行政院國家科學委員會專題研究計畫 成果報告
國內鮮牛乳β-lactoglobulin 之大量損失對於生化及生理
功能之影響
計畫類別: 個別型計畫 計畫編號: NSC93-2313-B-009-002- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立交通大學生物科技學系(所) 計畫主持人: 毛仁淡 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 8 月 9 日
前言
牛乳whey protein (乳清蛋白,去除 casein 之部分) 中β-lactoglobulin (LG) 約佔總蛋白
質的10~15%,它是一個最容易被小腸所吸收之牛乳蛋白質 (1)。在西元 2000 前之 60 年間,
有文獻非常清楚的報導LG 生化特性 (2),但對其特定的生理功能卻不十分清楚 (3)。最近
的結果顯示其生理功能可以降低膽固醇 (4),2003 年 review 報告 (5) 指出絕大部分之乳蛋 白皆有抗菌之功用。本年度 2003 更指出乳清蛋白 (whey protein) 中之 cystein- rich 蛋白 (LG 即是其中之一) 與細胞中之 glutathione (GSH) 新陳代謝機制略同,除了具有抗氧化功 能之外,並可在動物中能誘導p53 及阻礙新血管形成 (neoangiogenesis),所以有抗癌作用 (6)。LG 是一個很特殊的乳蛋白,可以抗酸及抗蛋白酵素之攻擊並且可直接經由腸胃道吸 收進入人體中。初生嬰兒在斷奶後服用牛乳,可在血液中找到約μg/L level 的 LG (7)。與 牛不同的是在人乳中並不存在 LG,但母親哺乳期間飲用牛乳,在其母乳中也會發現 LG (8)。如上所述 LG 可抗氧化及抗癌又能直接被人體所吸收,並提供嬰兒快速成長所需的胺 基酸 (9, 10). 因此牛乳中之 LG 成分具有特殊的重要性。我們的實驗室過去兩年成果中指 出 (11, 12, 13.),LG 是一個高效能的抗氧化分子,其抗氧化機制在於第 121 胺基酸序列位 置中cys 的 free-SH。其扮演一個極重要的角色 (LG 有五個 cys,其他四個分別在 66-160 及106-119 位置形成雙硫鍵鍵結)。利用化學修飾 (chemical modification) 將第 121 位置之 free-SH group 修飾後(carcoxylmethylation),其抗氧化的功能也隨之消失 (11)。我們另外在 一篇論文(13)中也同樣指出國內所有市售廠商的鮮乳中,80%之 LG 已經不存在(figure 2), 其主要的原因是國內鮮乳嚴重過度加熱 ,並且發現其機制是 free-SH group 早已與其他蛋 白質鍵結形成高分子聚合物 (12)。但若將-SH 先 block 則 LG 即不能與其他的蛋白質鍵結 或自行形成polymers,但有趣的結果是所測的美國鮮乳中 LG 卻極完整 (100%)。 至於如何去檢測thermal denatured 之 LG,我們也在這兩年中 (2001-2002)發展出超快 速五分鐘晶片檢測法及ELISA,在 900 個奶粉單株抗體中,我們篩選到 4 株只能辨認奶粉 不能辨認生乳之專一性抗體。此4 株體可以區分奶粉與鮮奶,在實驗中若將生乳摻雜奶粉 即可立即用抗體晶片發現 (如 figure 25),並且利用光電效果將其數位化後可以定量。此法 靈敏度高只需要10nL (1*10-9L) 的乳品即可檢測。在鮮乳加工廠中可在加熱滅菌前先行檢
驗即可得之有無摻雜奶粉,另外100%生乳若以 UHT 滅菌處理或處理時間過長 (如保久乳 或市面之”鮮乳”), LG 受到嚴重破壞也可以利用我們的快速檢測法測出。本實驗已清楚的 指出國內市售鮮乳中之品質問題。當然未做田野實驗研究 (field study),我們無法預設立 場,究竟是國內市場的鮮乳過度加熱還是摻雜奶粉?雖然我們無法在此時下定論,但是國 內市售鮮乳的確有改善的空間。 在過去兩年度我們發現國內所有市售奶粉中之LG 在加工過程中 80%以上遭到嚴重破 壞,因為LG 已與鮮乳中之 albumin,lactoalbumin 及 casein 結合成大分子聚合物。由於 LG 可直接在腸胃道中被吸收進入循環系統 (14,15,16),到底這些大分子聚合物是否可以被直 接吸收? 以及 LG 被吸收後具有何種生理功能(如生長,過敏及抗氧化),因此本計畫研究 目標如下。 研究目標: 1. 在實驗室中利用不同溫度及時間加熱處理 LG,並決定其結合之形成及利用生化方法 分析LG 與何種蛋白結合。 2. 以小鼠為動物模式比較國內市售牛乳加工前 (LG 未結合其他蛋白) 及加工後 (LG 結 合其他蛋白) 之 LG 在血中之吸收量。完成後再與美國市售鮮乳比較,並同時進行兩 種鮮乳對小鼠成長速率之影響分析。 3. 比較加熱 (LG 破壞) 及未加熱 (LG 未破壞)之牛乳對小鼠之過敏反應,以血液短暫期 (transient) 之 Ig E 為指標 (因為加熱後會擴大 cross-linking 之大分子,胃腸無法吸收)。 4. 決定 LG 在生乳中之抗氧化能力並定出其比例。同時進行調查市售鮮乳在 LG 損壞下 之總抗氧化力。 5. 宣導國內乳品加工方式之改善。
中文摘要 β-lactoglobulin (LG) 是牛乳中主要蛋白之一,占全部蛋白的 10-15%。實際上,在 人體中LG 可在腸胃道被吸收。雖然 LG 的生化特性已被研究地相當透徹,但其生理功 能目前仍在部分瞭解的階段。最近(2000-2003),LG 被發現有降低膽固醇及防癌的功 效。過去2 年間,我們的實驗結果發現在台灣市售的鮮奶中,LG 的含量幾乎完全地消 失,推測有 2 種可能性,第一、在加工的殺菌過程中,生乳被過度加熱;第二、市售 鮮乳中摻雜有還原乳。為了辨識奶粉與鮮奶中 LG 的差異性,我們發展出可以區分生 乳與還原乳的專一性單株抗體,利用抗體晶片的技術,我們可以在 5 分鐘內判別鮮乳 中是否有添加奶粉。目前我們正深入研究其高專一性辨別能力的作用機制,及製造過 程中LG 在生化特性及結構上的改變。基本上,主要是因素為 LG 與牛乳中其他的蛋白 質產生了相互結合的現象,如lactoalbumin、免疫球蛋白、albumin 以及 casein 等。 由於LG 也會藉由自身共價鍵結的方式,形成多單元結構,而在台灣 80%以上的 市售鮮乳中LG 結構上完全改變,目前也尚無法瞭解其改變是否會影響 LG 生理角色的 影響。值得注意的是牛奶已逐漸成為國人飲食中,胺基酸的主要攝取來源之一,因此 本研究也就益顯重要。
英文摘要
β-lactoglobulin (LG) is a major milk whey protein containing about 10-15% of total.
Essentially, in the gastro-intestinal tract, LG as an intact form can be absorbed directly into
the circulation in humans. Although the biochemical properties of LG have studied in
details prior to year 2000, the physiological functions have been partially elucidated. More
recently (2000-2003), LG is found to be associated with hypocholesterol and anticancer
effects. In the past two years, our studies show that the LG of cow milk sold in the
this episode: First, the raw milk is overheated during the manufactured process for
sterilization. Second, the fresh milk on the market might be containing some powdered
milk. To detect the deficient of LG or freshly milk containing the powdered form of milk,
we have recently developed a specific monoclonal antibody that can distinguish between the
powdered and raw milk. Using the antibody chip technique, we can essentially detect the
dry milk in 5 minutes. We have also explored the mechanism involved in the specificity of
the monoclonal antibody and the biochemical changes of LG during the manufactured
process. Basically, it was due to the cross-linking of LG to the other milk protein fractions,
such as α-lactoalbumin, immunoglobulin, albumin, and casein.
Because of the covalently cross-linking into large polymers and the damage of LG (> 80%)
in the milk of our domestic market, it is not know whether or not such substantial loss of LG
may affect its physiological role of the total LG uptake of ingested milk. Particularly, milk
has become one of the major amino-acid supplements in our routine diet.
