Pathogenicity of a baculovirus infection causing
white spot syndrome in cultured penaeid shrimp
in Taiwan
Hsin-Yiu C h ~ u ' ~ * ' * * ,
Chang-Yi Huangl, Chung-Hsiung wang2,
Hsien-Choung Chiang3, Chu-Fang Lo4,**
'
Department of Aquaculture, National Taiwan Ocean University, Keelung. Taiwan 20224, ROC *Department of Plant Pathology, National Taiwan University, Taipei, Taiwan 106, ROCI-Lan Livestock Disease Control Center, I-Lan, Taiwan 260. ROC Department of Zoology, National Taiwan University, Taipei, Taiwan 106, ROC
ABSTRACT. Outbreaks of a disease causing serious losses among populations of cultured penaeid shrimp, including Penaeus monodon, P japonicus and P penicillatus, in Taiwan are characterized by obvious white spots on the carapace, appendages and the inside surface of the body. For this reason, we propose the name 'white spot syndrome' (WSS). Healthy juvenile kuruma shrimp P japonjcus were exposed by immersion to epidermal filtrate from diseased P japonicus and L? monodon which exhibited marked white spot signs. These experimentally infected shrimp resembled the spontaneously infected ones. Cumulative mortalities reached 100% within 5 to 7 d and were significantly affected by catching and temperature stress. A non-occluded rod-shaped viral particle was found by electron microscopy in the epidermis of both spontaneously and experimentally infected kuruma shrimp. Virions were enveloped, 330 + 20 nm in length and 87
*
7 nm in diameter. The close resemblance in external signs and viral morphology between spontaneously diseased and experimentally infected shrimp indicated that the rod-shaped virus may be the main causative agent of the disease characterized by white spot syndrome in Taiwan.KEY WORDS: Baculovirus . Pathogenicity . Penaeid shrimp. White spots . Taiwan
INTRODUCTION
Since 1992, outbreaks of a new disease leading to
serious mortality among populations of cultured kuruma shrimp Penaeus japonicus have occurred in northern Taiwan. The disease is characterized by obvious white
spots on the carapace, appendages and the inside sur- face of the body, and cumulative mortality reaches 100%
within 2 to 7 d. The diseased shrimp also display signs of lethargy and reddish coloration of the hepatopancreas. In 1993, white spot syndrome (WSS) in cultured giant tiger prawn P monodon and redtail prawn F! penicillatus was observed. Serious damage to penaeid shrimp pro- duction from WSS in Taiwan has been reported (M. C. Tung, J. H. Chen, G . H. Cheng pers. comm.).
'E-mail: [email protected] "Addressees for reprint requests
A n epizootiological survey of kuruma shrimp in Japan reported similar findings (Nakano et al. 1994). According to the evidence from electron microscopy and the results of challenge tests with the filtrate from diseased shrimp lymphoid organs, the causative agent was a virus that was temporarily designated RV-PJ, a rod-shaped nuclear virus of Penaeus japonicus (Inouye et al. 1994, Takahashi e t al. 1994).
To date, the prevalence of baculoviruses in cultured penaeid shrimps has been well documented (Lightner 1983, 1985). Among these penaeid baculoviruses, monodon baculovirus (MBV), baculoviral mid-gut necrosis virus (BMNV), Baculovirus penaei (BP) a n d yellowhead disease virus (YHV) were considered to be the most important because they have on occasion caused serious losses in infected shrimp populations (Couch 1974. Lightner & Redman 1981, Sano et al. 1981. Lightner et al. 1983, 1987, Boonyaratpalin e t O Inter-Research 1995
Dis aquat Org 23: 165-173, 1995
al. 1993). The penaeid shrimps of Southeast Asia are severely threatened by viral infection.
Baculovirus-like viral particles were observed in the penaeid shrimp spontaneously diseased with WSS (Tung et al, pers, comm.). This virus may possibly be the main causative agent for the WSS that has occurred in Taiwan in penaeid shrimp. The present study describes our electron microscope observations of diseased shrimp. A filtrate was made from the epi- dermis of these diseased shrimp a n d the presence of viral particles was confirmed by negative staining. Direct inoculation of this virus-containing filtrate into fish cell lines showed no cytopathic effect. Challenge tests used this filtrate on different sized kuruma shrimp. The effects of catching and temperature stress were also investigated.
