國
立
交
通
大
學
生 物 科 技 學 系
博
士
論
文
利用 C 型肝炎病毒次基因系統研究花生四烯酸及銻化物
的抗病毒作用
Using HCV Subgenomic Replicon System to study the anti-HCV activity
of the Arachidonate and Antimonial Compounds
研 究 生:呂光洲
指導教授:林苕吟 博士
徐祖安 博士
利用 C 型肝炎病毒次基因系統研究花生四烯酸及銻化物的抗病毒作用
Using HCV Subgenomic Replicon System to study the anti-HCV activity
of the Arachidonate and Antimonial Compounds
研 究 生:呂光洲 Student:Guang-Zhou Leu
指導教授:林苕吟 Advisor:Tiao-Yin Lin
徐祖安 Co-Advisor:John T-A. Hsu
國 立 交 通 大 學
生 物 科 技 學 系
博 士 論 文
A Thesis
Submitted to Department of Biological Science and Technology College of Biological Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Philosophy
in
Biological Science and Technology
July 2005
Hsinchu, Taiwan, Republic of China
博碩士論文授權書
本授權書所授權之論文為本人在_國立交通大學_大學(學院)_生物科技學_系所 _______組_九十三_學年度第_二_學期取得_博_士學位之論文。 論文名稱:_利用C型肝炎病毒次基因系統研究花生四烯酸及銻化物的抗病毒作用_ 指導教授:_林苕吟 博士 、 徐祖安 博士________ 1. □同意 □不同意 本人具有著作財產權之上列論文全文(含摘要)資料,授予行政院國家科學委員會科學技 術資料中心(或改制後之機構),得不限地域、時間與次數以微縮、光碟或數位化等各種 方式重製後散布發行或上載網路。 本論文為本人向經濟部智慧財產局申請專利(未申請者本條款請不予理會)的附件之 一,申請文號為:______________,註明文號者請將全文資料延後半年再公開。 2. □同意 □不同意 本人具有著作財產權之上列論文全文(含摘要)資料,授予教育部指定送繳之圖書館及國 立交通大學圖書館,基於推動讀者間「資源共享、互惠合作」之理念,與回饋社會及學 術研究之目的,教育部指定送繳之圖書館及國立交通大學圖書館得以紙本收錄、重製與 利用;於著作權法合理使用範圍內,不限地域與時間,讀者得進行閱覽或列印。 本論文為本人向經濟部智慧財產局申請專利(未申請者本條款請不予理會)的附件之 一,申請文號為:______________,註明文號者請將全文資料延後半年再公開。 3.5同意 □不同意 本人具有著作財產權之上列論文全文(含摘要),授予國立交通大學與台灣聯合大學系統 圖書館,基於推動讀者間「資源共享、互惠合作」之理念,與回饋社會及學術研究之目 的,國立交通大學圖書館及台灣聯合大學系統圖書館得不限地域、時間與次數,以微縮、 光碟或其他各種數位化方式將上列論文重製,並得將數位化之上列論文及論文電子檔以 上載網路方式,於著作權法合理使用範圍內,讀者得進行線上檢索、閱覽、下載或列印。 論文全文上載網路公開之範圍及時間 – 本校及台灣聯合大學系統區域網路: 95 年 03 月 01 日公開 校外網際網路: 95 年 09 月 01 日公開 上述授權內容均無須訂立讓與及授權契約書。依本授權之發行權為非專屬性發行權利。依本 授權所為之收錄、重製、發行及學術研發利用均為無償。上述同意與不同意之欄位若未鉤選, 本人同意視同授權。 研究生簽名: 學號: 8728806 (親筆正楷) (務必填寫) 日期:民國九 十 四年 七 月二十二日國家圖書館博碩士論文電子檔案上網授權書
本授權書所授權之論文為本人在_國立交通大學大學(學院)_生物科技學系所 ______組_九十三_學年度第_二_學期取得_博_士學位之論文。 論文名稱:利用C型肝炎病毒次基因系統研究花生四烯酸及銻化物的抗病毒作用 指導教授:_林苕吟 博士 、 徐祖安 博士_________ 5同意 □不同意 本人具有著作財產權之上列論文全文(含摘要),以非專屬、無償授權國家圖書館, 不限地域、時間與次數,以微縮、光碟或其他各種數位化方式將上列論文重製, 並得將數位化之上列論文及論文電子檔以上載網路方式,提供讀者基於個人非營 利性質之線上檢索、閱覽、下載或列印。 上述授權內容均無須訂立讓與及授權契約書。依本授權之發行權為非專屬性發行 權利。依本授權所為之收錄、重製、發行及學術研發利用均為無償。上述同意與 不同意之欄位若未鉤選,本人同意視同授權。 研究生簽名: 學號: 8728806 (親筆正楷) (務必填寫) 日期:民國九 十 四年 七 月二十二日 1. 本授權書請以黑筆撰寫,並列印二份,其中一份影印裝訂於附錄三之 一(博碩士論文授權書)之次頁﹔另一份於辦理離校時繳交給系所助 理,由圖書館彙總寄交國家圖書館。For My wife, my sons and my family
Guang-Zhou Leu
August 2005
誌
謝
在這走得不算順利的博士班生涯中,我要衷心的感謝 林苕吟博士與 徐祖安 博士的指導與教悔,以及在我關鍵的時候拉我一把,使得今天得以順利畢業,當 然,最需要感謝的是默默支持我及我的家庭的太太—陳玲貴。如果沒有她的支持 與鼓勵,就沒有今天的我,在此我要說聲:老婆,謝謝妳。
利用
C 型肝炎病毒次基因系統研究花生四烯酸及銻化物的抗病毒作用
學生:呂光洲
指導教授
:林苕吟 博士
徐祖安 博士
國立交通大學生物科技學系﹙研究所﹚博士班
摘
要
C 型肝炎病毒是引起肝硬化與肝癌的主要元兇之ㄧ,根據世界衛生組織在一九
九七年統計的結果估算,全世界約有百分之三的人是
C 肝病毒帶原者,也就是將
近有一億七千萬人是染有慢性
C 型肝炎病毒,就我國的衛生署疾病管制局的統計
結果顯示,目前為止,約有七十萬國人染有
C 型肝炎,大約佔全台總人口的 3.5
﹪,遠高於歐、美等先進國家,再加上
C 型肝炎病毒並無疫苗可以防治,也沒有
特效藥可以治療,目前針對慢性
C 型肝炎的治療都是合併使用干擾素(IFN-α)
及雷巴威林(ribavirin),此一療程的最大問題在於有些人會有嚴重的副作用,
諸如發燒、發炎等症狀,此外,低的治癒率亦是此一治療方式需要克服的地方,
因此,為有效治療慢性
C 型肝炎,研發高效率及低副作用的藥物是當務之急。
C 型肝炎病毒自一九八九年被鑑定出來後,其相關的研究在世界各地展開,包
括有病毒學、致病機制、治療藥物的開發等,其中最大的突破之ㄧ應該是在西元
二千年左右的
C 型肝炎病毒次基因體複製模式的發現,使人類可以更加了解此一
病毒在宿主細胞內的情形,也使得針對病毒複製所開發的相關抑制藥物得以進展
做進一步的研究,醣酸銻鈉(Sodium Stiboglucognate;SSG)為抗寄生蟲用藥,
SSG 具有降低 C 型肝炎病毒基因的能力,且與干擾素合併使用有協同性加成的
抑制效果,不過此一藥物抑制病毒的機制尚待做進一步的研究,本報告中的另一
具有抑制
C 型肝炎病毒基因的物質是多元不飽和脂肪酸,多元不飽和脂肪酸普遍
存在於動植物中,常用於食品添加的營養補充劑,研究的結果顯示,多元不飽和
脂肪酸可以抑制含
C 型肝炎病毒次基因體細胞中的 C 型肝炎病毒基因,而且和
干 擾 素 一 起 使 用 抑 制 效 果 更 好 , 經 由 進 一 步 的 結 果 顯 示 , 以 花 生 四 烯 酸
(Arachidonic Acid;AA)為例,隨著花生四烯酸的處理濃度提高,抑制含 C 型
肝炎病毒次基因體細胞中的
C 型肝炎病毒基因效果越明顯,同時細胞內的脂肪性
過氧化物的產物也隨之提高,此過氧化產物也會抑制體細胞中的
C 型肝炎病毒基
因,此外,經由試管外的酵素活性測試結果顯示,花生四烯酸可抑制
C 型肝炎病
毒的蛋白脢的活性,本論文的研究成果顯示,由臨床用藥及天然食品中找出具有
病毒抑制效果的物質,將有助於藥物的發展,希望藉由這樣的成果,讓我們在
C
型肝炎防治上能有新天地。
Using HCV Subgenomic Replicon System to study the anti-HCV activity of the
Arachidonate and Antimonial Compounds
Student:Guang-Zhou Leu
Advisors: Dr. Tiao-Yin Lin
Dr. John T-A. Hsu
Department﹙Institute﹚of Biological Science and Technology
National Chiao Tung University
ABSTRACT
Hepatitis C virus causes severe liver diseases, such as liver cirrhosis and hepatocellular carcinoma (HCC). Acute infections in individuals often lead to development of chronic hepatitis. In 1997, it was estimated by WHO that more than 170 million, approximate 3% of worldwide population people suffer from chronic HCV infection. Pegylated interferon-α plus ribavirin is the only treatment option to combat HCV, but low curring rates and severe side effects are problems. There is no effective vaccine in the world to prevent HCV infection. Thus, more effective therapy is in urgent need due to the sever side effect and unsatisfactory curing rate of the current therapy. In this disertation, we found that polyunsaturated fatty acids (PUFAs) and sodium stibogluconate were able to exert anti-HCV activity by using HCV subgenomic replicon system. When replicon cells were treated with either compound in combination with interferon-α, synergistic anti-HCV activity was observed. The anti-HCV mechanism of sodium stibogluconate is still not clear. Whereas, the PUFAs were found to increase cellular lipid oxidative products exerting anti-HCV activity. PUFAs also abolish NS3/4A activity in vitro. Results in this study might be helpful for discovery of effective medicine to combat HCV infection.