實驗方法
1. 在實驗室中利用不同溫度及時間加熱處理 LG,並決定其結合之形成及利用生化方法 分析 LG 與何種蛋白結合。 在我們實驗室中之先導實驗及過去兩年之報告 (13) 清楚地證明國內市售鮮乳 LG 已經達到80%之破壞,這種 LG 破壞對於牛乳品質在生化及生理造成何種影響,其機制 為何? 所以本目標之一要先了解國內鮮乳 LG 破壞後與何種蛋白結合 (figure 2),並擬定 改善辦法,因為國外鮮乳並無此問題,所以在此方面研究極少,而絕大部分國外研究室是將純化後之 LG 先加熱後再研究,我們最近一年研究發現國外直接加熱法與利用牛乳
直接加溫所得之結果差異極大。第一:純化後之LG 加溫後只能自行形成 dimer ,trimer
及polymer。第二:牛乳直接加溫,LG 可與乳蛋白交互作用形成其他之結合體。大部分這
種結合物 (crossly linking products) 分子量極大,超過 300,000~500,000 dotons,在 PAGE 中以commassie blue 染色無法觀察,只能以 Westernblot 找到 LG 之部分(figure 4)。最近 一 年 我 們 已 發 展 一 種 特 殊 形 式 之 2D gel electrophoresis , 即 每 一 dimension 皆 用 SDS-PAGE,但第二次 dimension 中加入 reducing reagent :mercaptoethanol,如此 LG 與 其 他 在 雙 硫 鍵 鍵 結 下 的 結 合 蛋 白 即 可 輕 易 的 觀 察 到 (figure 15),也就是在 linear regression curve 中 45。
角度之內的蛋白,從figure 15 可發現鮮乳中有 lactoalbumin,albumin (極少),及 casein 為主要結合蛋白。我們預測其結合形成絕對不是一對一形式 ( 1:1 molar ratio),而是一個結合蛋白帶有極多之 LG 分子。實驗設計上:
(1) 首先將國內市售之乳蛋白通過 Gel-filtration HPLC column,再以 ELISA 方式決定 LG 之聚合物分布情形。 (2) 市售乳蛋白經過 mercaptoethanol 處理打斷其雙硫鍵鍵結再經過 HPLC,此時 LG 在理 論上應該全部elute 至 18K 的位置。 (3) 計算 LG 在每一高分子 fraction 之含量,求出其分子比例 (stochiometry)。 (4) 所有上述實驗再與生乳做比較統計,並純化及定性 LG 結合之蛋白,而定性可用 Westerm blot 及胺基酸定序來決定。 本實驗之可行性:由於最近兩年本實驗室有博士班學生兩名及碩士班學生兩名同時 進行牛乳之生化及免疫化學研究,所有工具如 LG 單株抗體及多株抗體之製作,ELISA 定量,HPLC 分析皆有相當成熟的經驗,對 LG 之分子結構及物理形狀 (conformation) 在 雙極光譜 (circular dichroic spectrum) 之改變及其 free-SH 之生化角色皆十分了解 (13), 對此項目執行將可順利完成。
2. 以小鼠為動物模式比較國內市售牛乳加工前 (LG 未結合其他蛋白) 及加工後 (LG 結
種鮮乳對小鼠成長速率之影響分析。 如上述(1)所陳述,我們認為 LG 加熱破壞後之 cross-linking 大分子可能會影響腸胃 道 (gastrointestinal) 之吸收能力。主要原因是大分子之 permeability 較弱,為了要證明此 假說,我們將以小鼠 (balb/c) 進行 in vivo 動物模式實驗。初步構想及設計如下: (1) 將 balb/c 分為三組﹔分別為對照組 (n=12),餵食市售鮮乳組 (n= 20)及生乳組 (n= 20)。餵食方式可以強迫餵食 (一般 gavage) 或摻雜在供水瓶中,為防止水中牛奶酸 敗,我們將採用前者方式。 (2) 在不同時間採血並利用 ELISA 及本實驗室最近研發之抗體晶片測定 Balb/c 血中 LG 之含量。文獻報導在人類母親實驗中以口服牛奶,只需要在1~2 小時後 LG 即可在母 奶中找到(8.),估計小鼠血液中之 pharmacokinetic 應該有類似人類之結果。小鼠將用 capillary tube 在眼角進行採血,本實驗室一次可採到 50μL,而只需要用 10 nL 在晶 片上即可定性定量。因在嬰兒採血實驗中約有 40%在血中有 LG 表現(7)。所以本實 驗在老鼠數目上可能依形況需要而增加。另外我們在餵食時間上要執行至血液中LG 量達到steady state 為主。 (3) 統計分析上,以上結果將採用 student t-test 分析,並以 p<0.05 為分界點。 (4) 若兩組有顯著差異可以證明加熱後之 LG 會結合大分子有礙腸胃道之 permeability 及 吸收,則國內市售鮮乳在加溫滅菌上應予改進。 (5) 若本實驗在(4)中證實,我們將繼續在實驗室中以生乳做不同溫度處理。首先以生化 方法定出加溫及時間變數上對大分子形成之 break-point。我們估計約在 80℃加熱 15 分鐘,LG 即可達到 50%之破壞率,再以同法與 in vivo 之 bioavailability 進行比較來 證明何種溫度加熱滅菌不會破壞腸胃道吸收率。本結果將提供國內乳業界參考。 (6) 本特定目標將繼續進行 in vivo 生理功能實驗,我們將以美國市售鮮乳 (具有 100%完 整LG) 及國內市售鮮乳 (僅具有 20%完整 LG) 進行小鼠試驗。同樣實驗與 (1) 一樣 分成三組,在母鼠尚未懷孕前進行餵食然後觀察懷孕生產後嬰鼠 (infant mice) 的生 長速率。由於在斷奶 (weaning) 前,嬰鼠處於母親保護期,所以生長速率難測,所以 在斷奶後再繼續檢測其生長,估計整個實驗將進行兩個月。
3. 比較加熱 (LG 破壞) 及未加熱 (LG 未破壞)之牛乳對小鼠之過敏反應,以血液短暫期 (transient) 之 Ig E 為指標 (因為加熱後會擴大 cross-linking 之大分子,胃腸無法吸
收)。
本實驗室已在進行小鼠對 LG 之過敏反應進行初步實驗,目前可以觀察到 LG 在經
過腹腔注射後可產生對LG 具有專一性的 Ig E,如 Fig 7 dot blot 的陽性表現,正常小鼠 血中成陰性反應。據文獻報告(23) 指出某些牛乳飲用者在免疫反應失調下,腸胃道可啟 發免疫反應而產生 Ig E,並暗示 (implicate) LG 為主要禍首 (24,.25)。當然其他如 lactalbumin 及 casein 也參予部分過敏反應(26,27,28,29.)。在人體實驗中也有報導加熱過 後之牛奶似乎可以降低免疫反應 (30,31)。所以國內市售鮮乳雖然 LG 已被破壞 (80%), 但對牛奶過敏之使用者可能有正面之影響。日常生活中也顯示飲用鮮乳過敏者,在食用 其他乳製品時並未發現有過敏反應。我們的假說是乳製品中之 LG 可能已經被完全破 壞,即LG 與其他乳蛋白 100%形成高分子,這種 LG 高分子無法透過腸胃道直接吸收而 降低其免疫反應。 本實驗之設計為: (1) 在特定目標 (2) 中的小鼠血液中除了測 LG 血中濃度並進行 LG 之 Ig E 抗體進行濃度 測定,其測定可使用本實驗室之抗體晶片。此法可測出小鼠在國內外鮮乳中LG 在血 中吸收率對 Ig E 是否有正向關係 (postive correlation)另外之過敏反應測定法將依 standard protocol (32,33) (2) 四周大的 BALB/C 小鼠將購置於國家實驗動物中心。購得後先妥善安置於動物房兩 週,待其六週大時才進行致敏實驗,其間飲食禁絕一切含奶類製品。實驗共分三組皆 進行腹腔注射,第一組為對照組完全不施打任何藥劑;第二組採用 5 µg 的 BLG 以
1:1 的比例跟佐劑 IFA(Incomplete Freund’s Adjuvant)充分混合,注射的總體積為 0.2 mL;第三組只施打含 IFA 藥品作為對照;第四組採用 10 µg 的 OVA 以 1:1 的比例跟 佐劑IFA(Incomplete Freund’s Adjuvant)充分混合,注射的總體積為 0.2 mL。 (3) 同樣實驗將用純化後的 LG 進行,由於人與小鼠在基因數目上只差約 300 個 (34),同
過 敏 者 可 在 日 常 生 活 中 飲 用 即 高 溫 處 理 的 保 久 乳 作 為 胺 基 酸 攝 取 之 平 衡 飲 食 (balance diet)。
(4) 如果上述實驗順利,我們將利用國內保久乳進行過敏反應測定。
4. 決定 LG 在生乳中之抗氧化能力並定出其比例。同時進行調查市售鮮乳在 LG 損壞下
之總抗氧化力。
