工作,但因對酵素科技感到興趣便想進 修相關的知識。感謝李耀坤教授能給我這個機會進入酵素科技實驗室,從對分子 生物實驗操作的一無所知到熟悉相關實驗操作,對我來說實是巨大的挑戰。所幸 在三年半多的夜晚與假日,實驗室清冷的燈光下,總有學長姐、同學、學弟妹給 予極大的幫忙與鼓勵,如岳進、惜禾、家瑜、媛婷、裕國、晟富、模沅、彰璟、 安芃、丞偉、曉眉、信宏、嘉郡、智仁等,尤其是假日見到家瑜、模沅、彰璟、 晟富、丞偉出現在實驗室的身影總令人感到心安,對藥品的擺放位置、儀器的操 作、實驗的內容問題總可得到滿意的答案或解決的辦法。 感謝爸爸、媽媽、明彰表哥、惠美與小采霓的支持與鼓勵,一路走來形單 心不單,在李耀坤老師的指導下完成本篇論文。在此 致上萬分感謝。目錄
摘要 ... I Abstract... I 誌謝辭 ... II 目錄 ...III 圖目錄 ...V 表目錄 ...VI 第一章 緒論 ...1 1-1 蛋白質接合酵素簡介 ...1 1-2 Sortase A 的功能 ...2 1-3 Sortase A 活性區域的結構 ...4 1-4 Sortase A 的活性部位與作用機制 ...5 1-4-1 Sortase A 辨認受質的活性部位 ...5 1-4-2 鈣離子對Sortase A 的影響 ...7 1-4-3 Sortase A 與受質反應的作用機制 ...8 1-4-4 溫度對Sortase A 的影響 ...9 1-5 Sortase A 之活性檢定 ...11 1-6 研究目的 ...12 第二章 實驗材料與方法 ...14 2-1 實驗藥品與儀器 ...14 2-2 Sortase A 質體的取得與定點突變 ...16 2-2-1 野生型表現系統的建立 ...16 2-2-2 定點突變 ...18 2-2-3 酵素的誘導 ...20 2-2-4 酵素的純化 ...21 2-2-5 決定蛋白質分子量與純度 ...22 2-2-6 蛋白質身份的鑑定 ...23 2-2-7 酵素的保存 ...24 2-2-8 蛋白質濃度的測定 ...24 2-3 Sortase A 反應機制之研究 ...25 2-3-1 水解活性的檢測與比較 ...25 2-3-2 接合活性的檢測與比較 ...26 第三章 結果與討論 ...28 3-1 酵素的基因序列、表現與純化 ...28 3-1-1 酵素的基因序列 ...28 3-1-2 酵素的表現與純化 ...293-1-3 Sortase 野生株與突變株的分子量 ...32 3-1-4 野生型酵素濃度測定 ...32 3-2 野生型與突變型酵素的活性檢測 ...34 3-2-1 Sortase A 野生株的活性檢測 ...34 3-2-2 水解活性與比較 ...36 3-2-3 接合活性的檢測 ...37 3-2-4 接合活性的比較 ...40 3-2-5 野生株與突變株的活性比較討論 ...43 3-3 Sortase A 四點突變株之應用 ...44 第四章 結論 ...46 第五章 未來研究方向 ...47 References...49 附錄一:DNA 序列與胺基酸序列對照表 ...55 附錄二:MALDI-TOF 分析的分子量圖譜 ...58 附錄三:Abz-LPETG-Dnp 被 Sortase A 水解的螢光量值對時間紀錄表 ...59 附錄四:不同時間點的MRM 的面積圖譜 ...64
圖目錄
圖 1-1:Sortases 種類示意圖 ...2
圖 1-2:Sortase A 功能示意圖 ...3
圖 1-3:Sortase A△N59 的結構 ...4
圖 1-4 :Sortase AΔN59 的電性分佈圖 ...5
圖 1-5:Srt A△N59 活性部位結合 LPETG 的 X-ray 晶體結構圖 ...6
圖 1-6:鈣離子對 Sortase A 作用示意圖 ...7
圖 1-7:鈣離子結合對 β6/β7 區域的影響示意圖 ...8
圖 1-8:“LPXT-酵素”中間物的形成示意圖 ...8
圖 1-9:Sortase A 的水解與接合機制示意圖 ...9
圖 1-10:Circular Dichroism (CD) Spectroscopy ...10
圖 1-11:Abz-LPETG-Dnp 之結構式 ...11 圖 1-12:GGGGGK-FITC 結構圖 ...12 圖 1-13:Sortase A 定點突變位置圖 ...13 圖 2-1:Sortase A 野生型表現系統質體的建立 ...16 圖 2-2:Sortase A 野生型表現系統質體的驗證 ...17 圖 3-1:Sortase A 野生型及突變株胺基酸定序表 ...29 圖 3-2:Sortase A 野生株及突變株的 SDS-page 蛋白質分子量 ...30 圖 3-3:Sortase A 野生株的 MALDI-TOF 分析圖譜 ...31
圖 3-4:Sortase A 野生株的 Mascot Score...31
圖 3-5:蛋白質標準品對 OD595 之標準檢量線 ...33
圖 3-6:Abz_LPETG_Dnp 被 Sortase A 分解的 LC/MS 圖譜 ...35
圖 3-7:Sortase A 野生株與突變株水解 Abz-LPETG-Dnp 的螢光值對時 間關係圖...36
圖 3-8:Sortase A 野生株與突變株水解活性的反應初速率 ...37
圖 3-9:Sortase A 野生型對 Abz_LPETG_Dnp 與 GGGGGKFITC 的反應 質譜圖...38 圖 3-10:生成物荷質比 691 訊號的 MSMS 分析 ...39 圖 3-11:Abz-LPETGGGGGK-FITC 的 MRM 分析 ...41 圖 3-12:時間對 MRM 面積的關係圖 ...42 圖 3-13: Sortase A 野生株與突變株接合活性的反應初速率 ...42 圖 3-14:SrtPro 在 QCM 上的應用示意圖 ...45 圖 3-15:接合 GGGGGK-FITC 的 QCM 晶片圖 ...45 圖 5-1:Sortase A 的 FRET 研究示意圖 ...47
表目錄
表 1-1:Sortase A 及其突變株的活性檢測 ...10 表 1-2:Sortase A 野生株與突變株結構改變的臨界溫度 ...11 表 2-1:單一突變點寡核苷酸模版引子 ...18 表 2-2:PCR 反應條件 ...19 表 2-3:四個突變點寡核苷酸模版引子 ...20 表 2-4:SDS 膠片配置表 ...22表 2-5:Sample loading buffer 配置表 ...23
表 2-6:LC-MASS 的分析條件表 ...27 表 3-1、不同濃度 BSA 標準品與 OD595 之吸收值 ...32 表 3-2:Sortase A 野生株與突變株的濃度 ...34 表 3-3:Abz_LPETGGGGGKFITC 訊號的 MSMS 分析表 ...40 表 3-4:每隔 5 分鐘的 Abz-LPETGGGGGK-FITC 之 MRM 面積積分值 ...41 表 3-5:野生株與突變株的活性比較表 ...43 表 3-6:Sortase A 水解反應研究結果之相對誤差 ...44
第一章 緒論
1-1 蛋白質接合酵素簡介
蛋白質接合酵素可將不同的胜肽段或不同功能的蛋白質與以接合後,形成新的多功能 性蛋白質(或稱嵌合蛋白),可應用在蛋白質的研究、生化檢測或醫療上藥物傳送系統的研 究等。 以化學操作方式將不同蛋白質接合的多種方法被發表出來,如Dawson 在 2000 年提到 以化學合成方法連接不同的胜肽或蛋白質(1)、根據不同策略利用內含子做蛋白質的接合 (2)、以半胱氨酸或其它胺基酸的特殊殘基之親合性來接合(3)。但這些接合方式需要高度的 操作技術,而且容易被非相關的胺基酸殘基或蛋白質阻擋接合位置,而影響蛋白質的接合。 在生物化學操作上最常表現嵌合蛋白的方法,是設計融合的基因序列,再表現出嵌合 蛋白。但此方法常因融合基因中的組成基因序列各有其適合的表現系統與條件,故易遇到 嵌合蛋白質不易表現、大量表現但表現在涵包體、或表現在溶解態卻因折疊不正確而無活 性的狀況出現(4)。 為避免融合基因的表現限制,在其組成基因中各加入特殊蛋白質序列的基因,讓組成 基因能在原本的表現系統與條件中表現且保持原正確的構形,再透過蛋白質間特殊的序列 (N 端為 LPETG,C 端為 GGGGG),讓轉譯後的蛋白質經酵素Sortases 作用而互相連接以 形成新的嵌合蛋白質(5, 6)。 Sortases 是在革蘭氏陽性菌中發現的半胱氨酸轉移胜肽酶,依其辨認序列的同源性及功能性可分為四種類型,分別為Sortase A、SortaseB 、SortaseC 、SortaseD,如下圖 1-1(7,
8)。Sortase A 透過 LPXTG(Leucine-Proline-X-Threonine-Glycine; X 為任意胺基酸)的特殊 序列,能催化表面蛋白與細菌細胞壁的共價連結,此反應與細菌粘附、免疫反應、細菌感 染,或噬菌體結合的受體功能有關(9)。Sortase B 透過 NPQTN 的特殊序列,讓細菌能獲得
鐵離子(10)。Sortase C 透過 LPXTG 的特殊序列催化細菌表面鞭毛的形成(11)。Sortase D 透 過LPNTA 的特殊序列與孢子的形成有關(9, 12)。 圖 1-1:Sortases 種類示意圖 (9) Sortases 藉著辨認特殊序列,催化不同胜肽或蛋白質連結的特性,為多功能崁合蛋白 的合成提供一個有別於化學合成或生物化學上表現融合基因的方法。其中Sortase A 被廣泛 的研究,應用在不同功能蛋白的連接上。
1-2 Sortase A 的功能
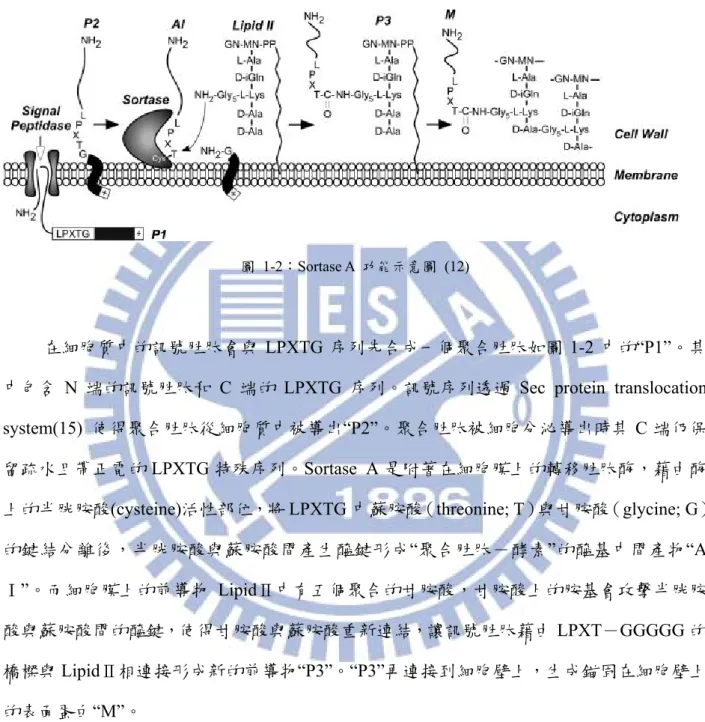
在革蘭氏陽性菌上Sortase 扮演重要的角色,若缺少了 sortases 的作用,則細菌將喪失 感染的能力,如金黃色葡萄球菌若無sortase A 的作用,則不會成為動物的感染源(13)。 在革蘭氏陽性菌中,Sortase A 其 N 端在細胞膜內,其 C 端在細胞壁中,能辨認細菌 表面蛋白或訊號胜肽中LPXTG 的特殊序列(X 為任意胺基酸),讓表面蛋白與與細菌的細 胞壁肽聚糖的GGGGG 序列以共價鍵接合(12, 14)。以圖1-2 Sortase A 功能示意圖來介紹攜帶 LPXTG 的胜肽段如何被 Sortase A 連結在細 胞壁上。
圖 1-2:Sortase A 功能示意圖 (12)