MATERIALS AND METHODS
Shrimp. Diseased kuruma shrimp were collected from a culture farm at Wu-Chieh in I-Lan Prefecture in northern Taiwan. The healthy kuruma shrimp used for challenge tests were obtained from a hatchery and a shrimp farm in southern Taiwan where no viral disease had been reported. All of the kuruma shrimp were maintained in aerated 25 to 28°C aquaria and fed on artificial, commercially obtained shrimp food twice daily.
Light microscopy. Both normal kuruma shrimp a n d individuals displaying marked white spot signs were preserved in Davison's fixative (Bell & Lightner 1988). After 48 h in Davison's fixative, specimens were trans- ferred to 5 0 % ethanol, then processed routinely for histology to 5 pm paraffin wax sections, a n d stained routinely with hematoxylin a n d eosin (H&E).
Transmission electron microscopy. Samples of the epidermis covering the gill chamber underneath the carapace were removed from naturally and experi- mentally Infected live kuruma shrimp, and immedi- ately prefixed in 2.5% glutaraldehyde in 0.1 M cold phosphate buffer solution (PBS, pH 7.4) for 2 h at 4OC. Subsequently, samples were washed several times in cold PBS, then postfixed in 1 % osmium tetroxide for 3 h a t 4°C. The samples were dehydrated and embed- ded in Spurr's resin. Ultrathin sections were prepared on a Reichert-Jung Ultracut-E ultratome, and stained with uranyl acetate and lead citrate. The sections were observed with a Hitachi H-600 transmission electron microscope.
Negative staining. The epidermis from infected Penaeus monodon was removed and homogenized in brackish water at 4OC at a ratio of 1:9. After being centrifuged at 8510 X g (Sigma 2K15 rotor 12141) for 5 rnin, the supernatant fluid was filtered through a
0.45 pm membrane. The filtrate was centrifuged at 14 549 X g (Sigma 2K15 rotor 12139) for 1.5 h and the resulting pellet was resuspended in sterilized brackish water prior to negative staining. For negative staining, 1 drop of suspension was mixed with 4 drops of the mixture of 0.1 % bovine serum albumin and 2 % phos- photungstic acid (1 : 2, pH 7.0). The mixture was placed on a grid for 30 to 60 s and excess suspension was removed with filter paper. The preparation was allowed to dry before being examined. The result was observed under a Hitachi H-600 transmission electron microscope.
Cytopathology assay. EPC (epithelioma papulosum cyprini), CHSE-214 (chinook salmon embryo), FHM (fathead minnow) and SSE-5 (sockeye salmon embryo) cells were seeded in 24-well microplates. A filtrate was made from the epidermis of the diseased shrimp and was diluted from 1/20 to U12500 in 5-fold dilutions. Diluted solutions were inoculated into the 4 fish cell lines and these cells were observed over 2 wk at a n incubation temperature of 20°C.
Challenge test I. An infection trial was performed using the filtrate of the epidermis from live or frozen naturally infected Penaeus japonicus and P monodon. The filtrate was diluted 500 to 750 times in brackish water in order to be used as a waterborne inoculum. Two replicates of 35 juvenile kuruma shrimp (1 mo old, mean weight 0.08 g ) were immersed in these diluted filtrates for 2 h. Two other populations were similarly exposed, either to the filtrate from healthy P monodon epidermis or to Grace's insect medium: these served as controls. After inoculation, shrimp were kept in aerated glass aquaria. Water temperature and salinity were 25 to 28°C and 25 to 30 ppt, respectively, through- out the experiment. Mortality was observed daily and moribund shrimp were collected and examined by transmission electron microscopy.
Challenge test 11. Infection trials using 3 different sized groups of juvenile kuruma shrimp proceeded as above. Twenty to thirty juvenile kuruma shrimp (mean weights 0.08, 0.16 and 0.26 g ) were exposed for 1 h to inoculum from diseased Penaeus japonicus prepared a s above and then kept in aerated glass aquaria. Water temperature and salinity were 25 to 28°C and 25 to 30 ppt, respectively, throughout the experiment. Mor- tality was observed daily.