目
錄
誌 謝
Viii
中文提要
ix
英文提要
xi
Chapter 1 Overview of Hepatitis C Virus (HCV) 1
1.1 HCV Viral Biology 1
1.1.1 Core (C) Protein 2
1.1.2 Envelope Proteins:E1 and E2 3
1.1.3 Nonstructural Protein 2 (NS2) 4
1.1.4 Nonstructural Protein 4 (NS3) 4
1.1.5 Nonstructural Protein 4 (NS4A and NS4B) 6
1.1.6 Nonstructural Protein 5 (NS5A and NS5B) 7
1.1.7 HCV subgenomic Replicon System 9
Chapter 2 Antiviral Targets of HCV 11
2.1 General reviews 11
2.2 Antiviral drug Screening Using HCV subgenomic replicon system 12
2.3 NS3 as an antiviral target 13
2.4 RNA-dependent RNA polymerase (RdRp ; NS5B ) as an antiviral target 14
Chapter 3 Aims of this Dissertation 16
Chapter 4 Materials and Methods 17
4.1 Reagents 17
4.2 Plasmid constructions and generation of lentiviral vectors 17
4.3 Isolation of stably transfected Ava5 cells expressing EG(∆4AB)SEAP 18
4.4 Detection of SEAP activity released from Ava5-EG(∆4AB)SEAP cells 18
4.5 Cell culture 19
4.6 Assay for inhibition of HCV subgenomic RNA 20
4.7 Cytotoxicity assay 20
4.8 Western Blotting 21
4.9 Synergistic statistics 22
4.10 Sustained anti-HCV response after drug removal 23
4.11 Northern Blotting 23
4.12 Lipid Peroxidation Assay 24
Chapter 5 Inhibition of Hepatitis C Virus Replication by Antimonial Compounds 25
5.1 Results 26
5.1.1 Effects of antimonial compounds on HCV replication and cellular toxicity 26
5.1.2 Synergistic anti-HCV activity of SSG and IFN-α combination 27
5.1.3 Sustained anti-HCV response after drug removal 28
5.2 Discussion 30
Chapter 6 Anti-HCV Activities of Selective Polyunsaturated Fatty Acids 33
6.1 Results and Discussion 34
6.1.1 Effects of PUFAs on HCV RNA replication 34
6.1.2 Arachidonic acid reduced HCV RNA level dose-dependently 34
Chapter 7 Dual Action of Inhibition of Hepatitis Virus C Replication by
Arachidonic Acid 37
7.1 Results and Discussion 40
7.1.1 Inhibition of HCV replication by AA does not involve new gene
expression 40
7.1.2 Lipid peroxidation in AA-induced anti-HCV activity 41
7.1.3 Suppression of HCV replication in replicon cells by 4-hydroxynonenal
(HNE), one of AA’s oxidative products 42
7.1.4 Inhibition of NS3/4A protease activity by AA in vitro 44
Chapter 8 Conclusion remarks and Future Works 47
Chapter 9 References 49
Chapter 10 Tables and Figures 71
Chapter 1: Overview of Hepatitis C Virus (HCV)
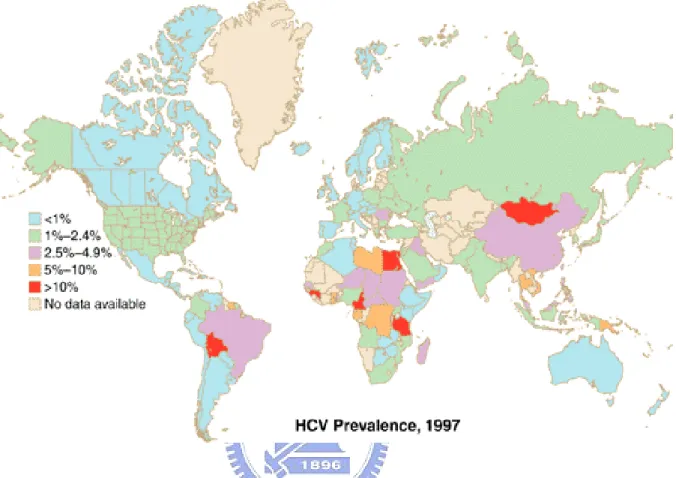
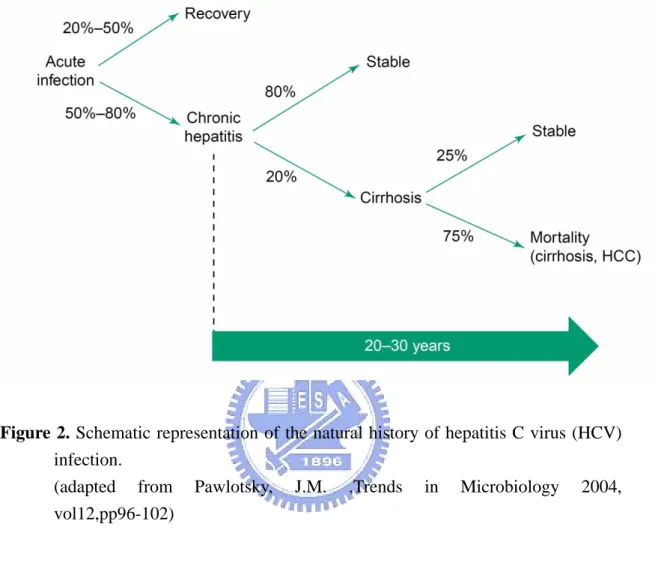
HCV, a member in the Flaviviridae family, was identified in 1989 as the causative agents of non-A, non-B hepatitis (29). As estimated by the World Health Organization (WHO), approximately 170 million, that is ~3% of the worldwide population were infected with HCV(1, 197). In Taiwan, approximately 3.5% population suffer from chronic HCV infection, as announced by CDC in June 2005. The global distribution of HCV infection was shown in Figure 1. About 70% of acute hepatitis C virus infection results in chronic infection. About 20% of chronic HCV patients would develope liver cirrhosis within 20 years. Approximate 1-5% of established cirrhosis cases would lead to hepatocellular carcinoma (HCC) per year (142, 164) as shown in Figure 2. The disease sequel is promoted by other factors such as age, alcohol abuse, HCV genotypes, and co-infection with other viruses such as hepatitis B virus (HBV), Epstein Bar virus (EBV), cytomegalovirus (CMV), and human immunodeficiency virus type I (HIV-1) (122).
