探討轉錄因子HBP1調控3T3-L1脂質生合成的角色; The Role of HBP1 Specific Regulation During Adipocyte Differentiation
108
0
0
全文
(2) 目錄 縮寫對照表 ................................................................................................................................ I 中文摘要 ..................................................................................................................................III 英文摘要 ................................................................................................................................. IV 第一章 前言 ..............................................................................................................................1 第二章 文獻探討 ......................................................................................................................2 一、脂質生合成 (Adipogenesis) ....................................................................................2 二、3T3-L1 前脂肪細胞 ..................................................................................................3 三、脂肪分化轉錄因子和下游調控基因的介紹 ............................................................5 四、HBP1 (HMG(High-mobility group)-Box Protein 1) ..............................................11. (一) HBP1 對細胞週期的調控 ..................................................................................11 (二) HBP1 抑制 Wnt 訊息傳遞路徑 ......................................................................14 (三) HBP1 對細胞分化的調控 ...................................................................... 15 五、槲皮素 (Quercetin) ................................................................................... 16 六、實驗目的...................................................................................................... 19 第三章 材料與方法.................................................................................................... 19 一、實驗設計...................................................................................................... 20 二、實驗藥品與儀器.......................................................................................... 21 三、細胞培養...................................................................................................... 25 四、細胞存活率 (Viability) 測定 ..................................................................... 28 五、油紅染色(Oil red O stain) ......................................................................... 29 六、質體 (plasmid) 的製備 .............................................................................. 30 i.
(3) 七、冷光酵素報導基因分析法.......................................................................... 33 八、Transient HBP1 knockdown 之3T3-L1 細胞株 ........................................ 34 九、RNA萃取分離--使用RNeasy® Mini Kit …………………………………..37 十、RT-PCR (Reverse Transcriptase PCR)--使用SuperScriptTMⅢ One-Step RT-PCR System with Platinum® Taq DNA Polymerase Kit ....................... 38 十一、Quantiative -PCR(Quantiative Reverse Transcription-PCR)使用 Taq Man® One-Step RT-PCR Master Mix Reagents .................................................. 40 十二、蛋白質萃取.............................................................................................. 41 十三、蛋白質定量 .............................................................................................. 42 十四、Western blot.............................................................................................. 43 十五、流式細胞儀 .............................................................................................. 47 十六、統計分析 .................................................................................................. 49 第四章 結果................................................................................................................ 50 第五章 討論................................................................................................................ 80 第六章 結論................................................................................................................ 85 參考資料...................................................................................................................... 86. ii.
(4) 圖目錄 圖 2-1 脂質生合成 ....................................................................................................... 3 圖 2-2 分化實驗流程 ................................................................................................... 5 圖 2-3 脂質生合成過程 ............................................................................................... 6 圖 2-4 脂質生合成的轉錄調控 ................................................................................... 6 圖 2-5 PPAR 之結構特徵............................................................................................. 8 圖 2-6 PPAR 之基因轉錄作用機制.............................................................................. 9 圖 2-7 HBP1 的結構圖 .............................................................................................. 11 圖 2-8 HBP1 對細胞週期的調控 .............................................................................. 12 圖 2-9 p38 對 HBP1 的調控 ..................................................................................... 13 圖 2-10 p38 對細胞凋亡的作用 ................................................................................ 14 圖 2-11Wnt 訊息傳遞與 HBP1 的作用 .................................................................... 15 圖 2-12 Quercetin 結構式 ........................................................................................... 16 圖 3-1 RNAi 之作用原理 ........................................................................................... 35 圖 4-1 HBP1 蛋白質在脂肪分化過程的表現 .......................................................... 58 圖 4-2 HBP1 mRNA 在脂肪分化過程的表現 ........................................................... 59 圖 4-3 Insulin 活化 HBP1 蛋白質的表現 ............................................................... 60 圖 4-4 Insulin 誘發 HBP1 -2kb promoter 的活性 ................................................... 61 圖 4-5 HBP1 siRNA 影響 3T3-L1 細胞中 HBP1 蛋白質(A)、mRNA(B)的表現量62 圖 4-6 HBP1 siRNA 對脂肪細胞分化之影響 ........................................................... 63 圖 4-7 HBP1 siRNA 影響脂肪分化相關因子的表現 ............................................... 64 圖 4-8 HBP1 siRNA 對脂肪細胞 leptin 表現量之影響 ............................................ 65 圖 4-9 HBP1 siRNA 影響 MCE 期細胞週期的進行................................................ 66 圖 4-10 HBP1siRNA 影響 MCE 期 C/EBP β轉錄因子 mRNA 的表現 ............... 67 iii.
(5) 圖 4-11 HBP1 的起動子上兩個可能的 PPAR γ連結位點 ................................... 68 圖 4-12 PPAR γ促進劑 TZD 對 HBP1 -2kb promoter 活性的影響 ...................... 69 圖 4-13 PPAR γ抑制劑 GW9662 對 HBP1 -2kb promoter 活性的影響 .............. 70 圖 4-14 PPAR γ抑制劑 GW9662 對 HBP1 mRNA 表現之影響 .......................... 71 圖 4-15 Quercetin 對 3T3-L1 細胞脂肪滴形成之影響 ........................................... 72 圖 4-16 Quercetin 影響 3T3-L1 脂肪細胞分化相關轉錄因子 PPAR γ和特定 基因 aP2 的表現 ............................................................................................ 73 圖 4-17 Quercetin 對 MCE 期 3T3-L1 細胞生長之影響........................................ 74 圖 4-18 Quercetin 影響 3T3-L1 前脂肪細胞 MCE 期細胞週期的進行................. 75 圖 4-19 Quercetin 影響 3T3-L1 MCE 期 C/EBP β和 C/EBP δ mRNA 的表現 76 圖 4-20 Quercetin 對 3T3-L1 前脂肪細胞 HBP1 蛋白質表現之影響 .................. 77 圖 4-21 Quercetin 對 HBP1 -2kb promoter 活性之影響 .......................................... 78 圖 4-22 Quercetin 影響 MCE 期 HBP1 mRNA 的表現……………………………79. iv.
(6) 表目錄 表一 槲皮素(quercetin) 化學性質………………………………………………17. v.
(7) 縮寫對照表 縮寫. 全名. ACC. Acetyl-CoA carboxylase. ADD1/SREBP1. Adipocyte differentiation factor 1/sterol regulatory element binding protein. aP2. Adipocyte Specific Fatty acid binding protein. AF-1. Activation fuction 1. AF-2. Activation fuction 2. C/EBPs. CCAAT/enhancer binding protein. CDK. Cyclin dependent kinase. DBD. DNA binding domain. DEX. Dexamethasone. Dsh. Dishevelled. DKK. Dickkopf. E2F. Early gene 2 factor. GPDH. Glycerophosphate dehydrogenase. HBP1. HMG(high-mobility group)-Box Protein 1. IGF-1 receptor. Insulin-like growth factor receptor. JNK. c-Jun N-terminal kinase. LBD. Ligand binding domain. LPL. Lipoprotein lipase. MCE. mitotic clonal expansion. MIX. Methylisobutylxanthine. I.
(8) pRb. Phospho Retinoblastoma. PPRE. Peroxisome proliferator responsive element. P38 MAPK. P38 mitogen activated protein kinase. PARP. Poly-ADP-ribose polymerase. p-ERK. Phospho-extracellular signal-related kinase. PPAR γ. Peroxisome proliferators-activated receptorγ. sFRPs. secreted Frizzled Related Proteins. TG. Triglyceride. TZDs. Thiazolidinediones. II.
(9) 摘要 肥胖與許多慢性疾病的發生息息相關,防治肥胖的研究從未間斷,尤其,有 關脂質生合成中關鍵標地的研究更顯重要。早期研究發現,轉錄因子HBP1的表 現在脂肪細胞分化完成後有明顯增加,但其調控機制並未被探討。因此,我們想 藉由此次的研究,探討HBP1在脂肪細胞分化過程的角色和相關調控機制。首先, DEX/MIX協同Insulin誘導3T3-L1前脂肪細胞分化為成熟脂肪細胞的8天過程中, HBP1的表現在分化初期2天的複製擴增(Mitotic Clonal Expansion, MCE)期無明 顯改變,但在4到6天的分化後期(terminal differentiation),HBP1的表現明顯增 加,結果顯示,HBP1在分化過程的不同階段可能有不同的功能,同時,insulin 亦有誘發HBP1表現的能力。為了進一步瞭解 HBP1的階段性功能,在分化初期, 我們利用槲皮素 (Quercetin)誘發HBP1的表現增加,導致了複製擴增期的生長 停滯於G1期,進而影響了脂肪細胞的最終分化,相對地,當HBP1的表現以 HBP1siRNA降解時,促進了複製擴增期的細胞生長,表示HBP1在分化的複製擴 增期有調控細胞週期進行的作用。有趣地是,HBP1 siRNA雖然加速分化前期的 複製擴增,卻依然造成最終分化的不完全,表示HBP1在分化後期的增加表現是 必要的,因此,我們假設HBP1可能是分化後期PPARγ的調控標地。當我們利用 CHIP-Mapper搜尋時,發現HBP1的啟動子上有兩個可能的PPARγ連結點,尤其, PPARγ抑制劑GW9662不但抑制HBP1啟動子的活性,也抑制了HBP1 mRNA的 表現,相對地,PPARγ活性促進劑TZD造成HBP1啟動子的活化。結論顯示, HBP1在脂質生合成過程中的複製擴增期和分化後期功能不同。我們希望此次的 研究結果,可以支持HBP1在未來作為一個肥胖相關疾病重要指標和調控標地的 可能性。. III.
(10) Abstract Obesity is an epidemic problem in most societies around the world. The obesity-related study has become the important topics in nutrition research. One early study showed that the transcription factor HBP1 is highly expressed in the adipose tissue in rat, wherein the HBP1 mRNA is more abundant in the fully differentiated adipocytes than those of the undifferentiated ones. In the current study, we confirm the previous finding that HBP1expression is not increased until the late stage of the 3T3-L1 adipocyte differentiation. These observations lead us to hypothesize that HBP1 may have stage-specific function during adipocyte differentiation. To test the hypothesis, in the early Mitotic Clonal Expansion (MCE) stage, we treated 3T3-L1 cells with querceuin to increase HBP1 expression, resulting in the growth arrest in G1 phase and consequent incompletion of adipocytes differentiation. Oppositely, HBP1 siRNA which down-regulates HBP1 leads to accelerated cell growth with early appearance of G2/M phase in MCE. These results indicate that HBP1 is involved in cell cycle regulation in MCE phase. Interestingly, although HBP1knockdown causes early MCE completion, the full adipogenesis process is impaired. This indicates that HBP1 activation during terminal differentiation stage is necessary for full adipogenesis. We further hypothesize that HBP1 is a potential target of PPARγ function during terminal differentiation. First, the CHIP-Mapper search discovers 2 putative PPARγbinding sites in HBP1 promoter. Moreover, GW9662 (PPARγ antagonist) inhibits the activation of a -2kb HBP1promoter and decreases HBP1 mRNA expression. Taken together, the results of the current study will further characterize HBP1 as a key marker of adipocyte differentiation, which may serve as a potential biological target in the intervention of obesity treatment.. IV.