Effect of catching stress o n experimental infection trials. Catching stress was applied to juvenile kuruma shrimp (n = 20, mean weight 0.16 g ) after exposure to brackish water only or to the inoculum from diseased Penaeus japonicus prepared as above. After transfer to glass aquaria, shrimp of experimental and control groups were caught by net and transferred to another glass aquarium once a day. Mortalities were recorded daily.
Effect of temperature stress o n experimental infec- tion trials. Twenty juvenile kuruma shrimp (mean
weight 0.16 g ) were transferred to 33OC after 1 h of exposure at room temperature (25 to 28°C) to the inoculum from diseased Penaeus japonicus prepared as above. A control group was exposed to brackish water at room temperature for 1 h and then transferred to 33°C. The shrimp were observed daily for mortality.
RESULTS
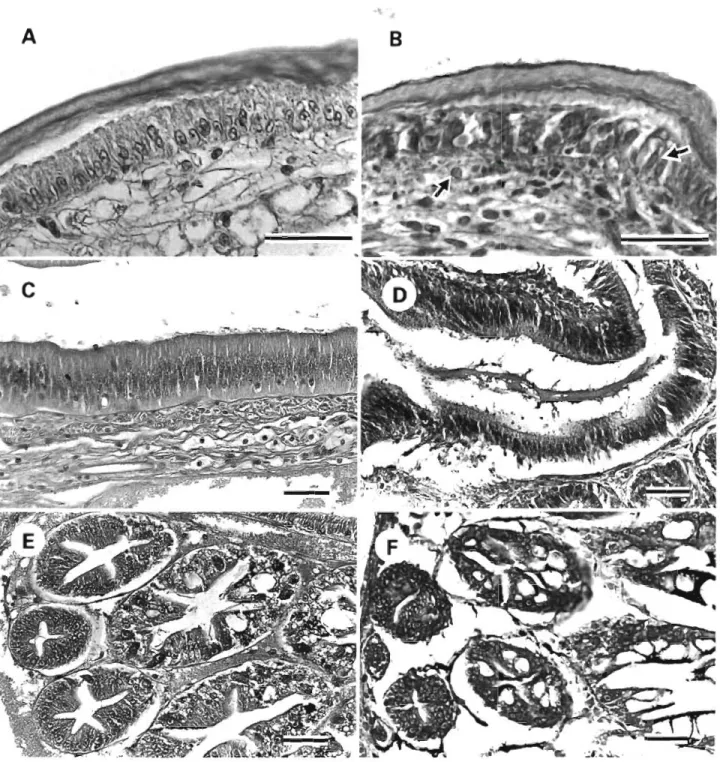
Histopathological changes in tissues from kuruma shrimp naturally infected with white spot syndrome were observed (Fig. 2A to F). Degenerated cells char- acterized by hypertrophied nuclei were observed in the cuticular epidermis a n d connective tissue. Cellular necrosis and detachment of intestinal epithelium tissue were also observed. Fig. 2F shows severe atrophy of the hepatopancreas in these diseased shrimp.
Light microscopy The ultrathin sections of the underlying epidermis of the cuticle from spontaneously diseased and experi- The typical external signs of spontaneously diseased mentally infected kuruma shrimp revealed numerous
Penaeus monodon and l? japonicus in these outbreaks non-occluded, enveloped viral particles with rod-
are the obvious white spots on the carapace, append- shaped morphology (Fig. 3A, B ) . The viral particles ages and the inside surface of the body (Fig. l A , B). were 330 k 20 nm in length and 87 * 7 nm in diameter (n = 30). The electron-dense central
core of the viral particle was nucleo- capsid, approximately 220 X 70 nm in size. Rod-shaped virions as large aggregations in the nuclei of infected eplthelial cells were also examined under transmission electron micro- scopy (Fig. 4 ) . No differences in virion morphology between spontaneously diseased and experimentally infected shrimp were recognized.
