Peritoneal Dialysate Effluent during Peritonitis Induces Human Cardiomyocyte
Apoptosis by Regulating the Expression of GATA-4 and Bcl-2 Families
Hsin-Hui Wang1,2*, Ping-Chun Li3*, Hsiao-Ju Huang4, Tzong-Yann Lee5,6, Ching-Yuang Lin4,7
1
Department of Pediatrics, Section of Nephrology, Taipei Veterans General Hospital,2Department of Pediatrics, Faculty of Medicine, National Yang-Ming University, Taipei,
3
Department of Surgery, Division of Cardiovascular Surgery, China Medical University Hospital,
4
Clinical Immunological Center, China Medical University Hospital,
5
Department of Internal Medicine, Section of Nephrology, En Chu Kong Hospital,
6
Department of Integrated Diagnostics & Therapeutics, National Taiwan University Hospital,
7
College of Medicine, China Medical University *Contributed equally to this work.
Running title: PD effluent induces cardiomyocyte apoptosis
Address for correspondence and reprint requests
Ching-Yuang Lin, M.D., Ph.D.
Clinical Immunological Center, and Division of Pediatrics Nephrology, China Medical University and Hospital
No. 2, Yuh-Der Road, Taichung, Taiwan 40402 TEL: 886-4-2207 1501
FAX: 886-4-2207 1352
ABSTRACT
Cardiovascular event and infection are leading causes of death from peritoneal dialysis (PD). This
study examined in vitro cellular mechanism for cardiotoxicity induced by PD-related peritonitis.
Cultured human cardiomyocytes were treated with PD effluent (PDE) during peritonitis (PPDE),
and effects of PPDE on cultured cardiomyocytes in terms of apoptosis, with expression its related
genes assessed. Results showed PPDE treatment of cardiomyocyte leading to onset of apoptosis, as
confirmed by phosphatidylserine exposure plus DNA fragmentation and damage. This apoptosis is
mediated by reduced Bcl-2/Bax and Bcl-xL/Bax ratios, as well as reduced expression of GATA-4,
an important cardiomyocyte survival factor, at the level of transcription. These changes activated
pro-apoptotic pathways. PPDE treatment also inhibited ERK signals, contributing to cardiotoxicity.
Our findings revealed that PPDE contains potent pro-apoptotic factors that regulate expression of
GATA-4 and Bcl-2 families, inducing cultured cardiomyocyte apoptosis. This pinpoints a key role
of apoptosis in PD-associated cardiovascular events, along with a potential therapeutic target.
INTRODUCTION
Cardiovascular event and infection are the first and second leading causes of death in the
peritoneal dialysis (PD) populations [1-4]; both events are closely related. PD-related peritonitis is
the crucial infection in PD patients [5, 7]. Peritoneal toxin should be absorbed to the systemic
circulation and might induce cardiotoxicity. After an episode of severe infection in dialysis patients,
risk of death from cardiovascular events is increased seven-fold for 6 months and continues to rise
for up to 48 months [5, 6]. It has been considered to play a significant role in up to one sixth of
patient deaths occurring during the course of PD therapy [8]. In 41.5% of patients with
peritonitis-related mortality, immediate cause of death was a cardiovascular event [9]. Clinical
findings indicate that a peritonitis episode may culminate in cardiovascular event [5, 8]: high
incidence of peritonitis is accompanied by greater risk of death [8, 10, 11], and cardiovascular
events contribute to risk of peritonitis-related death in patients undergoing PD [12-14]. However,
the possible mechanisms connecting PD-related peritonitis and cardiac mortality have not been
addressed.
Growing evidence implicates cardiomyocyte apoptosis as a mechanism contributing to various
types of heart disease [15-17]. Cardiomyocyte apoptosis could result in a loss of contractile tissue,
compensatory hypertrophy of myocardial cells, reparative fibrosis, and heart failure. In animal
models, endotoxin [18, 19], exotoxin [18, 20], and inflammatory mediator [21] play important roles
mediators and cytokines increase in PD effluent (PDE) and correlate with treatment outcome
[22-23]. Yet there are no data on effects of peritonitis PD effluent (PPDE) on cardiomyocytes
viability and apoptosis.
Bcl-2 protein family members are the best characterized proteins that are directly involved in
the regulation of apoptosis [24]. Bcl-2 and its closest homologues, Bcl-xL and Bcl-w, potently
inhibit apoptosis in response to many cytotoxic insults. Bax and Bak are well known proapoptotic
members of the Bcl-2 protein family. Regulation of apoptosis is highly dependent on the ratio of
anti-apoptotic to pro-apoptotic proteins. Conditions that induce myocardial stress cause complex
alterations in levels of Bcl-2 family proteins [25].
Cardiac Bcl-2 gene expression has been shown to be regulated by GATA-4 both in vitro and in
vivo [26]. GATA-4 is a transcription factor enriched in cardiac tissue that is essential for various
cardiomyocyte physiological and adaptive responses. An early event in the cardiotoxicity induced
by the antitumor drug doxorubicin is GATA-4 depletion, which in turn causes cardiomyocyte
apoptosis [27, 28]. GATA-4 has also been shown to upregulate transcription of the anti-apoptotic
genes Bcl-2 [26] and Bcl-xL[27, 28] in cardiomyocytes, and to play a central role in regulating the
survival or apoptosis of cardiomyocytes. Although previous studies have suggested the importance
of apoptosis regulation and GATA-4 expression in various heart diseases, their role in PD
peritonitis-related cardiotoxicity has not been elucidated.