1.1 HCV Viral Biology
HCV is a positive-stranded, enveloped RNA virus whose virion is about 60 nm in diameter. Its genome is ~9,600 nucleotides in length containing an open reading frame (ORF) encoding a polyprotein of ~3,000 amino acid residues (9, 62, 93, 143). The ORF is flanked by 5’ untranslated regions (UTR) close to 340 nucleotides and 3’ UTR of approximate 230 nucleotides. The HCV 5’ UTR contains a highly conserved internal ribosomal entry site (IRES), which serves for mediating cap-independent translation.
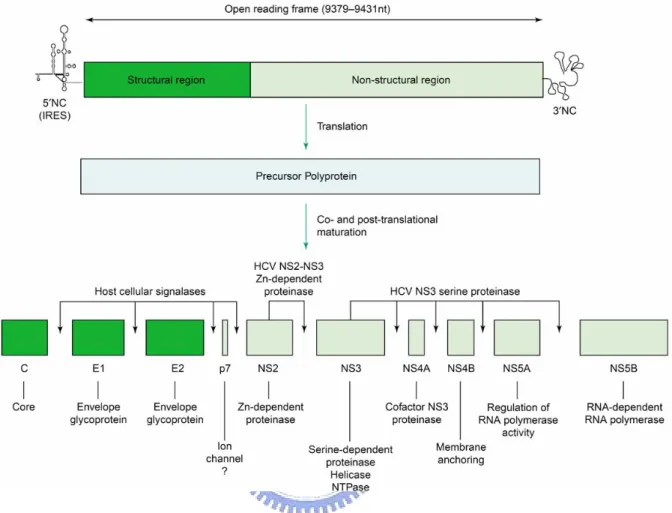
HCV IRES has three stem-loops with a pseudoknot neighbor on the 3’ boundary of the stem-loop III. This highly conserved pseudoknot plays a critical role for IRES activity. The 3’ end of HCV RNA contains three distinct regions, a variable region, a poly(U) segment, and a highly conserved 98 nucleotides X region. The poly(U) stretch and X region are critical elements for HCV replication in vivo. The mature viral proteins are cleaved by host signal peptidases and viral proteases into about 10 viral proteins: the core (C), two enveloped glycoproteins E1 (E1) and E2 (E2), p7, nonstructural protein 2 (NS2), NS3, NS4A, NS4B, NS5A, and NS5B (figure 3). C, E1, and E2 are structural proteins and others are non-structural proteins. Viral RNA genome is encapsidated with core protein and packed into viral envelope glycoprotein complex E1-E2 to form viral particles. Due to lack of an efficient in vitro culture system and a convienient small-animal model for HCV study and very low titers in clinical samples, the mechanisms of HCV replication and pathogenesis are still not clear.
1.1.1 Core Protein
HCV core protein, located on the N-terminus of HCV polyprotein, is released by host signal peptidase (126, 157). Core protein is the viral capsid protein and has DNA and RNA binding activity (157, 158). HCV core protein is cytoplasmic, located around some lipid vesicles on endoplasmic reticulum membranes. HCV core protein can modulate cellular signal transduction pathways (101, 102). Recently, many reports reported that HCV core protein is a cell growth and viablility modulator (193). The HCV core protein promote primary hepatocyte immortalization (159) and upregulate cyclin E expression (27) and perturbate the cell cycle of hepatobalstoma cell line (163).
The hepatitis C virus core protein plays an important role for the hepatocellular carcinoma (HCC) formation (69, 171). Oxidative stress response induced by HCV is one of the important issues of HCV-induced hepatocarcinogenesis. Huh-7 cells express HCV core protein and increase cellular reactive oxygen species (ROS) (111) and injure hepatocyte mitochondria (138). Therefore, HCV core protein induce oxidative injury may serve a role for HCV pathogenesis.
1.1.2 Envelope Proteins:E1 and E2
HCV enveloped glycoprotein E1 and E2 assemble into heterodimer and are the main components of viral envelope (139). The E1 and E2 are type I transmembrane
proteinswith a large N-terminal ectodomain and a C-terminal hydrophobicanchor and
they are N-linked glycoproteins that contain multiglycosylation sites. E1 is a 31 kDa protein, with about 6 potential N-linked sites, whereas E2 is an approximately 65-70 kDa glycoprotein, with about 11 potential N-linked sites (65, 139). These proteins play important role in virus entry and assembly. Using in situ immunoloblot study and glycan analysis, HCV envelope protein E1 and E2 are located in endoplasmic reticular (ER) membrane. They are necessary for the formation of the HCV viral particle.
Virus infection begins with interaction between the virion and the surface of the host cell. Virus envelope proteins are important in the early steps of the viral life cycle. HCV envelope E1E2 heterodimer complex is very important for HCV infection. Low density lipoprotein (LDL) receptor, scavenger receptor class B type 1 (SR-B1), glycosaminoglycans, and CD81 are putative receptors for HCV infection (11, 12, 44, 54,
55, 168). The CD81 cellular surface protein has been demonstrated interactive with HCV E2 protein that plays an important mediator for HCV binding to host cell. Use of the infectious HCV pseudotype particle as a model for studying HCV binding and entry host cell, CD81 plays as a coreceptor for HCV entry host cell (35, 211).
1.1.3 Nonstructural Protein 2 (NS2)
The nonstructural protein 2 (NS2), a 23-kDa protein, is one of viral proteases, which is required for NS2-NS3 cleavage and separate NS2 protein from the viral polyprotein precursor. This nonglycosylated integral membrane protein (165) is not essential for the virus replication in the replicon system (19, 117). NS2 protease activity is inhibited by alkylating agents and EDTA. The EDTA could abolish NS2 activity that
can be reverse by addition ZnCl2 and CdCl2 (147). The amino acids His-143 and
Cys-184 are essential for NS2 protease activity which has been demonstrated by site-directed mutagenesis (68). The NS2 protein not only has proteinase activity but also has apoptosis inhibition (51) and gene transcript modulation (45) activity. The NS2 protease is phosphorylated by protein kinase CK2 and degraded by proteosome (58).
1.1.4 Nonstructural protein 3 (NS3)
The nonstructural (NS) proteins are generated by co- and post-translational cleavage of the polyprotein by two virally-encoded proteases (10). One is NS2 metal-dependent protease and the other is NS3-NS4A serine protease complex. NS4A is cofactor for the NS3 protease and stabilizes NS3/NS4A complex. The HCV NS3/NS4A chymotrypsin-like serine protease is required for the maturation of the viral polyprotein,
cleaving it at the NS3-NS4A, NS4A-NS4B, NS4B-NS5A, and NS5A-NS5B junction sites. NS3 viral protein contains 631 amino acid residues, approximately 70 kDa, and contains two functional domains: the serine protease domain is located at the N-terminal one-third(72) of NS3 viral protein. The susequcent two-thirds in the NS3 encodes a NTPase/Helicase domain (96, 205). The NS3 viral protein is distributed in the cytoplasm and nucleus when expressed alone, but when co-expressed with NS4A, the majority of NS3 protease is then found on the ER membrane (200). The NS4A N-terminal hydrophobic domain forms a transmembrane (TM) segment which is required to target and to anchor NS3 to ER membrane. X-ray crystallography (97, 203) and NMR spectroscopy studies (124) have been determine the three-dimensional structure of the NS3/NS4A protease. The structure of the NS3 serine protease domain include a chymotrypsine-like fold, with two six-stranded β-barrel subdomains (41, 70). Similar to other trypsin-like serine proteases, NS3/NS4A is made of two domains both composed of a β-barrel and two short α-helices. The catalytic triad is formed by residues in the two β-barrels of the same loops: His-57 and Asp-81 N-terminal β-barrel and Ser-139 in the
C-terminal β-barrel. The NS3/NS4A serine protease requires Zn++ for activity and
structure stability (39). The tetrahedral Zn++ binding site is formed by residues Cys-97, Cys-99, Cys-145 and His-149.