(11) 第一章 前言. 第一章、 前言. 隨著工商業的發展,飲食精緻化走向,加上國人飲食習慣的改變,肥胖已是 國人常見之健康問題,在九十六年國人的十大死因中,與肥胖相關疾病心臟疾病、 腦血管疾病與糖尿病分別位居第二、三、四名. (1, 2). ,因此,肥胖的預防與治療將. 會是營養上不容忽視之重要課題。. 同時,天然食品及其活性成分用來進行養生保健、預防疾病之觀念也愈來愈 受到重視,許多體內(in vivo)和體外(in vitro)之研究亦證實,許多食物及其 活性成分的確具有預防肥胖之功效(3)。Quercetin 又稱槲皮素,屬於天然化合物類黃酮分子(flavonoid),廣泛地分佈於植物界中,如日常食用之蔬菜、水果及綠 茶等(4),研究發現,Quercetin 具抑制脂肪分化的功效,然而關於 Quercetin 抑制 脂肪分化的作用機轉尚不明確。. 更重要的是,早期的研究發現 HBP1的表現在脂肪分化後期明顯增加,但其 扮演的角色並不清楚,因此,我們想藉由此次的研究來探討 HBP1在脂肪細胞分 化過程中的角色,以及 HBP1 參與 Quercetin 調控脂質生合成之重要機制,最 終,希望我們的研究探討可以提供HBP1轉錄因子在未來肥胖的研究上作為一個 重要指標或肥胖相關疾病重要標定的可能性。. 1.
(12) 第二章 文獻探討. 第二章、文獻探討. 一、脂質生合成 (Adipogenesis). 脂肪是哺乳動物體內儲存及供應能量的重要器官,過多的糖類和脂肪攝取, 可以轉變合成為三酸甘油脂 (triglyceride,TG),儲存於脂肪細胞中。而前脂肪細 胞形成成熟脂肪細胞的過程稱之脂質生合成 (Adipogenesis),此過程可分為三個 階 段 , 包 括 前 脂 肪 細 胞 增 生 階 段 (preadipocyte proliferation) 、 分 化 階 段 (differentiation)和分化末期階段 (terminal differentiation),分別說明如下:(i) 前 脂肪細胞增生階段:前脂肪細胞進行細胞生長使細胞數目增加,當前脂肪細胞長 滿形成 monolayer 後,細胞生長停滯,以 Insulin、Dexamethasone(DEX) 和 Methylisobutylxanthine(MIX) 刺激進入 mitotic clonal expansion (MCE)期,再進行 1~2 次的有絲分裂,之後細胞生長停止並開始進入分化階段,Entenmann 等學者 指出,人類初代脂肪細胞不須經過細胞分裂即能完成脂肪分化,但在 3T3-L1 脂 肪細胞分化過程則必須經過 MCE 期才能完成分化(5, 6)。(ii)分化階段:當細胞受 到誘發進行分化後,細胞型態開始改變,由紡錘型轉變成圓亮的油滴,並伴隨三 酸甘油脂的生成 ,而 LPL (lipoprotein lipase)可作為分化早期的指標(7),之後, C/EBPs (CCAAT/enhancer binding protein)和 PPARγ(peroxisome proliferators activated receptor γ)表現增加(8)。(iii)分化末期階段:分化末期 TG 不斷的累積, 使細胞內油滴變大,形成成熟脂肪細胞,而成熟脂肪細胞為內分泌器官,會分泌 許多脂肪細胞特定之荷爾蒙,如 leptin、resistin 等蛋白質(9)。. 2.
(13) 第二章 文獻探討. Cell seeding confluence Post-confluence(Day 0) Day2. Day4. Day6. Day8. MCE. 前脂肪細胞增生階段. 分化階段. 分化末期階段. 圖 2-1 脂質生合成 資料來源:(10). 二、3T3-L1 前脂肪細胞. 近來研究指出,前脂肪細胞株如 3T3-L1、Ob1771、3T3-F422A(11)和初代脂 肪細胞等為研究脂肪細胞分化常用之體外模式,3T3-L1 在 1974 年由 Green 和 Kehinde 從 Swiss mouse embryo cloning 出來具高度分化為成熟脂肪細胞潛力的 前脂肪細胞株(12),這些細胞株在適當的刺激下會分化成脂肪細胞,並表現其生 理功能,若將 3T3-L1 前脂肪細胞殖入小鼠體內,此前脂肪細胞會自發行成脂肪 細胞,且不易與小鼠原有的脂肪區別(13)。 典型的 in vitro 的分化模式中,當細胞在培養皿長滿單層後,更換培養液為 添加數種化學藥品或荷爾蒙的分化培養基(differentiation medium),以促進分化, 分別如下說明: (一) Dexamethasone(DEX) DEX 為一種合成的 glucocorticoid agonist,會與 glucocorticoid receptor 作用,. 3.
(14) 第二章 文獻探討. 促使 C/EBP β和 C/EBP δ形成 heterodimers,進而活化 PPAR γ表現(14, 15, 16)。 (二) Methylisobutylxanthine(MIX) MIX 和 DEX 同樣具備促進脂肪細胞分化的作用,主要是透過增加 C/EBP β 表現,藉由抑制 c-AMP phosphodiesterase 及競爭抑制 A1 adenosine receptor,並 透過阻斷 inhibitory regulatory protein G1 刺激 adenyl cyclase 活性,以提高細胞 內 cAMP 的含量(17)。. (三) Insulin Insulin 主要是與 insulin-like growth factor receptor (IGF-1 receptor)作用,活化 其傳遞路徑以達到脂肪細胞分化,亦可促進 deoxylglucose 汲取,將 glucose 轉換 成 CO2 及 lipid,另外 p38 mitogen activated protein kinase (MAPK) 可促進前脂 肪細胞自發性分化作用(5),加上 insulin 可以刺激 p38 MAPK 活性(18),所以可以 促進分化,同時在分化後的第三或第四天起 insulin binding 活性會增加 6-10 倍 (19). 。. (四) Biotin Biotin 為脂肪酸合成之關鍵酵素 ACC (acetyl-CoA carboxylase) 的輔酶,進而 促進脂肪酸合成。. 4.
(15) 第二章 文獻探討. 2 days post-confluent. 0. ~8. 2. Mitotic Clonal Expression (MCE). (MDI). (Day). Terminal Differentiation. Insulin、Biotin. 1.3-isobutyl-1-Methylisobutylxanthine(IBMX) 2.Dexamethasone(DEX) 3.Insulin. 圖 2-2 分化實驗流程. 三、脂肪分化轉錄因子和下游調控基因的介紹. 前脂肪細胞株分化成脂肪細胞時,數種關鍵轉錄因子會相互作用影響,扮演 極重要角色(20),一般在 3T3-L1 分化過程中,細胞會相互接觸形成 confluence, 此時分裂階段處於 cell cycle G0/G1 期,接著細胞會產生 contact inhibition 現象,並有 LPL (lipoprotein lipase)出現,當加入 DEX、MIX 和 insulin 誘導分化後的一小時,early response 基因 c-myc、c-fos、c-jun 和 c-junB 開始表現, 隨後,參與分化重要的轉錄因子陸續增加表現,首先被誘發的為 C/EBPβ和 C/EBPδ(21),而 C/EBPβ和 C/EBPδ並共同啟動 PPAR γ的轉錄(22),adipocyte differentiation factor 1 /sterol regulatory element binding protein (ADD1/SREBP1)藉 由啟動內生性的 PPAR γ ligand,進而活化 PPAR-γ,之後 C/EBPα與 PPARγ 間不斷的相互作用。最後,共同啟動脂肪細胞特定基因的表現,如 leptin、fatty acid binding protein (aP2)、adipsin、GPDH (glycerophosphate dehydrogenase) 和 ACC,造成促使細胞內 TG 的生成和油滴的堆積(23)。. 5.
(16) 第二章 文獻探討. 圖 2-3 脂質生合成過程 資料來源:(10). 圖 2-4 脂質生合成的轉錄調控 資料來源:(25). 6.
(17) 第二章 文獻探討. (一) 脂肪分化轉錄因子- C/EBPs(CCAAT/enhancer binding protein) C/EBPs 為 basic leucine zipper 成員之一,包括了 C/EBP α、C/EBP β、 C/EBP δ等,最早於 1998 年, Landschulz 等學者在一些病毒的基因中 CCAAT 及相似結合的蛋白實驗中發現第一個成員 C/EBP α,而 C/EBPs 家族可調節許 多細胞中 mRNA 的轉錄作用,在脂肪細胞生合成過程中,C/EBPs 家族的表現 深深影響脂肪細胞的發展。在 3T3-L1 前脂肪細胞誘導分化時,C/EBP β和 C/EBP δ是最早且短暫表現的基因,C/EBP α則在分化後期被誘發出來,進而 轉錄活化 Adipocyte specific gene 的啟動子(26)。. 在 Gain- of – function 的體外實驗研究中顯示,C/EBP β在沒有分化劑荷爾 蒙的催化下,異位表現的 C/EBP β能夠誘發 3T3-L1 細胞的分化作用,但對 C/EBP δ而言,仍需有促分化劑的存在. (27). 。另外,近年來有一些轉錄因子可能. 與脂肪細胞生合成有關,但不像 PPAR γ和 C/EBPs 為主要調控者,它們可能 是做為脂肪細胞生合成某些層面的調節者,如 GATA 家族、FOXC2 和 LXR α。. 根據目前的研究發現,生長停滯的前脂肪細胞 3T3-L1 經由賀爾蒙刺激分化 時會快速增加細胞中 C/EBP-β 的表現。然而,C/EBP β 與 DNA 結合的能力是出 現在分化過程的 MCE 期,此時細胞進入細胞週期 S 期,由於,C/EBP β 在體外 會受到氧化作用而促進與 DNA 結合的能力,所以,在脂質分化過程中,氧化還 原態在 MCE 期及 C/EBP β 與 DNA 結合能力上扮演重要的角色。當 3T3L-1 受到 H2O2 及賀爾蒙刺激時,PPAR γ 表現增加進而促進脂質分化,細胞週期從 S 期快 速地進入 G2/M 期,同時,免疫螢光染色亦顯示,H2O2 使得 C/EBP β 與其調控 片段結合的現象提早出現,而給予抗氧化劑(50 μM genistein、50 μM resveratrol) 後,則會使細胞停留在細胞週期 S 期,進而抑制脂質生合成(28)。. 7.
(18) 第二章 文獻探討. (二) 脂肪分化轉錄因子- PPAR (peroxisome proliferator activated receptor) 在 1990 年的一篇文獻回顧中有提到,Issemann 與 Green 利用固醇類荷爾蒙 核受器 (steroid hormone nuclear receptor) 基因相似性質,自小鼠肝臟 cDNA library 選殖出與過氧化相關之 PPAR α(29),並發現可促使過氧小體增生,故稱 之 為 過 氧 化 體 增 殖 劑 活 化 受 器 (peroxisome proliferator-activated receptor , PPAR)。 目前,在不同之脊椎動物中已鑑定出 3 種不同型態之 PPAR,分別為 PPAR α(NR1C1)、PPAR δ/β(NR1C2) 和 PPAR γ(NR1C3),此三種 PPAR isoforms 在結構和功能上有相似的特性,主要由四個 Functional domains 中的六個 Structural regions A/B、C、D 與 E/F 所組成,N 端的 A/B domains 為 PPAR 的可 磷酸化區段 – independent activation fuction 1 (AF-1) (30);C domain /或 DNA binding domain (DBD) 為 PPAR 與目標基因啟動子上 peroxisome proliferator responsive element (PPRE) 結合(31, 32);D site 為 Cofactors 的 Docking domain;E domain 或 Ligand binding domain (LBD) 則為對 Ligand 具專一性的連結區(33),而 位在 E/F domain – independent activation fuction 2 (AF-2)具活化 PPAR ,促進基因 的轉錄(34)。. 圖 2-5 PPAR 之結構特徵 資料來源:(35). 8.