To clarify the relationship between PD-related peritonitis and high cardiac mortality, we
PD-related peritonitis, proapoptotic pathways are activated in cardiomyocytes. To test this
hypothesis, human cardiomyocytes were cultured and treated with PPDE. The possible underlying
SUBJECTS AND METHODS
Human cardiomyocytes culture
This research was approved by the China Medical Hospital Institutional Revien Board. Written
informed consent was obtained from each individual. Human cardiomyocytes obtained from the
myocardial ventricular resection specimens of patients undergoing cardiac surgery were isolated as
previously described [29]. Cells were cultured for a period of eight days, and culture medium was completely replaced every threedays.Cultured medium wasDulbecco’smodified Eaglemedium (Hyclone, UT, US) containing equal Volume of F12, 1% L-glutamine, 25% fetal bovain serum, 1%
non- essential amimo acid, 1% pyruvate, 250μl hydrocortisone, 250μl interferrin and insulin 250 μl in each 500ml. For keeping its cardiomyocytes property, the cells were passaged before 90 percent confluent about 2-3 days.
Doxorubicin (Sigma, MO, US) (DOXO) was prepared as 50μM stock solution in DMSO and then
diluted to 0.5 and 1μM in 1×PBS for working concentration.
We also measured electrophysiological character of cultured human cardiomyocytes, including
action potential duration (APD) and peak L-Type calcium current (IcaL) [30].
Immunostaining of human cardiomyocytes and confocal microscopic imaging
To characterize cardiomyocytes in culture, cells were incubated with a desmin monoclonal antibody
polyclonal anti-CAPON antibody (Santa Cruz Biotechnology, Inc. CA, USA), followed by a
FITC-labelled IgG (Santa Cruz Biotechnology, Inc.) secondary antibody. Cells were double labeled
with WGA (Invitrogen, San Francisco, USA). Staining was assessed and photomicrographs were
obtained using a Leica TCS SP2 Confocal Spectral Microscope.
Patient population and peritonitis
Turbid PPDE prior to antibiotic treatment were collected in 8 culture positive peritonitis episodes
from PD patients treated in our hospital. Microorganisms were gram-positive bacteria in 5 episodes
and gram-negative bacteria in 3 episodes. All patients had a Tenckhoff peritoneal catheter inserted
and were treated with the standard double-bag system (Baxter Healthcare Corp., Deerfield, IL).
These 5 men and 3 women had a mean age of 47.8 years. Causes of renal failure included chronic
glomerulonephritis, reflux nephropathy, obstructive nephropathy and renal hypoplasia. Time on PD
ranged from 10 to 56 months. Peritonitis was defined as presence of two of the following criteria:
microorganisms on gram staining, subsequent positive culture of PD fluid; cloudy fluid (leukocyte
count >100 cells/mL with >50% polymorphonuclear cells) and/or peritoneal inflammation
symptoms. Exclusion criteria included: a) tunnel-tract or exit site infections, b) completion of
antibiotic therapy for peritonitis within 28 days of study enrollment, c) the presence of peritonitis
attributed to fungus or mycobacterial infection or negative culture, d) drug sensitivity showing
resistance to initial antibiotic therapy, e) previous immunosuppressive therapy, f) anemia from a
Overnight dwell bags of PDE samples (SPDE) were collected also from 5 stable PD without
peritonitis patients (2 men, 3 women, mean age = 50.23 ± 11.67 years) treated in our hospital. No
patient was diabetic, and no peritonitis episode was noted over the past year. Causes of renal failure
included chronic glomerulonephritis, obstructive nephropathy andpolycystic kidney disease. Patients
signed an informed consent form for these studies.
Collection of PDE
The PDE were collected as previously described [31]: PDE was centrifuged at 400 g, 4C for 10 min. After centrifugation, supernatants were collected, refrigerated, then lyophilized and dried to powder using Freeze Dryer (Eyela, Tokyo, Japan). Powder was stored at -70C until analysis. The lyophilized PPDE samples were reconstituted as a 625 mg/ml stock solution with 1×PBS, and
filtrated by 0.45 μM filter. PPDE stock solutions were storage at o
C
20
and diluted with 1× PBS to working concentration before use.MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay
Cardiomyocyte cell viability was determined by a MTT assay. In a 96-well microplate, 2.5x104
cells/well were incubated in 100 l of culture media and exposed to different concentrations of DOXO or PPDE for varying time periods. 12.5μM MTT dye (stock ; 5 mg/ml) were added into
each well and the cells were further incubated at o
C
37
for 4hr. Viable cells could convert MTT dye dark blue product, the cells were lysed with dimethyl sulfoxide (DMSO) (Sigma, MO, US) andusing a microplate reader (mode FL 331, Bio-Tek Instruments, Winooski, VT).
Apoptosis assays by flow cytometry
Apoptosis was quantitatively gauged by detecting phosphatidylserine exposure on cell membrane
with Annexin V staining, as described above [32]. Cells were simultaneously stained with Annexin
V-FITC (25 ng/ml; green fluorescence, R&D Systems, Minneapolis, MN, USA) and dye exclusion
(propidium iodide, 20μg/ ml, red fluorescence). Data were obtained by flow cytometry analysis
with FACS-SCAN(Becton-Dickinson) FACS Canto in cell populations from which debris was
gated out and analyzed.