在細胞質中的訊號胜肽會與 LPXTG 序列先合成一個聚合胜肽如圖 1-2 中的“P1”。其
中包含 N 端的訊號胜肽和 C 端的 LPXTG 序列。訊號序列透過 Sec protein translocation
system(15) 使得聚合胜肽從細胞質中被導出“P2”。聚合胜肽被細胞分泌導出時其 C 端仍保
留疏水且帶正電的LPXTG 特殊序列。Sortase A 是附著在細胞膜上的轉移胜肽酶,藉由酶
上的半胱胺酸(cysteine)活性部位,將 LPXTG 中蘇胺酸(threonine; T)與甘胺酸(glycine; G) 的鍵結分離後,半胱胺酸與蘇胺酸間產生醯鍵形成“聚合胜肽-酵素”的醯基中間產物“A Ⅰ”。而細胞膜上的前導物 LipidⅡ中有五個聚合的甘胺酸,甘胺酸上的胺基會攻擊半胱胺 酸與蘇胺酸間的醯鍵,使得甘胺酸與蘇胺酸重新連結,讓訊號胜肽藉由LPXT-GGGGG 的 橋樑與LipidⅡ相連接形成新的前導物“P3”。“P3”再連接到細胞壁上,生成錨固在細胞壁上 的表面蛋白“M”。 利用 Sortase 的特性,在不同功能的胜肽或蛋白質中,以基因工程分別在 N 端加入 LPXTG 序列與 C 端加入 GGGGG 序列,就可將兩種不同的胜肽或蛋白質連接,形成新的 胜肽或嵌合蛋白(14, 16, 17)。其接合受質非常廣泛如螢光蛋白(18, 19)、胜肽核甘酸(20)、糖 蛋白連結(21)、聚合物(22)、固態物質(23, 24)、脂質(25, 26)、微小粒子(27)等。 其應用方法,如利用 Sortase A 接合不同的抗體在特定載體上作探針進行免疫檢查
(28)、製造傳遞蛋白(29)或標示特定領域以利核磁共振的觀察(30)等,顯示 Sortase A 的催化 加成反應有廣泛的應用性(31, 32),促使吾人對利用 Sortase A 生產嵌合蛋白的優化條件有更 深入的探討與研究。
1-3 Sortase A 活性區域的結構
Sortase A 有 206 個胺基酸,其 N 端在細胞質中可將 Sortase A 錨固在細胞膜,而 C 端 的部份在細胞壁中是催化反應的區域(13, 14)。 將缺少 N 端的 24 個胺基酸(Sortase A△N24)或 59 個胺基酸的基因重組 Sortase A ( Sortase A△N59)被證實能在大腸桿菌之水溶狀態細胞質液中大量表現(16, 33, 34),且讓 具有LPXTG 序列(X 可以是任意胺基酸)與 GGGGG 的胜肽段相互連結(35)。 圖 1-3:Sortase A△N59 的結構 (35)Sortase A△N59結構如圖1-3,其核心主要有 8 段 β 折板,一個 α 螺旋與 2 個連結 β 折 板的3 折螺旋(β3/β4 與 β6/β7)。其中 β4、β7、β8 形成一個疏水性的孔洞,而連結 β6、β7 的螺旋線段具有彈性,使得sortase 能像捕手手套一樣能在活性條件下改變結構捉住受質。
-4 Sortase A 的活性部位與作用機制
(Leucine-Proline-Glutamate -Threonine-Glycine) 相結合時其電性分佈圖如下圖1-4(35)。1
1-4-1
Sortase A 辨認受質的活性部位
Sortase A△N59與胺基酸序列LPETG 圖 1-4 :Sortase AΔN59 的電性分佈圖 (35) 圖中深綠色部份為疏水區、紫色為極性的殘基,青色為正電區域,紅色是酸性殘基。因LPETG 序列具疏水性且帶正電,故會與 Sortase A△N59結構中的β4、β7、β8 形成的疏 水性
ortase A△N59相親合後,各活性部位殘基與LPETG 作用的 X-ray 晶體結
構如下圖1-5 (16)。 孔洞相親合。 當LPETG 與 S 圖 1-5:Srt A N59△ 活性部位結合LPETG 的 X-ray 晶體結構圖 (16) 和 Gln-172 則無被發現與催化作用有關(16)。 7/β8 (綠)區域與 GGG 的辨認有關(35, 38)。 在圖1-5 中,當 LPETG(黃色)被束縛在活性部位時,各 β 折板上活性部位與 LPETG 有如下作用:β3/β4 (藍)區域與鈣離子的結合有關(36)。β6/β7(紅)區域與辨認 LPXTG 中
的LP 殘基有關(35-37),其中 Val-168 和 Leu-169 被認為是重要的識別殘基、Glu-171 為鈣
離子結合後的作用有關,而Gly-167、 Asp-170
1-4-2
鈣離子對 Sortase A 的影響
當β3/β4 區域上的殘基 Glu-105, Glu-108, Asp-112, and Asn-114 與鈣離子結合時,會透
過鈣離子拉住β6/β7 區域上的 Glu-171,如圖 1-6,讓原本具有彈性的 β6/β7 區域改變成有
次序的構形(36)。
圖 1-6:鈣離子對 Sortase A 作用示意圖 (36)
而β6/β7 區域改變成有次序的構形時,有助於辨認 LPXTG 序列,如圖 1-7。當 LPXTG
與Sortase A 結合後,β7 上的 Cys184 能裂解 LPXTG 序列中的蘇胺酸(threonine; T)與甘
圖 1-7:鈣離子結合對 β6/β7 區域的影響示意圖 (36)
1-4-3
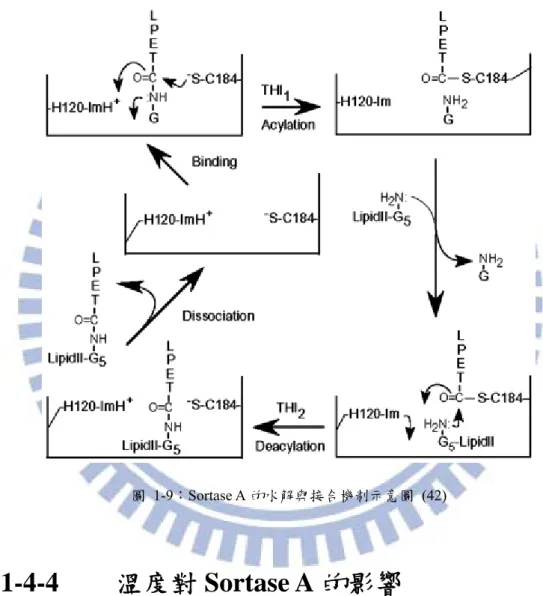
Sortase A 與受質反應的作用機制
當LPXTG 與 Sortase 疏水性孔洞相親合時,亮胺酸 L 與脯胺酸 P 被 β6/β7 區域辨認, 且分佈在Sortase A 疏水孔洞中的三個重要活性部位:分別為在 β7 上的 C184、在 β8 上的 R197 與在 β4 上的 H120,開始對蘇胺酸 T 與甘胺酸 G 作用(39) ,其作用示意圖如圖 1-8。 半胱胺酸Cys184 的氫硫鍵會對 LPXTG 序列中蘇胺酸 Threonine 上碳鏈上的羰基產生親核 攻擊,在酵素與受質間形成硫脂鍵(thioester),羰基與氧的共價鍵電子被擠壓至氧上,Arg197 的氨基因帶正電,提供庫倫吸引力吸住電子電,但因氧原子的陰電性大於氨基,故電子再 回振到羰基上形成碳氧原子間的共價鍵,此時為滿足八隅體的穩定,碳原子與氮原子間的 鍵結被切斷,電子轉移到 His120 殘基,His120 釋出氫離子,進而產生“LPXT-酵素”的中 間物(40, 41)。 圖 1-8:“LPXT-酵素”中間物的形成示意圖 (40)當“LPXT-酵素”中間物遇到五甘胺酸(pentaglycine)時,甘胺酸上的氨基對 “LPET -酵素”中間物產生親核性攻擊,使“LPXT-酵素”中間物的蘇胺酸 T 與五甘胺酸的 G 產生 醯胺鍵(amide bond)的連結,形成 LPXT-GGGGG 的生成物。而 Sortase A 再度被釋放於 溶液中,進行下一個循環的反應(42),如圖 1-9。
圖 1-9:Sortase A 的水解與接合機制示意圖 (42)
1-4-4
溫度對 Sortase A 的影響
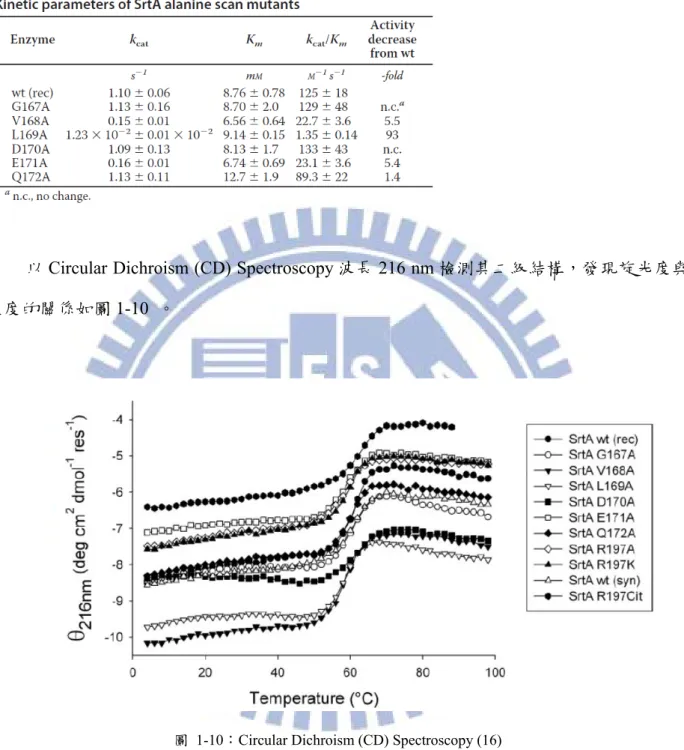
根據 Matthew L. Bentley 等人的研究(16),Sortase A 及其突變株在 37℃下在水解
表 1-1:Sortase A 及其突變株的活性檢測 (16)
以Circular Dichroism (CD) Spectroscopy 波長 216 nm 檢測其二級結構,發現旋光度與
溫度的關係如圖1-10 。
圖 1-10:Circular Dichroism (CD) Spectroscopy (16)
其結構改變的臨界溫度最低在56 ℃以上,如表 1-2。由此推測 Sortase A 及其突變株
表 1-2:Sortase A 野生株與突變株結構改變的臨界溫度 (16)
1-5 Sortase A 之活性檢定
Sortase A 同時具有水解反應與接合反應的特性,根據研究(44)指出,Sortase A 的反應 依循“乒乓機制” (ping-pong mechanism) (45)。即 Sortase A 與 LPETG 先形成醯基脢 (acyl-enzyme)中間物後,GGGGGC 才會產生親核攻擊,最後形成 LPET-GGGGGC 的產物。
故 其 活 性 檢 定 分 兩 階 段 , 首 先 利 用 自 消 螢 光 胜 肽 o-aminobenzoyl-
Leucine-Proline-Glutamate-Threonine- Glycine-2,4-dinitrophenyl(以下簡稱 Abz-LPETG-Dnp)
來檢測Sortase A 的水解活性並用 Liquid Chromatograph/Mass Spectrometer (LC/MS)驗證其
反應物與生成物分子量的訊號。
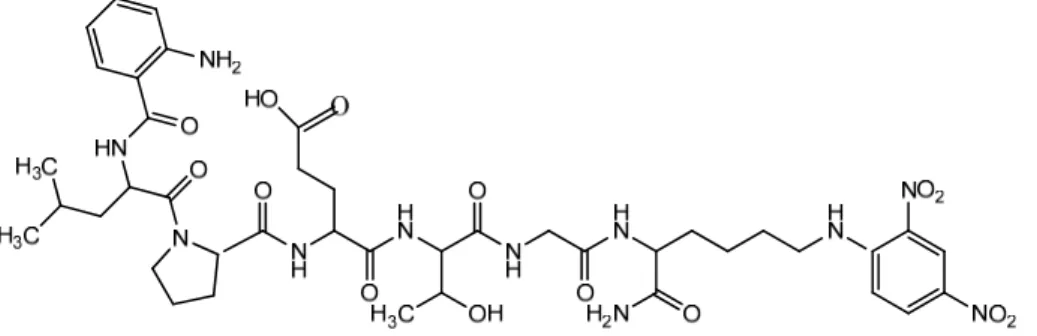
再 取 胜 肽 段 Glycine-Glycine- Glycine- Glycine- Glycine-Lysine- Fluorescein isothiocyanate (以下簡稱 GGGGGK-FITC)與 Abz-LPETG-Dnp 反應,Sortase A 會將 G-Dnp
從 Abz-LPETG-Dnp 切開,再將 Abz-LPET 轉移接合到 GGGGGK-FITC 上,最後形成
Abz-LPETGGGGGK-FITC。以 LC/MS 檢測生成物 Abz-LPETGGGGGK-FITC 的分子量訊 號,並分析其訊號量值來了解Sortase A 的接合反應。 圖 1-12:GGGGGK-FITC 結構圖
1-6 研究目的
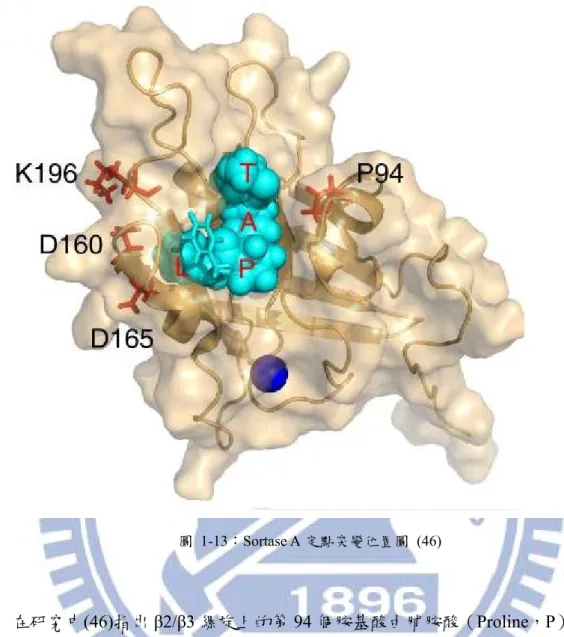
有研究指出在Sortase A 的三個螺旋上有四個胺基酸的突變點會影響其接合效率(46)。 這四個突變點分別為在β2/β3 縲旋上的 P94,在 β6/β7 的連接縲旋上的 D160 和 D165,與在 C 端的 β7/β8 連接縲旋上的 K196。這些位置都在 Sortase A 的表面,且靠近 LPXTG 結合位 的疏水孔洞,如圖1-13。圖 1-13:Sortase A 定點突變位置圖 (46)
在研究中(46)指出 β2/β3 縲旋上的第 94 個胺基酸由脯胺酸(Proline,P)突變成絲胺
酸(Serine,S),簡稱P94S;在 β6/β7 縲旋上的第 160 個胺基酸由天冬胺酸(Aspartic acid,