(19) 第二章 文獻探討. 其中 PPAR 含三種 isoform 分別是 PPAR α、PPAR β/δ和 PPAR γ,其 中 PPAR γ專一表現在脂肪組織,在γ1、γ2、γ3 三種不同的轉錄產物中,脂 肪細胞主要表達為 PPAR γ2 (36),而 PPAR α與 PPAR γ1 主要表現於可代謝脂 肪的組織,如肝、腎、心等,調控脂質相關代謝基因轉錄,另外,有研究指出, PPAR α在血管內皮細胞、單核細胞與巨噬細胞亦有表現,而 PPAR δ則分布於 全身各組織。 臨床上治療糖尿病的藥物Thiazolidinediones (TZDs) 類藥物,如pioglitazone (Actos® )、rosiglitazone (Avandia® )等(37,. 38). ,胰島素增敏劑 (agonist) 可經由活化. PPAR γ來降低胰島素阻抗進而改善血糖。目前已知 TZDs 的作用標的在PPAR γ 接受體,PPAR-γ 是細胞核內 PPARs的一種接受器,含有接合子結合區 (ligand binding domain )和輔助活化子接合區 (coactivator binding site),與核內另 一接受體 retinoid X receptor( RXR )結合成異質雙合體(heterodimer )的結構,維持 在不活化狀態,而當接合子與PPAR-γ 接合,則促使構象變化( conformational changes ),經由與特定基因對應的 DNA sequences 作用,調節基因轉錄表現(39)。. 圖2-6 PPAR之基因轉錄作用機制 資料來源:(39). 9.
(20) 第二章 文獻探討. 研究指出,TZDs 會促進脂肪細胞分化,因此,在治療糖尿病、降血糖的同 時,服用 TZDs 的病患易有體重增加的副作用(40, 41,42),動物實驗證實,當老鼠脂 肪不表達 PPAR γ時,TZDs 降血糖的效果並不明顯,另一方面,與血糖調控相 關的二大組織 – 肌肉和肝臟,若未表達 PPAR γ,該動物仍有降血糖的功效(43,44), 由此得知,PPAR γ為脂肪組織發展所必需。. (三) 下游調控基因- leptin 瘦體素 1994 年,Zhang 等學者在 ob/ob 肥胖鼷鼠上發現瘦體素(leptin),leptin 主要 在體內脂肪組織表現,由 167 個胺基酸所組成,分子量約 16 kDa 蛋白質(45)。文 獻指出,ob/ob 肥胖鼷鼠為 ob 基因發生突變,無法合成正常具有功能的瘦體素, 導致肥胖,當給予 ob/ob 肥胖鼷鼠瘦體素後,它們的攝食量、體脂肪、高血糖和 高胰島素血症有明顯改善的症狀(46,. 47, 48). ,因此,瘦體素被認為是一個調脂因子. (lipostatic factor) (48)。 另一方面,對人類而言,當脂肪細胞儲存過多的能量時,分泌大量的脂肪激 素 (adipokines) (49),其中瘦體素主要作用為調控食慾,抑制神經胜肽 Y 和 AGRP 的活性,進而降低攝食量和促進能量消耗維持體脂肪的恆定,因此,脂肪組織不 只是一個能量儲存的場所,更是調節能量代謝的重要調控因子。 更進一步地,Mac Djougald 等學者指出,胰島素和 leptin 的調控具相關性, 在糖尿病老鼠的脂肪組織上施打 insulin,可促進 leptin mRNA 的表現(50)。此外, 有許多報告亦指出,大部分肥胖的人,血中 leptin 的濃度較高,表示肥胖呈現出 leptin 抗性階段,可能與第二型糖尿病有關或與胰島素訊息傳遞發生問題、leptin 受器出現問題具相關性(51)。. 10.
(21) 第二章 文獻探討. 四、HBP1(HMG(Hihg-mobility group)-Box Protein 1). HBP1 屬於HMG (high-mobility group) 家族的成員,為一轉錄因子,大小約 513個胺基酸,其結構包含: --HMG-box DNA 結合區 (HMG-box DNA binding domain) --抑制區 (repression domain) --Rb 結合區 (Rb binding domain) --p38 結合區 (p38 binding domain). 圖 2-7. HBP1 的結構圖. 資料來源:(52). HBP1 的生物功能近年來陸續地被發掘探討,HBP1 參與細胞生長的調控主 要著重於 HBP1 抑制細胞週期進行以及 HBP1 調控肌肉細胞分化的角色。. (一) HBP1 對細胞週期的調控 HBP1 與 Rb 結合抑制細胞週期 G1 的進行 HBP1影響細胞週期與 pRb (retinoblastoma,also called pocket protein) 對G1 期的調控功能相似。pRb 家族為細胞週期調控的核心之一,包括 pRb1/p110、. 11.
(22) 第二章 文獻探討. pRb2/p130 和p107 三個,它可以使細胞週期停滯於 G1期, pRb 及其他成員 p107 和p130 可分別與轉錄因子 E2F (early gene 2 factor) 結合而抑制 E2F的轉 錄功能(53),當pRb 被cyclin 與CDK (cyclin dependent kinase)磷酸化後,Rb與E2F 分離,E2F 釋放而使細胞週期得以進行。研究證實,HBP1可和Rb家族成員中的 Rb與p130 作用,進而抑制細胞週期G1 期的進行. (53, 54, 55). 。另外,早期研究指出,. Rb(56)、E2F(57)和 CDK inhibitor(58) 與3T3-L1 細胞分化時生長停滯有所關聯。. 圖 2-8 HBP1 對細胞週期的調控 資料來源:(52). HBP1 為p38 MAPK 路徑抑制細胞週期之直接作用目標 p38 MAPK 訊息傳遞路徑與細胞凋亡和細胞週期停滯有關,最近的研究顯 示,p38 MAPK 可藉由直接磷酸化而穩定HBP1的表現,造成細胞週期停滯於G1 期,而抑制乳癌細胞的生長。 12.
(23) 第二章 文獻探討. 綜觀,HBP1調解細胞週期的標的基因包括N-myc、c-myc 以及cyclin D1。在 HBP1 過度表現的細胞中,可誘導細胞週期停滯在G1/S 期;而在肝細胞再生模式中, 當基因轉植小鼠的肝臟 HBP1 表現增加時,能延遲G1期的發展。最後,最近的研 究指出,p38 MAPK 調控HBP1 的蛋白表現,造成G1期的停滯(52)。因此,在細胞及 動物的細胞週期實驗確立了HBP1 在調控G1 期發展的角色,這樣的發現,也顯示 出HBP1藉由抑制細胞週期的活化因子,進而抑制細胞增生。. 圖 2-9. p38 對 HBP1 的調控. 資料來源:(52). 13.
(24) 第二章 文獻探討. 圖 2-10. p38 對細胞凋亡的作用. 資料來源:(52). (二) HBP1抑制Wnt 訊息傳遞路徑 Wnt 訊息傳遞路徑與腫瘤發生、癌症生成有密切的關係,近年來, Wnt 在 代謝相關疾病(如脂質生合成、糖尿病)的影響亦開始廣泛地被探討。典型的 Wnt 訊息傳遞路徑開始於 Wnt 蛋白質與細胞膜上的受體 Frizzled 及 LRP 結合形 成複合體,並傳遞訊息到細胞質中使 Dishevelled(Dsh)影響 APC、GSK-3β、 Axin 三者無法和β-catenin 形成複合體,使得β-catenin 不被分解,β-catenin 進 而轉移至細胞核進行調控下游基因的轉錄(59,60,61)。同時,轉錄因子 HBP1 在細胞 生長調控的研究上,發現可藉由與β-catenin 競爭 Lef/Tcf 結合至 cyclin D1、c-myc 之啟動子(promoter)來抑制 Wnt 訊息傳遞路徑的基因活化作用。. 14.
(25) 第二章 文獻探討. 圖 2-11. Wnt 訊息傳遞與 HBP1 的作用. 資料來源:(59). (三) HBP1 對細胞分化的調控 HBP1 不但調節細胞週期的進行,而抑制細胞的增生。而且,在肌肉細胞 C2C12 中,細胞週期停滯的情況下,HBP1的活化更會促使肌肉細胞的分化。此外,. Lesage 等學者發現,在大鼠組織中,HBP1 mRNA表現在脂肪組織、肺臟含量較 多,另外,在Ob1771 (preadipocyte) 和C2C12 (muscle)的分化過程中,HBP1 mRNA 的表現在分化的組別中顯著高於未分化的組別,同時,在Ob1771細胞分化後, 隨著insulin 濃度增加,HBP1mRNA的表現明顯增加(62),顯示HBP1參與了脂肪 細胞分化的調控,但在脂肪細胞分化過程, HBP1參與的機制尚不明確。. 15.
(26) 第二章 文獻探討. 五、Quercetin 槲皮素. 代謝症候群為心血管疾病及糖尿病等疾病的危險因子,症狀包括腹部脂肪堆 積、動脈粥狀硬化、血脂異常、血壓增高、血糖升高等。雖然惡性腫瘤、心臟病、 心血管疾病與糖尿病等慢性病發生機轉是多因的,但不可否認地,飲食因子是其 中一個相當重要的因素,所以如何排除或減少飲食中與慢性病有關的危險因子或 增加保護因子的攝食,也就成為預防這些疾病的最好方法。近年來,蔬菜及水果 中的天然化學成分,比如:phenols, isoflavones, allium organosulfur compounds及 indoles等,無論在流行病學或臨床研究上,都顯示對人類疾病的產生具有不等程 度的預防作用,同時也可達到改善體質、調節生理機能、增進健康之目的。 蔬果、榖類、豆類中含有許多抗氧化活性的物質,quercetin 是主要的類黃 酮成分,常以配醣體的型式存在於多種的花、果實和葉子(63),亦存在於日常食 用的蔬菜、水果中,如洋蔥、綠茶、蘋果和草莓(64),其在預防癌症、心血管疾 病、體重控制、免疫調節等醫療效用亦受到相當之重視(65)。. 圖 2-12 quercetin 結構式 資料來源:(66). 16.
(27) 第二章 文獻探討. 學名. 槲皮素(quercetin). 化學名. 2-(3,4-Dihydroxyphenyl)3,5,7-trihydroxy-4H-1-Benzopyran-4-one3,3’,4’,5,7-pentahydroxyflavone. 分子式及分子量. C15H10O7,分子量 302.23. 物理及化學性質. 1、於 314℃被分解。 2、UVλmax (aLc):258,375 nm。 3、含二分子水的黃色針狀結晶(稀醇), 於 95-97℃成無水化合物,可溶於冰醋酸, 鹼性溶液下呈黃色,幾乎不溶於水,醇性 溶液嚐起來有苦味。. 表一. 槲皮素(quercetin) 化學性質. 資料來源:(66). 近年來,探討 quercetin 調控脂肪生合成之研究陸續出現, quercetin 影響脂 質生合成之機制包括抑制脂肪於脂肪細胞堆積,Shisheva 等學者使用 47μM quercetin 給予大鼠初代脂肪細胞,發現可以抑制初代脂肪細胞約 50%lipogenesis 反應,投予 110μM quercetin 則可抑制 100%(67),此外,在 3T3-L1 前脂肪細胞, Hsu 等學者則認為 quercetin 抑制脂肪細胞分化可能與細胞凋亡(apoptosis)的 調控有關,向下調控 PARP(Poly-ADP-ribose polymerase)及 Bcl-2 蛋白質,增 加 caspase-3、Bax 等蛋白質的表現,促進細胞凋亡(68)。進一步地,最近的一篇研 究指出,添加 10、50 及 100 μM 的 quercetin 促使 casapse-3、casapse-9 活性增 加,導致成熟脂肪細胞凋亡比率增加,且隨著 quercetin 濃度的增加亦會降低脂 肪細胞脂質生合成,並有效減少脂肪粒數目及大小,另外 quercetin 之添加亦會 17.