在胃潰瘍(stomach ulcer)病患中,由於胃細胞中之自由基可刺激 lesion 及免疫反應 (35),文獻中亦指出(36),潰瘍之大小與自由基的多寡有正向關係,服用抗潰瘍藥物 lansoprazole 再利用 NMR 方法證實其機制為除去自由基(scavenging free radicals)。在酒精 誘導潰瘍之大鼠動物模式中也發現抗氧化劑quercetin 可以降低其 mast cells 及潰瘍之區 域 (37) ,至於牛奶在治療或防止潰瘍已行之有年 (38) ,在對 aspirin 有反應之 ulcer 之 病患中也建議服用 aspirin 之同時喝牛乳,但在 ulcer 之動物模式中來證實牛乳是何種成 分保護 ulcer 則非常不清楚,建議是脂肪或蛋白質為保護機制者皆有 (39,40,41,42) ,在 這方面研究非常稀少﹔估計原因是以牛乳為基礎發展藥物獲利不高。最近報導牛乳中之 α-lactoalbumin 可以保護酒精及壓力所引起之潰瘍 (42)。另外也有報導牛乳中具有-SH 鍵之蛋白有很強的保護作用 (43) ,其原因如上所述-SH 蛋白與 glutathione 略同是一抗氧 化分子。我們在先導實驗及過去兩年中也證實 LG 是一個有效的抗氧化分子,但在過度 加熱氧化後或經過化學修飾則失去抗氧化能力。本特定目標要決定: 第一:LG 在牛奶中到底佔有多少之抗氧化能力。我們的方法大致如下: (1) 先大量製造 LG 之多株抗體 (form rabbits)
(2) 純化 anti-LG 之 IgG,再備製即一支 anti-LG affinity column
(3) 將牛乳通過此 affinity column,可將牛乳中之 LG 全部除掉,此時的牛乳稱為 LG-deficient milk
(4) 比較一般牛奶與去除 LG 牛奶之間抗氧化力的差別 第二: 我們要決定國內市售鮮乳加工前後之抗氧化能力 第三: 我們要決定國內市售及美國市售鮮乳之抗氧化能力
第四: 我們要決定市售鮮乳與市售奶粉之抗氧化能力 第五: 我們要決定市售鮮乳與市售奶粉之抗氧化能力
以上實驗因為各種待測之parameter 很多,如加熱時間,溫度等…,所以在實驗流程
尚為考慮各種因素。 抗氧化能力測試:
抗氧化實驗可用我們實驗過去報告之方法 (44, 45),以 LDL 為 lipid peroxide 之 substrate, 分別在其中加上不同量之抗氧化劑以 probucol 及 vitamin 為 internal standact。自由基之 initiatior 可用 Cu+2或AAPH (水溶性之 A2O-化合物) 來誘導 LDL 之氧化,抗氧化強度則 將以IC50 (inhibitory contraction at 50% )來表示。 5. 宣導國內乳品加工方式之改善。 國內大部分的廠商所採用的加熱殺菌方式為UHT(135℃加熱殺菌 2-3 秒),僅少部 分廠商使用HTST 加熱殺菌方式(75℃加熱殺菌 15 秒),因此在宣導上有極大的困難。 所以我們將先與農委會協調,並利用本實驗室所製作出的辨認生乳及還原乳的單株抗 體,幫助農委會制訂國家檢測標準,在標準中規定加熱殺菌不得超過 75℃連續加熱 15 秒鐘,並提供檢測方式。標準制訂後再進一步協調廠商改善加工方式,因此本特定目標 實施方式如下: 1. 本實驗室所研發出的抗體,可分辨出超出 80℃加熱 1 分鐘以上的牛乳,應可用於制 訂一套規範加熱殺菌的檢驗標準。 2. 與農委會協調將此規範加熱殺菌的檢驗標準列為國家檢驗標準。 3. 利用已研究出的成果與廠商協調改善乳品加工方式 4. 協助農委會檢驗牛乳加工品質 5. 幫助農委會杜絕品質不良的乳品。
結果
1. 建立台灣目前市面鮮乳之部分品質指標:
利用native-PAGE, figure 1 顯示市面鮮乳(processed milk)加工後,whey protein fraction 中之 lactoglobulin (LG)大量減少。由於 LG 在腸胃中有降膽固醇之功能(4),所
以 LG 之大量減少破壞或失性(denaturation)後可影響牛乳之品質。另外台灣 5 種名牌
與美國市場鮮乳比較(figure 2),發現美國產品中之LG 與生乳(raw milk)無顯著差異。
而進口之澳洲、紐西蘭(除丹麥外)奶粉中其 LG 之流失則相當有限,因此台灣本土之
鮮乳產品有待大家繼續努力研究。在我們的實驗中發現:加熱過程可以使 LG 大量改變
其負電荷之生化特性(figure 3),而在 Westernblot 中(figure 4)更發現加熱可使 LG 形 成聚合大分子(polymerization)。這些聚合分子在美國市售之鮮奶中並無發現。
2. 建立生乳中摻雜奶粉的檢測方式:
生乳中摻雜奶粉的問題,在台灣已存在四十多年皆無法解決,申請人利用在單株抗 體的技術預解決此問題,因此申請人先製作出對還原乳(奶粉沖泡之牛奶)高專一性之 單株抗體,進而利用ELISA 逐一篩選分別與生乳及還原乳反應,進一步篩選出與兩種乳 有不同反應之單株抗體,此抗體即可區分生乳與還原乳。如 figure 5 結果顯示初步篩選 出之單株抗體共八株,將titer 高的抗體株單株化,所得四個單株抗體分別為 1B5F2, 2F2D9, 1C10F10 及 1D8F8。由結果得知此單株抗體只能與還原乳中之蛋白質產合,不能與生乳 中之蛋白質結合。因此能將此抗體運用於區分生乳及還原乳。在深入研究中我們將奶粉 直接以不同比例摻雜入生乳中,利用此抗體偵測,其結果發現摻雜10%以上的奶粉入生 乳中可順利的偵測出。如figure 6。另一方面將生乳經加熱後,此抗體只辨認加熱後的生 乳而不辨認沒加熱過之生乳。如figure 7。此結果說明所製作出的單株抗體僅辨認受溫度 改變之乳中蛋白,因此我們再將生乳經不同溫度不同時間加熱,利用此偵測發現在80℃ 加熱一分鐘以上之生乳才會被此抗體偵測出。如figure 8,說明此抗體辨認的是加熱過度 的牛乳,此結果說明在學術上可用於研究因加熱導致結構改變的蛋白,在產業上可運用 於制訂牛乳製成中加熱的標準,進一步監控牛乳品質。利用Westernblot 分析此四株單株 抗體分別辨認牛乳中何種蛋白,分析結果得知所篩選出之單株抗體全部辨識LG 如 figure 9,此結果更可證明 LG 為熱敏感性蛋白與 Regester GO 等人所做之理論相同(17),因此 LG 在乳蛋白中扮演一個相當重要的角色,可用於當作熱處理後的指標,並且進一步運用於監控牛乳的品質。
3. 完成 LG 之 epitope mapping
蛋白質經過加熱後會導致結構的改變,結構上變化可以利用抗體技術來做區別鑑 定;LG 是牛乳中主要對熱敏感之蛋白,LG 經加熱處理後,其二級結構將嚴重改變,計畫 申請人已成功製作出只辨認奶粉而不辨認生乳之單株抗體,而此單株抗體epitope 為 LG 經加熱所產生結構上改變的區段。為了更進一步分析此單株抗體epitope,我們利用四種方式來分析此單株抗體epitope 位於 LG 的確切位置: 1. 酵素分解法(enzyme digestion) 2. 化學修飾法(chemical modification) 3. 胜肽矩陣法(peptide array) 4。胜肽合成(peptide synthesis)。
1. 酵素分解(enzyme digestion ) :
酵素分解分成兩部份進行,分別利用trypsin 及 CNBr 處理 LG 蛋白分子,將 trypsin 及CNBr 處理後之蛋白質片段利用 Werstern blot 進行分析,其結果如 figure 10 和 figure 11。 在 trypsin 所處理後之蛋白質片段,可發現當反應時間越長,其單株抗體越無法辨 認,故推測會被trypsin 切割的胺基酸 (Arg 和 Lys)可能被包含在 epitope 內,因此 trypsin
作用時一併將epitope 破壞,導致單株抗體無法辨識。而在 CNBr 分解作用後之蛋白質片
段可發現一個主要抗體辨識片段約9 kDa,因此可以推測會被 CNBr 切割的胺基酸 (Met)
沒有被包含在epitope 內,並且證明此 epitope 位於 9 kDa 之片段內。另外,利用 N 端蛋 白質定序,所得序列結果為AASDIS,此數據更加證明 LG 的 epitope 是經 CNBr 處理後 之最大片段(9kDa)。
2. 化學修飾法(chemical modification) :
單株抗體對於 epitope 的胺基酸結構具有專一性,因此利用化學修飾法改變胺基酸
結構,試圖找出影響抗體結合的化學修飾,即可以推測包含在epitope 內之胺基酸。為了
證實tryptic digestion 所提出的假設,我們使用 acetylation modification 方式將乙醯基共價 鍵結在Lys 或 Arg 的胺基上,利用 competitive ELISA 的方式,分析修飾後 LG 與單株抗 體間之結合能力,顯示經acetyl-modfication 之 LG 與單株抗體間之結合能力明顯降低, 證明Lys 或 Arg 位於 epitope 內。
3.胜肽矩陣(peptide array) :
根據酵素分解和化學修飾法所得之結果整合分析後,我們推測出 epitope 位於
residue 25 到 residue 107 間約 9 kDa 片段之中,而且 Lys 或 Arg 是構成 epitope 之必要條 件。 因此利用胜肽矩陣法分析此 9 kDa 片段,設計各為 15 個胺基酸共 12 條胜肽在 peptide array 上,如 figure 12 所示,將此 peptide array 與能辨別奶粉和生乳之單株抗體
反應後,經壓片感光處理後,共有3 點呈現反應,分別是編號 4, 8, 12 之胜肽。將此
3 條胜肽與原胺基酸序列比對,發現此 3 條胜肽之胺基酸序列有重疊之現象,且點 8 和 12 反應強度相同,相較之下點 4 的反應強度較弱。 推測點 4 胜肽之胺基酸序列僅包含
部分 epitope,導致與單株抗體間的結合能力較弱,因此分析出 epitope 之序列為