Evaluation of apoptosis by TUNEL and DAPI staining
Cultured human cardiomyocytes were plated at approximately 2 x105 cells/well in 12-well plates with DOXO or PPDE and incubated at 37˚C for 24 h. Cell nuclei were stained with 4, 6-Diamidino-2-phenyindole (DAPI, Sigma) and DNA fragments labeled with
AlexaFluor 488 dye-labeled anti-BrdU antibody (Apo-Brd UTM TUNEL assay kit. Invitrogen).
Cells were observed and photomicrographs were obtained using a Leica TCS SP2 Confocal
Spectral Microscope. An individual blinded to the experimental conditions counted at least 300
cells in 6 different high power fields for each experiment.
Comet assay
peritonitis for 24 h at 37˚C, isolated, then examined for DNA damage by Comet assay previously
described [33]. Treated cells were embedded in situ in 1% agarose, then placed in lysis solution for
30 min. Cell nuclei were subsequently electrophoresed for 20 min at 1 V/cm, followed by staining
with PI and visualization with a fluorescence microscope.
RNA isolation and reverse transcription
RNA was extracted from cells using RNAzol B (TEL-TEST, INC.). Resulting RNA suspension was forthwith converted to cDNA by reverse transcription, cDNA samples stored at -70C for analysis.
Real-time PCR with SYBR green assay
Five 贡l of cDNA (1-10 ng) was mixed with SYBR green PCR core reagent or master mix reagent
(Aplied Biosystems, CA, US). The thermal cycling conditions were determined according to rules of‘Thermalcycling parametersforprimeroptimization’.Each RNA sampleswasalso analyzed with β-actine which serve as internal control for correcting relative specific gene expression levels.
Primers were designed using Primer Express Primer Design software, as follows:
Bcl-2: sense primer: ATGTGTGTGGAGAGCGTCAA
antisense primer: ATCACCAAGTGCACCTACCC
Bcl-xL: sense primer: ACAGCAGCAGTTTGGATGC
Bax: sense primer: TTGGGTGAGACTCCTCAAGC
antisense primer: CACTGTGACCTGCTCCAGAA
GATA-4: sense primer: AGCTCCTTCAGGCAGTGAGA
antisense primer: CTGTGCCCGTAGTGAGATGA β-actine : sense primer : CAGGTATGCACCCAGAGTGA
antisense : GATATGGAGAAGATTTGGCA
According to amplification plot, cycle number over the threshold means Ct value. The Ct value of
non-template control is 45. The relative expression ratio among untreated RNA and different RNA
samples could be calculated with 2-Ct.
Western blotting
10-50μg protein extracts were separated by 10-12% SDS-PAGE and transferred to nitrocellulose
(PVDF) membranes blocked overnight with 1×TBS buffer containing 5% skim milk. Membranes
were incubated with optimal concentrations of primary antibodies: anti-GATA-4 mAb (Abcam,
Abgent, San Diego, CA, USA), anti-extracellular signal–regulated kinase (ERK) mAb (Cell Signaling Technology, Beverly, MA, USA), and anti-actin mAb (Sigma) in 1×TBS buffer containing 5% skim milk. Membranes were washed and then incubated with appropriate secondary
antibody (goat anti-mouse mAb conjugated with HRP), and visualized by enhanced
chemiluminescence ECL detection kit (Perkin Elmer, MA, US). The antibody for GATA-4 (1:100),
purchase from cell signaling (CA, US).
Statistics
All data are presented as mean ± standard deviation (SD). Differences between groups were
RESULTS
Characterization of human cardiomyocytes in primary culture
To characterize cardiomyocytes, muscle markers desmin and myocyte-specific protein α
-sarcomeric actinin were detected (Figure 1). CAPON, recently documented as endogenous protein
expressed in guinea pig cardiomyocytes, interacts with nitric oxide synthase to accelerate cardiac
repolarization by inhibition of L-type calcium channels. Expression of endogenous CAPON protein
in cultured cardiomyocytes was detected by immunofluorescent staining and confocal microscopy
(Figure 1). Both action potential duration (APD) and peak L-Type calcium current (IcaL) were
APD10, APD50, APD75 and APD90 : 95.4±10.6, 289.2±15.6, 308.2±15.4, and 318.4±16.4 ms,
respectively, with peak IcaL density of -10.2±0.9 pA/pE at + 10mV (n=6).