The NTPase/Helicase activity of NS3 viral protein is another important function for viral replication. The amino acids in the NS3 helicase domain is highly conserved between various HCV strains with more than 80% sequence identity. The NS3 NTPase/Helicase domain have identified their multiple roles in RNA binding,
RNA-stimulated NTPase activity, and unwinding of RNA regions with secondary structure and NTP hydrolysis by using mutagenesis and structural analysis. For replication HCV use positive-strand RNA template to synthesis the negative-strand. Because the positive and negative RNA strands are complimentary, the strand separation requires NS3 helicase activity. Other functions NS3 helicase in HCV replication include relax the positive-strand RNA to increase translational efficiency or faciliatate access of highly stable secondary structures such as IRES element and the 98-bp 3’X element (99, 185).
In addition to the functions of NS3 viral protein in the processing of the HCV polyprotein and HCV replication, NS3 was shown to affect intracellular signal transduction mediated by PKA and PKC (20-22). PKC and PKA catalytic sites interact with NS3 and results in inhibition of kinase activity in vivo and in vitro. The intact
NH2-terminus of NS3 carry out an important regulatory function in the
hyperphosphorylation of NS5A p56 and generation of its p58 form (133). The presence of the authentic N-terminus may be necessary for directed NS3 and NS5A interaction. The mechanisms of how NS3 promote NS5A hyperphosphorylation remain unclear.
1.1.5 Nonstructural proteins 4 (NS4A and NS4B)
HCV NS4A is a small hydrophobic protein containing 54 amino acid residues, which acts as cofactor for NS3 protease activity by forming stable complex with NS3 protein and anchoring this complex on the ER membrane as an essential component for the HCV polyprotein processing (84, 97). Moreover, the interaction of NS4A with NS3
will promotes the interaction of NS3 to NS5B. On the other hand, the NS4A may bind directly to NS5B and enhance the polymerase activity, with need to be elucidated in future.
The NS4B protein has 261 amino acid residues for HCV genotype 1b and its molecular weight is about 30 kDa. The functions of NS4B are still not clear. Some reports have pointed out that NS4B is highly hydrophobic and plays a role in the association of HCV replication complex to the ER (112). To determined localization and topology of NS4B protein by using recombinant HCV NS4B constructs. The protein was localized in the endoplasmic reticulum (ER), but also induced a pattern of cytoplasmic foci positive for markers of the ER. Computer predictions of the membrane topology of NS4B suggested that it has four transmembrane segments (120). Introduction of site-directed mutations demonstrate N-terminal alpha-helix in NS4B is very important for HCV replication (48). Kate et al. pointed out that HCV NS4A and NS4B proteins have an effect on translational inhibition (92). It will help HCV virus infection and survival in a host cell.
1.1.6 Nonstructural proteins 5 (NS5A and NS5B)
NS5A protein has 447 amino acid residues which is phosphorylated and membrane-associated protein of unclear structure and functions. Many studies have shown that NS5A plays an important role in regulating interferon-induced cellular antiviral response (26, 46, 60, 80, 131, 136, 148, 150, 167, 188, 189). The NS5A protein inhibit eIF2-α and eIF4E phosphorylation to block cellular translation (75). NS5A was
also found to associate with NS5B, the RNA-dependent RNA polymerase, RdRp, and modulate their activity (173). Sequence analysis of NS5A reveals that NS5A contains the interferon-sensitive determining region, called ISDR (50, 61), which is important for viral resistance to interferon induced anti-viral effect (182). NS5A is a serine rich protein and can be phosphorylated on the multiple sites (94, 161, 182). The hyperphosphorylated form of NS5A protein shifts molecular weight from 56 kD to 58 kD (78, 133). Double-strand RNA is an intermediate product of viral replication and activates specific transcriptional factors, including IFN-α regulatory factors, IRF-1 and IRF-3. Jill Pflugheber (145) has shown that NS5A could influence HCV persistence by blocking IRF-1 activation and disrupt host antiviral pathway. PKR is induced by interferon to play the antiviral and antiproliferative activities. The inductuion of NF-κB by double-strand RNA dependent protein kinase is mediated by IKK activation (210).
The terminal non-structural HCV viral protein is called NS5B, which is a 68 kDa protein and is identified as the RNA-dependent RNA polymerase (RdRp) that play a central role in virus replication. The HCV RdRp mediated insertion of sequence crosses the ER phospholipids bilayer as a TM segment has been demonstrated recently (85). By double label immunofluorescence analyses (169), NS5B was found in the endoplasmic reticulum (ER) when expressed alone or in the context of the entire HCV polyprotein. The carboxyl-terminal 21 amino acid residues were necessary and sufficient to target NS5B to the cytosolic side of the ER membrane. This hydrophobic domain is highly conserved among 269 HCV isolates analyzed and predicted to form a transmembrane alpha-helix. Based on a structural model and the amino acid conservation among
different HCV isolates, Moradpour et al. (127) have designed a panel of insertion sequence mutants and analyzed their membrane association and RNA replication. They demonstrated that the membrane association of the RdRp is essential for HCV RNA replication. The crystal structure of NS5B (24, 108) showed a catalytic center followed by a C-terminal extension that connects to the TM segment via active-site groove. Most substitutions of these conserved motif residues severely reduced enzymatic activity (116). Oligomerization of NS5B might be important for modulating the activity of polymerase (154). Indeed, NS5B has been demonstrated to directly interact with NS3, NS4A, and other components of the replication complex (77, 84, 146, 173), and these interactions regulate RdRp activity (146). Otherwise, recombinant RdRp protein expressed in insect cells using baculovirus vector (17, 116) or E. coli (3, 186, 202, 209) showed RdRp activity could be assayed in vitro. This is very important for setting up an in vitro assay system of RdRp activity for evaluation of new drugs against HCV.
1.1.7 HCV subgenomic Replicon System
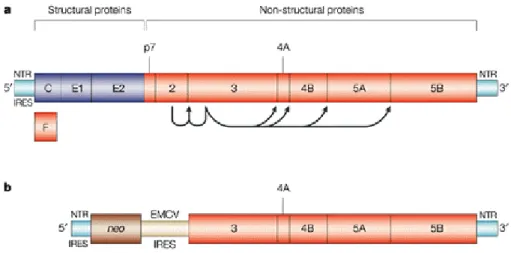
HCV was identified in 1989 but study of the mechanism of HCV replication have been hampered due to the lack of an efficient animal model and culture system. Not until 1999, Lohmann, et al.(117) developed the HCV subgenomic replicon system for studing the HCV replication in cultured cells. The HCV replicon is a bicistronic RNA molecule, which contains HCV IRES in front of neomycin phosphotransferase gene. The second cistron in the RNA molecule contains an encephalomyocarditis virus (EMCV) IRES, followed by the HCV nonstructural proteins (NS3 through NS5B) and terminating with the HCV 3’ UTR (figure 4). Replicon RNA molecules are maintained during numerous
passages after Huh-7 cells are transfected with in vitro transcripted HCV replicon RNAs. The benefit of the HCV subgenomic replicon system is that it is easy to detect viral genome by nucleic acid technology such as RT-PCR and Northern blot because of high level of viral RNA replication. Since the structural proteins (Core, E1 and E2) are not included in the subgenomic RNA construct, no virion can be formed. Thus, the HCV replicon system only partially recapitulated the entire viral life cycle in nature. Recently, important progress has been made to provide an efficient HCV cell culture system for studing HCV. This novell HCV culture system relies on the role of a clonal viral genome. (113, 194, 213).
Chapter 2: Antiviral targets of HCV
2.1 General Reviews
Virus binding, internalization, cytoplasmic releasing and uncoating, IRES-mediated translation, polyprotein processing, virion packing, assembly, maturation, and release of infectious particles protease are the important steps for in the viralreplication life cycle. All steps of virus life cycle are the targets for anti-viral drug discovery. The complete HCV life cycle is difficult to investigate due to the lack of a suitable in vitro and in vivo model system for HCV infection. Inadequate virion productions for study are the main problems of the model system.