(28) 第二章 文獻探討. 降低 phospho-extracellular signal-related kinase (P-ERK)、c-Jun N-terminal kinase (JNK) 蛋白質之表現,顯示 quercetin 可經由 MAPK 訊息傳遞路徑造成脂肪細胞 凋亡之發生,而降低脂肪細胞的分化 (69) 。綜合以上所述,quercetin 可能抑制 3T3-L1 前 脂 肪 細 胞 及 脂 肪 細 胞 增 生 和 脂質 生 合 成 , 活 化 促 細胞 凋 亡 因 子 (pro-apoptoic factor)-Bax、Bcl-XS、Bad、aspase-3、caspase-9…等之表現,增 加 PARP 蛋白質斷裂現象;抑制抗細胞凋亡因子 Bcl-2 家族中 Bcl-2、Bcl-XL… 等之表現所導致,造成細胞週期停滯及誘導細胞凋亡發生,預防脂質生合成。. 18.
(29) 第二章 文獻探討. 實驗目的 許多研究曾指出 quercetin 富含於日常生活中蔬菜、水果中,具有抑制脂肪 細胞分化之效用,而早期的研究發現 HBP1 的表現在脂肪分化後期明顯增加, 然而關於其作用機轉之研究尚十分缺乏。因此,本實驗以 3T3-L1 前脂肪細胞株 體外分化模式,探討 HBP1 在脂肪細胞分化過程中的角色,以及 HBP1 參與 quercetin 調控脂質生合成之重要機制。. 19.
(30) 第三章 材料與方法. 第三章、材料與方法. 一、實驗設計. 3T3-L1 Cell. Aim 1. 探討 HBP1 在脂肪分化(adipocyte differentiation)過程的表現. Aim 2 HBP1 的表現量影響脂肪細胞分化過程. 利用 siRNA 將細胞中 HBP1 表現降 解,觀察 HBP1 對脂肪分化的影響. Aim 3 Quercetin 透過對 HBP1 的調控影響 脂肪分化過程. Quercetin 是否可抑制 脂質生合成. 20. Quercetin 階段性調控 HBP1 影響脂質生合成.
(31) 第三章 材料與方法. 二、實驗藥品與儀器 藥品: 名. 稱. 廠. 牌. 產 地. 貯 存 溫 度. A Acrylamide. Genepure. USA. 4℃. Agarose. AMRESCO®. USA. 室溫. Ampicillin. Bio Basic Inc.. Canada. 4℃. Ammonium Persulfate. J. T. Baker. USA. 室溫. β-mercaptoethanol. J. T. Baker. USA. 室溫. Bovin Serum Albumin. USBiological. USA. 4℃. Bromophenol Blue. Bio Basic Inc.. Canada. 室溫. chloroform. TEDIA. USA. 室溫. Deoxycholate. Sigma. US. 室溫. DMSO. Sigma. US. 室溫. Dulbecco’s Modified Eagle. Gibco. USA. 4℃. Dexamethasone. Sigma. US. -20℃. Dual light. Applied. USA. 4℃. Germany. -20℃. B. C. D. Medium. Biosystems DNase I stock solution. Qiagen. 21.
(32) 第三章 材料與方法. E ECL. PerkinElmer. US. 4℃. EcoR I. BioLabs. US. -20℃. EDTA. J. T. Baker. USA. 室溫. Ethanol. 東華化工. Taiwan. 室溫. Etidium Bromide. Genepure. USA. 4℃. Fetal Bovine Serum. Gibco. USA. -20℃. Glycine. USBiological. USA. 室溫. Hind III. Biolabs. US. -20℃. Isopropanol. Sigma. US. 室溫. Insulin. Sigma. US. -20℃. 3-isobutyl-1-methyl-xanthine. Sigma. US. -20℃. L-Glutamine. Gibco. USA. -20℃. Luria Bertani medium. Himedia. India. 室溫. Luria Bertani Agar. Himedia. India. 室溫. Lipofectamine TM 2000. Invitrogen. USA. 4℃. Lipofectamine TM RNAi. Invitrogen. USA. 4℃. F. G. H. I. L. MAX. 22.
(33) 第三章 材料與方法. N NaCl. AMRESCO®. USA. 室溫. 10x NEBuffer EcoR I. Biolabs. US. -20℃. OPTI-MEM®. Gibco. USA. 4℃. Oil red O. Sigma. US. 室溫. 10x PBS. UniRegion. USA. 室溫. O. P. Bio-Tech Penicillin/Streptomycin. Gibco. USA. -20℃. Phosphatase Inhibitors. Sigma. US. -20℃. Polybrene. Sigma. US. -20℃. Protease Inhibitors Cocktail. Sigma. US. -20℃. Protein Assay Dye Reagent. Bio Rad. CA. 4℃. SDS. Merck. Germany. 室溫. Sodium Bicarbonate. Sigma. US. 室溫. SUPERase-In. Ambion. USA. -20℃. 50x TAE. UniRegion. USA. 室溫. Cocktail. S. T. Bio-Tech TEMED. J. T. Baker. USA. 室溫. Tris-base. USBiological. USA. 室溫. I 23.
(34) 第三章 材料與方法. TRIzol®. Invitrogen. USA. 4℃. Triton X-100. J. T. Baker. USA. 室溫. Trypsin. Gibco. USA. -20℃. Tween 20. Showa. Japan. 室溫. 儀器: 名稱. 廠牌. 倒立式顯微鏡. Olympus. 離心機. Sigma. 微量離心機. DENVILLE. iCycler. BioRad. MultilmageTM Light Cabinet. Alpha Innotech Corporation. Mulpid® -2 plus electrophoresis. ADVANCE. CLC-110 chemiluminescence detector. TOHOKU. CLC-10 CL counter. TOHOKU. PowerPacTM Basic Power Supply. BioRad. Thermo Scientific NanoDrop 1000. Thermo. Spectrophotometer Mini Trans-Blot® Electrophoretic. BioRad. Transfer Cell. 24.
(35) 第三章 材料與方法. 三、細胞培養 資料來源:10 (一) 細胞株 3T3-L1前脂肪細胞株:購自ATCC® Catalog NO. CL-173TM,組織來源:小 鼠纖維母細胞。 HEK-293T人類胚胎腎臟上皮細胞株,由賴志河老師實驗室機構所贈。. (二) 試劑配置 DMEM ( high glucose) Dulbecco’s Modified Eagle Medium (DMEM) 粉末溶於800 ml 的無菌水中, 加入1.5 g Sodium Bicarbonate (Sigma),再以1N HCl 調整pH值為7.1~7.2之間,定 量1 L後以過濾膜 (0.22 μm) 過濾,加入 10%Bovine serum (3T3-L1 cell) /Fetal Bovine Serum (293T cell)、2% L-Glutamine 與1% Antibiotic Antimycotic,而成為 最終之細胞培養液。 1X磷酸緩衝液(Phosphate-buffer saline;PBS) 以10X PBS (UniRegion Bio-Tech)以滅菌水稀釋為 1X PBS,經 121℃、30分鐘高 壓滅菌後使用並存放於室溫備用。. (三) 繼代培養 (Subculture) 細胞長滿後,將培養皿中之培養液吸除後,加入 10 ml 滅過菌之PBS 清洗 後,吸除PBS並加入trypsin 1 ml 溶液後,放入37℃培養箱中反應5 分鐘,於倒 立顯微鏡下觀看細胞是否全數脫落。確定細胞完全脫落後,加入10 ml 培養液以 抑制 trypsin 的活性,並將細胞液取出放入15 ml 離心管中,以1000 rpm 速度離 心5 分鐘後,吸除上清液,加入適量的培養液將細胞打散後,依比例稀釋分至裝 有培養液之培養皿中,置於37℃、5% CO2之培養箱內培養。. 25.
(36) 第三章 材料與方法. (四) 細胞冷凍 細胞長滿時,將培養皿中之培養液吸除後,加入 10 ml 滅過菌之PBS 清洗 後,小心地吸除。加入1 ml trypsin 溶液後,放入37℃培養箱中反應5 分鐘,於 倒立顯微鏡下觀看細胞是否全數脫落。確定細胞完全脫落後,加入10 ml 培養液 以抑制trypsin 的活性,並將細胞液取出放入15 ml 離心管中,以1000 rpm 速度 離心5 分鐘後,吸除上清液,每管加入3ml 含有5% DMSO 的培養液,分裝1 ml 至冷凍保存管中,放入4℃冰箱裡10 分鐘後,移至-20℃放置30 分鐘後,放到-80 ℃中24 小時,最後移至液態氮筒中保存。. (五) 細胞分化 3T3-L1 前脂肪細胞培養confluents的單層(monolayer)後,兩天後即 Day 0時, 加入分化的培養液0.25μM Dexamethasone/0.25 mM 3-isobutyl-1-methyl-xanthine/10 μg/mL insulin 於 high glucose DMEM 培養兩 天,每兩天更換 10 μg/mL Insulin / 100 μg/mL Biotin 培養液,持續培養至分 化第八天,細胞即慢慢分化完成,隨著脂肪細胞的分化,於顯微鏡下觀察,細胞 由紡錘狀變為球形,細胞內會堆積許多圓亮的脂肪滴。 藥品配置 1、0.25mM Dexamethasone(DEX):1000x stock 藥品. 需要量. conc.. Dexamethasone(Sigma). 0.098g. 0.25M. DMSO. 1 mL. 先將0.098 g DEX power 溶於 1 mL DMSO中,即為0.25M. 26.
(37) 第三章 材料與方法. 藥品. 需要量. 0.25M DEX. 10 μl. DMSO. 10 mL. Stock conc.. conc.. 0.25 mM. 0.25μM. 先將0.25M DEX以DMSO 稀釋為0.25 mM DEX,分裝於滅菌 eppendorf中, 貯存於-20℃。. 2、0.5 M 3-isobutyl-1-methyl-xanthine(MIX) :1000x stock 藥品. 需要量. Stock conc.. conc.. 3-isobutyl-1-methyl-xanthine(MIX). 0.111g. 0.25 M. 0.25 mM. (Sigma) DMSO. 1 mL. 先將0.111g MIX power 溶於 1 mL DMSO中,分裝於滅菌 eppendorf中,貯存 於-20℃。. 3、100μg/ mL Biotin:1000x stock 藥品. 需要量. Stock conc.. conc.. Biotin(Sigma). 0.01 g. 100 μg/ mL. 0.1 μg/ mL. 二次水. 100 mL. 先將0.01g biotin power 溶於 100 mL二次水中,以0.2μm filter過濾分裝於 eppendorf中,貯存於-20℃。. 27.