AQKKIIAEK,此結果證實先前實驗結果與假設,共有 3 個 Lys 包含在 epitope 內。 4. 胜肽合成(peptide synthesis) :
然而我們已分析出epitope 之胺基酸序列後,為了更加證實 epitope 為 peptide array
所分析之結果,利用合成水溶性胜肽方法,合成一條 10 個胺基酸之胜肽,其序列為
AQKKIIAEKT,利用 compeptitive ELISA 證實是否此胜肽具有與 LG 競爭結合單株抗體
之能力,結果顯示此合成胜肽能有效與 LG 競爭結合單株抗體,所以證實 epitope 是
AQKKIIAEKT,位於 LG 之 D 股上(figure 4)。
4. 分析出 LG 加熱後結構改變的機制
此epitope 之胺基酸序列在 LG 三級結構位置上有二個特點(figure 13):一、此 epitope在native LG 之三級結構中,呈現出穩定的二級結構(β-sheet)。二、此 epitope 和 LG 的 C 端形成雙硫鍵鍵結(disulfide bond linkage)。 然而,我們所分析出之 epitope 需在 LG 經加
熱後破壞結構才會被單株抗體所辨識,尚未破壞 LG 之結構並不會被此單株抗體所辨
識,因此我們假設此epitope 在 native LG 內,由於雙硫鍵之關係能夠形成一個穩定的二
級結構,但加熱後雙硫鍵被破壞造成此epitope 不再是穩定的二級結構,所以結構趨向亂
無章法之 disordered structure 時,才會被單株抗體所辨識,因此我們假設加熱導致此
epitope 結構改變,為了證實這個假設,我們設計了 2 個實驗:一、化學修飾法(chemical modification)。二、旋光雙極圖譜分析(circular dichroism analysis)。
1. 化學修飾法(chemical modification):
我們假設加熱會導致雙硫鍵破裂,使得此epitope 結構趨向 disordered structure 以利 於單株抗體辨識。 因此利用 carboxymethylation modification 之方式直接在 Cys 上加一個 甲基羧基,使得兩個Cys 彼此無法形成雙硫鍵鍵結。 修飾過後的 LG 利用 compeptitive ELISA 之方式來觀察是否會與加熱後 LG 競爭結合單株抗體。明顯發現,經化學修飾之 LG 結合單株抗體的能力比加熱過後之 LG 遠超出 10 倍以上,此結果證明我們的假設, 破壞雙硫鍵鍵結後之抗原決定基會更易於單株抗體的辨識。
2. 旋光雙極圖譜分析(circular dichroism analysis):
我們已確定LG 內雙硫鍵鍵結會影響 epitope 的結構,促使其與單株抗體的結合,然 而,是否如同我們的假設,此epitope 在缺乏雙硫鍵鍵結的情況下,其會形成 disorder 之 結構,不論加熱或化學修飾皆會形成此結構,為了釐清這個疑點我們利用旋光雙極光譜 分析證明我們的假設,旋光雙極光譜之特點即是可分析出蛋白質之二級結構的組成。其 結果如figure 14 所示,無論是加熱過後或是化學修飾後之 LG 皆導致 disorder 之結構形 成,進一步分析所合成之胜肽(AQKKIIAEKT)也為 disorder 之結構。此結果證明加熱導 致epitope 結構改變,由原本之 sheet 變成 disorder 之結構,此外更說明此結構改變造成 此單株抗體能夠區分奶粉或生乳之最主要原因。
5. 利用胜肽矩陣法(pepride array)分析出 epitope 上 4 個胺基酸對抗體辨識
的重要性
我們所分析出之 eptiope 上包含 4 個胺基酸帶有電荷(3 正 1 負),我們假設抗體和
epitope 間存在離子吸引力來穩定兩者結合,利用胜肽矩陣之技術製作出 12 條點突變的
胜肽,分析epitope 中哪些胺基酸會影響抗體結合的能力。胜肽設計上,將 eptiope 中所
包含之Lys 全部置換成 Ala,或置換同樣為正電荷的 Arg,而 Glu 則置換成 Ala 或同樣為 負電荷的Asp,此外,亦同時將兩個連續的 Ile 置換成 Ala。結果如 figure 15 所示,發現 將Lys69、Glu74、Ile71 及 Ile72 經置換後,抗體便無法辨認,即使置換成同樣電荷的胺 基酸 (Lys→Arg,Glu→Asp)。因此我們證明帶電荷的胺基酸的確提供離子吸引力與抗體結
合,由Lys69 與 Glu74 所提供離子吸引力具有不可置換性,Ile71 和 Ile72 亦具有不可置 換性。
6. 分析出 pH 對 LG 及抗體辨認的影響
除加熱或者化學修飾 (carboxymethylation)會造成 LG 結構的改變外,我們也發現 pH 亦會影響LG 之結構,我們利用旋光雙極圖譜分析 LG 於不同 pH 下其二級結構之改變, 結果如figure 16 所示,發現當 pH 值在 8~10 之間 LG 之二級結構有些微改變,依據文獻 報告,說明LG 上之 EF loop 於鹼性環境中會出現結構上的變化,反映在 LG 結合疏水性 分子的能力上,當LG 於鹼性環境中 EF loop 會將袋口處打開,以利疏水性分子進入袋型 結構;而在酸性環境下 EF loop 會將袋口處關上,使得疏水性分子無法進入袋型結構, 因此pH 在 LG 之生理功能扮演相當重要的調節因子。LG 在生乳中(pH 6.5)攜帶疏水性分 子通過胃 (pH 2),此時 LG 將這些疏水性分子緊密包覆在袋型結構裡不受強酸破壞,然 來進入小腸(pH 8) LG 會將這些疏水性分子釋放出,以幫助小腸對疏水性分子之吸收。另 一方面,pH 亦會影響抗體辨識 LG 之能力,明顯地發現在低 pH 時 (pH< 3),抗體與 LG 之結合能力降低,pH 4~7 有較穩定的結合能力,當 pH 越高則抗體結合能力略為降低, 利用二級抗體與一級抗體的直接反應,分析是否所有的抗原及抗體反應,都與我們產生 之單株抗體呈現在不同pH 下的結合程度,結果發現抗體與抗原之間的結合能力在 pH< 3 時最差,但於pH 4~10 並不會有所改變,因此在 pH 高於 8 時,我們產生之單株抗體與LG 的結合能力下降,再次證實 figure 15 所得到之結果,Lys69 是 epitope 中能被抗體辨 別到之重要的胺基酸,Lys 的 pKR是10.53,所以當 pH 超過 8 時,Lys 側鏈上之氫離子
逐漸地解離,導致Lys 不帶電,因此影響了抗體的結合能力。
7. 決定出 LG 的 D 股扮演的生理角色
LG 雖然生理功能未明確,但有文獻指出 LG 具有攜帶疏水性分子之功能,如維生素 A 和D(vitamin A 和 D)、棕梠酸(palmitic acid)、膽固醇(cholestrol)、膽紅素(bilirubin)等。而
文獻指出位在D 股上的 Lys69 具有穩定 LG 與疏水性分子結合之功能,然而,此 Lys69
之結合能力,且更進一步探討是否加熱後之 LG 還具有攜帶疏水性分子的功能。我們利
用兩種疏水性分子分別是維生素 A 及棕梠酸,主要因為此兩種分子已被證實能夠與 LG
結合,其次,棕梠酸帶有一負電可同時分析和 Lys69 上的正電荷之關係,由 3D 結構中
(figure 17) Lys69 帶正電荷之側鏈(side chain)與棕梠酸之羧基距離僅 3.88Å 大小,因此 Lys69 在與棕梠酸結合中應扮演重要之角色。此結果已被我們設計之兩個實驗證明。 1. 證明出 LG 於不同 pH 值擁有不同之結合能力 利用螢光光譜分析儀分析LG 在不同 pH 值中與疏水性分子之結合能力,其結果如 figure 18 所示,LG 在 pH 8~10 之條件下與維生素 A 有最好的結合能力,而在 pH 8 與棕 梠酸有最好的結合能力,隨著pH 越高則結合能力越低。因為 Lys 之 pKR為10.53,當 pH 值在10.53 時 LG 上之 Lys 將不帶電,所以當 pH 值高於 8 時,LG 與棕梠酸的結合能力 會逐漸下降。 2. 證明加熱後之 LG 喪失與疏水性分子結合能力 先前實驗結果得知加熱後之LG 會導致結構改變,使原本排列有序之二級結構變成 混亂無章的disordered structure。 因此利用螢光光譜分析儀分析加熱是否會影響 LG 結合 疏水性分子的能力。其結果如figure 19 所示,證明加熱(80℃或 100℃)後之 LG 不論對維 他命A 或棕梠酸結合能力皆明顯下降甚至喪失。 綜合以上結果,pH 會影響 LG 之結合能力,一般來說,pH 在 8~9 之間 LG 具有較 佳的結合能力。另外,值得注意的是,LG 與棕梠酸的結合程度受 Lys69 的調控,於高 pH 值(pH >8)時,導致 Lys69 不帶電,而無法與帶負電之棕梠酸形成離子吸引力,因此 證明Lys69 在 LG 結合功能上之重要性。當加熱溫度達 80℃以上會明顯降低 LG 與疏水 性分子之結合能力。所以我們所產生的單株抗體能夠分辨 LG 是否受到加熱破壞其結合 營養物質的能力。
8. 發展測定 LG 與 Vit D 結合程度之方法
在先前報告得知一個LG 與兩個分子維生素 D 結合,我們先利用螢光光譜分析儀測 定維生素D 與 LG 結合的分子數比值,再將 LG 與維生素 D 形成完全結合的複合體,比 較 LG 及複合體在螢光光譜分析儀中,給予 287nm 激發光後產生 332nm 發射光值不同