IRES at the 5’ region of HCV RNA genome is also a target for anti-HCV drug discovery (183, 195). Ribozyme and antisense oligonucleotides for viral RNA are currently being evaluated in clinical trials (183). RNA interference technologies are used for specific purposes against HCV viral genome recently (66, 100, 155, 199, 208). HCV E2 glycoprotein is very important for viral attachment to host cell, which is another potential anti-viral target. Monoclonal antibody against HCV E2 protein has been assessed in clinical phase Ib (183). Until the subgenomic replicon system has been developed (19, 117), partial HCV life cycle could not be studied in vitro. Although the subgenomic replicon system could help us to observe highly viral RNA replication, no virion could be obtained.
NS3 viral protease and NS5B RdRp are viral specific proteins and has been conserded as good candidates as anti-viral targets. Several biochemical and cell-based
assay systems have been established to identify selective inhibitors, for NS3/4A protease and NS5B polymerase. The success of protease and polymerase inhibitors in controlling human immunodeficiency virus (HIV) infection justfies the conviction that the HCV NS3 and NS5B be the choice of drug targets. Moreover, the combination therapy of protease and reverse transcriptase inhibitors has been observed the most effective treatment against HIV infections. Structure-activity relationship (SAR) studies and in-depth investigations on biochemical property of target proteins provided the necessary information for designing inhibitors and for optimization of their activity. Indeed, a NS3 protease inhibitor and a NS5B inhibitor have reached phase II clinical studies (183). The antiviral targets of the HCV have been reviewed recently (40, 176, 201). We’ll majorly address on replicon system, NS3/4A viral protease and NS5B polymerase.
2.2 Antiviral drug Screening Using HCV Subgenomic Replicon System
The HCV subgenomic replicon system facilitates a robust anti-HCV drug screening plateform. Numerous studies report the discovery of novell anti-HCV compounds/drugs based on the HCV subgenomic replicon system (109). The HCV replicon system enables high-level replication of viral subgenomic RNA. Although the life cycle of the HCV could not be fully evaluated by this system, informations on HCV replication and host factors involved in HCV replication could be obtained. The disadvantage of screening drugs based on the replicon system is that compounds identified need to be further investigated for their modes of actions.
2.3 NS3/4A as an antiviral target
NS3 viral protein is a multifunctional enzyme, which contains a protease and a helicase/NTPase domain. The N-terminal one-third part, adjacent to the C-terminus of NS2, is the protease domain and the other two-third is the helicase/NTPase domain. Both the protease activity and helicase activity are commonly considered as targets for anti-viral drug discovery. Many assay systems of both enzyme activities have been well established for screening of inhibitors. Inhibitors could be identified through molecular modeling based on the NMR and X-ray crystallographic studies (144, 149). Three strategies for developing inhibitors of the HCV NS3/4A protease have been attempted: abolishing the NS3/NS4A interaction, interfering with zinc binding and preventing substrate binding in the active site. Typically, the serine protease inhibitors derived from its own substrates by substitution of the scissile amide bond with an electrophilic moiety is able to form a covalent adduct with the catalytic serine residues (47). The penta- or hexapeptides NS3 protease inhibitors derived from the N-terminal NS3 cleavage products were named peptidomimetic inhibitors (83, 140, 212). The biopharmaceutical classes have been reported that structurally diverse NS3-protease substrate-based inhibitors including α-ketoamide (73, 74), Pyrrolidine-5,5-trans-lactams (4, 174, 175), azapeptides (6, 212), α-ketoacids (33, 132, 137), and boronic acids (153). VX-950 (LY-570310) is a lead NS3-protease inhibitor compound and is in preclinical stage (183) that is identified by using structure-based computational and combinatorial-chemistry techniques. Using structure-activity relationship study another important type of peptidomimetic inhibitors derivative was found from the N-terminal cleavage products by NS3-protease (114). BILN 2061 is a macrocyclic peptidyl
carboxylic acid (53, 115) that is a very potent competitive inhibitor of the NS3/4A protease. BILN 2061 has good bio-availablity and inhibits HCV replication in cell-based assay (103, 162). BILN 2061 was at phase I clinical trials without serious adverse events and did not damage liver enzymes, only minor gastrointestinal disturbances were observed. These results indicate that BILN 2061 may be safe and effective in the treatment of hepatitis C infection. However, the BILN 2061 resistant mutant of the HCV has been found (141). Only one amino acid change in the NS3 protease region would result in the development of resistant strain..
2.4 RNA-dependent RNA polymerase (RdRp ; NS5B ) as an antiviral
target
Viral polymerases are attractive targets for antiviral therapy as demonstrated by the clinical success of nucleoside and non-nucleoside inhibitors of HBV and HIV replication. According to the chemical structure and mechanism of action, three categories of the viral polymerase inhibitors could be classified (38): (i) nucleoside analogues (49), (ii) non-nucleoside inhibitors (15, 16, 79, 166), and (iii) pyrophosphate mimics (180). Nucleoside analogues are substrate analogues that need to be phosphorylated to their corresponding nucleotide in cytoplasm of infected cells. Then, the prodrug becomes active against the viral polymerases. The nucleotide could be incorporated by the polymerase during progressive gene synthesis, leading to early termination of the elongation reaction and thus inhibition of the viral replication. HCV NS5B polymerase is also an anti-HCV target (201) since the NS5B activity is essential for HCV viral replication. The biochemical properties (110, 119, 214) and crystal structures (24, 108) of
NS5B have been characterized. HCV NS5B uses di- or tri- nucleotides to initiate RNA replication and forms replication complex at 3’-end of HCV RNA genome. The replication complex contains NS5B polymerase, template, primer, nucleotides, and other associated factors. NS5B polymerase forms complex with nucleotides is needed for de novo initiation (23, 156). De novo initiation must then be followed by RNA elongation, termination of polymerization and release of nascent strand. In principle, each of these steps could be seen as a target for anti-viral therapy. SAR studies have led to the identification of both catalytic and regulatory nucleotide binding site in the HCV RdRp (160) such information is essential for the design of novel nucleotide analogues inhibitors.
.
Benzimidazole and bezothiadiazine derives are the two major classes of the non-nucleoside type inhibitors of NS5B polymerase. These inhibitors comprise structurally heterogenous compounds, which usually bind to a site on the enzyme surface away from active site, such as an allosteric site (191). These two structural classes of inhibitors abolish the NS5B activity by binding to different allosteric interaction sites (192). A series of diketo acids were reported to selectively and potently inhibit the HCV NS5B polymerase elongation activity in vitro (181). The mechanism of action of these related compounds was found to be noncompetitive with respect to both the RNA template and to the nucleotides.
Chapter 3 : Aims of this Disertation
Interferon-α (or PEGylated interferon-α) plus ribavirin is the only recommended treatment for HCV currently. Severe adverse effects and low curing rate are problematic for the IFN-based treatment. Thus, it is important to discover more effective and specific antiviral drugs for HCV therapy currently. In this report, we utilize the HCV subgenomic replicon system to investigate and to screen the in-house collected and commercially available chemical liberaries for novel anti-HCV compounds. We found that antimonial compounds, sodium stibogluconate (SSG), and polyunsaturated fatty acids (PUFAs) could inhibit HCV replication in replicon system. Moreover, combination treatment of these agents with interferon-α on the replicon system exerted synergistic anti-HCV activity. Thereafter, we investigated the anti-HCV mechanisms of these anti-viral agents and found that the cellular oxidative status induced by arachidonate correlates with its anti-HCV activity. We then used the oxidative product of the arachidonate (HNE) to treat the replicon cells and found that HNE exhibits anti-viral activity in a dose-dependent manner. On the other hand, we found that arachidonate could inhibite NS3 protease in vitro. The results of the current study might prove to be helpful for novel treatment against HCV infection.