PPDE induces cell death in human cardiomyocytes
Cardiac cell death is believed to play a major contributory role in development and progression of
myocardial dysfunction [17]. To assess whether PPDE treatment induced cardiac cell death, cell
viability were evaluated by MTT assay. Doxorubicin-induced cardiotoxicity, which has been well
described [35], was used as a positive control. MTT assay showed PDE during peritonitis- and
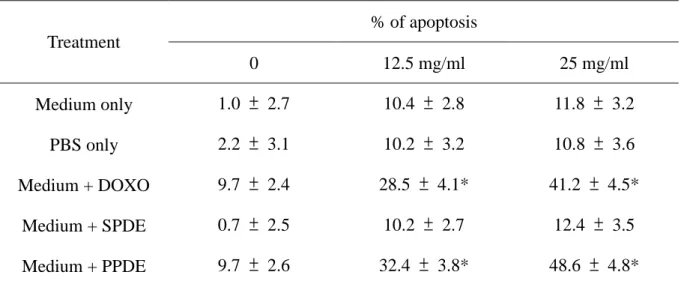
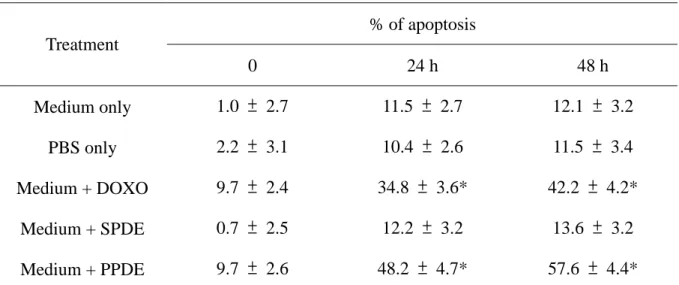
doxorubicin-induced human cardiomyocyte cell death as both dose- (Figure 2A and Table 1) and
time-dependent (Figure 2B and Table. 2). When cardiomyocytes were pre-exposed to 12.5, 18, or 25 mg/ml PDE during peritonitis for 24 hr, cell viabilities were 70.65.7%, 58.79.7%, and
41.67.8%, respectively, all significantly lower than in cardiomyocytes without pre-treatment (P<0.05) (Figure 2A). This change was even more profound in the 48 hr treatment group (Figure
2B). When cardiomyocytes were pre-exposed to 25 mg/ml PDE from stable PD patients for 24 and
48 hr, cell viabilities were similar with cardiomyocytes without pre-treatment (data not shown).
PPDE induces apoptosis in human cardiomyocytes
The above lend substantial evidence of apoptosis playing a critical role in cardiomyocyte cell
death associated with several cardiac diseases [15, 17]. To explore whether PPDE during peritonitis
challenge induces human cardiomyocyte apoptosis, we assessed apoptotic cell death by flow
cytometry. TUNEL staining and Comet assays were performed for determination of DNA damage.
Doxorubicin, which can induce cardiomyocyte apopotosis [33], was used as a positive control. After
cell incubation with 25 mg/ml PPDE peritonitis for 24 hr, apoptosis was detected by flow cytometry
(Tables 1 and 2). TUNEL staining and confocal microscopy confirmed cardiomyocyte apoptosis
(Figure 3A). Analyses indicated little cardiomyocyte apoptosis with non-exposed condition (control
group) and exposure with 25 mg/ml PPDE from stable PD patients (Tables 1 and 2). By contrast,
doxorubicin and PPDE induced apoptosis in 32.3-49.7% of human cardiomyocytes after treated for
24 hours. Finally, PPDE induced DNA damage was determined by Comet assay (Figure 3B): higher
concentrations of PPDE resulted in greater numbers of damaged cells.
The Bcl-2 family of proteins are key regulators of the stress-induced apoptotic pathway [25]; to
determine their role in regulation of PPDE induced cardiomyocyte apoptosis, mRNA concentrations
of prosurvival proteins Bcl-2 and Bcl-xL and proapoptotic protein Bax were measured in human
cardiomyocytes by quantitative real-time RT-PCR(Figure 4A). In cardiomyocytes exposed to PPDE
4 hrs, the Bcl-2/Bax and Bcl-xL/Bax ratios were 0.300.02 and 0.44 0.02, respectively (Figure
4B). Compared to the no-exposure control group, Bcl-2/Bax and Bcl-xL/Bax ratios dropped by
30-45% following 4 hrs of PPDE treatment (Figure 4A; P<0.05 versus control). Western blotting
analysis for Bcl-2, Bcl-xl and Bax protein expression in the same experimental conditions obtained
similar results (Figure 6A). These data indicated that PPDE treatment decreased Bcl-2/Bax and
Bcl-xL/Bax ratios, resulting in increase Bax expression in human cardiomyocytes.
Transcription factor GATA-4 has been identified as a specific myocardial survival factor which
induces transcription and expression of Bcl-2 and which is associated with cell survival [27-29, 36].
To characterize mechanisms underlying PPDE activity in human cardiomyocytes, mRNA and
protein expression of GATA-4 were measured. For cardiomyocytes exposed to PPDE, GATA-4
mRNA expression decreased fivefold relative to no-exposure control cells by quantitative real-time
RT-PCR (P<0.05) (Figure 5A). Western blots of nuclear GATA-4 protein expression in PPDE
exposed human cardiomyocytes also showed lower levels than the control group (Figure 5B, C),
suggesting that PDE during peritonitis treatment decreases levels of GATA-4 gene expression in
PPDE does not contain inflammatory mediators
To evaluate whether PPDE was enriched in pro-apoptotic mediators, TRAEL, FasL, TNFα, IL-6
and IL-1 were rated by enzyme-linked immunoassay by comercial ELISA kit. Cultured supernatant
from peripheral blood mononuclear cells stimulated with lippolysaccharide was used as positive
control; TRAEL, FasL, TNFα and IL-1 were undetectable in 25mg/ml PPDE (data not shown).
The lower limit of sensitivity was 0.70 pg/ml.