(figure 20)。除利用上述方法來測定 LG 與維生素 D 是否結合外,未來希望探討加熱後 LG 是否具有結合維生素 D 之能力,若喪失結合能力,是否結合兩分子維生素 D 之能力 皆喪失。
9. 分析出 LG 加熱後與牛乳中的 casein、lactalbumin(La)及 BSA 產生結合
還原乳中與生乳加熱後在SDS 或 native-PAGE 中分析,結果發現 LG 大量減少(figure 1),利用 Western blot 分析發現加熱後的生乳,LG 與乳中的蛋白質形成大分子(figure21)。而此大分子是LG 與乳中何種蛋白質結合目前無直接的證據證明,因此我們嘗試利
用IEF 2D-PAGE 及本實驗室所研發出的 2D-PAGE 分析,在 IEF 2D-PAGE 中並無發現明
顯的不同,除LG 明顯減少外,如 figure 22。而在本實驗室所研發出的 2D-PAGE 分析中
發現,大量減少的LG 是與牛乳中的其他蛋白產生大分子的結合,其乳中蛋白為 casein、
La 及 BSA,如 figure 23,結合的機制是利用 SH group 相互結合,因本實驗室所研發出
的2D-PAGE 中加入還原劑使利用 SH group 相互結合結合大分子變成單一蛋白質分子, 所以能清楚且直接的分析出LG 與乳蛋白中的 casein、La 及 BSA 結合,並分析出結合機 制是用雙硫鍵結合。
10. 建立蛋白質晶片偵測牛乳品質
本實驗室致力發展蛋白質晶片已二年,現階段已相當成熟,不僅是樣品的快速檢測 或是不同疾病的檢測皆已研發完成其結果如figure 24。因此在牛乳的品質檢驗我們結合 蛋白質晶片用以快速檢驗大量的牛乳樣品,其原理為利用單株抗體與 gold 結合成之immunoglold 與吸附於 nitrocellulose strip 上之生乳、還原乳反應,immunogold 利用毛細 現象與吸附於nitrocellulose strip 上之蛋白質結合,形成紅色為還原乳,無法呈色為生乳。 如figure 25,將生乳中摻有不同比例的奶粉,分別吸附於 nitrocellulose strip 上,與 gold 結合之單株抗體反應其結果在二分鐘內即可偵測出生乳中摻雜奶粉,而在結果發現摻雜
比例越高顏色越深,摻雜比例 5%以上即可偵測出。此結果說明我們已成功結合蛋白質
晶片用於牛乳品質的檢測上,我們將可運用此技術大量檢測國內牛乳品質。
11. 分析出 LG 具有抗氧化的功能
LG 的功能試驗中,我們發現 LG 具有抗氧化的功能如 figure 26,取 IC50的LG 做長
時效的抗氧化實驗,在SDS-PAGE 中我們發現隨著時間的增加 monomer 越來越少,dimer
越來越多。所以我們推測其抗氧化的機制是利用蛋白質本身被氧化,保護其他蛋白質不
會因氧化壓力而氧化。為了證明我們看到的是 LG 而不是其他雜蛋白,利用本實驗室所
製造出anti-LG 的單株抗體證明其確實為 LG。氧化後 LG 形成 dimer 或是 polymer 的型 態與加熱過後的形式非常相似如figure 27,根據文獻指出 dimer 的型態是利用 LG 的 SH group 形成,所以利用化學修飾及加熱的方式將 free-SH group 修飾,發現 LG 就會失去 抗氧化的功能,如figure 28。所以 LG 主要是利用其為在第 121 胺基酸位置的 free-SH 來 抵抗氧化的壓力,藉由犧牲自己來避免LDL 的氧化。
12. 建立小鼠牛乳過敏動物模式
本實驗室為了研究牛乳所造成的過敏,我們建立了小鼠動物模式,將均質過之牛乳 直接注射入小鼠腹腔,相隔14 天在注射一次,此時會造成小鼠有強烈的過敏反應,我們 利用血液分析結果得知血液內含有IgE。蛋白質晶片分析亦得到相同的結果如 figure 29。 進一步分析造成牛乳過敏的主要乳蛋白為LG。所以造成牛乳過敏的主要蛋白質為 LG。 由上述的結果本實驗室已成功的建立小鼠牛乳過敏動物模式,我們將利用此動物模式更 深入的研究LG 造成小鼠過敏的機制。β-LG β-LG α-LA
A B
Figure 1:本實驗室利用 15% Native-PAGE 之電泳膠片分析生乳及鮮乳 圖。A Lane 為生乳。B Lane 為加工過之生乳。其中在 Lane A 上之兩種 蛋白質(β-LG)經加溫處理後大量消失。
1 3
2 8
4
5
6
7
9
10
11 12 13
Figure 2:利用 15% Native-PAGE 之電泳膠片分析台灣不同廠牌之市售鮮 乳及奶粉。其中Lane 1 為直接由牧場取得之生乳、Lane 2 至 Lane 6 為台
灣市售鮮乳、Lane 7 至 Lane 9 為進口奶粉分別來自丹麥、澳洲及紐西蘭、
Lane 10 至 Lane 13 為來自美國市售鮮乳。在鮮乳之β-LG 明顯減少,而 奶粉與美國市售鮮乳中β-LG 仍然存在。
1 3
2
4
5
6
7
β-LG β-LG α-LA Figure 3:利用 15% Native-PAGE 之電泳膠片分析不同時間熱處理(95℃) 之α-LA 及β-LG 之變化。 Lane 1 至 7 為不同熱處理時間分別為 0、30、 60、120、240、480 及 720 秒。 β-LG 之變性與時間成正比A. Native - PAGE
B. SDS - PAGE
7.4 84 120 205 36 52 30 M 0 480 90015 30 60 120 240 0 15 30 60 120 240 480 900Figure 4:利用 Western blot 分析在不同時間熱處理後之β-LG 變化。(A) 15% Native-PAGE。(B) 15% SDS-PAGE。分析與β-LG 抗體之反應,Lane 左至右為不同熱處理時間分別為0、15、30、60、120、240、480 及 900 秒。β-LG 明顯在加熱時間越長蛋白濃度逐漸降低。並且發現高分子聚 合物 (panel B)。
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0 2 4 5 10 20 40 60 80 100
Percentage of dry milk added into raw milk
OD at 405 n m 0 0.1 0.2 0.3 0.4 0.5 0.6 1 2 3 4 5 6 7 8 Primary clones OD at 405 nm Raw milk Dry milk Figure 5:利用 ELISA 初步所篩選出只會辨認還原乳不會辨認生乳。篩選 出有高titer 的抗體株單株化分別為 1, 2, 3 及 5 (1B5F2, 2F2D9, 1C10F10 及1D8F8) Figure 6:利用 ELISA 分析辨認還原乳單株抗體在生乳中加入不同比例的 還原乳的免疫結合能力。由結果得知加入還原乳比例越高所分析的讀值越 高,因此可利用此單株抗體解決生乳中還原乳的摻雜問題。
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 1B5F2 2F2D9 1C10F10 1D8F8 2B4B4 Clones OD at 405 nm
Raw milk Raw milk heated
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45 0 50 60 70 80 90 Temperature OD at 405nm 15secs 30secs 1mins 2mins 4mins 8mins 16mins Figure 7:利用 ELISA 分析辨認牛乳的單株抗體與生乳加熱後的結合力。 生乳加熱後加(熱 95℃15 分鐘)與此單株抗體的結合力增加五倍以上, 此結果可推測加熱後會暴露出新的epitope。2B4B4 的單株抗體為同時辨認 生乳與還原乳。 Figure 8:利用 ELISA 生乳經不同時間不同溫度加熱後此辨認還原乳單株 抗體的免疫結合力。將生乳用50, 60, 70, 80 及 90℃加熱 15’’, 30’’, 1’, 2’, 4’, 8’ 及 16’。結果當生乳加熱 80℃以上 1 分鐘即可被此抗體辨認。因此可用 此抗體訂定國家標準。
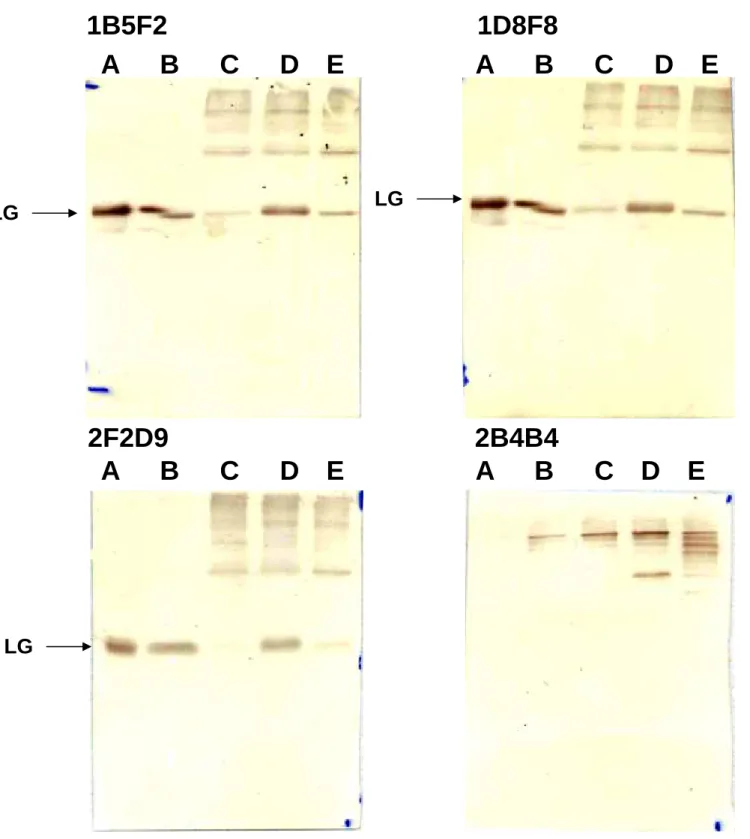
Figure 9:利用 Western blot 分析還原乳的專一性單株抗體(1B5F2, 2F2D9
及1D8F8)及缺乏專一性的單株抗體的特性。每一個 lane 放入 10ug 的乳
蛋白。Lane A:為 native LG、lane B:為生乳、lane C:為市售鮮乳、lane D:為還原乳及 lane E:加熱 95℃15 分鐘後的生乳。
1B5F2 1D8F8
A B C D E
2F2D9 2B4B4
LG
LG
LG
A B C D E
A B C D E
A B C D E
Figure 10:利用 western blot 分析由 trypsin 分解 LG 之片段與抗體結合情形。每個 well 加入濃度為 10 µg 之蛋白質。LG 和 trypsin 量之比例為 500:1。 Lane M:marker。 Lane A:LG + trypsin 反應 5 mins。 Lane B:LG + trypsin 反應 10 mins。Lane C:LG + trypsin 反應 30 mins。結果發現此抗體無法辨認到被 trypsin 分解之 LG 片段。