(38) 第三章 材料與方法. 4、10 mg/ mL Insulin:1000x stock 藥品. 需要量. Stock conc.. conc.. Insulin(Sigma). 0.1 g. 10 mg/ mL. 10 μg/ mL. 0.01N HCl. 10 mL. 先將0.01g biotin power 溶於 10 mL0.01N HCl 中,以0.2μm filter過濾分裝於 eppendorf中,貯存於-20℃。. 四、細胞存活率 (Viability) 測定 資料來源:70 (一) 原理 正常細胞由於細胞膜完整,遇到加入 trypan blue 染劑時,則不會被染色, 呈現亮點為活細胞;反之,死細胞或是受損的細胞則因細胞膜通透性已被破壞, 染劑可進入細胞內加以染色,成為藍色細胞。. (二) 實驗步驟 藥品配置 100 mM Quercetin 分子量為 338.26,取 2.96 g 之 Quercetin (sigma) ,移至無菌操作台,加入 10 ml DMSO,以 0.22 μm 的濾膜過濾後放置 - 20℃ 儲存。 實驗步驟 在 6 孔盤中種下 5×105 顆的 3T3-L1 細胞,等至細胞貼附長滿後二天時 (MCE期),加入10、50 和 100 μM 不同濃度的 quercetin 處理 48 小時,將細胞 以 trypsin-EDTA 打下後,取細胞懸浮液中的 50 μl 再加上 100 μl 的 trypan blue, 混合均勻後,以血球計數器來計算未被染色的 3T3-L1細胞數目。. 28.
(39) 第三章 材料與方法. 細胞計數 將細胞懸浮液與 trypan blue 按比例體積混合,取出 20 μl 混合液注入血球計 數盤凹槽中,於倒立式顯微鏡下觀察計數。計算計數盤中上下共十大方格中細胞 總數後除以十,乘以稀釋倍數,最後再乘以 104 (血球計數盤每一方格的體積為 10-1 mm3),及為每毫升中細胞懸浮液之細胞數。. 五、油紅染色(Oil red O stain) 資料來源:71 (一) 原理 染脂肪細胞內油滴的生成。. (二) 實驗步驟 藥品配置 取Oil-red O 212 mg加入isopropanol 60 mL中,蓋上蓋子,stir at 4℃ overnight, 以 Whatman 3MM paper過濾,過濾後Oil-Red O solution:dd water=3:2混勻後, 4 ℃ overnight , 最 後 再 以 Whatman 3MM paper 過 濾 , 4 ℃ 待 使 用 。 10 % formalin/PBS 是取 27.3 mL 37% formaldehyde solution+ 72.97 mL PBS。 細胞染色 將分化至第八天的3T3-L1 脂肪細胞,吸除培養液,以 PBS潤洗兩次,加入 2 mL 的10% formalin/PBS固定細胞一小時,再以dd water清洗數次,加入 1.5 mL Oil-red O(蓋過細胞即可)染10 min,吸除染劑,以dd water 清洗數次,室溫倒扣 晾乾於顯微鏡下即可照相。. 29.
(40) 第三章 材料與方法. 六、質體 (plasmid) 的製備 資料來源:72 (一) 原理 質體 DNA 為獨立於染色體DNA外的環狀小分子DNA,通常只有數千個鹽 基對(bp) ,具有自行複製的能力,因此,在生物技術應用上,質體DNA常被用 來作為載體以選殖特定的DNA以及表現特定的蛋白質用。抽取質體DNA的方法 有很多種,其中「鹼性溶解法」最常用的方法,此方法效率高、價廉、簡單為。 其原理是利用鹼處理質體DNA和染色體DNA,使兩者雙股打開呈單股狀態,再 加入酸中和鹼使得單股DNA回復成為雙股DNA。細菌以NaOH及SDS分解,並使 蛋白質及DNA變性,之後以酸中和。因此,小分子質體DNA在中和後恢復原態, 但大部份的細菌染色體DNA則無法完全復原而與SDS-K+ 所形成之複合物一起 沉澱下,可以離心去除,而上清液中所含的質體則可以酒精或異丙醇(isopropanol) 使之沉澱。此外,將分離下來的質體,以不同限制酵素作用,經電泳分離DNA 片段後,比較各DNA片段之分子量,檢定質體之正確性。. 使用的質體 (乾燥) Name. Relevant characteristics. PGL3-HBP1-LUC. transient overexpressing HBP1. RSV-β-gal (Rous sarcoma virus β-galactosidase). internal vector control. 質體來源:Dr. Yee Lab 機構.. (二) 實驗步驟 轉型 (Transformation) 將 DH5α competent cells (ECOS 101;Yeastern Biotech) 自-80℃取出後回. 30.
(41) 第三章 材料與方法. 溫,取 33 μl 至eppendorf 中,加入 3 μl 質體DNA (取決於濃度) 後置於冰 上10 分鐘後,將含有質體DNA 及competent cells 之eppendorf 放入42℃水浴45 秒,此時勿晃動,Heat shock 後立即放回冰上2 分鐘。接著,加入300 μl 之Luria Bertani medium,放入37℃培養箱,以100 rpm 速度震盪培養1 小時,取200 μl 混合液均勻塗抹在含特定抗生素的 Luria Bertani Agar 上,置於37℃培養箱24 小 時。. 挑選菌落 以滅菌過的牙籤或無菌tip輕輕挑起培養皿上單一菌落,連同牙籤或無菌的 tip一起放入含有50 μg/ml 之Ampicillin 之Luria Bertani medium 的培養管中, 再度放入37℃培養箱,以200 rpm 速度震盪培養24 小時。. 質體小量製備--使用QIAGEN® Plasmid Mini Kit (QIAGEN) 將上述培養 24 小時之菌體離心15 分鐘 (6000g,4℃) 後,將上清液倒掉, 加入 300 μl Buffer P1 後震盪均勻後,加入300 μl Buffer P2,上下搖勻4~6 次 後,置於室溫下5 分鐘,接著加入300 μl Buffer P3,混勻後放置冰上5 分鐘後, 離心10 分鐘 (最大速度,4℃),保留上清液。QIAGEN-tip 20 先以1 ml Buffer QBT 通過column 之後,再加入前述之上清液,待其全部通過column 後,以2 ml Buffer QC清洗2 次,接著再以0.8 ml Buffer QF 將DNA elute 至eppendorf 中,加入0.56 ml isopropanol 後離心30 分鐘 (13000 rpm,4℃)。之後,小心地將上清液抽掉, 以1 ml 70%酒精清洗,離心10 分鐘 (10000 rpm,4℃),將上清液抽掉,剩餘的 DNA pellet 使其乾燥5~10 分鐘後,以50 μl 二次水將其溶解,保存於-20℃。. 31.
(42) 第三章 材料與方法. 質體鑑定--以限制酶切割 EcoR I digestion: Component. Volume. Plasmid DNA. 15μl. 10x NEBuffer EcoR I. 5μl. EcoR I (BioLabs). 0.5μl. ddH2O. X μl. total volume. to 50μl. DNA電泳分析 酶切反應物於 37℃水浴反應 24 小時後,每管加入 6 X 的 DNA loading dye 混合均勻,注入含有 Ethidium bromide 之 1% agarose 之膠片中,將膠片置於電 泳槽中,於電泳槽中以 100 伏特進行電泳,以 MultilmageTM Light Cabinet 分析 影像並儲存。. 質體大量製備--使用QIAGEN Plasmid Maxi Kit (QIAGEN) 將上述質體鑑定正確之小量質體,放入含有Ampicillin之LB培養液300 ml 培 養 overnight,離心15 分鐘 (6000 g,4℃) 後,將上清液倒掉,加入10 ml Buffer P1 後震盪均勻後,加入10 ml Buffer P2,上下搖勻4~6 次後,置於室溫下5 分鐘, 接著加入10 ml Buffer P3,混勻後放置冰上20 分鐘後,離心30分鐘 (最大速度, 4℃),保留上清液。QIAGEN-tip 500 先以10 ml Buffer QBT 通過column 之後, 再加入前述之上清液,待其全部通過column 後,以30 ml Buffer QC清洗2 次, 接 著 再 以 15 ml Buffer QF 將 DNA elute 至 eppendorf 中 , 加 入 10.5 ml 之 isopropanol 後離心30 分鐘 (13000 rpm,4℃)。之後,小心地將上清液抽掉,以 5 ml 70%酒精清洗,離心10 分鐘 (10000 rpm,4℃),將上清液抽掉,剩餘的DNA 32.
(43) 第三章 材料與方法. pellet使其乾燥5~10 分鐘後,以200~300μl 二次水將其溶解,保存於-20℃。. 七、冷光酵素報導基因分析法--Luciferase Gene Reporter Assay 資料來源:73 (一) 原理 生物發光現象是 Luciferin 在 Luciferase 的催化下進行氧化反應所產生。 Luciferase + O2 Luciferin-----------------------------------products + CO2 + light Other+ elements (二) 實驗步驟 轉染 以 lipofectamineTM 2000 轉染 0.4 μg HBP1 -2kb promoter 和 0.2 μg RSV-β -gal (internal control)質體至培養於 24 well 每一孔盤中的 293T 細胞,24 小時後, 給予不同濃度的 insulin、TZD 和 GW9662 24 小時後,以 luminometer 分析 Luciferase 活性。 打破細胞 使用 4℃的 PBS 清洗細胞兩次,加入含有 DTT 的 Lysis buffer(DTT final 0.5mM),24 well 每一個 well 25μl,在搖晃機上搖 30 分鐘(冰上) ,將細胞放置 eppendorf,以 11800 rpm,離心 2 分鐘,將上清液放置新的 eppendorf,放到-80 ℃冰箱中保存備用。 分析 加入 12.5μl 的 buffer A 至每一 well,將 2-10μl(5μl)的 test sample 加入 至 well,等待 10 分鐘,等待的時間配置 bufferB,稀釋 galacton-plus substrate 至 bufferB (1:100) ,加入 buffer B,每個 well 加入 50μl,測量 Luciferase。放置 室溫 30 分鐘,加入 50μl Accelerator II,等 1-2sec,立刻測量β-galactosidase。. 33.
(44) 第三章 材料與方法. 八、Transient HBP1 knockdown 之3T3-L1 細胞株 資料來源:74 (一) RNA干擾術(RNA interference;RNAi) 背景 RNAi 最早於 1990 年時被發現,植物學家 Jorgensen 等將查爾酮合酶 (chalcone synthase) 的基因置於一個 promoter 後面,於矮牽牛花中,希望藉由增 加一套基因使花色更鮮豔,結果發現不但多一套的基因沒有表現,原本存在的那 一套基因反而失去功能而出現白花,當時稱此現象為「共同抑制 (co-suppression)」。1995年,Guo 和Kemphues 利用反義股RNA (anti-sense RNA) 阻斷了parf-1 肌肉蛋白的表現,同時,在對照組實驗中,發現 sense RNA 亦可 抑制parf-1的表現。1998 年,Fire 與Mello 在線蟲(Caenorhabditis elegans) 上也 發現這種RNA 干擾現象,並首度被稱為RNAi。直到2001 年初,Thomas Tuschl 證明了在哺乳動物細胞中,亦可以使用RNAi 來抑制特定基因的表現,因此RNAi 技術也由植物延伸到了哺乳類動物,對於其機制也有了更深入的了解。. 原理 RNAi 是利用細胞內的雙股RNA 所引發的基因沈默作用。長鏈的雙股RNA 先經由Dicer (一種RNase) 將其切割成多個小段,每段約19~25nt,稱為小干擾 RNA(small interfering RNA),也就是所謂的siRNA。siRNA 會被細胞質中的核糖 核酸誘導沉默複合體 (RNA-induced silencing complex;RISC) 辨識並結合,使雙 股的siRNA解開,形成sense 與antisense 兩個單股的RNA,並將其中的正義股 (sense) 分解,保留反義股 (antisense)。而保留的反義股則會引導RISC結合至互 補的 mRNA上,之後,mRNA會被RISC切斷進而降解,使其無法轉譯成蛋白質, 因此,目標基因遭到抑制並失去功能。. 34.