Role of ERK pathway in PPDE induced cardiotoxicity
We next examined possible signaling mechanisms regulating PPDE-induced cardiomyocyte
apoptosis. The GATA-4 molecule contains putative ERK phosphorylation sites, and recent studies
have shown that some survival factors [36-38] induce activity of GATA-4 via MEK/ERK-dependent
phosphorylation. Therefore, we explored activity of MEK/ERK signaling pathways in PPDE treated
cardiomyocytes. Figure 6 shows ERK phosphorylation significantly reduced in cells exposed to
PPDE peritonitis, suggesting that PPDE inhibits the ERK signaling pathway, consistent with the
DISCUSSION
Our study demonstrates that PPDE contains potent pro-apoptotic factors and causes an
imbalance between proapoptotic and prosurvival pathways, inducing apoptosis in human
cardiomyocytes. This study revealed a possible mechanism of PD-related, peritonitis-induced
cardiotoxicity. These novel findings constitute the first direct evidence linking PD peritonitis and
cardiomyocyte apoptosis. Cardiovascular events are the major cause of death in PD patients with
peritonitis. Our findings demonstrate the central role of apoptosis in PD peritonitis-associated
cardiovascular events, and provide an explanation for the high incidents of cardiovascular events in
PD-related peritonitis.
Cardiomyocyte death is important in the pathogenesis of cardiac disease in end stage renal
disease [3]. Cardiomyocyte death induces LV dilatation with compensatory LV hypertrophy, and
eventually leading to systolic dysfunction. LV hypertrophy appears to be an important, independent,
determinant of survival in patiens with end stage renal diseases [39]. Our study yields direct cellular
evidence of PPDE from PD patients as cardiotoxic. In end-stage renal disease, cardiomyocyte death
may be caused by continual LV overload, decreased large and small coronary vessel perfusion,
hyperparathyroidism, and malnutrition [3]. Our data provide another possible cause of cardiac cell
death in patients undergoing PD.
Recent studies have proven various cardiac pathological states associating with cardiomyocyte
when diverse pathogens induced peritonitis (data not shown). Toxicity to cardiomyocytes was most
profound with exposure to Staphylococcus aureus, Staphylococcus epidermidis, and Streptococcus
viridans. Gram negative pathogens such as Escherichia coli and Proteus vulgaris were less cytotoxic.
Prior animal study shows both endotoxin and exotoxin exposure inducing myocardial depression
[18-20]. Interestingly, pretreatment of animals or cells with lipopolysaccharide (LPS) induced LPS
tolerance and largely reduced inflammation and dysfunction of the rat myocardium [40, 41]. Further
study should delineate effects of pathogen-related toxins on the PPDE-induced cardiotoxicity.
A second possible cause of cardiomyocyte apoptosis is inflammatory or pro-apoptotic mediators
and cytokines in PPDE. Expression of inflammatory mediators and cytokines in PPDE is reported
to increase during acute peritonitis, then return to control levels [22-24]. Inflammatory mediators
and cytokines have been cited as playing a role in cardiomyocyte apoptosis and clinical cardiac
dysfunction [42], with TRAL, FasL, TNFα, or IL-1 undetectable in 20 mg/ml PPDE. Inflammatory
mediators and cytokines in PPDE may not contribute greatly to cardiomyocyte apoptosis.
GATA-4 is a survival factor in terminally differentiated cardiac myocytes [27, 28] and may be
an essential component of adaptive response of the adult heart [28]. Experiments have shown 50%
reduction in GATA-4 levels impairing drug-induced stress responses, yet not interfering with
normal embryonic and postnatal mouse development [28]. Our results indicate a mechanism by
which PPDE inhibits transcription of the GATA-4 gene: lower GATA-4 levels in cardiomyocytes
exposed to PPDE would impair their ability to respond to cardiac work or to stimuli that produce
of GATA-4 depletion may prove effective cardioprotection in PD patients with peritonitis.
The essential role of GATA-4 as a survival factor may be explained, in part, by its function as an
upstream activator of the antiapoptotic Bcl-2 gene family. Previous research [43] has shown that
over expression of Bcl-2 attenuates myocardial apoptosis. In the present study, exposure of
cardiomyocytes to PPDE caused decreases in the Bcl-2/Bax and Bcl-xL/Bax ratios. Moreover, Bcl-2
isamajorregulatorofmitochondrialpermeability and hencepreventsactivation ofthe“intrinsic” mitochondria-dependent apoptosis pathway [44]. The intrinsic apoptotic pathway is activated in
cardiac myocytes in response to various extracellular and intracellular stimuli [44]. Our results
provide evidence that activation of the intrinsic apoptotic pathway may be a possible mechanism for
PPDE induced cardiomyocyte apoptosis.
Several studies have suggested that in a subset of cell types, activation of ERK can protect
against pro-apoptotic stimulants [45]. However, inhibition of apoptosis by ERK depends upon cell
type and stimuli. The present study found that the ERK pathway is involved in protection against
PPDE-induced cultured cardiomyocyte apoptosis. Interestingly, GATA-4 contains putative ERK
phosphorylation sites. Prior studies show HGF and endothelin-1 activating GATA-4 via MEK-ERK
pathway-dependent phosphorylation, with dominant MEK mutant blocking HGF-induced Bcl-xL
expression in cardiac muscle cells [29, 36]. Studying phosphorylation of GATA-4 by ERK pathway
will elucidate the role of MEK/ERK/GATA-4 pathway in PPDE-induced cardiomyocyte apoptosis.