D)突變成天冬醯胺(Asparagine,N),簡稱 D160N;第 165 個胺基酸由天冬胺酸突變成丙 胺酸(Alanine),簡稱 D165A;在 β7/β8 縲旋上的第 196 個胺基酸由賴胺酸(Lysine,K) 突變成蘇胺酸(Threonine,T),簡稱 K196T,被證實能增進 Sortase A 的效率(46),應是此 四個突變造成縲旋圈的構型改變,使 sortase 能更容易與受質結合。其中 P94S、D160N、 D165A 與在 LPXT 上的結合有關;K196T 與 GGGGG-胜肽的接合有關。 本研究期望在Sortase A 的野生株(簡稱 SrtW)上進行四個點突變 P94S、D160N、D165A 與 K196T。取得四點突變株(簡稱 SrtPro)後,進行突變酵素的催化反應研究,期待在室 溫的環境中找出能提高蛋白質接合效率的突變型酵素,以應用在嵌合蛋白的接合中。
第二章 實驗材料與方法
2-1 實驗藥品與儀器
實驗所需之藥品來源與儀器規格敘述如下: 一. 藥品: 1. 配製菌種培養液之藥品購自 Merck、Difco、Zymeset 公司 2. 菌種轉型用之藥品均購自 Invitrogen、Sigma、Merk 公司 3. 配製緩衝液之藥品均購自 Sigma、聯工公司 4. 各種 Kit 購自 GeneMark、Viogene 公司 5. 限制酶購自 NEB 公司 6. 轉型之菌種購自益生公司 7. 胜肽 Abz-LPETG-Dnp:購自 anaSpec 公司 8. 胜肽 GGGGC-FITC:由明欣生物科技公司合成 二. 儀器 恆溫培養箱(EYELA NDO-450ND)搖動培養箱(Firstek, Scientific, orbital shaking incubator Model-S302R) UV 吸收光譜儀(AGILEMT 8453)
螢光光譜儀(Hitachi f 4500)
Elisa reader(Themo,Multiskan GO Microplate Spectrophotometer) 聚合酶連鎖反應器(GeneAmp PCR system 2400, 9700)
桌上型離心機(Hermle) 高速離心機(Kubata 7820)
超音波震盪器(Misonix, ultrasonic processor)
高效能液相層析儀(High Performance Liquid Chromatography) HPLC column:Inertsil 7 ODS-3(5μm,4.6 mm×250 mm)
三. 層析管柱:
HPLC system(Pharmacia Biotech FPLC3467) HiTrap Desalting column(Pharmacia ,5mL) HiTrap Ni coumn(Pharmacia)
四. 寡核甘酸引子與定序工作:委託明欣生物科技公司。
2-2 Sortase A 質體的取得與定點突變
2-2-1
野生型表現系統的建立
一、質體的重組
將Sortase A 野生型的基因片段,以限制酶 AseⅠ與限制酶 HindⅢ將包含 Sortase A 活
性部份(第 25 個胺基酸 H 到第 206 個胺基酸 K)的 DNA 片段共 546 鹼基對 (base pair,bp) 切出來(14, 16, 17)。再以限制酶 NdeⅠ與限制酶 HindⅢ將 pet28a 的表現載體 5318 bp 切出
來,因為AseⅠ與 NdeⅠ的切位有 TA 兩個 DNA 序列碼重疊,故上述的兩個線性基因片段
可用T4DNA 接合酶接合共 5864 bp,在 22 ℃下進行聚合脢鏈反應(polymerase chain reaction,
PCR)反應兩小時,得到 Sortase A 野生型表現系統的質體。
+
HindⅢ AseⅠ HindⅢ NdeⅠ 圖 2-1:Sortase A 野生型表現系統質體的建立因AseⅠ與 NdeⅠ接合後切位點消失,故以含 Sortase A 活性部份的 DNA 片段的兩切
位點NcoⅠ與 HindⅢ,來作反應,以 DNA 電脉膠片來檢驗質體的大小應符合下列理論值:
pet28a 的 DNA 大小為 5246 bp ,含 His-tag 標櫼的 Sortase A 野生株 DNA 大小為 618 bp,如
b.p 750 500 1000 5000 3000 6000 圖 2-2:Sortase A 野生型表現系統質體的驗證 二、勝任細胞的製備:大腸桿菌菌種DH5α 與 BL21(DE3) 取大腸桿菌菌種加入2 ml 之 LB 中,置入 37℃培養箱 12 hr。稀釋 50 倍至 100 ml,再 置入37 ℃培養箱中養菌,直到 OD 值在 0.4~0.6 間。以 1500 rpm、4 ℃離心 6 min,去上 清液。以0 ℃、50 ml、100 mM 的 CaCl2回溶(原0.5 倍體積),每隔 5 min 以手輕拍之且
置於冰上30 min。再以 1500 rpm、4 ℃離心 6 min,去上清液。加入 10 ml 含 30%的 glycerol
的CaCl2(7 ml、100 mM CaCl2與3 ml 的甘油)回溶(原 0.1 倍體積)。最後取 100 μl 分
裝至eppendorf。存放至-80 ℃備用。
三、轉型
將已建立完成的表現質體以熱休克反應(heat shock response)轉型至大腸桿菌 DH5α
的菌株中,操作如下:取勝任細胞DH5α 放在冰上溶解後,加入表現質體 5~10 μl,在冰上
養液,在37 ℃培養箱養菌 30 分鐘後,在無菌操作臺上塗盤(含 kanamycin 50 μg/ml)。將
塗盤後的培養皿置入 37 ℃培養箱中培養 16 小時。挑單株菌於 5 ml 的液養液中(含
kanamycin 50 μg/ml)置入 37℃培養箱中培養 16 小時,取菌液 4.5 ml 抽出 Sortase A 野生
型質體,將DNA 質體於-20 ℃中保存。且將菌液 200 μl 加入 200 μl 培養液與濃度 50%的
400 μl 甘油(甘油最後濃度 25%),放入-80 ℃中保存。
取5 μl 質體以 NcoⅠ及 HindⅢ限制酶水解之,並進行 DNA 電泳分析,檢驗 DNA 質體
的大小,並做DNA 定序以確定基因片段與載體已成功接合。
2-2-2
定點突變
一、單一定點突變 取得Sortase A 野生株的質體後,設計在胺基酸位置第 165 與第 196 的突變點寡核苷酸 模版引子如表2-1。 表 2-1:單一突變點寡核苷酸模版引子 Name Sequence( 5’ to 3’)D165A-F GTG AAA CCA ACG GCC GTG GAA GTC CTG
D165A-R CAG GAC TTC CAC GGC CGT TGG TTT CAC
K196T-F TGG GAA ACC CGT AAA ATC TTC
K196T-R TTT ACG GGT TTC CCA CAC ACC
各取1 μl 的引子與 Sortase A 的質體 1 μl(濃度約 5 ng/μg)混合濃度 10 μM
,利用聚合酶連鎖反應(polymerase chain reaction, PCR)以核苷酸模版引子將 Sortase A 的
表 2-2:PCR 反應條件
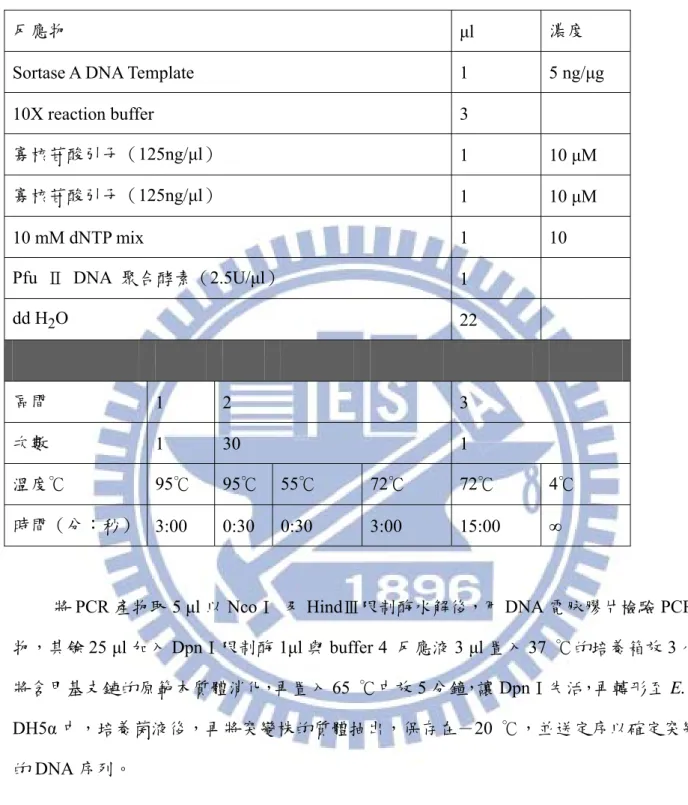
反應物 μl 濃度
Sortase A DNA Template 1 5 ng/μg
10X reaction buffer 3
寡核苷酸引子(125ng/μl) 1 10 μM
寡核苷酸引子(125ng/μl) 1 10 μM
10 mM dNTP mix 1 10
Pfu Ⅱ DNA 聚合酵素(2.5U/μl) 1
dd H2O 22
區間 1 2 3
次數 1 30 1
溫度℃ 95℃ 95℃ 55℃ 72℃ 72℃ 4℃
時間(分:秒) 3:00 0:30 0:30 3:00 15:00 ∞
將PCR 產物取 5 μl 以 NcoⅠ 及 HindⅢ限制酶水解後,用 DNA 電脉膠片檢驗 PCR 產
物,其餘25 μl 加入 DpnⅠ限制酶 1μl 與 buffer 4 反應液 3 μl 置入 37 ℃的培養箱放 3 小時 將含甲基支鏈的原範本質體消化,再置入65 ℃中放 5 分鐘,讓 DpnⅠ失活,再轉形至 E. coli DH5α 中,培養菌液後,再將突變株的質體抽出,保存在-20 ℃,並送定序以確定突變株 的DNA 序列。 二、四個定點突變 為增加至四個突變點在表現質體pet28a 上,本研究先縮短範本質體長度後,做點突變,
最後再將四個點突變的Sortase A 質體接回原表現質體 pet28a 上。故先去除 Sortase A 野生
(如表2-3 所示)將野生型的胺基酸序列從第 60 個胺基酸到第 206 個胺基酸的 DNA 序列
利用PCR 夾出,以 NdeⅠ 及 HindⅢ限制酶水解後,用 DNA 電脉膠片分離 PCR 產物,回
收462 bp 質體片段後,用 T4DNA 接合酶接入有相同切位的 pRSET A 載體(2756 bp),如此
可將原本長5864 bp 的質體,縮短為長 3218 bp 的質體,以增加點突變的成功率。再以表
2-3 中的核苷酸模版引子,依序如單一定點突變的方式取得 Sortase A 四個突變的基因序列。
表 2-3:四個突變點寡核苷酸模版引子
Name Sequence( 5’ to 3’)
primerA AGC CATATG CACCACCACCACCACCACCAAGCCAAA
primerB CGC AAG TTA TTT CAC TTC GGT
P94S-F GGT CCG GCT ACT TCT GAG CAA CTG AAT
P94S-R ATT CAG TTG CTC AGA AGT AGC CGG ACC
D160N-F ACC TCC ATC CGT AAC GTG AAA CCA ACG
D160N-R CGT TGG TTT CAC GTT ACG GAT GGA GGT
D165A-F GTG AAA CCA ACG GCC GTG GAA GTC CTG
D165A-R CAG GAC TTC CAC GGC CGT TGG TTT CAC
K196T-F TGG GAA ACC CGT AAA ATC TTC
K196T-R TTT ACG GGT TTC CCA CAC ACC
將取得的四個突變點Sortase A 的 DNA 序列與 pet28a 以限制酶 NdeⅠ 及 HindⅢ水解
後,用T4DNA 接合酶接合,取得四個突變點的表現質體,簡稱 SrtPro。