(45) 第三章 材料與方法. 應用 RNAi 技術的應用相當廣泛,在疾病治療應用上,許多疾病的發生是因為特 定基因表現異常或過度表現,我們可以依所需來關閉非必要或致病基因的功能, 以抑制這些異常或過度活化的基因。理論上說,若能關閉致病基因的表達,則很 多疾病將被治癒。因此,利用RNAi 技術可應用於抑制病毒感染與複製能力、對 抗癌症、心血管與腦血管疾病、糖尿病…等各種疾病的新療法。. 圖3-1 RNAi 之作用原理 資料來源:(75). 35.
(46) 第三章 材料與方法. (二) 使用的siRNA StealthTM RNAi Negative Universal Control Duplexes Lot. 130942A01做為控制組 Mouse HBP1 siRNA購自Thermo SCIENIFIC,其target Sequence如下: TARGETplus SMARTpool L-054764-01-0005,Mouse HBP1,NM_153198,5 nmol ON-T TARGETplus SMARTpool siRNA J-054764-09,HBP1 Target Sequence:AAUAAAGGUUGGUCGUCAU ON-T TARGETplus SMARTpool siRNA J-054764-10,HBP1 Target Sequence:CCUCAGAUAUACCGGAAAA ON-T TARGETplus SMARTpool siRNA J-054764-11,HBP1 Target Sequence:GGUCAAGAGUUUUCGGGAU ON-T TARGETplus SMARTpool siRNA J-054764-12,HBP1 Target Sequence:CUGAAAGGCACGCGGCUGU. (三) 實驗步驟 轉染至3T3-L1細胞 將3T3-L1細胞培養至10 cm 培養皿中,待長滿後進行繼代培養2~3次,進行 實 驗 。將100 nM siRNA 與serum free medium(Opti-MEM® )混合 ,加入 RNAi duplex-Lipofectamine TM RNAi/MAX反應10-20分鐘後,進行Reverse Transfection, 接著加入500μl含有400,000顆3T3-L1細胞於3.5 cm 培養皿,置於37℃、5% CO2 之培養箱內培養,待細胞長滿後即可移至6 well培養皿進行分化和並抽取RNA和 蛋白質以做鑑定。. 36.
(47) 第三章 材料與方法. 九、RNA萃取分離--使用RNeasy® Mini Kit (QIAGEN) 資料來源:76 (一) 原理 RNA抽取原理,是以Chomczynski和Sacchi 之單一步驟抽取RNA方法修改 而成,以高濃度之guanidine salts及urea等變性劑,使細胞溶解,並破壞胞器所釋 放之RNase的活性,所得之細胞溶解液再以酸性之phenol/chloroform萃取,分成 兩層,DNA及蛋白質會在有機層或介面層,而RNA在水層,如此可有效分離出 RNA。將萃取得之total RNA以isopropanol沉澱,並溶於RNase-free的水中即可。. (二) 實驗步驟 細胞先用PBS清洗一次後,加入1 ml TRIzol® ,以pipet 來回吸放數次,將細 胞沖下並打破,將TRIzol® 細胞液移至15ml離心管,放置-80℃冰箱待後面分析使 用,自-80℃冰箱拿出後,立即快速將細胞液37℃水浴回溫,加入400μl chloroform 震盪均勻後,放置室溫2-3分鐘,使蛋白質變性,再以12000 g、4℃離心15分鐘, 取上層水層至另一eppendorf,在加入600 μl 70% DEPC-Alcohol後混勻,使RNA 沉澱,取700 μl 至RNeasy spin column(放在2 ml collection tube 上) 後離心15 秒 (10000 rpm),將下層的液體倒掉後,再將剩下的700 μl 至RNeasy spin column 後離心15 秒 (10000 rpm),將下層的液體倒掉,加入350 μl RW1,離 心15 秒 (10000 rpm) 後將下層的液體倒掉,加入10 μl DNase I stock solution至 70 μl Buffer RDD(Qiagen supplied with RNase-free DNase set)至RNeasy spin column膜上,放置室溫15分鐘,再加入350 μl RW1離心15 秒 (10000 rpm),加 入500μl RPE,離心15 秒 (10000 rpm),將下層液體倒掉後,再加入一次RPE, 離心2分鐘 (10000 rpm),將column 放到空的2 ml collection tube 裡,離心1 分鐘 (13000rpm),接著換到含有SUPERase-In(RNase inhibitor) 1.5 ml collection tube, 加入30 μl RNase-free water,於室溫下靜置1分鐘,離心1 分鐘(10000 rpm),再. 37.
(48) 第三章 材料與方法. 加入一次20μl RNase-free water,室溫下靜置1分鐘並離心1 分鐘 (10000 rpm) 後,即可將RNA 放到-80℃冰箱中保存。取2 μl 的RNA以Thermo Scientific NanoDrop 1000 Spectrophotometer測A260/A280的比值及RNA濃度,RNA溶液在 260 nm下吸光值為1.0時濃度相當於40 μg/ml,A260/A280的比值為1.8-2.0為 佳。. 十、RT-PCR (Reverse Transcriptase PCR)--使用SuperScriptTMⅢ One-Step RT-PCR System with Platinum® Taq DNA Polymerase Kit (Invitrogen) 資料來源:77 (一) 原理 RT-PCR (Reverse Transcriptase PCR,反轉錄酶PCR),其原理是將一段待測 的RNA序列經反轉錄酶的作用轉錄成cDNA,再利用PCR技術將基因片段以幾何 級數倍增的方式增加到數十萬倍,所形成的PCR基因產物經一種叫 Ethidium bromide 的化學物質作用,利用其會與DNA嵌合,經紫外燈照射時會發出肉眼可 見的螢光,即在電泳膠片上會呈現具有特定分子量的PCR基因片段的產物。. (二) 實驗步驟 Reaction set up: Component. Volume 12.5 μl. 2x Reaction Mix Template RNA (1 μg). X μl. Sense primer (10 μM). 0.5 μl. Anti-sense primer (10 μM). 0.5 μl. SuperScriptTM III RT/Platinum® Taq Mix. 0.5 μl to 25 μl. DEPC-water. 38.
(49) 第三章 材料與方法. Reaction protocol: Temperature. Time. Cycle. 30 min. 1. 2 min. 1. cDNA synthesis 55℃ Denaturation 94℃ PCR amplification 94℃. 15 s. 55℃. 40 s. 68℃. 45 s. 40. Final extension 68℃. 5 min. 4℃. ∞. 1. 將 RNA、primer 與試劑混合後,放入PCR 專用之小管中,放入iCycler, 設定上述之反應條件即可。反應完成後,每管加入6 倍之DNA loading dye,再 以含有0.26%之EtBr 之1% Agarose 跑膠,並以MultilmageTM Light Cabinet 分析影 像並儲存。. 39.
(50) 第三章 材料與方法. The primer sequence and products size for PCR experiment mRNA 18S. Primer sequence. length (bp). Forward 5’-GGGAGCCTGAGAAACGGC-3’ Reverse. HBP1. 5’-AATCCTTGGCCCTCTGAGAT-3’. 154bp. 5’- GGCTCGGGCAGCTGCTT-3’ 87bp. 5’- TTACTCCACTGCCCACCTGT-3’. Forward 5’-AAGACAGCTCCTCCTCGAAGGTT-3’ Reverse. 225 bp. 5’- GCCATGGCCTTGACCAAGGAG-3’. Forward 5’- AAAGTGCAGGCTTGTGGACT-3’ Reverse. aP2. 198 bp. Forward 5’- GCAAGAGCCGCGACAAG-3’ Reverse. C/EBPδ. 5’-AGCAGTGCCAAATGGTGGAT-3’. Forward 5’- GAACAGCAACGAGTACCGGGTA-3’ Reverse. C/EBPβ. 501bp. Forward 5’-TTTTCAAGGGTGCCAGTTTC-3’ Reverse. CEBP/α. 5’-CCGCTCCCAAGATCCAACTAC-3’. Forward 5’-ACTCCTGTGATGAGCACATGGA-3’ Reverse. PPAR-γ. 262bp. 183 bp. 5’-GCGTAAATGGGGATTTGGTCACCA-3’. 十一、Quantiative -PCR(Quantiative Reverse Transcription-PCR)使用Taq Man® One-Step RT-PCR Master Mix Reagents (Roche) 資料來源:78 (一) 原理 Quantiative RT-PCR(即時螢光定量 PCR) 其原理為在 PCR 反應系統中,加入 螢光基團,利用螢光信號積累即時監測整個 PCR 過程,最後以標準曲線對未知 的樣品進行定量分析的方法。Ct 值的含義是 C 為 Cycle,t 代表 threshold,每個 40.
(51) 第三章 材料與方法. 反應管內的螢光信號到達設定的閾值時所經歷的 cycle 數,因此,即時螢光定量 PCR 是一種採用外標準曲線定量的方法。. (二) 實驗步驟 Reaction set up: Component. Volume. TaqMan® 2X Universal PCR Master Mix. 20 μl. 40X MultiScribeTM and RNase Inhibitor Mix. 1 μl 1.6 μl. 25X primer&probe. X μl. RNA (60 ng). to 40μl. DEPC-H2O. 將 RNA、primer 與試劑混合後,放入 real time PCR 96 well 專用之小管中, 放入 Applied Biosystems PRISM® 7900HT,上機,設定所需之反應條件即可。. 十二、蛋白質萃取. (一) 原理 利用細胞刮除法使細胞破裂並使用 RIPA 裂解液讓細胞快速裂解,使蛋白 溶解變性並抑制蛋白酶的活性使蛋白穩定,再利用超高速離心,取上清液蒐集總 蛋白。. (二) 實驗步驟 藥品配置. 41.
(52) 第三章 材料與方法. RIPA buffer: Component. Concentration. NaCl. 150mM. Tris buffer. 10mM. EDTA. 5mM. SDS. 0.1%. Triton X-100. 1%. Deoxycholate. 1%. 細胞以 PBS 清洗過後,加入適量的 RIPA buffer (含 1:100 protease inhibitors cocktail 與 phosphatase inhibitors cocktail) , 以 細 胞 刮 杓 將 細 胞 刮 下 , 放 至 eppendorf,置於冰上 30 分鐘,將細胞打破,以 11800 rpm 轉速離心 30 分鐘,取 上清液,放到-80℃冰箱中保存備用。. 十三、蛋白質定量 資料來源:79 (一) 原理 蛋白質染劑可與蛋白質結合形成藍色複合物,並可於 595nm 測得此複合物 之吸光值。藉由 BSA 標準品所形成的定量標準曲線,即可換算出待測樣品的蛋 白質濃度。. (二) 實驗步驟 藥品配置. 42.