LG
SDS-PAGE
Western blot
M
A B
C
M A
B
C
Figure 11:利用 western blot 分析由 CNBr 所分解 LG 之片段與抗體結合情形。每個 well 加入濃度為 30 µg 之蛋白質。Lane M:marker。 Lane A: LG。 lane B:LG + CNBr。從結果得知 epitope 應位於 9 kDa 之片段內。
M A
B
SDS-PAGE Western blot
M A
B
9 kDa
LG
Figure 12:利用 peptide array 之技術在 membrance 上合成各為 15 個胺基酸共 12 條胜肽,並
利用抗體分析。結果顯示胜肽4. 8. 12 會與抗體結合,將結果和序列對照可以發現一個重疊的
區域 (黑框內),其序列是 AQKKIIAEK.
Figure 13: 利用 LG 之 3D 結構分析 epitope,發現我們所分析出之 epitope 位於 D strand 上 (綠色),此外更發現 D strand 與 C 端有雙硫鍵鍵結 (Cys66-Cys160)來穩 定 β-sheet 二級結構。
Figure 14: 利用旋光雙極圖譜分析 LG 經不同處理 (加熱,化學修飾)及合成胜肽。結果發 現除native LG 保有完整之β-sheet (215nm 有最低 ellipticity),其他三個皆傾向混亂無章 disorder 之結構。
Figure 15: 利用 peptide array 技術製作不同點突變之胜肽,分析抗體與 epitope 結合之特性。 結果發現Lys69 和 Glu74 不論是突變成不帶電的 Ala 或是帶相同電荷的 Arg 或 Asp,皆無法
與抗體結合產生反應,相較之下Lys70 及 75 卻不受突變而影響與抗體的結合能力,另外 Ile71
和 Ile72 突變也會影響抗體的結合。總結而言 Lys69、Glu74、Ile71 and Ile72 構成 epitope 中能否被抗體辨識之重要胺基酸。
Figure 17: 利用這 3D 結構圖分析 epitope 與棕梠酸(PA)間之相互關係。發現 epitope 上之 Lys69 與 Ile71 具有穩定 LG 與棕梠酸(PA)結合的功能。Lys69 提供一正電荷與棕梠酸(PA)
上的負電荷形成離子吸引力,另一方面Ile71 則和棕梠酸(PA)上疏水性部分提供了疏水 性作用力。 Figure 16: 利用旋光雙極圖譜分析 pH 對 LG 結構之影響。發現當 pH 值高於 8 時,LG 之結構稍微有點變化,此改變可能與LG 於鹼性環境中 EF loop 會將袋口處打開有關, 在高pH 值時,有利於疏水性分子進入,反之低 pH 值時緊密地將疏水性分子保護在 LG 袋口內,藉此方式來控制疏水性分子結合。
Figure 19: 利用螢光光譜分析儀分析加熱是否會影響 LG 結合疏水性分子的能力。發
現當LG 加熱溫度高於 80℃時,與維生素 A 或棕梠酸的結合能力大幅下降,亦證明結
構遭受到破壞時,同時喪失結合疏水性分子之功能。
D. Palmitic acid binding
0 20 40 60 80 100 120 native LG 80℃ 100℃ R e la tiv e e n h a n c e m e n t (% )
Palmitic acid
native LG
80℃
100℃
B. Retinol binding 0 20 40 60 80 100 120 native LG 80℃ 100℃ R e la ti v e i n te n s it y ( % )native LG
80℃
100℃
Retinol
Figure 18: 利用螢光光譜分析儀分析 LG 結合維生素 A 或棕梠酸是否會受到 pH 值之影 響。結果發現LG 在 pH 6 以下之環境,幾乎沒有任何能力結合維生素 A 或棕梠酸,相對 LG 在弱鹼環境下 (pH 8-9) 具有較佳的結合能力.不過對棕梠酸而言,當 pH 達到 10 時,結 合能力為原有一半,可能原因與LG 上之 Lys 側鏈上氫離子解離有關係。 A. Retinol binding -5 0 5 10 15 20 25 0 2 4 6 8 10 12 E m issi o n i n te n s it y at 470 n m 2 4 6 8 10 pH -5 0 5 10 15 20 25 0 2 4 6 8 10 12 pH % C h an g e o f e m issi o n a t 332 n m C. Palmitic acid pHA. Native - PAGE B. SDS - PAGE
0
15 30 60 120 240
480 960
LG
7.4 84 120 205 36 52 30M
0
15 30 60 120 240
480 960
LG
Figure 21:利用Western blot 分析在不同時間熱處理後之β-LG 變化。(A) 15% Native-PAGE。(B) 15% SDS-PAGE。分析與β-LG 抗體之反應,Lane 左至右為不同熱處理時間分別為0、15、30、60、 120、240、480 及900秒。β-LG 明顯在加熱時間越長蛋白濃度逐漸降低。並且發現高分子聚合物 (panel B)。 0 200 400 600 800 250 300 350 400 450 500 Wavelength (nm) E m is s io n in te n s it y 5µM LG 5µMLG + 10µM VitD3 0 200 400 600 800 250 300 350 400 450 500 Wavelength (nm) E m is s io n in te n s it y 5µMLG 5µMLG + 10µM VitD2 Figure 20: 利用螢光光譜分析儀分別比較 LG 及 LG 與維生素 D2或維生素D3完全結合形成 之複合體在332nm 發射光值。將可利用此方法定量結合多少比率維生素 D 或藥物。
Figure 23:利用本實驗室研發出的 2D-PAGE 分析。牛乳經加熱後 LG 與乳中哪幾 種蛋白質結合。A:並未加熱的生乳。B:加熱後的生乳。結果發現生乳經加熱 後LG 與 BSA、Casein 及 LA 結合成大分子。
LG
Figure 22:利用 IEF 2D-PAGE 分析牛乳經加熱後 LG 與乳中哪幾種蛋白質結合。 A:並未加熱的生乳。B:加熱後的生乳。結果並未發現有不相同的地方。
A B
LG BSA Casein LAA B
大分子 小分子 小分子 小分子Figure 24:利用蛋白質晶片快速檢測不同病人有無得到疾病。此晶片大小為 1.5 cm x 2 cm,晶片上共有 100 病人檢體,此結果在二分鐘以內即可診斷出。
Figure 25:利用蛋白質晶片快速檢測牛乳中有無摻雜奶粉。由右至左摻雜濃度為 100, 90, 80, 70, 60, 50, 40, 30, 20, 10, 5 及 0%,最右邊為 positive control。用此晶片
0 0.2 0.4 0.6 0.8 1 1.2 0 10 20 30 40 50 60 70 80 90 100 Protein concentration (µM) Absorbance at 540nm probucol Vitamine E LG heated LG BSA PBS Figure 26:不同物質及β-LG 的抗氧化能力分析。LG 在濃度 80ug 達到最大抗氧化 效力,加熱過的LG 則無抗氧化的能力。
Figure 27:利用 SDS-PAGE 分析加熱後與氧化後之 LG。Lane M:為 maker。 Lane A:為 native LG。 Lane B:為加熱後之 LG。Lane C:為氧化後之 LG。在加熱後 與氧化後之LG 會形成 dimer 的結構。 KDa 119 79 46 24 19 31 M A B C
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0
20
40
60
80
100
120
LG conentration(µM)
Absorbance at 540nm
LG modify LG Figure 28:利用化學修飾法將 LG 修飾後分析其抗氧化力。經化學修飾後之 LG 抗氧化力明顯比native LG 低。證明 SH group 為主要參與 LG 抗氧化機制。 Figure 29:利用蛋白質晶片診斷過敏小鼠血液中有無 IgE。A:未過敏之小鼠血液。 B:牛乳過敏之小鼠血液。結果得知本實驗室已成功誘發牛乳過敏。A
B
參考文獻:
1. Desjeux JF, Heyman M. Milk proteins, cytokines and intestinal epithelial functions in
children. Acta Paediatr Jpn. 1994 Oct;36(5):592-6.
2. Sawyer L, Kontopidis G. The core lipocalin, bovine beta-lactoglobulin. Biochim Biophys
Acta. 2000 Oct 18;1482(1-2):136-48.