Sortase A 野生株與突變株的 DNA 序列與胺基酸序列對照表見附錄一。
2-2-3
酵素的誘導
抗生素kanamycin 的培養基上。置入 37 ℃的培養箱中約 16 小時。分別取單株菌落置入 5 ml
的培養液中(含1 mM/ml 的 kanamycin)中約 12 小時,得到菌液。
分別取Sortase A 野生株與突變株之菌液 1 ml 置入 500ml 的培養液中,待菌液濃度至
OD600 = 0.4 時 加 入 異 丙 基 - β -D- 硫 代 吡 喃 半 乳 糖 苷 ( Isopropyl
β-D-1-Thiogalactopyranoside,IPTG)(最後溶液1 mM/1ml)誘導 12 小時後,以 15000 rpm、
4 ℃離心後去上清液,留下菌體。加入貯存的 Buffer 1(50 mM tris,150 mM NaCl,pH=8.0)10 ml。 以超音波在冰上振破菌液(強度4.5,頻率 50Hz)振盪 3 分鐘,打破細胞壁,再以 15000 rpm、4 ℃離心 30 min 後,取得胞內粗提液。
2-2-4
酵素的純化
利用親和性管柱層析法將酵素由胞內粗提液中純化,親和層析法是利用蛋白質和固相 的基質有特異性的結合而使其他雜質蛋白流過,通常會在固定相的小球上修飾和蛋白質有 強接合能力的分子。現今可以利用基因重組技術將Sortase A 的 N 端加上具親和性的 6 個組 胺酸(Histidine)稱為 His-tag,且六個組胺酸分子量很小較不會影響原有蛋白質的性質。組胺酸的imidazole ring 上的氮 (N)帶有兩個單獨電子對,於是鎳離子(Ni2+)粒子就可以吸附
上去,蛋白質因此就可以和固相的基質結合。最後要溶離出所要的蛋白質時就用未接有任
何東西的imidazole 來競爭,這是一個很溫和的方法,在溶離的過程中,蛋白質不容易變性。
首先配置4 種溶液,分別為 Buffer 1 (50 mM tris,150 mM NaCl,pH 8.0)、Buffer 2 (50 mM
tris,150 mM NaCl,50 mM imidazole,pH 8.0)、Buffer 3 (50 mM tris,150 mM NaCl,100 mM imidazole,pH 8.0)、Buffer 4 (50mM tris,150 mM NaCl,250 mM imidazole,pH 8.0)。先將層析管
柱內的溶液置換成Buffer 1,再以低流速將酵素粗提液流過管柱,Sortase A 因具有 His-tag,
故會與管柱中的鎳離子親合,再依次用Buffer2、Buffer3,讓雜質蛋白流過,最後利用 Buffer4
2-2-5
決定蛋白質分子量與純度
純化的Sortase A 蛋白質分子量(MW)與均質度(homology)之粗略估計,可藉由十
二 烷 基 硫 酸 鈉 聚 丙 烯 醯 胺 凝 膠 電 泳 法 (sodium dodecyl sulfate polyacrylamide gel
electrophoresis,SDS-PAGE)來觀測,這個系統是由 Laemmli 在 1970 年提出(48),此項技 術的原理是利用介面活性劑 SDS(十二烷基硫酸鈉)破壞蛋白質的二級結構,使其變性形 成線性長條分子,並包覆變性蛋白質,使其帶有一致的負電荷,再利用檢體中蛋白質分子 量大小的不同,使其在電泳膠片中分離,形成不同的帶狀,分子量愈大者,移動距離越短, 依此可用以檢驗蛋白質的純度,是最常用的蛋白質分析技術之一。 首先製備下列4 項膠片與藥品:
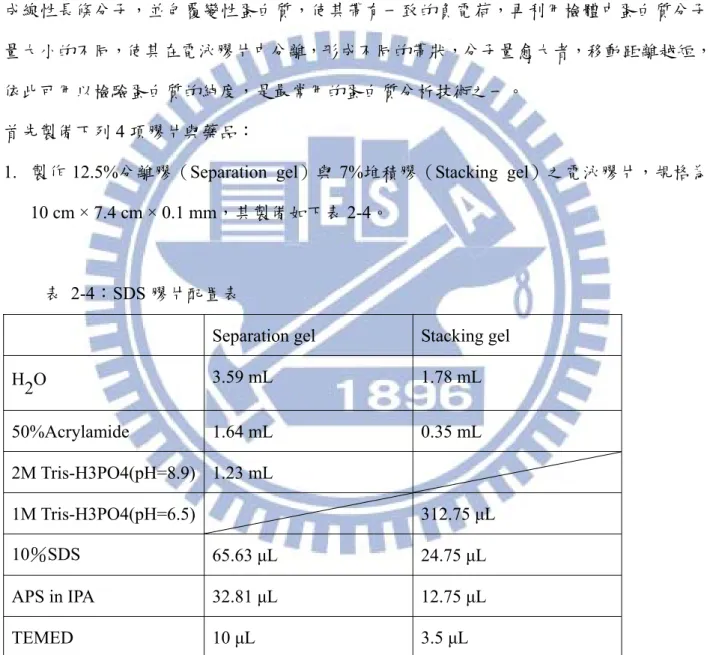
1. 製作 12.5%分離膠(Separation gel)與 7%堆積膠(Stacking gel)之電泳膠片,規格為 10 cm × 7.4 cm × 0.1 mm,其製備如下表 2-4。
表 2-4:SDS 膠片配置表
Separation gel Stacking gel
H2O 3.59 mL 1.78 mL 50%Acrylamide 1.64 mL 0.35 mL 2M Tris-H3PO4(pH=8.9) 1.23 mL 1M Tris-H3PO4(pH=6.5) 312.75 μL 10%SDS 65.63 μL 24.75 μL APS in IPA 32.81 μL 12.75 μL TEMED 10 μL 3.5 μL
2. 製備 Sample loading buffer,配置內容如下表 2-5. 表 2-5:Sample loading buffer 配置表
藥品 配置溶液體積 1M Tris-HCl(pH=6.8) 0.6 mL 50%甘油 5 mL 2-mercaptoethanol 0.5 mL 1%bromophenol blue 1 mL 10%SDS 2 mL H2O 0.9 mL Total Valume 10 mL
3. Stain solution:200 mL 醋酸,500 mL 異丙醇,3 L 去離子水,0.6 克 Coomassie blue 4. Destain solution:400 mL 醋酸,400 mL 異丙醇,3.2 L 去離子水
將純化之Sortase A 野生型與突變型各取 10 μl 溶液,加入 5 μl 的 Sample loading buffer
混合後,在95℃加熱 5 分鐘,取 5 μl 的溶液與 5 μl 的 protein maker 注入 SDS-page 的孔洞,
以電壓170 伏特進行電脉 1 小時,再將 SDS-page 浸泡在 Stain solution 中 2 小時,再以 Destain
solution 褪去背景色,可看到酵素在 SDS-page 中的位置與 protein maker 比對可估算蛋白質 的分子量與均質度。
將 純 化 之 野 生 型 與 突 變 型 的 Sortase A 以 除 鹽 管 柱 除 塩 後 進 一 步 以 MALDI
(matrix-assisted laser desorption ionization)-TOF 質譜儀鑑定蛋白質分子量。
2-2-6
蛋白質身份的鑑定
取微量SDS-page 中酵素的膠片,先依如下步驟處理:
2. 加入 50 μl wash buffer (ABC buffer,50% ACN),放置>15 min,將 gel 洗到白色。 3. 將澄清液吸出後,tube 蓋子打開,讓殘餘液體揮發。
4. 加入 50 μl 100% ACN 約 5 min。 5. 重覆步驟 3, RT 室溫約 3 min。
6. 加入 3 μl 濃度 20 μg/μl 的 trypsin (in ABC buffer)在 4 ℃下靜置 20 min。 7. 放入 37 ℃烘箱 O/N。
8. 將澄清液吸出後,tube 蓋子打開,讓殘餘液體揮發。 9. 加入 3 μl 50% ACN, 1%TFA ,sonication 20 min。
最後將處理後的樣本,用MALDI-TOF 質譜儀鑑定蛋白質的身份。
2-2-7
酵素的保存
純化之Sortase A 溶液保存在 4℃待用。或以除鹽管柱置換成 Tris buffer (50mM),經液
態氮急速冷凍後,以冷凍乾燥機抽乾,可得乾燥的粉末,冰在-20℃保存。
2-2-8
蛋白質濃度的測定
採用 Bradford Assay 吸收值方法(49),進行蛋白質濃度的測定。此定量方法利用
Coomassie Brilliant Blue G-250 與蛋白質結合的特性,取 G-250 與牛血清蛋白(bovine serum albumin,BSA)結合後,G-250 的顏色會從紅色轉變成為藍色,在 595nm 波長有強吸收, 利於偵測。
以不同濃度的蛋白質標準品建立標準測量線,再利用內插法計算待測蛋白質的濃度。 其步驟如下:
1. 從 4 ℃冷藏中拿出 2 ml 的 Bradford reagent,在室溫中回溫。
2. 取 2 μl 的 BSA 標準品(10 mg/ml)在 microtube 中,以 Tris buffer 稀釋到 100 μl 最後得
到標準品的濃度為200 μg/ml。
盤中BSA 的質量分別為 0,0.4,0.8,1.2,1.6,2,3,4 μg。 4. 加入 0.2 ml 的 Bradford reagent,在室溫中反應 2 分鐘。
5. 在 ELISA reader 上,以波長 595nm 的光測量之,可得到一標準曲線。
6. 取純化的酵素 1 μl 置於檢驗盤中,加入 Tris buffer 至 20 μl,再加入 Bradford reagent 0.2 ml
在室溫中反應2 分鐘。
7. 在 ELISA reader 上,以波長 595 nm 的光測量之,可得到酵素的吸收值,再參考標準品 的吸收值與濃度曲線,可測得酵素的濃度。
2-3 Sortase A 反應機制之研究
2-3-1
水解活性的檢測與比較
Sortase A 有水解與接合兩階段反應。以 Abz-LPETG-Dnp 來檢驗 Sortase A 野生株與突 變株的水解作用。
Abz-LPETG-Dnp 的 Abz 端具螢光性質,Dnp 端具消光性質,當連接兩者的 LPETG 被 Sortase A 切開分離成 Abz-LPET 與 G-Dnp 時,Abz 端的螢光不再被 Dnp 端扺消而呈現出來。
透過螢光光譜儀,可測量得到Abz 端的螢光會增加(42, 44)。藉由此特性,可檢測 Sortase A
的水解活性。
在200 μl 反應溶液(Tris 5 mM、NaCl 15 mM、鈣離子 2 mM)中,固定 Abz-LPETG-Dnp
的量為50 μM(溶解在 dimethyl sulfoxide 中),Sortase A 為 4 μM,設定螢光光譜儀中激發
波長為320 nm、散射波長為 420 nm,觀察 Abz-LPET 的螢光值會隨著反應時間的變化。
再以 LC/MS 做分子量的驗證。Abz_LPETG_Dnp 的分子式 C41H57N11O14,分子量為
927,在 LC/MS 檢測中應會出現帶 1 正電的分子量 928 訊號。被 sortase A 切斷後,Abz_LPET
被分解成Abz_LPET(分子式 C27H38O8N5分子量560 不帶 OH 與不帶+1 電)與 G-Dnp(分
Abz_LPET 分子量 578(加 OH 及帶+1 電)的訊號與 G-Dnp 的分子量為 369(加 H 及帶+ 1 電)的訊號。
由此紀錄比較野生株與突變株的反應速度,了解何者的水解效率最佳。