Apoptosis is no simple, linear chain reaction. Recent study found sustained inflammation activating
on possible extrinsic apoptotic pathway involvement and relative importance of intrinsic and
extrinsic pathways in PPDE-induced cardiomyocyte apoptosis. Unlike necrosis, viewed as basically
irreversible, the step-by-step nature of apoptosis suggests existence of reversible checkpoints. Our
results indicate therapeutic regimens that block intrinsic apoptosis signaling pathways presumably
attenuate PPDE-induced cardiotoxicity.
In conclusion, this study demonstrates for the first time that PPDE contains potent pro-apoptotic
factors that regulate expression of GATA-4 and Bcl-2 families, inducing cultured cardiomyocyte
apoptosis. Findings illustrate a pivotal role of apoptosis in PD peritonitis-associated cardiovascular
events, explain high cardiac mortality in PD-related peritonitis, and pinpoint apoptotic events as a
ACKNOWLEDGEMENT
REFERENCES
1. 2008. U.S. Renal Data System, USRDS 2008 Annual Data Report: Atlas of Chronic Kidney
Disease and End-Stage Renal Disease in the United States, National Institutes of Health,
National Institute of Diabetes and Digestive and Kidney Diseases, Bethesda, MD.
2. Go AS, Chertow GM, Fan D, McCulloch CE, Hsu CY. 2004. Chronic kidney disease and the
risks of death, cardiovascular events, and hospitalization. N Engl J Med 351: 1296-1305
3. Parfrey PS, Foley RN. 1999. The clinical epidemiology of cardiac disease in chronic renal
failure. J Am Soc Nephrol 10: 1606-1615
4. Schiffrin EL, Lipman ML, Mann JF. 2007. Chronic kidney disease: effects on the
cardiovascular system. Circulation 116: 85-97
5. Bender FH, Bernardini J, Piraino B. 2006. Prevention of infectious complications in
peritoneal dialysis: best demonstrated practices. Kidney Int Suppl S44-54
6. Ishani A, Collins AJ, Herzog CA, Foley RN. 2005. Septicemia, access and cardiovascular
disease in dialysis patients: the USRDS Wave 2 study. Kidney Int 68: 311-318
7. Aslam N, Bernardini J, Fried L, Burr R, Piraino B. 2006. Comparison of infectious
complications between incident hemodialysis and peritoneal dialysis patients. Clin J Am Soc
Nephrol 1: 1226-1233
8. Fried LF, Bernardini J, Johnston JR, Piraino B. 1996. Peritonitis influences mortality in
9. Perez Fontan M, Rodriguez-Carmona A, Garcia-Naveiro R, et al. 2005. Peritonitis-related
mortality in patients undergoing chronic peritoneal dialysis. Perit Dial Int 25: 274-284
10. Maiorca R, Cancarini GC, Brunori G, Camerini C, Manili L. 1993. Morbidity and mortality
of CAPD and hemodialysis. Kidney Int Suppl 40: S4-15
11. Piraino B. 1998. Peritonitis as a complication of peritoneal dialysis. J Am Soc Nephrol 9:
1956-1964
12. Digenis GE, Abraham G, Savin E, et al. 1990. Peritonitis-related deaths in continuous
ambulatory peritoneal dialysis (CAPD) patients. Perit Dial Int 10: 45-47
13. Firanek CA, Vonesh EF, Korbet SM. 1991. Patient and technique survival among an urban
population of peritoneal dialysis patients: an 8-year experience. Am J Kidney Dis 18: 91-96
14. Lupo A, Tarchini R, Carcarini G, et al. 1994. Long-term outcome in continuous ambulatory
peritoneal dialysis: a 10-year-survey by the Italian Cooperative Peritoneal Dialysis Study
Group. Am J Kidney Dis 24: 826-837
15. Olivetti G, Abbi R, Quaini F, et al. 1997. Apoptosis in the failing human heart. N Engl J Med
336: 1131-1141
16. Narula J, Pandey P, Arbustini E, et al. 1999. Apoptosis in heart failure: release of
cytochrome c from mitochondria and activation of caspase-3 in human cardiomyopathy.
Proc Natl Acad Sci U S A 96: 8144-8149
17. Haunstetter A, Izumo S. 1998. Apoptosis: basic mechanisms and implications for
18. Natanson C, Danner RL, Elin RJ, et al. 1989. Role of endotoxemia in cardiovascular
dysfunction and mortality. Escherichia coli and Staphylococcus aureus challenges in a
canine model of human septic shock. J Clin Invest 83: 243-251
19. Ramana KV, Willis MS, White MD, et al. 2006. Endotoxin-induced cardiomyopathy and
systemic inflammation in mice is prevented by aldose reductase inhibition. Circulation 114:
1838-1846
20. Sibelius U, Grandel U, Buerke M, et al. 2000. Staphylococcal alpha-toxin provokes
coronary vasoconstriction and loss in myocardial contractility in perfused rat hearts: role of
thromboxane generation. Circulation 101: 78-85
21. Mann DL. 1999. Inflammatory mediators in heart failure: homogeneity through
heterogeneity. Lancet 353: 1812-1813
22. Wang HH, Lin CY. 2005. Interleukin-12 and -18 levels in peritoneal dialysate effluent
correlate with the outcome of peritonitis in patients undergoing peritoneal dialysis:
implications for the Type I/Type II T-cell immune response. Am J Kidney Dis 46: 328-338
23. Lai KN, Lai KB, Lam CW, et al. 2000. Changes of cytokine profiles during peritonitis in
patients on continuous ambulatory peritoneal dialysis. Am J Kidney Dis 35: 644-652