3. Perez MD, Calvo M. Interaction of beta-lactoglobulin with retinol and fatty acids and its
role as a possible biological function for this protein: a review. J Dairy Sci. 1995 May;78(5):978-88.
4. Nagaoka S, Futamura Y, Miwa K, Awano T, Yamauchi K, Kanamaru Y, Tadashi K, Kuwata
T. Identification of novel hypocholesterolemic peptides derived from bovine milk beta-lactoglobulin. Biochem Biophys Res Commun. 2001 Feb 16;281(1):11-7.
5. Lonnerdal B. Nutritional and physiologic significance of human milk proteins. Am J Clin
Nutr. 2003 Jun;77(6):1537S-1543S.
6. Bounous G, Molson JH. The antioxidant system. Anticancer Res. 2003
Mar-Apr;23(2B):1411-5.
7. Kuitunen M, Savilahti E, Sarnesto A. Human alpha-lactalbumin and bovine
beta-lactoglobulin absorption in infants. Allergy. 1994 May;49(5):354-60.
8. Sorva R, Makinen-Kiljunen S, Juntunen-Backman K. Beta-lactoglobulin secretion in
human milk varies widely after cow's milk ingestion in mothers of infants with cow's milk allergy. J Allergy Clin Immunol. 1994 Apr;93(4):787-92.
9. Kuitunen OO, Savilahti E, Sarnesto A. Human alpha-lactalbumin and bovine
beta-lactoglobulin absorption in premature infants. Pediatr Res. 1994 Mar;35(3):344-7.
10. Minehira K, Inoue S, Nonaka M, Osada K, Yamada K, Sugano M. Effects of dietary
protein type on oxidized cholesterol-induced alteration in age-related modulation of lipid metabolism and indices of immune function in rats. Biochim Biophys Acta. 2000 Jan 3;1483(1):141-53.
β-Lactoglobulin (2004) (To be submitted)
12. W. L. Chen, M. T. Hwang, H. C. Liu, C. W. Li, and S. J. T. Mao, Distinction between dry
and raw milk using monoclonal antibodies prepared against dry milk proteins. (Submitted)
13. W.L. Chen, C.Y. Liau, and S. J. T. Mao. Analyses of β-Lactoglobulin as a thermal marker
in raw and commercially processed milk using native gel electrophoresis, Westernblot, and circular dichroic spectrum. (Submitted)
14. Lovegrove JA, Osman DL, Morgan JB, Hampton SM. Transfer of cow's milk
beta-lactoglobulin to human serum after a milk load: a pilot study. Gut. 1993 Feb;34(2):203-7.
15. Mao FC, Bremel RD. Enzyme-linked immunosorbent assays for bovine alpha-lactalbumin
and beta-lactoglobulin in serum and tissue culture media. J Dairy Sci. 1991 Sep;74(9):2946-51.
16. Mao FC, Bremel RD, Dentine MR. Serum concentrations of the milk proteins
alpha-lactalbumin and beta-lactoglobulin in pregnancy and lactation: correlations with milk and fat yields in dairy cattle. J Dairy Sci. 1991 Sep;74(9):2952-8.
17. Regester GO, Pearce RJ, Lee VW, Mangino ME. Heat-related changes to the
hydrophobicity of cheese whey correlate with levels of native beta-lactoglobulin and alpha-lactalbumin. J Dairy Res 1992 Nov;59(4):527-32.
18. Sakurai K, Goto Y. Manipulating monomer-dimer equilibrium of bovine Beta
-lactoglobulin by amino acid substitution. J Biol Chem 2002 Jul 12;277(28):25735-40.
19. Taulier N, Chalikian TV. Characterization of pH-induced transitions of beta-lactoglobulin:
ultrasonic, densimetric, and spectroscopic studies. J Mol Biol 2001 Dec 7;314(4):873-89.
20. Ikeuchi Y, Nakagawa K, Endo T, Suzuki A, Hayashi T, Ito T. Pressure-induced
denaturation of monomer beta-lactoglobulin is partially irreversible: comparison of monomer form (highly acidic pH) with dimer form (neutral pH). J Agric Food Chem 2001 Aug;49(8):4052-9.
21. Cho Y, Singh H, Creamer LK. Heat-induced interactions of beta-lactoglobulin A and
22. Havea P, Singh H, Creamer LK. Heat-induced aggregation of whey proteins: comparison
of cheese WPC with acid WPC and relevance of mineral composition. J Agric Food Chem 2002 Jul 31;50(16):4674-81.
23. Jarvinen KM, Chatchatee P, Bardina L, Beyer K, Sampson HA. IgE and IgG binding
epitopes on alpha-lactalbumin and beta-lactoglobulin in cow's milk allergy. Int Arch Allergy Immunol. 2001 Oct;126(2):111-8.
24. Selo I, Negroni L, Creminon C, Yvon M, Peltre G, Wal JM. Allergy to bovine
beta-lactoglobulin: specificity of human IgE using cyanogen bromide-derived peptides. Int Arch Allergy Immunol. 1998 Sep;117(1):20-8.