Transmission electron microscopy
Negative staining and cytopathology assay
The result of negative staining of the
in Fig. 5. Viral particles with rod-
shaped morphology can be seen. These a r e similar to the viral particles
---
-mQ&
observed in ultrathin sections of
-
spontaneously diseased shrimp. Nopellet from the filtrate of diseased Penaeus monodon epidermis is shown
-
bacteria were observed.TT*=-
Cytopathic effects were not found inany of the 4 tested fish cell lines; the filtrate which was used a s a water- borne inoculum had no cytotoxicity.
Challenge test
Healthy shrimp were exposed to epi-
Fig. 1. External appearance of spontaneously diseased (A) Penaeus monodon dermal diseased
and (B) P japonicus cultured in Taiwan. White spots (arrows) appear on the laponicus a n d l? monodon which ex-
Dis aquat Org 23: 165-173, 1995
Fig. 2. Penaeus japonicus. Histopathological changes in the tissues of kuruma shrimp naturally Infected with white spot syn- drome. ( A , B) Cuticular epidermis of normal and diseased shrimp. Degenerated cells characterized by hypertrophied nuclei (arrows) were observed in the cuticular epidermis and connective tissue. (C) Normal intestinal tissue compared with (D) cellular necrosis and detachment of Intestinal epithelium tissue in d~seased shrimp. (E, F) Hepatopancreas of normal and diseased shrimp.
Severe atrophy of the hepatopancreas in the diseased shrimp was observed. H&E. Scale bars = 40 pm
These experimentally infected shrimp resembled the pathogenic to the smallest shrimp tested (mean weight spontaneously affected ones (Fig 6), a n d cumulative 0.08 g ) ; all of these shrimp died within 5 d . Only 35% mortalities reached 100% within 5 to 7 d (Fig. 7 ) , while cumulative mortality was found in the 0.16 g shrimp no shrimp died in the control groups. As Fig. 8 shows, group after 7 d , although mortality reached 100% in the inoculum from diseased P japonicus was highly 12 d. 10% mortality was observed in the group of
Fig. 3. Penaeus japonicus. Epidermis of shrimp spontaneously diseased with white spot syndrome. (A) Observed aggregation of viral particles in the nucleus and in the cytoplasm. Scale bar = 2 pm. (B) High magnification of viral particles with rod-shaped
Dis aquat Org 23: 165-173, 1995
Fig. 4 . Penae~rs japonicus. Rod-shaped viral particles in the epidermis of shrimp experimentally infected with the filtrate obtained from diseased P monodon. Scale bar = 200 nm
Fig. 5. Penaeus monodon. Negatively stained micrograph of the pellet from filtrate of diseased giant tiger s h r ~ m p epidermis Viral particles (arrows) with rod-shaped morphology were observed. Scale bar = 500 nm
Fig. 6. Penaeus japonjcus. White spot (arrow) on the removed carapace of experimentally infected shrimp
largest shrimp (mean weight 0.26 g ) within 2 wk. No shrimp died in the control groups. Accordingly, the 0.16 g shrimp were chosen to estimate the effect of stress.
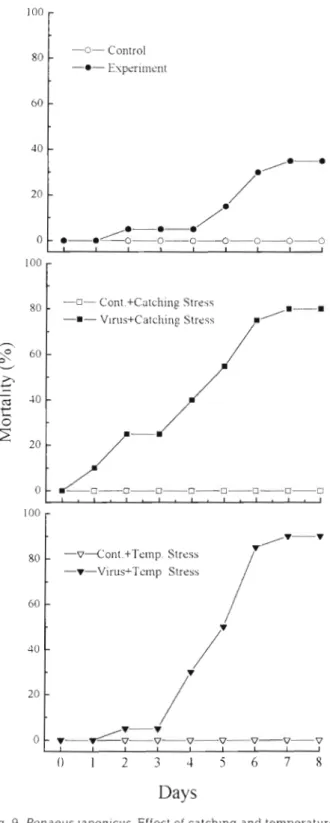
Effect of stress on experimental infection trials T h e results presented in Fig 9 show the effect of catching a n d temperature stress on experimental infection trials. The shrimp in the experimental group under catching stress began to die 1 d after inocula-
Days
Fig. 7. Penaeus japonicus. Cumulative mortalities ('%) of shrimp (0.08 g average weight) experimentally infected by immersion in filtrates from diseased P japonicus and J?