Chapter 4:Materials and Methods
4.1 Reagents
Dulbecco's modified Eagle's media (DMEM) high glucose, Fetal Calf Serum (FCS), TRIZOL™ Reagent, and G418 (geneticin) were purchased from Invitrogen (Carlsbad, CA). Eicosatetraynoic acid (ETYA), 8-iso prostaglandin E2 (8-iso PGE2), and 8-iso prostaglandin F2α (8-iso PGF2α) were obtained from Cayman Chemical Co. (Ann Arbor, MI). MK-886 was purchased from Calbiochem (EMD Bioscience, San Diego, CA). arachidonic acid (AA; 20:4, n6), docosahexaenoic acid (DHA; 22:6, n3), eicosapentaenoic acid (EPA; 20:5,n3), α-linolenic acid (18:3,n3), γ-linolenic acid (18:3,n6), and linoleic acid (18:2,n6) , oleic acid (18:1,n9), myristic acid (14:0), palmitic acid (16:0), and Steric acid (18:0), actinomycin D, vitamine C, vitamine E, indomethacin, ibuprofen and 1-aminobenzotriazole (1-ABT) were obtained from Sigma-Aldrich Inc. (St. Louis, MO). Trolox was obtained from BIOMOL Research Laboratories Inc. (Plymouth
Meeting, PA). The α-P32-dCTP was purchased from Amersham Bioscience Corp.
(Piscataway, NJ). Human hepatoma cells (Huh-7) was purchased from Japanese Collection of Research Bioresources (JCRB, JCRB0403) and Huh-7 cell clone containing HCV replicon (Ava5) was provided by Apath, Inc. (St. Louis, MO).
4.2 Plasmid constructions and generation of lentiviral vectors
The generation of pLenti-EG(∆4AB)SEAP plasmid has been described (JVM, 2004 paper). In short, the forward primer, 5’-CCA CCG CCA CCA TGG TGA GCA AGG
GC-3’, and the reverse primer, 5’-TCA TGT CTG CTC GAA GCG GCC-3’, were used. PCR products was then inserted into pLenti6/V5-TOPO plasmid (Invitrogen, Carlsbad, California, USA). Recombinant lentiviral vector was generated by a transient plasmid transfection as described (43). The 293FT cells were co-transfected with
pLenti-EG(∆4AB)SEAP and ViraPowerTM Packaging Mix using Lipofectamine 2000
according to manufacturer’s instructions. Two days after transfection, lentiviral particles were collected from conditioned medium and cellular debris was cleared off by low-speed centrifugation.
4.3 Isolation of stably transfected Ava5 cells expressing
EG(∆4AB)SEAP
In 24-well plates, Huh-7 or Ava5 cells were seeded at a density of 2 × 104 cells per
well. After incubation at 37℃ overnight, cells were transduced with serial dilutions of lentiviral particles in the presence of 6 µg/ml polybrene (Sigma, St. Louis, MO, USA). After incubation for 24 h, virus-containing medium was replaced by fresh medium and the cells were incubated for another 2 days. Clonal cells surviving under blasticidin selection were maintained carefully. Cells were examined for expression of EG(∆4AB)SEAP by Western blot analysis and extracellular SEAP activity. Positive cell lines were thus produced and designated as Ava5-EG(∆4AB)SEAP.
4.4 Detection of SEAP activity released from Ava5-EG(∆4AB)SEAP
cells
mg/ml G418 and 10 µg/ml blasticidin. Cells were seeded in six-well plates at a density of 5×104 cells per well. After incubation at 37 ◦C for 1 day, cells were treated with IFN-α (0,0.78, 1.56, 3.13, 6.25, 12.50 and 50.00 IU/ml). IFN-α was obtained from Sigma (St. Louis, MO, USA). Two days later, culture medium was replaced with fresh phenol red-free DMEM/10% FBS containing the same concentration of drugs and cells were incubated for one more day. Culture media were collected from each well and SEAP activities were measured using Phospha-Light assay kit (Tropix, Foster City, CA, USA) according to manufacturer’s instruction.
4.5 Cell culture
Dulbecco’s modified Eagle’s medium (DMEM) high glucose, Fetal Calf Serum (FCS), G418 (geneticin) and blasticidin were purchased from Invitrogen (Carlsbad, CA). Human hepatoma cells (Huh-7) and HCV sub-genomic replicon cells were obtained from Apath, LLC (St. Louis, MO) (19). A reporter-based cell line, Ava5-EG(∆4AB)SEAP (105, 106), for HCV drug screening was derived from HCV replicon cells (Ava5). EG(∆4AB)SEAP is a reporter gene consisting of enhanced green fluorescent protein (EG), the NS3-NS4A protease decapeptide recognition sequence (∆4AB), and secreted alkaline phosphatase (SEAP). In the reporter cell line, a reporter gene, EG(∆4AB)SEAP was stably integrated in the Ava5 cells to generate Ava5-EG(∆4AB)SEAP cells. In Ava5-EG(∆4AB)SEAP cells, SEAP activity in the culture medium can be used to reflect anti-HCV activity (105). Cells were maintained in a humidified atmosphere containing
5% CO2. For Ava5 and Ava5-EG(∆4AB)SEAP cells, the culture medium was
blasticidin, respectively, to maintain selection pressures for sustaining the expression of exogenous genes.
4.6 Assay for inhibition of HCV subgenomic RNA
Ava5-EG(∆4AB)SEAP cells were seeded in 96-well plates at a density of 5×103 per well.
After incubation for 1 day, cells were treated with drugs for 48 h. At the end of incubation, the culture medium was replaced with fresh phenol red-free DMEM containing 10% FBS and the same concentration of drugs. Incubation was continued for one additional day. Culture media were collected and SEAP activities were measured using the Phospha-Light assay kit (Tropix, Foster City, CA) according to the supplier’s instruction. IFN-α, Sb2O3, and SbCl5 were from Sigma-Aldrich (St. Louis, MO) and
SSG (21 % [wt/wt] SbV) was from Wuhan Shengmao (Hubei, China,
http://my.ecplaza.net/Wuhancorp/).
4.7 Cytotoxicity assay
Cell viability was determined by the MTS assay essentially as described (82). In short, for a 96-well microtiter plate, 10 ml of reagent containing phenol red-free DMEM, MTS (tetrazolium compound [3-(4,5-dimethylthiozol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4- sulfophenyl)-2H-tetrazolium, inner salt]; Promega, Madison, WI) and PMS (phenazine methosulfate; Sigma, St. Louis, MO) in a ratio of 80:20:1, respectively. The mixed reagent was distributed to cells (100 µL/well). The plates were incubated for 1 – 4 hrs
490 nm. Compounds were also analyzed for their effects on cell cycle distribution. In this assay, cells were seeded in 6-well plates at the density of 1×106 cells per well, and
incubated in various concentrations ofSSG for 24 hrs. After drug treatment, adherent cells were harvested by trypsin digestion. Cells were then centrifuged, washed once in PBS, and resuspended in 200 µL phosphate buffered saline (PBS). Cells were then slowly added to 5 mL of ice-cold 70% ethanol, and stored at -20°C until analysis. Fixed cells were collectedby centrifugation, washed twice with PBS, resuspended in 1 mL of a solution containing 3.4 mM sodium citrate, 20 µg/mL propidium iodide, and 100 µg/mL RNase A, and stored in the dark for 1 hr. Cells were analyzed using a FACSVantage flow cytometer (Becton Dickinson Labware, Franklin Lakes, NJ). Cell cycle analysis was performed according to the mathematical model of Jett (87).
4.8 Western Blotting
Western blotting was performed essentially as described (105). Anti-HCV NS3 and anti-actin antibodies were obtained from LTK Biolaboratories (Taipei, Taiwan) and CHEMICON International Inc. (Temecula, CA), respectively. Briefly, Cells were seeded in 6 cm plate for two days at a density of 1×106 cells pre plate. Sequentially, cells were scrapped and collected by centrifugation and washed with phosphate buffered saline (PBS) and then lysed with lysis buffer containing 20 mM Tris (pH 7.5), 150 mM NaCl, 1 % Triton X-100 and protease inhibitor cocktail (Roche Diagnostic GmbH, Mannheim, Germany). The cell lysates were clarified by centrifugation and equal amounts of protein samples were applied to 10 % SDS-polyacrylamide gel electrophoresis. Proteins in gel were then transferred to Hybond-C membrane (Amersham Biosciences) using
semi-dry method (BioRad, Hercules, CA, USA). The membrane was blocked with 5 %
skim milk in Tris-buffered saline (TBS) with 0.1 % Tween 20 overnight at 4 and ℃
stained with rabbit polyclonal anti-NS3 antibody (LTK Biolaboratories, Taipei, Taiwan) or mouse monoclonal anti-actin antibody (CHEMICON International Inc. (Temecula, CA)). Bound antibody was further probed with horseradish peroxidase-conjugated anti-rabbit IgG (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) or horseradish peroxidase-conjugated anti-mouse IgG and Signals were revealed by the ECL Western blotting system (Amersham Biosciences, Piscataway, NJ) and detected by autoradiography.