24. Cory S, Adams JM. 2002. The Bcl2 family: regulators of the cellular life-or-death switch.
Nat Rev Cancer 2: 647-656
25. Bishopric NH, Andreka P, Slepak T, Webster KA. 2001. Molecular mechanisms of apoptosis
26. Kobayashi S, Lackey T, Huang Y, et al. 2006. Transcription factor gata4 regulates cardiac
BCL2 gene expression in vitro and in vivo. Faseb J 20: 800-802
27. Aries A, Paradis P, Lefebvre C, Schwartz RJ, Nemer M. 2004. Essential role of GATA-4 in
cell survival and drug-induced cardiotoxicity. Proc Natl Acad Sci U S A 101: 6975-6980
28. Suzuki YJ, Evans T. 2004. Regulation of cardiac myocyte apoptosis by the GATA-4
transcription factor. Life Sci 74: 1829-1838
29. Ancey C, Corbi P, Froger J, et al. 2002. Secretion of IL-6, IL-11 and LIF by human
cardiomyocytes in primary culture. Cytokine 18: 199-205
30. Kuan-Cheng Chang, Andreas S. Barth, Tetsuo Sasano, Eddy Kizana, Yuji Kashiwakura,
Yiqiang Zhang, D. Brian Foster, and Eduardo Marbán. 2008. CAPON modulates cardiac
repolarization via neuronal nitric oxide synthase signaling in the heart. PNAS March 18,
2008 vol. 105 no.11 4477-4482
31. Wang HH, Lee TY, Lin CY. 2008. Integrins mediate adherence and migration of T
lymphocytes on human peritoneal mesothelial cells. Kidney Int 74: 808-816
32. Chen Q, Gong B, Mahmoud-Ahmed AS, et al. 2001. Apo2L/TRAIL and Bcl-2-related
proteins regulate type I interferon-induced apoptosis in multiple myeloma. Blood 98:
2183-2192
33. Kim Y, Ma AG, Kitta K, et al. 2003. Anthracycline-induced suppression of GATA-4
transcription factor: implication in the regulation of cardiac myocyte apoptosis. Mol
34. Chang KC, Barth AS, Sasano T, et al. 2008. CAPON modulates cardiac repolarization via
neuronal nitric oxide synthase signaling in the heart. Proc Natl Acad Sci U S A 105:
4477-4482
35. Shan K, Lincoff AM, Young JB. 1996. Anthracycline-induced cardiotoxicity. Ann Intern
Med 125: 47-58
36. Kitta K, Day RM, Kim Y, et al. 2003. Hepatocyte growth factor induces GATA-4
phosphorylation and cell survival in cardiac muscle cells. J Biol Chem 278: 4705-4712
37. Kitta K, Clement SA, Remeika J, Blumberg JB, Suzuki YJ. 2001. Endothelin-1 induces
phosphorylation of GATA-4 transcription factor in the HL-1 atrial-muscle cell line. Biochem
J 359: 375-380
38. Morimoto T, Hasegawa K, Kaburagi S, et al. 2000. Phosphorylation of GATA-4 is involved
in alpha 1-adrenergic agonist-responsive transcription of the endothelin-1 gene in cardiac
myocytes. J Biol Chem 275: 13721-13726
39. Silberberg JS, Barre PE, Prichard SS, Sniderman AD. 1989. Impact of left ventricular
hypertrophy on survival in end-stage renal disease. Kidney Int 36: 286-290
40. Barroso-Aranda J, Schmid-Schonbein GW, Zweifach BW, Mathison JC. 1991.
Polymorphonuclear neutrophil contribution to induced tolerance to bacterial
lipopolysaccharide. Circ Res 69: 1196-1206
41. Madorin WS, Cepinskas G, Kvietys PR. 2001. Peritonitis induces rat cardiac myocytes to
lipopolysaccharide pretreatment. Crit Care Med 29: 1774-1779
42. Haudek SB, Taffet GE, Schneider MD, Mann DL. 2007. TNF provokes cardiomyocyte
apoptosis and cardiac remodeling through activation of multiple cell death pathways. J Clin
Invest 117: 2692-2701
43. Chen Z, Chua CC, Ho YS, Hamdy RC, Chua BH. 2001. Overexpression of Bcl-2 attenuates
apoptosis and protects against myocardial I/R injury in transgenic mice. Am J Physiol Heart
Circ Physiol 280: H2313-2320
44. Foo RS, Mani K, Kitsis RN. 2005. Death begets failure in the heart. J Clin Invest 115:
565-571
45. Jarpe MB, Widmann C, Knall C, et al. 1998. Anti-apoptotic versus pro-apoptotic signal
FIGURE LEGENDS
Figure 1. Characterization of human cardiomyocytes by immunostaining of cardiomyocyte markers.
Double labeling of cardiomyocytes with WGA (red) and -sarcomeric actinin (green ) (first line)or
desmin (green) (second line) or CAPON (green) (third line). Negative control using cultured human
skeletal muscle cells was stained with CAPON (last line).
Figure 2. Cell viability reduced after treatment with PPDE. Human cardiomyocytes were treated
with PPDE during peritonitis. Cell viability was determined by MTT assay. (A) Cells were
incubated with various concentration of peritoneal dialysate effluent during peritonitis (PPDE) (12,
5, 18, 25 mg/ml) (n=8) or doxorubicin (0.5, 1M; Doxo) as a positive control for 24 hr. () treated
with medium only () or Doxo pretreatment; (B) cell were treated with PPDE (25 ng/ml) for 24 or
48 hrs. (n=8) and controls Data are expressed as meanSD of 8 different PPDE. # P<0.01 versus
medium only control, * P<0.05, ** P<0.01.