monodon contrasted with healthy shrimp control groups
0 2 4 6 8 10 12 14
Days
Fig. 8. Penaeus japonicus. Cumulative mortalities ("h) of dif- ferent sized groups of juvenile shrimp (0.08, 0.16 and 0.26 g average weight) experimentally infected by immersion in
filtrates from diseased P japonicus
tion, a n d cumulative mortality reached 80 O/o within 7 d . Mortality among the experimentally Infected shrimp under temperature stress reached 90% within 7 d . Only 3 5 ' h mortality was observed in the experimen- tally ~ n f e c t e d shrimp without stress. However, no shrimp died in t h e control groups, whether stressed or not.
DISCUSSION
Disease outbreaks among cultured penaeid shrimp d u e to white spot syndrome have been observed in Taiwan slnce early 1992. The diseased shrimp have obvious white spots on the carapace, appendages a n d the inside surface of the body, and also display signs of lethargy a n d reddish coloration of the hepatopancreas. Thus, in this paper, w e propose the n a m e 'white spot syndrome' (WSS).
Vibriosis, viral infection, poor environmental m a n - a g e m e n t a n d nutrient imbalance h a v e all b e e n conjec- tured to b e the possible cause for these outbreaks (S. N. Chen, S. L. Huang, J . S. Uang, J S. Wang, W. Y Liou & C . H. Kou pers. comm.). Based on electron microscope observation, however, a rod-shaped virus was considered to b e the main causative a g e n t . In the present study, the pathogenicity of a virus from dis- eased Penaeus japonicus a n d l? monodon with white spot syndrome w a s investigated. Close resemblance of white spot signs a n d viral morphology between spontaneously diseased a n d experimentally infected shrimp demonstrated that this virus is indeed the causative agent of the outbreak. T h e virus is highly
Dis aquat Org 23: 165-173, 1995 -0- Control -0- Experiment -0- Cont.+Catching Stre.5~ -m- Vlrus+Catching Stress 100 -v--Cont.+Temp. Stress -V-Vims+Tcrnp Stress
Days
Fig. 9. Penaeuslaponicus. Effect of catching and temperature stress on cumulative mortalities (X) of juvenile shrimp (0.16 g average weight) experimentally infected by immersion in
filtrates from diseased P japonicus
pathogenic a n d constitutes a threat to shrimp. Informal pilot studies in which diseased shrimp w e r e fed to healthy specimens s.uggest that the virus may b e trans- mitted orally a s well a s via water
In J a p a n , a serious disease causing white spots on the body of cultured kuruma shrimp continues to b e re- ported. Serious damage to kuruma shrimp production has occurred a n d many shrimp farms in western J a p a n have temporarily been forced to close (Nakano et al. 1994). Moreover, the causative agent was considered to b e a virus that was shipped with imported kuruma shrimp from China (Momoyama e t al. 1994). Outbreaks of WSS amongst penaeid s h ~ i m p are evidently not a con- fined, local problem anymore. It has already brought the cultured shrimp industry in Asia to a critical condition. In order to classify the causative virus more clearly a n d develop a quick diagnostic method, data on the physicochemical characterization of this agent a r e necessary. Little data on the characteristics of crus- tacean baculoviruses exist. BP a n d MBV a r e t h e only 2 crustacean baculoviruses that have been characterized (Summers 1977, M a n e t al. 1993). The absence of marine crustacean cell lines a n d problems in t h e puri- fication of viruses a r e barriers to this study. In t h e pre- sent paper, w e attemped to m a k e a pellet of the agent from homogenate of infected shrimp a n d preliminary success was shown by the negatively stained virions from the products. Further studies need to b e per- formed to purify the a g e n t more effectively a n d to increase virus yield from the purification technique, a s well a s to determine t h e DNA structure a n d other specific characteristics of this virus. Furthermore, comparisons of the causative rod-shaped virus with other shrimp baculoviruses a r e required to establish whether the WSS-associated virus is really a n e w agent, or a mutation of a n already known virus.