4.9 Synergistic statistics (104)
The isobologram analysis was used to evaluate the effects of combined drug treatments (30, 31). Traditional isobologram analysis is a frequently used method for analyzing the effects of multiple drugs and for determining their additivity, synergism, or antagonism. Various doses of SSG and IFN-α were combined in a checker board manner to generate dose-response curves (isoboles) of 50% and 80% inhibition of HCV replication to evaluate the effect of drug combination. The synergism between two drugs was quantified by combination indices (CI) using the CalcuSyn computer program (Chou & Hayball, CalcuSyn for Windows, Biosoft, Cambridge UK, 1996) (30). CI values < 1 indicate a synergistic effect; CI = 1 an additive effect, and CI > 1 an antagonistic effect. The evaluation of drug combination based on a median-effect equation has been widely employed in the literature. On the other hand, cell was treated with arachidonic acid (mock, 0.78, 3.125, 12.5, 50, 200 µM) and interferon-α (mock, 0.78, 3.125, 12.5, 50, 200
IU/mL) used in a range of combination. Response surface plot for inhibitory
concentration for each combination was analyzed by sigmoid regression. EC90 (= 90%
inhibition effect of HCV replicon RNA) and EC50 (= 50% inhibition) used for plot. The
total RNA from each combination was isolated by TRIZOL reagent and analysed using Northern blot. The statistics data of each combination was triplicate, means ± SD using SigmaPlot.
4.10 Sustained anti-HCV response after drug removal
Ava5 cells containing HCV subgenomic replicon RNA were plated in 15-cm culture dishes at a density of 2.0 x 106 cells per dish. During this experiment, cells were treated
with the indicated drugs for 5 days. The culture medium containing drugs was
subsequently removed, and the cells were cultured in fresh medium containing 10% FBS
and1 mg/mL of G418 without drug. Cells were then allowed to grow for 1 month.
Surviving cells were visualized by staining with crystal violet.
4.11 Northern Blotting
Total RNA was isolated from cells using TRIZOL™ Reagent (one-step, guanidium
thiocyanate phenol-chloroform total RNA isolation reagent) per supplier’s instruction. RNA was isolated and concentration was determined by spectrophotometer. All reagents used for analysis are of ultra-pure grade. RNA samples were loaded onto 1 % TBE agarose gel, 10 µg each well, and separated by electrophoresis at 10 V/cm for 1.5 h according to Kevil et al. (95). The RNAs in the gel were then transferred to a positively
(Vacu. GeneXL, Pharmacia, MI). After drying, RNA was then cross-linked to the
membrane by UV irradiation using UV Stratalinker® 1800 (Stratagene, CA). The
membrane was probed separately with the NS5B gene fragment of HCV and human glyceraldehydes-3-phosphate dehydrogenase (GAPDH) fragment labeled with
[α-32P]dCTP by rediprimeTM II random prime labeling system (Pharmacia, MI) in
accordance with manufacturer’s instructions. Hybridization was carried out with denatured probes in Rapid-hyb hybridization buffer (Pharmacia) for 2 h at 65°C. After hybridization, membranes were washed once in 2X SSC-0.2% SDS for 20 min at 60°C and once in 1X SSC-0.2% SDS for 20 min at 60°C and twice in 0.1XSSC-0.2% SDS for 15 min at 65°C. The results of hybridisation were visualized by autoradiography. The Northern blotting procedure employed in this study is essentially the same as that has been used in previous studies (106, 109).
4.12 Lipid Peroxidation Assay
Ava5 cells (2x106) were seeded into 10-cm petri dish overnight, and then treated with different concentration of AA for 24 h. The cells were washed with phosphate buffered saline (PBS) twice and scraped off the culture plate. Cells were then collected by centrifugation (500 g, 5 min, 4 °C), and re-suspended in 500 µl of Tris buffer supplied with 5 mM butyled hydroxytoulene (BHT) as described in manufacture manual. The cell lysates were prepared by repeated freezing in dry ice or liquid nitrogen and thawing to 37 ℃ on water bath, and then centrifuged at 15,000 g for 30 min. The intracellular levels of malondialdehyde and 4-hydroxynonenal were analyzed with a lipid peroxidation assay kit (Calbiochem).
Chapter 5 :
Inhibition of Hepatitis C Virus Replication by Antimonial CompoundsThe recent development of a sub-genomic replicon system in Huh-7 cells (19, 117) provides a powerful tool for studying virus replication and for screening anti-HCV drugs. By screening a set of marketed drugs, we have discovered that arsenic trioxide (As2O3) is
a potent HCV inhibitor (82). Compounds identified through this approach should be considered as promising candidates for drug development because the toxicity and pharmacokinetic properties of these marketed drugs are well documented. In this study, we evaluated sodium stibogluconate (SSG) (an old drug containing antimony used in leshmania treatment) for its anti-HCV potential. Antimony and arsenic both belong to the group 15 (the nitrogen family) in the periodictable. We found that SSG, along with several other antimonial compounds, including Sb2O3 and SbCl3, were able to exert
potent anti-HCV activity at concentrations that did not affect cell viability. When SSG was combined with IFN-α, these two drugs acted synergistically to suppress HCV replication and to prolong antiviral activity.
5.1 RESULTS
5.1.1 Effects of antimonial compounds on HCV replication and cellular toxicity We had previously found that arsenic trioxide (As2O3), which is clinically used to treat
acute promyelocytic leukemia (APL) (5), could suppress HCV replication in the HCV subgenomic RNA system (replicon) at doses that did not causes cellular toxicity (82). Since antimony (Sb) and arsenic belong to the same group in the periodic table, we decided to evaluate whether antimonial compounds also possess anti-HCV activities. In
this study, we found that several antimony-containing compounds, including Sb2O3,
SbCl3, and SSG, could inhibit HCV replication in Ava5-EG(∆4AB)SEAP cells (Table 1).
Ava5-EG(∆4AB)SEAP cells were treated with 5 µM of Sb2O3, 5 µM of SbCl3, or 0.5
mg/mL of SSG (equivalent to 0.105 mg/mL or 860 µM of pentavalent antimony (SbV)
when considering that the %w/w of Sb in SSG is approximately 21%) according to methods as described. Compared to untreated control cells, the remaining SEAP activity of cells treated with Sb2O3, SbCl3, or SSG was 18, 31, and 38%, respectively.
The remaining SEAP activity was 19 and 23 % when cells were treated with 100 IU/mL of IFN-α and 1 µM of As2O3, respectively. There was no observed cellular toxicity, as
revealed by MTS analysis, when cells were treated with these agents for up to 72 hrs (data not shown).