( : treated with PBS only) ( : treated with medium and stable peritoneal dialysate effluent
(SPDE) without peritonitis) ( treated with medium and peritonitis peritoneal dialysate effluent (PPDE))
Figure 3. PPDE treatment induces apoptosis in cultured human cardiomyocytes. (A) APO-Brud
as a positive control for 24 h, then stained by the TUNEL method. Nuclei were counter stained with
AlexaFluor 488dye. Confocal microscopy of cardiomyocytes showed all nuclei (blue), including
apoptotic nuclei (vacuoles). (B) Cardiomyocyte DNA damage was determined by the Comet assay.
Cardiomyocytes were treated with PPDE (12.5 or 25 mg/ml) or Doxo (0.5M) as a positive control
for 24 h, and then the Comet assay was performed. (B) Western
Figure 4. (A) PPDE treatment upregulates Bax gene expression in cultured human cardiomyocytes.
Human cardiomyocytes were treated with or without PPDE (25 mg/ml), and then total RNA was
prepared following 4 h of treatment. Bcl-2, Bcl-xL, and Bax mRNA expression levels in
cardiomyocytes were determined by quantitative real-time RT-PCR. (n=8) Data are expressed as the
mean SD of 8 different PPDE.* P<0.01 versus control. (B)Bcl-2/Bax and Bcl-xL/Bax ratio of
experiment (A). Data are expressed as meanSD of 8 different PPDE. * P<0.01 versus control. :
no treatment, medium only ; : no treatment PBS only ; : medium + stable PDE without
peritonitis(SPDE) ( treated with medium and peritonitis peritoneal dialysate effluent (PPDE)) ;
Figure 5. Effects of PPDE on expression of cardiac GATA-4 mRNA and protein. Cultured human
cardiomyocytes were treated with or without PPDE (25 mg/ml), and then protein extracts and total
RNA were prepared following 4 h of treatment. (A) GATA-4 mRNA expression levels in cultured
human cardiomyocytes as determined by quantitative real-time RT-PCR. (B) Western blot showing
by densitometry. Data are expressed as meanSD of 8 different PPDE. * P<0.01 versus control.
Figure 6. PPDE treatment reduces ERK phosyhorylation, GATA4, Bcl-2, Bcl-xL expression and
enhances Bax expression in cultured human cardiomyocytes. Cardiomyocytes were treated with
medium only, PBS only, SPDE (25 mg/ml) for 2 and 4 hours and PPDE (25 mg/ml) for 2 and 4
hours. Cell lysates were separated by SDS-PAGE and specific monoclonal antibodies were used to
detect phosphorylated and total ERK, and Bax, Bcl-2, Bcl-xL, GATA4 expression. Representative
blots from 8 separate experiments were shown. Quantitative densitometry expressed as
phosphorylated protein relative to total protein. Data are expressed as the meanSD of 8 different
PPDE .* P<0.01 versus control. Medium only, PBS only and SPDE (25 mg/ml) in medium were
Table 1. Dose Dependent manner of PPDE induced cell apoptosis in cultured human cardiomyocytes. % of apoptosis Treatment 0 12.5 mg/ml 25 mg/ml Medium only 1.0 ± 2.7 10.4 ± 2.8 11.8 ± 3.2 PBS only 2.2 ± 3.1 10.2 ± 3.2 10.8 ± 3.6 Medium + DOXO 9.7 ± 2.4 28.5 ± 4.1* 41.2 ± 4.5* Medium + SPDE 0.7 ± 2.5 10.2 ± 2.7 12.4 ± 3.5 Medium + PPDE 9.7 ± 2.6 32.4 ± 3.8* 48.6 ± 4.8*
Cultured human cardiomyocytes were exposed to medium only, PBS only, medium plus DOXO
(0.5μM), medium plus SPDE (24 hours) and medium + PPDE (24hrs). Annexin V-FITC apoptosis
anlysis were determined by FACS. Each value is the mean from eight independent peritonitis
episodes.
Table 2. Time dependent manner of PPDE induced cell apoptosis in cultured human cardiomyocytes. % of apoptosis Treatment 0 24 h 48 h Medium only 1.0 ± 2.7 11.5 ± 2.7 12.1 ± 3.2 PBS only 2.2 ± 3.1 10.4 ± 2.6 11.5 ± 3.4 Medium + DOXO 9.7 ± 2.4 34.8 ± 3.6* 42.2 ± 4.2* Medium + SPDE 0.7 ± 2.5 12.2 ± 3.2 13.6 ± 3.2 Medium + PPDE 9.7 ± 2.6 48.2 ± 4.7* 57.6 ± 4.4*
Cultured human cardiomyocytes were exposed to medium only, PBS (phosphate buffer solution)
only, medium plus DOXO (0.5μM), medium + SPDE (stable peritoneal dialysate effluent) (25
mg/ml) and medium + PPDE (peritonitis peritoneal dialysate effluent) (25 mg/ml). Annexin V-FITC
apoptosis anlysis were determined by FACS. Each value was the mean from eight independent
peritonitis episodes.