25. Selo I, Clement G, Bernard H, Chatel J, Creminon C, Peltre G, Wal J. Allergy to bovine
beta-lactoglobulin: specificity of human IgE to tryptic peptides. Clin Exp Allergy. 1999 Aug;29(8):1055-63.
26. Yvon M, Levieux D, Valluy MC, Pelissier JP, Mirand PP. Colostrum protein digestion in
newborn lambs. J Nutr. 1993 Mar;123(3):586-96.
27. Savilahti E, Kuitunen M. Allergenicity of cow milk proteins. J Pediatr. 1992 Nov;121(5 Pt
2):S12-20.
28. Grzybowski J, Antos M, Sakiel S, Dobke MK. Antidietary antigen antibodies in the sera of
patients with burns as a potential marker of gut mucosa integrity failure. J Burn Care Rehabil. 1992 Mar-Apr;13(2 Pt 1):194-7.
29. Gotteland M, Crain-Denoyelle AM, Heyman M, Desjeux JF. Effect of cow's milk protein
absorption on the anaphylactic and systemic immune responses of young rabbits during bacterial diarrhoea. Int Arch Allergy Immunol. 1992;97(1):78-82.
30. Enomoto A, Konishi M, Hachimura S, Kaminogawa S. Milk whey protein fed as a
constituent of the diet induced both oral tolerance and a systemic humoral response, while heat-denatured whey protein induced only oral tolerance. Clin Immunol Immunopathol. 1993 Feb;66(2):136-42.
31. Davis PJ, Williams SC. Protein modification by thermal processing. Allergy. 1998;53(46
32. Nakagawa T. [IgE antibody determination] Nippon Rinsho. 2001 Oct;59(10):1950-4. 33. Paschke A, Zunker K, Wigotzki M, Steinhart H. Determination of the IgE-binding activity
of soy lecithin and refined and non-refined soybean oils. J Chromatogr B Biomed Sci Appl. 2001 May 25;756(1-2):249-54.
34. Pennisi E. Human Genome Project. And the gene number is...? Science. 2000 May
19;288(5469):1146-7.
35. Oshima A, Asayama K, Sakai N, Kitajima M. The role of endogenous free radical
scavengers on tissue recovery in the experimental ulcer model. J Clin Gastroenterol. 1990;12 Suppl 1:S58-64.
36. Biswas K, Bandyopadhyay U, Chattopadhyay I, Varadaraj A, Ali E, Banerjee RK. A novel
antioxidant and antiapoptotic role of omeprazole to block gastric ulcer through scavenging of hydroxyl radical. J Biol Chem. 2003 Mar 28;278(13):10993-1001. Epub 2003 Jan 15.
37. Kahraman A, Erkasap N, Koken T, Serteser M, Aktepe F, Erkasap S. The antioxidative and
antihistaminic properties of quercetin in ethanol-induced gastric lesions. Toxicology. 2003 Feb 1;183(1-3):133-42.
38. Mathewson M, Farnham C. Milk therapy in ulcer disease: yes or no? Home Healthc Nurse.
1984 Jul-Aug;2(4):8.
39. Dial EJ, Lichtenberger LM. A role for milk phospholipids in protection against gastric acid.
Studies in adult and suckling rats. Gastroenterology. 1984 Aug;87(2):379-85.
40. Dial EJ, Lichtenberger LM. Milk protection against experimental ulcerogenesis in rats. Dig
Dis Sci. 1987 Oct;32(10):1145-50.
41. Dial EJ, Romero JJ, Lichtenberger LM. Gastroprotection by dairy foods against
stress-induced ulcerogenesis in rats. Dig Dis Sci. 1995 Nov;40(11):2295-9.
42. Matsumoto H, Shimokawa Y, Ushida Y, Toida T, Hayasawa H. New biological function of
bovine alpha-lactalbumin: protective effect against ethanol- and stress-induced gastric mucosal injury in rats. Biosci Biotechnol Biochem. 2001 May;65(5):1104-11.
43. Rosaneli CF, Bighetti AE, Antonio MA, Carvalho JE, Sgarbieri VC. Efficacy of a whey
protein concentrate on the inhibition of stomach ulcerative lesions caused by ethanol ingestion. J Med Food. 2002 Winter;5(4):221-8.
44. Chi Feng Tseng, Chia Ching Lin, Hsing Yi Huang, Huei Chun Liu, Simon J.T. Mao*
Antioxidant role of human haptoglobin. (Submitted)
45. Mao S. J., Yates M. T., Jackson R. L., Antioxidant activity and serum levels of probucol
計畫成果
今年度計畫主要有五項執行目標,除目標二尚未完成其餘各項皆已完成,執行效率達 95%。其 主要成果如下:
國際及國內會議(international conference)
1. Chen WL, Ho JC, Hoang KC, and S. J. T. Mao. (2001) Differentiation between fresh and powdered milk using chemical, physical, and immunochemical analyses. The sixteenth joint annual conference of biomedical sciences. Programs & abstracts.
2. Chen WL, S. J. T. Mao. (2002) Analyses of β-lactoglobulin as a thermal marker in raw and commercially processed milk using a native gel electrophoresis and circular dichroic spectrum. Experimental Biology 2002 New Orleans, LA, USA. FASEB J. 16:A1187. 3. Liau CY, Chen WL, Wu YJ, Lin CC, and S. J. T. Mao. (2002) Differentiation of Milk
Proteins Using Phage Displayed Antibodies. The seventeenth joint annual conference of biomedical sciences. Oral presentation.
4. Chen WL, and S. J. T. Mao. (2003) Distinguish between dry and raw milk using monoclonal antibodies prepared against dry milk proteins: LG is a sensitive thermal marker in dry milk. Experimental Biology 2003 San Diego, USA. FASEB J. 17:A1127.
5. Chen WL, Huang MT, Liu HC, Li CW, and S. J. T. Mao. (2003) Production of conformationally dependent monoclonal antibody against thermal-denatured milk. The eighteenth joint annual conference of biomedical sciences. Oral presentation. P81
6. Liu HC, Huang MT, Chen WL, and S. J. T. Mao. (2003) Effect of disulfide bond for the antioxidant activity of β-lactoglobulin. The eighth conference on biochemical engineering. Oral presentation. P205
7. Song CY, Chen WL, Wang SS, and S. J. T. Mao. (2004) Thermal denaturation of milk β-lactoglobulin. The nineteenth joint annual conference of biomedical sciences. Programs & abstracts.
8. Chen WL, Huang MT, Li CW, Liu HC, and S. J. T. Mao. (2004) Thermal denaturation of β-lactoglobulin as probed by an monoclonal antibody. Experimental Biology 2004 Washington, DC, USA. FASEB. 18:A144.
9. Chen WL, Yang MC, Song CY, and S. J. T. Mao. (2005) Thermal denaturation of β-lactoglobulin as probed by specific monoclonal antibodies. The twentieth joint annual conference of biomedical sciences. Programs & abstracts. P396
10. Yang MC, Chen WL, and S. J. T. Mao. (2005) Ligand binding ability of β-lactoglobulin in native and thermal denatured state. The twentieth joint annual conference of biomedical sciences. Oral presentation. P230
11. Chen W L, Huang MT, and S. J. T. Mao. (2005) A novel two-dimentional gel electrophoresis for studing the cress-linking between β-Lactoglobulin and milk proteins. 2005 ADSA-ASAS-CSA joint annual meeting. Programs & abstracts. P144.
12. S. J. T. Mao, Chen WL, Yang MC, and Liu WT. Distinguish between native and thermally denatured β-lactoglobulin using a monoclonal antibody as a probe. 2005 ADSA-ASAS-CSA joint annual meeting. Oral presentation. P77.
13. Song CY, Yang MC, and S. J. T. Mao. Residues 69-74 of beta-lactoglobulin are responsible for a monoclonal antibody binding to thermal denatured lactoglobulin. 2005 ADSA-ASAS-CSA joint annual meeting. Oral presentation. P76.
發表之 SCI 論文
1. Chen WL, Huang MT, Liu HC, Li CW, S. J. T. Mao. (2004) Distinction between dry and raw milk using monoclonal antibodies prepared against dry milk proteins. J Dairy Sci., 87:2720-2729.
2. Song CY, Chen WL, Yang MC, Huang JP, S. J. T. Mao. (2005) Epitope mapping of a monoclonal antibody specific to bovine dry milk: involvement of residues 66-76 of strand D in thermal denatured beta-lactoglobulin. J Biol Chem. 280:3574-3582.
3. Chen WL, Hwang MT, Liau CY, Ho JC, Hong KC, S. J. T. Mao. (2005) {beta}-Lactoglobulin is a Thermal Marker in Processed Milk as Studied by Electrophoresis and Circular Dichroic Spectra. J Dairy Sci. 88:1618-1630.
4. Liu H. C and S. J. T. Mao. (2005) Effect of disulfide bond for the antioxidant activity of β-Lactoglobulin (Submitted to J Dairy Sci.)