Acknowledgements. The authors express their appreciation to Mr W. B. Yu and Mr W. T Hong, I-Lan Livestock Disease Control Center, for sampling assistance.
LITERATURE CITED
Bell TA, Lightner DV (1988) A handbook of normal penaeid shrimp histology. World Aquaculture Society, Baton Rouge
Boonyaratpalln S, Supamataya K, Kasonchandra J, D~rek- busaracom S, Ekpanithanpong U, Chantanachooklin C (1993) Non-occluded baculo-like virus, the causative agent of yellow head disease in the black tiger shrimp (Penaeus monodon). Fish Pathol 28: 103- 109
Couch JA (1974) An enzootic nuclear polyhedrosis virus of pink shnmp: ultrastructure, prevalence, and enhance- ment. J Invertebr Pathol 24:311-331
lnouye K, Miwa S, Oseko N , Nakano H, Kimura T (1994) Mass mortalities of cultured kuruma shrimp, Penaeus japonicus, In Japan in 1993 electron microscopic evidence of the causative virus. Fish Pathol 29:149-158 (in Japanese) Lightner DV (1983) Diseases of cultured penaeid shrimp. In:
McVey JP (ed) Mariculture. CRC Press, Boca Raton, p 289-320
Lightner DV (1985) A review of the diseases of cultured penaeid shrimps and prawns with emphasis on recent discoveries and developments. In: Taki Y e t a1 (eds) Pro- ceedlngs of the First International Conference on the Cul- ture of Penaeid Prawns/Shrimps. SEAFDEC Aquaculture Department, Manila, p 79-103
Lightner DV, Hedrick RP. Fryer JL, Chen SN. Liao IC, Kou GH (1987) A survey of cultured penaeid shrimp in Taiwan for viral a n d other important diseases. Fish Pathol 22:127-140 Lightner DV, Redman RM (1981) A baculovirus-caused dis-
ease of the penaeid shrimp, Penaeus monodon. J lnvertebr Path01 38:299-302
L ~ g h t n e r DV, Redman K M , Bell TA (1983) Observations on the geographic distribution, pathogenesis and morphology of the baculovirus from Penaeus monodon Fabncius. Aqua- culture 32:209-233
Mari J , Bonami JR, Poulos B, Lightner D (1993) Preliminary characterization and partial cloning of the genome of a baculovirus from Penaeus monodon (PmSNPV = MBV). Dis aquat Org 16:207-215
Responsible Subject Edltor: J. E. Stewart, Dartmouth, Nova Scotia, Canada
Momoyama K, Hiraoka M , Nakano H, Koube H, lnouye K, Oseka N (1994) Mass mortalities of cultured kuruma shrimp, Penaeus japonicus, in J a p a n in 1993: histopatho- logical study. Fish Pathol 29:141-148 (in Japanese) Nakano H, Koube H, Umezawa S, Mornoyama K , Hiraoka M ,
Inouye K, Oseko N (1994) Mass rnortalities of cultured k u r u n ~ a shrimp, Penaeus japonicus. in J a p a n in 1993: epizootiological survey and infection trials. Fish Pathol 29:135-139 (in Japanese)
Sano T, Nishimura T. Oguma K, Momoyama K, Takeno N (1981) Baculovirus infection of cultured kuruma shrimp Penaeus japonicus In J a p a n . Fish Pathol 15.185-191 Summers MD (1977) Characterization of s h r ~ m p baculovirus.
Ecological Research Series 600/3-77-130, US Environmen- tal Protection Agency. Washington, DC
Takahashi Y, Itami T. Kondo M , Maeda M. Fujii R, Tomon- a g a S, Supamattaya K, Boonyaratpalin S (1994) Electron microscopic evidence of bacilliform virus infection in kuruma shrimp (Penaeus japonicus). Fish Pathol 29: 121-125
h4anuscript first received: February 17, 1995 Revised version accepted: M a y 30, 1995