2-3-2
接合活性的檢測與比較
為比較Sortase A 野生株與突變株的接合活性,以 MRM(Multiple reaction monitoring)
方法來定義Abz-LPETGGGGGK-FITC 的產量(50)。以 GGGGGC-FITC 與 Abz-LPETG-Dnp
作為反應物,Sortase A 先將 Abz-LPETG-Dnp 水解成 Abz-LPET 後,再接合 GGGGGC-FITC,
最後形成Abz-LPETGGGGGC-FITC。
故取Abz-LPETG-Dnp 與 GGGGGC-FITC(最後濃度各為 100 μm)在反應溶液(最後
濃度Tris 5 mM、NaCl 15 mM、鈣離子 2 mM)中加入 Sortase A(最後濃度 4 μm),從 Sortase
A 加入後開始直到 60 分鐘,每隔 5 分鐘或 10 分鐘取出反應液,再加入 pH 值為 10 的 EDTA (ethylene diamine tetraacetic acid,乙二胺四乙酸,最後濃度 5 mM)(51),並在 95 ℃加熱 5
分鐘以中止反應,取得在不同時間點含Abz-LPETGGGGGC-FITC 的反應溶液。
以LC/MS 檢驗其反應物與生成物,在 MS 圖譜上除了看到 Abz-LPETG-Dnp 分子量 928
的訊號、Abz-LPET 分子量 578 的訊號、G-Dnp 分子量 369 的訊號,以外應還可看到反應物
的GGGGGK-FITC 的訊號及經 Sortase A 作用後 Abz_LPET 轉移接上 GGGGGK-FITC,形
成的生成物Abz-LPETGGGGGK-FITC 訊號。GGGGGK-FITC 的分子式 C37H40O12N8S 分子 量820,在 LC/MS 檢測中應會出現帶 1 個正電的質荷比 821 或帶 2 個正電的質荷比 411 訊號。 Abz-LPETGGGGGK-FITC 的分子式為 C64H78O14N11分子量為1380,在質譜中應會出現帶 1 個正電的質荷比1381 或帶 2 個正電的質荷比 691 訊號。 將反應溶液進行LC/MS 分析(分析條件如表 2-6)並對 Abz-LPETGGGGGC-FITC 進 行MRM 分析,透過特定碎片 FITC 胜肽段選定的 Abz-LPETGGGGGC-FITC 分子,計算其 質譜儀上的面積積分值,代表Abz-LPETGGGGGC-FITC 的生成量值。將所得的量值對時間 點作圖,可了解Sortase A 野生株與突變株的反應速率何者較佳。
表 2-6:LC-MASS 的分析條件表 時間(分) 流速(ml/min) 水 ACN (Acetonitrile,CH3CN) 0 1 90 10 1 1 80 20 7 1 35 65 20 1 10 90 30 1 0 100 32 1 90 10 35 1 90 10
第三章 結果與討論
3-1 酵素的基因序列、表現與純化
3-1-1
酵素的基因序列
在DNA 定序後,取得野生型與各突變株的胺基酸序列如下圖 3-1:SrtW 為 Sortase A
去除N 端 24 個胺基酸所表現的野生株(14, 16, 17)。以 SrtW 為模版做單一定點突變得到
Srt196 為 Sortase A 的 K196T 突變株,Srt165 為 Sortase A 的 D165A 突變株。為對 Sortase 進
行P94S、D160N、D165A、K196T 的點突變,本研究縮短質體長度去除 N 端 59 個胺基酸
保留有活性的區域(第60 個胺基酸 Q 至第 206 個胺基酸 K)(47)以利於點突變的疊加,
最後得到Sortase A 去除 N 端 59 個胺基酸且具有 P94S、D160N、D165A、K196T 點突變所
圖 3-1:Sortase A 野生型及突變株胺基酸定序表
3-1-2
酵素的表現與純化
一、Sortase A 野生株的表現與純化 Sortase A 野生株與突變株均能在胞內液中過量表現,純化後的酵素在 SDS-page 上估 測的分子量SortaseW、D165A、K196T 約 23000,如圖 3-2(A)中所示;SortasePro 約 17000, 如圖3-2(B)中所示。maker SrtW_crude 圖 3-2:Sortase A 野生株及突變株的 SDS-page 蛋白質分子量 二、Sortase A 野生株的 MALDI-TOF 分析 蛋白質經trypsin處理後,會得到不同長度的片段,因Trypsin會水解蛋白質中所有的由 賴氨酸Lys或精氨酸Arg的羧基所構成的肽鍵,不過如果這兩個部份在C-terminal方向連接的 是proline,它就不會水解這個地方的鍵結(52)。所以經trypsin作用之後得到的片段,以 MALDI-TOF質譜儀分析,比對Matrix Science線上資料庫中Sortase A的胺基酸序列後,胺基 酸序列涵蓋度(sequence coverage MS)為54.7%,有部份重疊如下圖3-3,且其Mascot score >52、significance threshold p<0.05如圖3-4,證實所純化的蛋白質為Sortase A。
SrtW D165A K196T 70K 70K 53K 53K 41K 41K 30K 30K SrtPro 22K 22K 10K
(A) (B)
maker圖 3-3:Sortase A 野生株的 MALDI-TOF 分析圖譜
3-1-3
Sortase 野生株與突變株的分子量
以MALDI-TOF 質譜儀檢測 Sortase A 野生株與突變株的分子量,分析圖譜如附錄二所 示,得到下列結果:SrtW 的分子量為 22970.9,其理論值 23070.79,相對誤差 0.43%;D165A 的分子量為22942.0,其理論值 23026.78,相對誤差 0.37%;K196T 的分子量為 22902.4, 其理論值23043.72,相對誤差 0.61%;Srt Pro 的分子量為 16643.7,其理論值 17540.81,相 對誤差 5.11%。檢測結果符合預測的分子量,故經蛋白質純化後取得之野生株與突變株的 分子量以檢測值為準來計算對應的酵素濃度。3-1-4
野生型酵素濃度測定
使用的蛋白質標準品為牛血清蛋白(BSA),濃度為 10 mg/ml。將蛋白質標準品與緩衝 液混合,形成不同濃度的標準品,總體積為20μl,再加入 Bradford reagent 至 200 μl,放置 在室溫下2 分鐘,以 UV 光譜儀讀取各樣品 OD595 nm 之吸收值如表 3-1。 表 3-1、不同濃度 BSA 標準品與 OD595 之吸收值 BSA (μg) OD595 nm 0 0.332 0.4 0.35 0.8 0.373 1.2 0.4 1.6 0.414 2 0.452 3 0.513 4 0.572根據UV 光譜儀讀取各樣品 OD595 nm 之吸收值劃出蛋白質標準品對 OD595 nm 之標
準檢量線,如圖3-5。故得到 OD595 nm 的量值(y 值)與 BSA 濃度(x 值)的關係為
y=0.0613x+0.3261(檢量公式)。 y = 0.0613x + 0.3261 R2 = 0.9966 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0 1 2 3 4 5 BSA(μg) OD5 9 5 圖 3-5:蛋白質標準品對 OD595 之標準檢量線 取1 μl 待測蛋白質溶液,所測得對應的 OD595 吸收值如下:Srt W 為 0.4896、D165A 為0.483、K196T 為 0.443、SrtPro 為 0.3825,以內插法帶入檢量公式,可得 1μl 待測蛋白質 溶液的濃度如下:Srt W 為 2.6672 (μg/μl)、D165A 為 2.5595 (μg/μl)、K196T 為 1.9070 (μg/μl)、
SrtPro 為 0.9201 (μg/μl),再參考 MALDI-TOF 分析儀鑑定 Sortase A 野生株與突變株的分子
量,如表3-2 所示,可得各溶液的莫耳濃度如下:Srt W 為 115.61 μM、D165A 為 111.16μM、
表 3-2:Sortase A 野生株與突變株的濃度 1(μl) y(od) x(μg) MW (μM) Srt W 0.4896 2.6672 23070.79 115.61 D165A 0.483 2.5595 23026.78 111.16 K196T 0.443 1.9070 23043.72 82.76 SrtPro 0.3825 0.9201 18000 51.11
3-2 野生型與突變型酵素的活性檢測
3-2-1
Sortase A 野生株的活性檢測
Sortase A 對 Abz-LPETG-Dnp 水解後,得到產物 Abz-LPET 與 G-Dnp,其 Abz-LPET 在 320 nm 的螢光激發下會有 420 nm 的波長散射,在螢光光譜儀可見螢光量值隨時間的增加
而增加,驗證Sortase A 在 pH 為 8 且常溫的環境中具有水解活性,如圖 3-7。
以LC-MASS 驗證其反應物與生成物。可見 Abz_LPETG_Dnp 荷質比 928 的訊號。被
Sortase A 水解後,也出現 Abz_LPET 荷質比 578 的訊號及 G_Dnp 荷質比 369 的訊號,如
0924SrtW_ALD Time 2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00 22.00 24.00 26.00 28.00 30.00 32.00 34.00 % 0 100 2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00 22.00 24.00 26.00 28.00 30.00 32.00 34.00 % 0 100 2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00 22.00 24.00 26.00 28.00 30.00 32.00 34.00 % 0 100 2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00 22.00 24.00 26.00 28.00 30.00 32.00 34.00 % 0 100
0924SrtW_ALD Scan ES+