(53) 第三章 材料與方法. Standard: 二次水. Concentration. BSA. 0 μg/ml. 0 μl. 800 μl. 200 μl. 2 μg/ml. 20 μl. 780 μl. 200 μl. 4 μg/ml. 40 μl. 760 μl. 200 μl. 6 μg/ml. 60 μl. 740 μl. 200 μl. 8 μg/ml. 80 μl. 720 μl. 200 μl. 10 μg/ml. 100 μl. 700 μl. 200 μl. total volume. protein assay dye(BIO-RAD). 1 ml. 標準品 BSA 以上述濃度作標準曲線,加入 200 μl protein assay dye (BIO-RAD),以達總體積 1 ml 震盪均勻靜置 5 分鐘,以 595 nm 波長測其吸光值 以 595 nm 的吸光值,作成標準線,分析最佳標準迴歸直線,求出此線的方程式(此 直線的迴歸分析結果之 R2>0.998 時,此直線方程式的值使可相信)。將樣品取 1 μl 的待測蛋白質溶液加入 799μl 二次水,再加入 200 μl protein assay dye (BIO-RAD),震盪均勻靜置 5 分鐘,測其吸光值再以樣品所測得 OD 值之結果, 帶入方程式即可求得蛋白質濃度。. 十四、Western blot. (一) 原理 西方墨點法是利用抗體和抗原專一性地結合,去標定電泳膠上的特定蛋白質, 而探針是一級抗體(Primary antibody),顯色用標記的二級抗體(Secondary antibody)。SDS 是介面活性劑,可使蛋白質變性,在分子表面均勻佈上一層負 電荷,利用由稀到濃的膠體配合梯度電泳,分離蛋白質,分子量越小的就跑的越. 43.
(54) 第三章 材料與方法. 下面,經過 SDS-PAGE 分離的蛋白質樣品,轉移到固相載體(PVDF),固相載體 以非共價鍵形式吸附蛋白質,保持電泳分離的多胜類型及生物活性不變,以固相 載體上的蛋白質作為抗原,與對應的一級抗體發生免疫反應,再與酶或同位素標 記的二級抗體反應,最後經過顯色或顯影檢測電泳分離的蛋白成分表現。. (二) 實驗步驟 藥品配置 4X Lower Tris: Component. Concentration. Tris-base (1.5M). 36.34 g. SDS (0.4%). 0.8 g. ddH2O. X ml. pH. 8.8. total volume. 200 ml. 4X Upper Tris: Component. Concentration. Tris-base (0.5 M). 12.11 g. SDS(0.4 %). 0.8 g. ddH2O. X ml. pH. 6.8. total volume. 200 ml. 44.
(55) 第三章 材料與方法. APS (0.1g/ ml) Component. Concentration. Ammonium persulfate (10 %). 1g. ddH2O. X ml. total volume. 10 ml. Running and stacking Stock solutions. 8%Resolving gel. 40%Acrylamide. 10%Resolving gel. 1.6 ml. 4%stacking gel. 2 ml. 0.39 ml. Upper Tris(4X). 1 ml. Lower Tris(4X). 2.00 ml. 2 ml. Water. 4.4 ml. 4 ml. 2.6 ml. APS (10%). 40 μl. 40 μl. 40 μl. TEMED. 4 μl. 4 μl. 4 μl. total volume. 8.044 ml. 跑膠(SDS-PAGE) 10X Running buffer: Component. Concentration. Tris-base (0.25 M). 30.3 g. Glycine (1.9 M). 144 g. SDS (1 %). 10 g. ddH2O. X ml. pH. 8.6. total volume. 1000 ml 45.
(56) 第三章 材料與方法. 轉印 10X transfer buffer: Component. Concentration. Tris-base (0.25 M). 30 g. Glycine (1.9 M). 144 g. ddH2O. X ml. pH. 8.3. total volume. 1000 ml. 免疫染色 10X Tris-base buffer saline: Component. Concentration. Tris-base (0.2 M). 0.2 M. Glycine (1.4 M). 80 g. ddH2O. X ml. pH. 7.6. total volume. 1000 ml. TBS/T: Component. Concentration. 10X TBS. 100 ml. Tween 20 (0.1%). 1ml. ddH2O. 900 ml. total volume. 1000ml. 46.
(57) 第三章 材料與方法. 實驗步驟 膠體的配製依蛋白質分子量的大小而決定配製成 8、10 % 的 SDS-PAGE。 待鑄好的 SDS-PAGE 組合放置電泳槽中,倒入適量的 1× running buffer。將製備 好的蛋白樣品以及標準品小心加入於固定於垂直電泳槽內的 stacking gel 的 well 內,以避免 sample 溢出,先以 90 伏特電壓跑 4 % stacking gel,20 分鐘後,調 整電壓至 100 - 120 伏特,約跑 2 – 3 小時,即完成蛋白分離。 轉印時,先將 PVDF 膜與 3M 濾紙裁剪與膠片大小相同。將轉漬夾打開後, 黑色面朝下,依序放上海棉墊、濾紙、SDS-PAGE gel、PVDF 膜 (先用 100% 甲 醇濕潤 2 分鐘、再放於 transfer buffer 中浸潤)、濾紙,最後放上一片海棉墊,確 定其緊密貼合後,將轉印夾慢慢地合起來,放到轉印槽中,倒入 1× transfer buffer, 以 400 mA 電流轉印 1 - 2 小時,使蛋白質轉印到 PVDF 膜上。 轉印完成的 PVDF 膜以 5% 脫脂牛奶 (以 TBS/T 配製) 搖晃 1 小時進行 blocking 後,以 TBS/T 清洗 5 分鐘 3 次後,加入適當稀釋倍數之一級抗體反應 overnight。將一級抗體以 TBS/T 清洗 5 分鐘 3 次後,依不同的一級抗體,加 入特異作用的二級抗體反應 1 小時,再以 TBS/T 清洗 5 分鐘 3 次,以感光試劑 Western lightening chemiluminescence reagent plus 進行呈色反應,利用 LAS - 4000 mini FUJIFILM 冷光影像系統分析。. 十五、流式細胞儀 資料來源:28 (一) 原理 細胞週期大致可分為四個不同時期,G1 phase、S phase、G2 phase 和 M phase。 每個時期的 DNA 含量會因為功能作用不同而有所差異,一般來說,G2 期為 G1 期的兩倍,則 S 期則基於兩者之間。因此可以利用細胞中的 DNA 含量來決定週 期,利用螢光物質來標示 DNA,之後再利用 FACS(fluorescence-activator cell sorter). 47.
(58) 第三章 材料與方法. 進行 DNA 含量分析。. (二) 實驗步驟 試劑配置. Component Propidium iodide TritonX-100 RNase A PBS. Stock solution. Final solution. volume. 100 μg/ml. 50 μg/ml. 250 μl. 10 %. 1%. 50 μl. 100 mg/ml. 100 μg /ml. 0.5 μl. -. -. 199.5 μl 500 μl. total volume. 固定細胞 移去培養基,以 PBS 細胞清洗一次,Trypsin 打下來,加入 5 ml 細胞培養液 將細胞收集在 15 ml 離心管中離心,1500 rpm,10min,倒掉上清液,將細胞完 全彈散後加入 10ml PBS 清洗,離心 1500rpm,10min,倒掉上清液,將細胞完全 彈散,加入 3ml 冰的 90% Methanol,進行細胞固定(以 Shake 3 速度震盪,將酒 精緩慢滴入),固定完成後放置-20℃冰箱。. 流式細胞儀分析 隔天,將 sample 從冰箱取出後離心,1500rpm,10min,去除上清液,不用彈 散(因為用酒精固定後細胞很脆弱) ,加入 10ml PBS 清洗離心(延管壁緩緩加入), 1500 rpm,10min,去除上清液,將細胞完全彈散,在每個離心管中加入 300~500μl 的 PI 染劑(視細胞數目增減),用 1ml pipette 在 15ml 離心管中抽吸數次,將細胞 用絹布製的篩子過濾到 FAS 管中,避光至於冰上反應 30min,以流式細胞儀進行 48.
(59) 第三章 材料與方法. 樣品分析(每秒不超過 300 顆細胞,每個數據收集 10000 顆細胞),數據以 Modfit LT 軟體進行分析。. 十六、統計分析 實驗結果均以平均值 ± 標準差 (Mean ± SD) 表示,利用 Student’s t test 或 Duncan’s multiple range test 比較組間差異。所有數據通過常態分佈檢定,否則 轉型為對數值 (Log)。統計分析以 SAS 套裝軟體 ( version 9.0, SAS institute, Cary, NC ) 分析,統計比較以 p< 0.05 為具有顯著之差異。. 49.
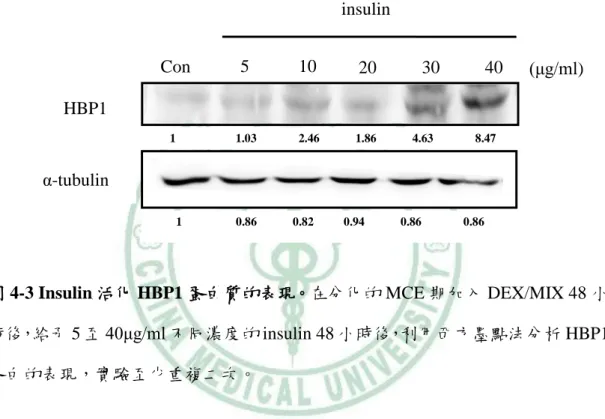
(60) 第四章 結果. 第四章、結果. 一、HBP1 在脂肪細胞分化過程的表現. (一) 脂肪細胞分化後期 HBP1 表現明顯增加 典型的 in vitro 脂肪分化模式中,前脂肪細胞株如 3T3-L1、Ob1771、 3T3-F422A 和初代脂肪細胞為常用的細胞,其中,3T3-L1 在 1974 年由 Green 和 Kehinde 由 Swiss mouse embryo cloning 出來,具高度分化為成熟脂肪細胞潛力的 前脂肪細胞株,當以 DEX/MIX 協同 Insulin 誘導時,3T3-L1 將分化為成熟脂肪 細胞,也是我們此次研究所選用的實驗細胞模式。 3T3-L1 細胞分化的特徵包括:1) 細胞型態由紡錘型變為球形,2) 細胞內 TG 的堆積,油紅染色可染出脂肪細胞內的油滴、3) 脂肪細胞分化特定基因的表 現增加,例如 PPAR γ、C/EBP α和 aP2 等。這些改變將是我們用來檢測脂肪 細胞分化重要的指標。 為了觀察 HBP1 在脂肪細胞分化過程中所扮演的角色,首先,我們檢視 HBP1 的表現是否隨著分化過程的進行而有所改變。當我們依照標準分化流程以 MDI 刺激 3T3-L1 前脂肪細胞後、於分化第 0、2、4、6 和 8 天分別收集其蛋白 和 total RNA,並以 western blot 和 RT-PCR 觀察 HBP1 變化。圖 4-1 和 4-2 指出, 在 3T3-L1 前脂肪細胞分化為成熟脂肪細胞的 8 天過程中,HBP1 蛋白的表現在 分化初期 2 天無明顯改變,但是,在第 4 到第 6 天,HBP1 的表現明顯增加。結 果顯示,HBP1 在分化過程前期的 MCE 期無增加現象,一直到分化後期,HBP1 的表現才增加,表示著,HBP1 在脂肪分化過程的不同階段可能有不同的功能。. 50.