369 3.93e7
9.93
29.39 12.60
0924SrtW_ALD Scan ES+
578 5.90e7
10.89
0924SrtW_ALD Scan ES+
928 6.01e7
12.54
0924SrtW_ALD Scan ES+
TIC 4.65e8 12.60 4.38 2.74 10.89 10.27 34.11 25.82 23.97 22.95 14.93 18.08 29.25 30.07 7.26 7.06 5.27 7.47 9.93 11.17 31.72 33.77 0924SrtW_ALD m/z 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100
0924SrtW_ALD 184 (12.604) Sb (3,40.00 ) Scan ES+
1.51e7 928.5 456.2 424.2 293.2 369.2 456.7 929.5 930.4 464.8 696.4 483.7 679.3 560.3 697.5 0924SrtW_ALD m/z 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100
0924SrtW_ALD 159 (10.891) Sb (3,40.00 ) Scan ES+
9.29e6 578.3 346.3 293.2 387.0 528.3 579.2 580.3 616.3 0924SrtW_ALD m/z 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100
0924SrtW_ALD 145 (9.932) Sb (3,40.00 ) Scan ES+ 5.80e6 369.2 293.2 293.4 369.4 370.2 293.6 352.4 387.0 688.2780.0794.6811.2892.0 991.3 圖 3-6:Abz_LPETG_Dnp 被 Sortase A 分解的 LC/MS 圖譜
3-2-2
水解活性與比較
固定Abz-LPETG-Dnp 濃度為 50 μM, Sortase A 野生株與突變株的濃度為均為 4 μM,由 螢光光譜儀的測量由加入酵素開始,可得到螢光值與反應時間的關係圖,如圖 3-7。(螢光 量值對時間的紀錄值,如附錄三)由圖中可見Sortase A 野生株與突變株在 120 秒內螢光值 漸漸平緩,以SrtPro(藍色線)與 D165A(粉紅色線)反應最快,SrtW(黃色線)次之, K196T(綠色線)最差,對照組(紫色線)無酵素作用,故螢光量值未有顯著的增加。0
500
1000
1500
2000
2500
3000
0
20
40
60
80
100
120
(sec)
SrtPro D165A SrtW K196T blank 圖 3-7:Sortase A 野生株與突變株水解 Abz-LPETG-Dnp 的螢光值對時間關係圖 取3~13 秒的數值作水解反應初速率的比較圖如下圖 3-8,由圖中可得到反應初速率的 斜率SrtPro 為 68.986、D165A 為 55.418、SrtW 為 21.997、K196T 為 18.665,而對照組的斜 率為2.856(野生株與突變株線型如圖例所示)。可知對Abz-LPETG-Dnp 水解反應初速率的 大小依次為SrtPro、D165A、SrtW、K196T。y = 68.986x + 830.99
y = 55.418x + 945.84
y = 21.997x + 710.73
y = 18.665x + 637.77
y = 2.8564x + 265.62
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
1
2
3
4
5
6
7
8
9 10 11 12 13 14
(sec)
SrtPro D165A SrtW K196T blank 線性 (SrtPro) 線性 (D165A) 線性 (SrtW) 線性 (K196T) 線性 (blank) 圖 3-8:Sortase A 野生株與突變株水解活性的反應初速率3-2-3
接合活性的檢測
以 GGGGGK-FITC 與 Abz_LPETG_Dnp 檢 測 SrtW 的 活 性 , 在 質 譜 上 可 見 GGGGGK-FITC 荷質比為 411 的訊號(帶 2 個正電),Abz-LPETGGGGGK-FITC 質荷比為 691 訊號(帶 2 個正電),如圖 3-9。 0924SrtW_reac Time 2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00 22.00 24.00 26.00 28.00 30.00 32.00 34.00 % 0 1000924SrtW_reac Scan ES+
TIC 5.17e8 15.07 13.01 12.26 9.80 8.49 14.45 14.11 34.18 24.87 23.43 22.47 21.92 7.67 6.10 10.00 18.77 20.21 29.45 31.10 33.91 33.09 2.81 2.12
0924SrtW_reac m/z 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100
0924SrtW_reac 211 (14.453) Sb (3,40.00 ) Scan ES+
1.21e7 690.9 690.7 691.2 691.4 691.7 473.7 383.7 293.3 468.3 574.7 573.7 0924SrtW_reac m/z 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100
0924SrtW_reac 220 (15.070) Scan ES+ 1.67e7 928.5 456.2 424.2 250.3 929.5 369.2 323.1 456.7 930.5 696.4 483.7 679.3 931.4 697.5 0924SrtW_reac m/z 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100
0924SrtW_reac 190 (13.015) Sb (3,40.00 ) Scan ES+
1.39e7 369.2 293.2 352.2 369.3 411.1 411.6 798.7 709.5 821.2 0924SrtW_reac m/z 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100
0924SrtW_reac 206 (14.111) Sb (3,40.00 ) Scan ES+
4.64e6 578.2 346.1 293.1 387.1 419.0 496.7 578.4 579.2 961.9 580.4 627.8 936.0 1075.7
對質荷比691 訊號的分子作 MSMS 分析,得到大量的荷質比 233.1、390.2、574.9、759.3 碎片,如圖3-10。 srtW-691MSMS m/z 100 200 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 % 0 100 2012120605 294 (10.339) Cm (293:296) Daughters of 691ES+ 3.11e5 574.9 390.2 233.1 227.2 575.1 575.4 390.3 574.5 759.3 575.7 391.2 691.5 619.3 759.6 760.5 392.4 328.0 233.5 761.1 461.9 560.3 761.6 199.1 821.4 923.6992.3 圖 3-10:生成物荷質比691 訊號的MSMS 分析 比對碎片的荷質比與胜肽片斷的理論值分子量,其中訊號強度較大的 233.1 的訊號來 自於胜肽片斷 Abz-L 分子式 C13H17O2N2分子量理論值 232,若帶 1 個正電,則荷質比為 233;390.2 的訊號來自於胜肽片斷 FITC 分子式 C21H12O5NS 分子量理論值 390;574.9 的 訊號來自於胜肽片斷PETGGGGGK-FITC 分子式 C51H60O18N11S 分子量理論值 1146,若帶 2 個正電,則荷質比為 574,考慮有 2 個 C 同位素的存在,則荷質比為 575;759.3 的訊號 來自於胜肽片斷PETGGGGGK 分子式 C30H48O13N10S 分子量理論值 756,若帶 1 個正電, 則荷質比為577,考慮有 2 個 C 同位素的存在,則荷質比為 579。其它訊號強度較弱的碎片, 也符合其理論值分子量的荷質比,如表 3-3。故在 LC-Mass 中荷質比 691 的訊號表示 Abz-LPETGGGGGK-FITC 被 Sortase A 合成。
表 3-3:Abz_LPETGGGGGKFITC 訊號的 MSMS 分析表 MS 中的荷質比 胜肽片斷的推測 胜肽片斷的理論值分子量 227.2 PE 226 233.1 Abz-L 232 328 PET 327 390.2 FITC 390 461.9 LPET 460 560.3 Abz-LPET 560.3 574.9 PETGGGGGK-FITC 1146 619.3 Abz-LPETG 617 691.5 Abz-LPETGGGGGK-FITC 1379 759.3 PETGGGGGK 756 821.4 GGGGGK-FITC 820 923.6 TGGGGGK-FITC 921 992.3 Abz-LPETGGGGGK 989
3-2-4
接合活性的比較