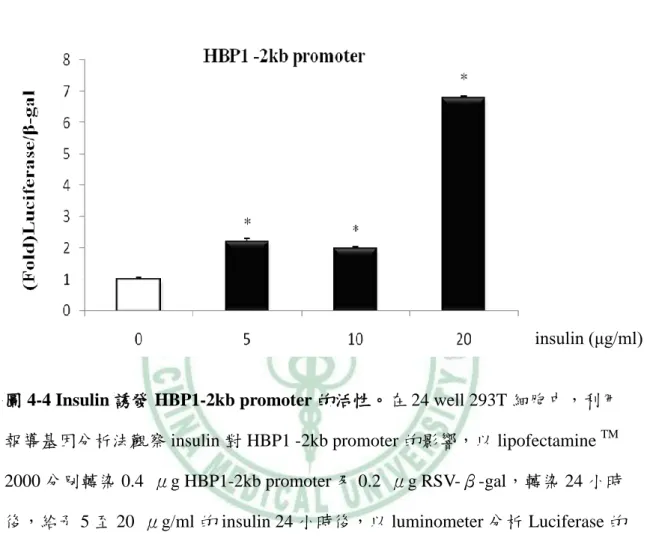
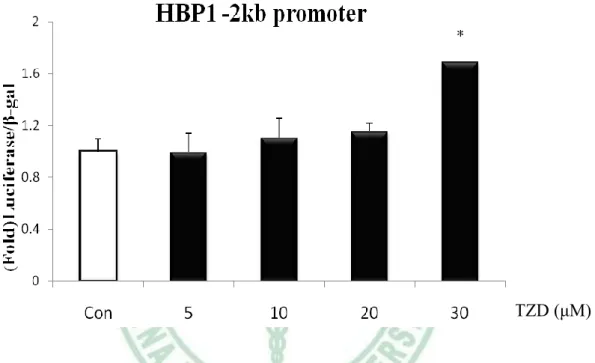
(61) 第四章 結果. (二) Insulin 誘導 HBP1 的表現 由於 HBP1 在分化第四天的表現量增加,加上之前的研究發現,在成熟的大 鼠脂肪細胞,insulin 增加了 HBP1mRNA 的表現,因此,我們假設,可能是 insulin 在分化後期誘導了 HBP1 表現。於是,我們在 3T3-L1 post-confluence 時,給予 DEX/MIX 二天後,分別給予 5 μg/ml、10 μg/ml、20 μg/ml、30 μg/ml 和 40 μg/ml 不同濃度的 insulin 48 小時後,以 Western blot 分析 HBP1 的蛋白表 現量。結果顯示,insulin 隨著濃度的增加,HBP1 蛋白的表現也隨之增加 (圖 4-3)。 此結果表示,insulin 具活化 HBP1 的能力,也可能是 HBP1 於 3T3-L1 分化後期 增加的主要原因。 由於 insulin 的給予增加了 HBP1 蛋白的表現,我們推測,insulin 可能藉由 活化 HBP1 的 promoter 增加 HBP1 的表現。因此,我們運用冷光酵素報導基因 分析法(Luciferase Gene Reporter Assay) 檢視其可能性,首先,我們轉染 0.4 μg 連接 HBP1 -2kb promoter 的 luciferase 基因的載體和 0.2 μg RSV-β-gal (internal control)質體至 24-well 的 HEK-293T 細胞,24 小時後,加入 0~20 μg/ml 不同 濃度 insulin,24 小時後,收集 cell lysate,以 luminometer 分析 luciferase 和β-gal 活性,圖 4-4 結果顯示,insulin 明顯誘導 HBP1 -2kb promoter 的活性。證實了我 們的假設;insulin 透過活化 HBP1 啟動子而增加 HBP1 蛋白的表現量。. 51.
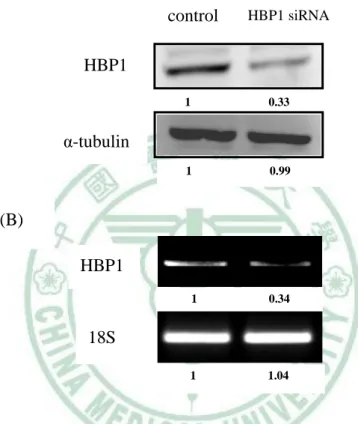
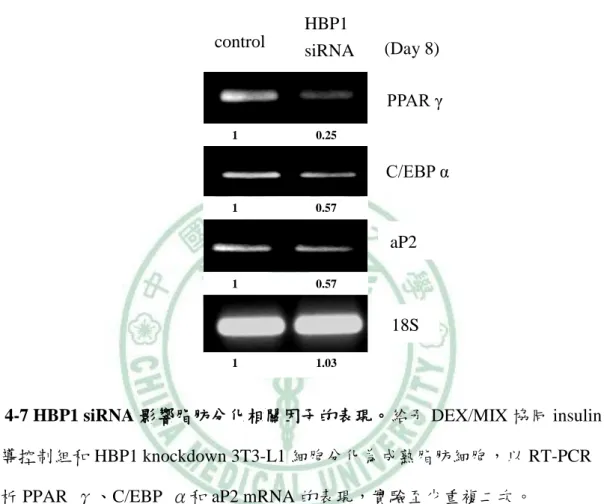
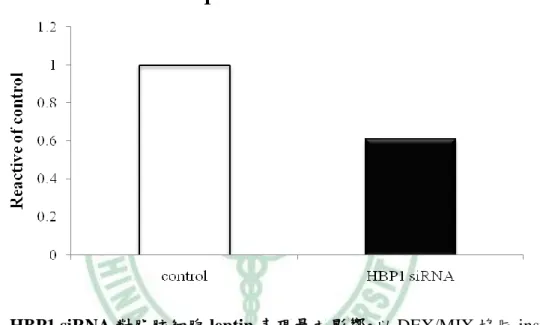
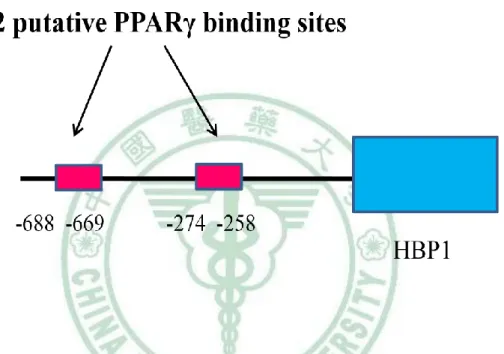
(62) 第四章 結果. 二、HBP1 的表現量影響脂肪細胞分化過程. (一) HBP1 reduction 造成脂肪細胞分化不完全 既然 HBP1 的含量隨著脂肪細胞分化的進行而有所改變,表示,HBP1 具調 控脂肪分化的潛在功能。為了驗證此假設,我們運用 RNA 干擾技術降低 3T3-L1 細胞內 HBP1 的含量,觀察脂質生合成是否受到影響。首先,我們利用 western blot 和 RT-PCR 確認 HBP1 專一的 siRNA 的確降解 3T3-L1 細胞中 HBP1 蛋白質和 mRNA 的表現(圖 4-5)。接著,我們分別給予 DEX/MIX 協同 Insulin 誘導控制組 和 HBP1 siRNA 3T3-L1 細胞分化為成熟脂肪細胞,於分化第八天,分別以 RT-PCR 分析 PPARγ、C/EBPα和 aP2 mRNA 的表現、 real time PCR 分析 leptin mRNA 含量和油紅染色觀察細胞內油滴的生成。圖 4-6 的結果為顯微鏡下觀察脂肪細胞 型態的改變情形;HBP1siRNA 組的細胞在分化實驗八天後,圓形型態細胞明顯 低於控制組,同樣地,圓形細胞內油滴的油紅染色,HBP1siRNA 細胞明顯低於 控制組。當我們進一步比較成熟脂質生合成的指標基因 PPARγ、C/EBPα和 aP2 的表現時,其 mRNA 的含量,HBP1 siRNA 組明顯低於控制組 (圖 4-7),leptin mRNA 的含量也具有相同的結果(圖 4-8)。此結果顯示,HBP1 表現量減少導致了 脂肪分化不完全,表示,HBP1 具調控脂質生合成的生理功能,但其調控機制需 進一步探討。. 52.
(63) 第四章 結果. (二) 於 MCE 期,HBP1 reduction 促進細胞週期進行和增加 CEBP/ β 和 CEBP/ δ 轉錄因子的表現 3T3-L1 脂肪細胞分化過程,依生理現象的不同,可簡單分為細胞繼續增生 的 MCE 期和細胞型態改變的分化後期。由以上結果得知,HBP1 的含量在這兩 期是不一樣的,表示,HBP1 在分化不同階段可能有不同的生理功能。雖然,我 們以 RNA 干擾技術降低細胞內 HBP1 的含量後,造成脂肪細胞分化的不完全, 但此結果並未解答 HBP1 在分化過程中的確實角色。為了進一步了解 HBP1 可能 的階段性生物功能,我們分別觀察它的表現量在 MCE 和分化後期的影響。 首先,我們觀察 HBP1 在 MCE 期的角色。有關 HBP1 生物功能探討的文獻 指出,HBP1 的主要生理角色是負責細胞週期 G1 期的調控;過多的 HBP1 表現 導致乳癌細胞週期停滯於 G1 期,而且,我們也發現,HBP1 的含量於 MCE 期( 0 到 2 天)並無增加的現象,所以,我們假設,HBP1 具調控 MCE 期細胞增生的功 能,換言之,HBP1 的表現量降低將使 MCE 期的細胞增生速度變快,伴隨著相 關基因的提早出現。結果如預期地,當我們給予 HBP1 siRNA 的 3T3-L1 細胞 MDI 分化劑 4 小時後,以流式細胞儀偵測細胞週期的進行,發現,HBP1 siRNA 組相較於控制組,明顯出現較多的 G2/M 期細胞分佈(圖 4-9),表示 HBP1 knockdown 造成較快的細胞週期進行,G2/M 期的較早出現。同樣地,HBP1 siRNA 也造成較高的 MCE 期相關分化基因 C/EBP β 和 C/EBP δ 表現(圖 4-10)。由此實 驗得知,HBP1 在 MCE 期的生理功能,如一般地,調控細胞週期和細胞增生。 值得探討的是,較快的 MCE 期細胞增生,應該有助 3T3-L1 脂肪細胞分化的完 成,但是,HBP1 knockdown 雖然出現 MCE 期細胞週期的促進,卻導致最終分 化的不完全。為追尋答案,我們進而探究 HBP1 在分化後期的角色。. 53.
數據

+7


相關文件
• Theorem: boosting tends to increase margins boosting tends to increase margins boosting tends to increase margins boosting tends to increase margins of training boosting tends
“ Numerical studies of hyperbolic IBVP with high-order finite difference operators satisfying a summation by parts rule.”.. “High-order finite difference methods,
• 接續之前的例子,若原為 0.288 pF 的液晶 電容 C LC ,再並聯一個亦為 0.288 pF 的電 容C st ,則電位保持的變化值為.
(Inheritance) ;c.動態繫合(Dynamic binding) ;d.動態範圍(Dynamic scoping).
(A)行百里者半於九十(B)步步要謹慎,因步步難行
Mass-to-light ratios of old stellar populations are uncertain; (1) the initial mass function (IMF) of star formation is poorly known; it may vary with radius in an individual galaxy
The Liouville CFT on C g,n describes the UV region of the gauge theory, and the Seiberg-Witten (Gaiotto) curve C SW is obtained as a ramified double cover of C g,n ... ...
Describe and analyze a recursive algorithm to reconstruct an arbitrary full binary tree, given its preorder and postorder node sequences as