Abz-LPETGGGGGK-FITC 為接合生成物,故取其中 FITC 片斷 390 的訊號峰值作為 Abz-LPETGGGGGK-FITC 的參照片斷,作 MRM 分析,如圖 3-11。在 10.39 分時檢測到 Abz-LPETGGGGGK-FITC 的產物,其 MRM 圖譜面積積分值表示產量,此面積之荷質比訊 號單純為691,表示此其產量的計算不受其它物質的干擾。圖 3-11:Abz-LPETGGGGGK-FITC 的 MRM 分析 每隔5 分鐘所取得的 MRM 圖譜面積積分值如表 3-4(各時間點的 MRM 圖譜見附錄四) 表 3-4:每隔 5 分鐘的 Abz-LPETGGGGGK-FITC 之 MRM 面積積分值 time(min) 0 5 10 15 20 25 30 35 SrtPro 3500 13348 19205 24671 31497 40319 44820 48597 D165A 2186 12029 15367 20792 26666 34726 38345 41400 SrtW 2533 8621 14496 19839 23694 34546 35922 40010 K196T 2735 4761 9962 15316 24594 27408 35631 33243 依此表作時間對MRM 面積的關係圖,如圖 3-12。可見以 SrtPro 的接合反應最佳,D165 次之,K196T 最差。
0
10000
20000
30000
40000
50000
60000
0
10
20
30
40
(min)
MR
M_
ar
ea
SrtPro
D165A
SrtW
K196T
圖 3-12:時間對 MRM 面積的關係圖 取0~30 分鐘的數值作接合反應初速率的比較圖如下圖 3-13,得到接合反應初速率的 斜率SrtPro 為 1358.5、D165A 為 1179.8、SrtW 為 1151.5、K196T 為 1133(各酵素的線型 如圖例所示),可知對 Abz-LPETG-Dnp 與 GGGGGFITC 的接合反應初速率的大小依次為 SrtPro、D165A、SrtW、K196T。y = 1358.5x + 4959.2
y = 1179.8x + 3747.5
y = 1151.5x + 2677.1
y = 1133x + 206.78
0
10000
20000
30000
40000
50000
0
5
10
15
20
25
30
35
min
MR
M_
ar
ea
SrtPro D165A SrtW K196T 線性 (SrtPro) 線性 (D165A) 線性 (SrtW) 線性 (K196T) 圖 3-13: Sortase A 野生株與突變株接合活性的反應初速率3-2-5
野生株與突變株的活性比較討論
以SrtW 的水解初速率與水解接合初速率為基準值,可得突變株的反應初速率倍數關 係,如下表3-5,令 SrtW 的水解倍數與接合倍數為 1;SrtPro 的水解倍數為 3.14, 接合倍數 為1.18;D165A 的水解倍數為 2.52, 接合倍數為 1.02;K196T 的水解倍數為 0.85, 接合倍 數為0.98。 表 3-5:野生株與突變株的活性比較表 種類 水解反應初速率 水解倍數 接合反應初速率 接合倍數 SrtPro 68.986 3.14 1358.5 1.18 D165A 55.418 2.52 1179.8 1.02 SrtW 21.997 1.00 1151.5 1.00 K196T 18.665 0.85 1133 0.98 比較水解倍數,可知對Abz-LPETG-Dnp 的水解活性大小依次為 SrtPro、D165A、Srt W、 K196T。由接合反應初速率的接合倍數,可知對 Abz-LPETG-Dnp 與 GGGGGFITC 的接合 活性大小,依次為SrtPro、D165A、SrtW、K196T。 而D165A 的水解倍數為 2.52,接合倍數為 1.02,顯示此突變有利於 LPETG 的水解反 應,但對GGGGG 端的接合反應並無有效的改善。K196T 的水解倍數為 0.85,接合倍數為 0.98,顯示此突變雖不利於 LPETG 的水解反應,但接合反應倍數與野生株相近,故 K196T的突變有利於GGGGGK-FITC 的結合。而 SrtPro 的水解倍數為 3.14,較 D165A 的水解倍
數2.52 為大,故推測 P94S 與 D160N 的突變點,也有利於 LPETG 的水解反應速率。
綜合以上討論P94S、D160N、D165A 能增快水解速率,有利於 LP 殘基的結合,K196T
則有利於GGGGG 殘基的結合。其中 D160N、D165A 位於 β6/β7 的螺旋上,K196T 位於 β7/β8
的螺旋上,與文獻探討中β6/β7 區域與辨認 LPXTG 中的 LP 殘基有關(35-37) ,β7/β8 區域
在Chen, et al.(2011)的研究中(46)指出 LPETG 被 SrtPro 催化的 Kcat(最大催化速率)
為4.8、SrtW 的 Kcat 為 1.5、D165A 的 Kcat 為 2.4、K196T 的 Kcat 為 1.2,令 SrtW 的催化
速率為基準值1,則 SrtPro 的催化倍數為 3.2、D165A 的催化倍數為 1.6、K196T 的催化倍數 為0.8。以此數據與本研究相比,其相對誤差可見 SrtPro 為-1.9%、K196T 為 6.3%,如下 表3-6。而 D165A 的相對誤差雖為 57.5%,但仍證明 D165A 的突變有助於水解反應初速率 的增加。故本研究實驗結果與之相符。 表 3-6:Sortase A 水解反應研究結果之相對誤差 種類 水解倍數 (本研究) 最大催化速率 (Irwin Chen) 催化倍數 (Irwin Chen) 相對誤差% [=(水解倍數-催化倍 數)/催化倍數*100%] SrtPro 3.14 4.8 3.2 -1.9 D165A 2.52 2.4 1.6 57.5 SrtW 1 1.5 1 0 K196T 0.85 1.2 0.8 6.3
3-3 Sortase A 四點突變株之應用
在石英晶體微量天平(QCM)上修飾 LPETGLPETG 的胜肽段(53),藉著 SrtPro 將 GGGGGK圖 3-14:SrtPro 在 QCM 上的應用示意圖
取SrtPro 濃度 50μM,GGGGGK-FITC 濃度 500μM,作用在修飾LPETGLPETG 的QCM
上反應12 小時,得到的顯微照片如下圖 3-15。可知當一端受質為固定端時,SrtPro 仍保有 活性。 對照組 實驗組 圖 3-15:接合 GGGGGK-FITC 的 QCM 晶片圖
SiO
2 O Si O OG
T
E
P
L
GGGGGK-FITC
O Si O OT
E
P
L
GGGGGK-FITC
SrtPro
第四章 結論
在取得 Srt W、D165A、K196T 與 SrtPro 的酵素過程中,發現 Sortase A 的野生株與
D165A 、 K196T 、 SrtPro 此 三 種 突 變 株 均 能 在 胞 內 液 中 大 量 表 現 , 且 均 能 水 解 Abz-LPETG-Dnp 後與 GGGGGK-FITC 接合,生成 Abz-LPETGGGGGK-FITC。故 Sortase A
的野生株與D165A、K196T、SrtPro 的突變株在水溶液中均能呈現水解活性與接合活性。 以螢光光譜儀檢測 Abz-LPETG-Dnp 被水解後的螢光值,發現突變株 D165A、SrtPro 能增快水解速率,且 SrtPro 的水解倍數 3.14 大於 D165A 的水解倍數 2.52,顯示 P94S、 D160N、D165A,有利於 LP 殘基的結合而增加水解的反應速率。K196T 水解倍數只有 0.85 但接合倍數為0.98 顯示 K196T 則有利於 GGGGG 殘基的結合,增加接合的反應速率。在比 較突變株的水解與接合反應初速率後,顯示突變株SrtPro 比野生株 SrtW 在水解初速率上大 3.14 倍,在接合初速率上大 1.18 倍。故可以用 SrtPro 取代 SrtW 進行反應。 研究過程中發現水解反應在 1 分鐘時已接近反應速率的最大值,而接合反應在 30 分 鐘 後 反 應 速 率 才 減 緩 , 故 可 知 Sortase A 的作用雖遵循「乒乓機制」(44) ,先水解
Abz-LPETG-Dnp 再接合 GGGGGFITC,但要縮短接合反應的時間較能有效縮短 SortaseA 的 作用時間。
最後將SrtPro 應用在 QCM 的研究發現,雖然 LPETG 端固定在 QCM 晶片上非在溶液
懸浮狀態,但懸浮在溶液中的GGGGGK-FITC 仍能被接合在 LPET 上,表示 SrtPro 在此狀
第五章 未來研究方向
在實驗過程中探討接合反應速率時,以每隔5 分鐘取得 MRM 面積的方法呈現酵素的
反應量值,可進一步設計以螢光共振能量轉移(Fluorescence resonance energy transfer, FRET)的方式,藉由觀察 FRET 的螢光量值了解 Sortase A 即時的接合速率反應量值。如
以 Rhodamine-QALPETG 與 GGGGGFITC 為 受 質 , 當 Sortase A 作 用 後 形 成
Rhodamine-QALPETGGGGGK-FITC,以 450nm 的螢光激發 FITC,若有產物則發散波長會
由530nm 轉移至 585nm,在螢光光譜上,可藉由觀察 585nm 波長的變化測得 Sortase A 接
合反應的速率 (54)。
圖 5-1:Sortase A 的 FRET 研究示意圖 (54)
而 本 研 究 試 以 Rhodamine-QALPETG 為 受 質 研 究 其 接 合 反 應 時 , 發 現
Rhodamine-QALPETG 在 Sortase A 活性溶液(Tris5mM、NaCl 15mM、鈣離子 2 mM)中螢
光會消失,將溶液中的Tris 降低至 5μM 則螢光才會又出現,但 Sortase A 活性在此低濃度
鹽 類 環 境 中 活 性 降 低 , 不 利 於 進 行 Sortase A 反 應 速 率 的 探 討 。 而 在 低 濃 度 鹽 類
Rhodamine-QALPETG 螢光消失的現象,未來可能是研究探討的方向。
實驗結論中顯示縮短接合反應的時間較能有效縮短 SortaseA 的作用時間,故在 β7/β8
(D185~K196)的螺旋上進行有利於辦認 GGGGG 受質的胺基酸突變,尚有很多的研究空
但最佳反應條件,如 pH 值、溫度、反應時間等,仍需再做研究。且雖然 SrtPro 具有接合
的專一性,但要修飾不同的抗體在 QCM 晶片上做為探針進行免疫檢查時,將胜肽段置換
References
1. Dawson, P. E., and Kent, S. B. (2000) Synthesis of native proteins by chemical ligation,
Annu Rev Biochem 69, 923-960.
2. David, R., Richter, M. P., and Beck-Sickinger, A. G. (2004) Expressed protein ligation.
Method and applications, Eur J Biochem 271, 663-677.
3. Ayers, B., Blaschke, U. K., Camarero, J. A., Cotton, G. J., Holford, M., and Muir, T. W.
(1999) Introduction of unnatural amino acids into proteins using expressed protein ligation, Biopolymers 51, 343-354.
4. Levary, D. A., Parthasarathy, R., Boder, E. T., and Ackerman, M. E. (2011)
Protein-protein fusion catalyzed by sortase A, PLoS One 6, e18342.
5. Strijbis, K., Spooner, E., and Ploegh, H. L. (2012) Protein ligation in living cells using
sortase, Traffic 13, 780-789.
6. Matsumoto, T., Tanaka, T., and Kondo, A. (2012) Sortase A-catalyzed site-specific
coimmobilization on microparticles via streptavidin, Langmuir 28, 3553-3557.
7. Dramsi, S., Trieu-Cuot, P., and Bierne, H. (2005) Sorting sortases: a nomenclature
proposal for the various sortases of Gram-positive bacteria, Res Microbiol 156, 289-297.
8. Comfort, D., and Clubb, R. T. (2004) A comparative genome analysis identifies distinct
sorting pathways in gram-positive bacteria, Infect Immun 72, 2710-2722.
9. Maresso, A. W., and Schneewind, O. (2008) Sortase as a target of anti-infective therapy,
Pharmacol Rev 60, 128-141.
10. Klebba, P. E., Charbit, A., Xiao, Q., Jiang, X., and Newton, S. M. (2012) Mechanisms of
iron and haem transport by Listeria monocytogenes, Mol Membr Biol 29, 69-86.
11. Swaminathan, A., Mandlik, A., Swierczynski, A., Gaspar, A., Das, A., and Ton-That, H.
(2007) Housekeeping sortase facilitates the cell wall anchoring of pilus polymers in Corynebacterium diphtheriae, Mol Microbiol 66, 961-974.
12. Marraffini, L. A., Dedent, A. C., and Schneewind, O. (2006) Sortases and the art of anchoring proteins to the envelopes of gram-positive bacteria, Microbiol Mol Biol Rev 70, 192-221.
13. Mazmanian, S. K., Liu, G., Jensen, E. R., Lenoy, E., and Schneewind, O. (2000)
Staphylococcus aureus sortase mutants defective in the display of surface proteins and in the pathogenesis of animal infections, Proc Natl Acad Sci U S A 97, 5510-5515.
14. Mazmanian, S. K., Liu, G., Ton-That, H., and Schneewind, O. (1999) Staphylococcus
aureus sortase, an enzyme that anchors surface proteins to the cell wall, Science 285, 760-763.
15. Driessen, A. J., Fekkes, P., and van der Wolk, J. P. (1998) The Sec system, Curr Opin
Microbiol 1, 216-222.
16. Bentley, M. L., Lamb, E. C., and McCafferty, D. G. (2008) Mutagenesis studies of
substrate recognition and catalysis in the sortase A transpeptidase from Staphylococcus aureus, J Biol Chem 283, 14762-14771.
17. Mao, H., Hart, S. A., Schink, A., and Pollok, B. A. (2004) Sortase-mediated protein
ligation: a new method for protein engineering, J Am Chem Soc 126, 2670-2671.
18. Popp, M. W., Antos, J. M., Grotenbreg, G. M., Spooner, E., and Ploegh, H. L. (2007)
Sortagging: a versatile method for protein labeling, Nat Chem Biol 3, 707-708.
19. Tanaka, T., Yamamoto, T., Tsukiji, S., and Nagamune, T. (2008) Site-specific protein
modification on living cells catalyzed by Sortase, Chembiochem 9, 802-807.
20. Pritz, S., Wolf, Y., Kraetke, O., Klose, J., Bienert, M., and Beyermann, M. (2007)
Synthesis of biologically active peptide nucleic acid-peptide conjugates by sortase-mediated ligation, J Org Chem 72, 3909-3912.
21. Samantaray, S., Marathe, U., Dasgupta, S., Nandicoori, V. K., and Roy, R. P. (2008)
Peptide-sugar ligation catalyzed by transpeptidase sortase: a facile approach to neoglycoconjugate synthesis, J Am Chem Soc 130, 2132-2133.
22. Parthasarathy, R., Subramanian, S., and Boder, E. T. (2007) Sortase A as a novel molecular "stapler" for sequence-specific protein conjugation, Bioconjug Chem 18, 469-476.
23. Chan, L., Cross, H. F., She, J. K., Cavalli, G., Martins, H. F., and Neylon, C. (2007)
Covalent attachment of proteins to solid supports and surfaces via Sortase-mediated ligation, PLoS One 2, e1164.
24. Clow, F., Fraser, J. D., and Proft, T. (2008) Immobilization of proteins to biacore sensor
chips using Staphylococcus aureus sortase A, Biotechnol Lett 30, 1603-1607.
25. Wu, Z., Guo, X., Wang, Q., Swarts, B. M., and Guo, Z. (2010) Sortase A-catalyzed
transpeptidation of glycosylphosphatidylinositol derivatives for chemoenzymatic synthesis of GPI-anchored proteins, J Am Chem Soc 132, 1567-1571.
26. Antos, J. M., Miller, G. M., Grotenbreg, G. M., and Ploegh, H. L. (2008) Lipid
modification of proteins through sortase-catalyzed transpeptidation, J Am Chem Soc 130, 16338-16343.
27. Wu, S., and Proft, T. (2010) The use of sortase-mediated ligation for the immobilisation of
bacterial adhesins onto fluorescence-labelled microspheres: a novel approach to analyse bacterial adhesion to host cells, Biotechnol Lett 32, 1713-1718.
28. Sakamoto, T., Sawamoto, S., Tanaka, T., Fukuda, H., and Kondo, A. (2010)
Enzyme-mediated site-specific antibody-protein modification using a ZZ domain as a linker, Bioconjug Chem 21, 2227-2233.
29. Popp, M. W., Dougan, S. K., Chuang, T. Y., Spooner, E., and Ploegh, H. L. (2011)
Sortase-catalyzed transformations that improve the properties of cytokines, Proc Natl
Acad Sci U S A 108, 3169-3174.
30. Refaei, M. A., Combs, A., Kojetin, D. J., Cavanagh, J., Caperelli, C., Rance, M., Sapitro,
J., and Tsang, P. (2011) Observing selected domains in multi-domain proteins via sortase-mediated ligation and NMR spectroscopy, J Biomol NMR 49, 3-7.
31. Pritz, S., Wolf, Y., Kraetke, O., Klose, J., Bienert, M., and Beyermann, M. (2009)
Enzymatic ligation of peptides, peptide nucleic acids and proteins by means of sortase A,
Adv Exp Med Biol 611, 107-108.
32. Proft, T. (2010) Sortase-mediated protein ligation: an emerging biotechnology tool for
protein modification and immobilisation, Biotechnol Lett 32, 1-10.
33. Ton-That, H., Mazmanian, S. K., Faull, K. F., and Schneewind, O. (2000) Anchoring of
surface proteins to the cell wall of Staphylococcus aureus. Sortase catalyzed in vitro transpeptidation reaction using LPXTG peptide and NH(2)-Gly(3) substrates, J Biol Chem
275, 9876-9881.
34. Ilangovan, U., Ton-That, H., Iwahara, J., Schneewind, O., and Clubb, R. T. (2001)
Structure of sortase, the transpeptidase that anchors proteins to the cell wall of Staphylococcus aureus, Proc Natl Acad Sci U S A 98, 6056-6061.
35. Zong, Y., Bice, T. W., Ton-That, H., Schneewind, O., and Narayana, S. V. (2004) Crystal
structures of Staphylococcus aureus sortase A and its substrate complex, J Biol Chem 279, 31383-31389.
36. Naik, M. T., Suree, N., Ilangovan, U., Liew, C. K., Thieu, W., Campbell, D. O., Clemens,
J. J., Jung, M. E., and Clubb, R. T. (2006) Staphylococcus aureus Sortase A transpeptidase. Calcium promotes sorting signal binding by altering the mobility and structure of an active site loop, J Biol Chem 281, 1817-1826.
37. Bentley, M. L., Gaweska, H., Kielec, J. M., and McCafferty, D. G. (2007) Engineering the
substrate specificity of Staphylococcus aureus Sortase A. The beta6/beta7 loop from SrtB confers NPQTN recognition to SrtA, J Biol Chem 282, 6571-6581.
38. Zong, Y., Mazmanian, S. K., Schneewind, O., and Narayana, S. V. (2004) The structure of
sortase B, a cysteine transpeptidase that tethers surface protein to the Staphylococcus aureus cell wall, Structure 12, 105-112.
Purification and characterization of sortase, the transpeptidase that cleaves surface proteins of Staphylococcus aureus at the LPXTG motif, Proc Natl Acad Sci U S A 96, 12424-12429.
40. Frankel, B. A., Tong, Y., Bentley, M. L., Fitzgerald, M. C., and McCafferty, D. G. (2007)
Mutational analysis of active site residues in the Staphylococcus aureus transpeptidase SrtA, Biochemistry 46, 7269-7278.
41. Connolly, K. M., Smith, B. T., Pilpa, R., Ilangovan, U., Jung, M. E., and Clubb, R. T.
(2003) Sortase from Staphylococcus aureus does not contain a thiolate-imidazolium ion pair in its active site, J Biol Chem 278, 34061-34065.
42. Ton-That, H., Mazmanian, S. K., Alksne, L., and Schneewind, O. (2002) Anchoring of
surface proteins to the cell wall of Staphylococcus aureus. Cysteine 184 and histidine 120 of sortase form a thiolate-imidazolium ion pair for catalysis, J Biol Chem 277, 7447-7452.
43. Adler, A. J., Greenfield, N. J., and Fasman, G. D. (1973) Circular dichroism and optical
rotatory dispersion of proteins and polypeptides, Methods Enzymol 27, 675-735.
44. Huang, X., Aulabaugh, A., Ding, W., Kapoor, B., Alksne, L., Tabei, K., and Ellestad, G.
(2003) Kinetic mechanism of Staphylococcus aureus sortase SrtA, Biochemistry 42, 11307-11315.
45. Frankel, B. A., Kruger, R. G., Robinson, D. E., Kelleher, N. L., and McCafferty, D. G.
(2005) Staphylococcus aureus sortase transpeptidase SrtA: insight into the kinetic
mechanism and evidence for a reverse protonation catalytic mechanism, Biochemistry 44, 11188-11200.
46. Chen, I., Dorr, B. M., and Liu, D. R. (2011) A general strategy for the evolution of
bond-forming enzymes using yeast display, Proc Natl Acad Sci U S A 108, 11399-11404.
47. Marraffini, L. A., Ton-That, H., Zong, Y., Narayana, S. V., and Schneewind, O. (2004)
Anchoring of surface proteins to the cell wall of Staphylococcus aureus. A conserved arginine residue is required for efficient catalysis of sortase A, J Biol Chem 279,