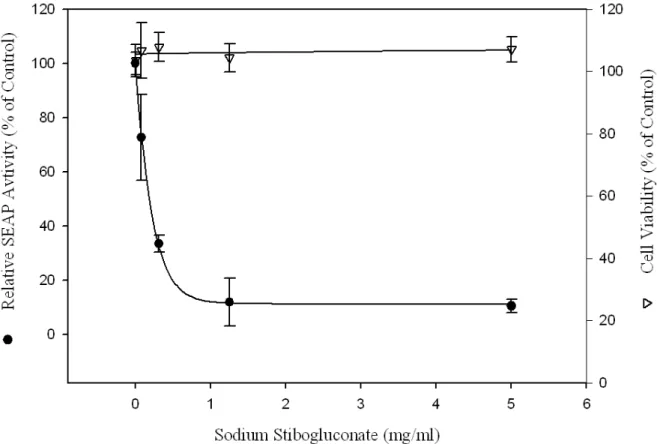
Because SSG is an existing drug, the anti-HCV effect of SSG was analyzed in more details. Ava5-EG(∆4AB)SEAP cells were treated with serially diluted SSG for 72 hrs and SEAP activities were analyzed to measure the relative copy number of the HCV
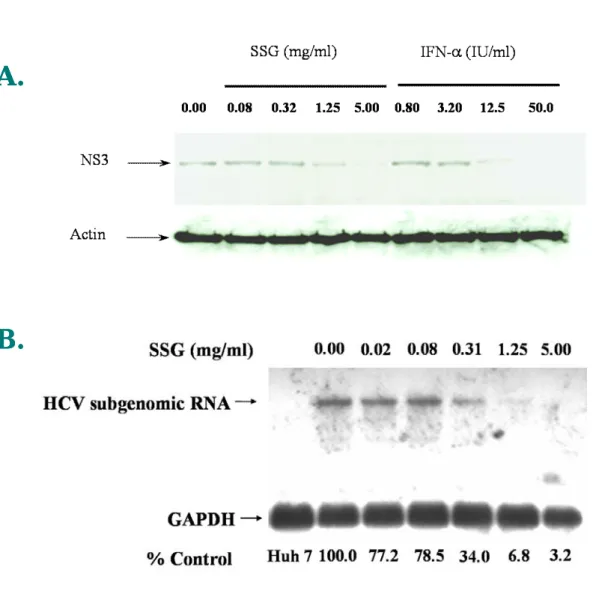
replicon (Fig. 5). SSG reduced SEAP activities in a dose-dependent manner. Activity was reduced to 20% of that in control and there was no cellular toxicity, as revealed by MTS analysis, when the cells were treated with up to 5 mg/mL (41 mM) of SSG. The
50% effective concentration (EC50) of SSG for HCV inhibition was 0.2 to 0.3 mg/mL
(equivalent to 0.042 and 0.063 mg/mL (345–517 µM) of Sb). Western-blotting analysis was also performed to confirm the results in Ava5-EG(∆4AB)SEAP reporter cell system. Ava5 cells were treated with various doses of IFN-α and SSG for 72 hours and cell lysates were collected and analyzed with anti-HCV NS3 antibody. As shown in Fig. 6A, the level of HCV NS3 protein decreased upon SSG treatment in a dose-dependent manner, which correlated with the results shown in Fig.5. In addition, results from Northern-blotting indicated that SSG at 1.15 or 5 mg/mL effectively reduced the level of HCV RNA in a dose-dependent manner (Fig. 6B). The mRNA level of GAPDH remained unchanged up SSG treatment up to 5 mg/mL and no HCV signal could be detected in the parental Huh-7 cells because of the lack of HCV subgenomic RNA in Huh-7 cells. Under the same conditions, no difference was observed when Ava5 cells were treated with various concentration of SSG for 24 h and analyzed for propidium iodide-stained DNA content by flow cytometry (results not shown).
5.1.2 Synergistic anti-HCV activity of SSG and IFN-α combination
We then assessed whether the combination of SSG and IFN-α exerts synergistic, additive, or antagonistic anti-HCV effects using the isobologram method (82, 109). In this method, synergism, additivity, or antagonism are represented by concave, linear, or convex isoeffective curves (isoboles), respectively. Inhibition of HCV replication was
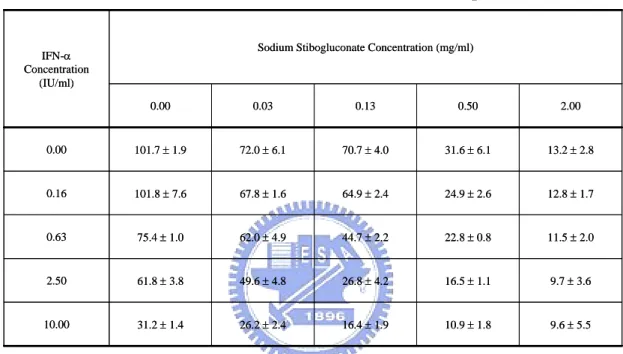
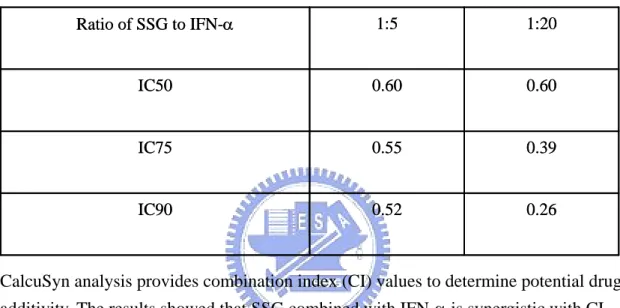
evaluated in Ava5-EG(∆4AB)SEAP reporter cells treated with various doses of SSG (0, 0.03, 0.13, 0.5, and 2 mg/mL) in combination with various doses of IFN-α (0, 0.16, 0.63, 2.5, and 10 IU/mL) (Table 2). The results presented in Table 2 were used to generate isoboles of 50% and 80% inhibition of HCV replication (Fig. 7). SSG and IFN-α exerted strong synergistic anti-HCV activities as revealed by the sharp concave isobole plots and by the CI values < 1 (range: 0.26 to 0.60), as calculated using CalcuSyn analysis (Table 3).
5.1.3 Sustained anti-HCV response after drug removal
Current IFN-based therapy suffers from an unsatisfactory Sustained Viral Response (SVR) rate. An SVR is defined as having undetectable HCV levels 6 months after the termination of antiviral treatment. In this study, we wished to evaluate if the addition of SSG might benefit IFN-based treatment by enhancing the sustained anti-HCV response using Ava5 subgenomic cells. Toward this end, Ava5 cells were firstly treated with drugs to suppress the replication of HCV subgenomic RNA. This was performed in the absence of G418 to remove the selection pressure in culture medium because the HCV subgenomic RNA in those cells contains a G418 resistant gene. Then, the drugs were removed and cells were allowed to recover in the presence of G418 and only those cells with sufficient HCV subgenomic RNA content would survive. If the HCV subgenomic RNA in drug-treated Ava5 cells “rebound” after drug removal, then cells would become resistant to G418 selection. In this experiment, Ava5 cells were treated with SSG (0.5 mg/mL), IFN-α (20 U/mL), or the combination of both for 5 days in the absence of G418. The drugs were removed and fresh culture media containing 500 µg/mL of G418 was
added. After cells were cultured for up to 30 days after drug removal, only control cells without drug treatment exhibited G418-resistant phenotype by forming colonies that can be observed under crystal violet staining (Fig. 8). All drug-treated cells failed to form G418-resistant colonies. However, at day 18 after drug removal, no drug-treated cells survived and grew under G418 selection. At day 24, no cell seemed to survive under G418 selection if cells were treated with the combination of IFN-α and SSG. A small amount of cells became stainable by crystal violet when cells were treated with IFN-α alone. Much more cells were stainable by crystal violet when cells were treated with SSG alone. At day 30, more crystal violet-stainable cells were observed when cells were treated with wither IFN-α or SSG alone. Whereas no cells could be observed after 30 days of additional culturing for those cells treated with combination of IFN-α and SSG for 5 days. These results indicated that the suppression of HCV subgenomic RNA in Ava5 cells could be sustained longer by treatment with the combination of IFN and SSG.
5.2 DISCUSSION
Currently, the only therapeutic option for treating chronic HCV infection is the IFN-based treatment. However, this therapy is often accompanied by severe side-effects, and the response rate of approximately 50% is not satisfactory. Thus, new and more effective therapeutic agents are needed to combat HCV infection. To shorten an otherwise lengthy process for drug discovery and development, we chose to screen drugs that are already in use for the treatment of other human diseases. Using an ex vivo system (liver biopsy samples), we have previously shown that SSG is effective at reducing HCV replication (206). SSG is a compound containing Sb. In this study, we examined the effect of SSG on HCV replication using the subgenomic replicon system. We showed
that several compounds containing Sb such as Sb2O3 and SbCl3 also possess potent
anti-HCV activities.
Giventhe absence of potent individual agentsagainst HCV, other thancombinations of
IFNwith ribavirin that possesspotential antiviral effects, it is important toexplore the feasibility and potential benefit of combinationtherapy with other new anti-HCV agents. The importance of combination therapy in the treatment of HCV infection has been demonstrated by the fact that combined IFN-α and ribavirin gave rise to a sustained viral response (SVR) in chronic HCV of 38 – 49% compared with 5 – 13% with IFN-α monotherapy (37, 125). In this study, we showed that there is a strong synergistic inhibitory effect of the combination SSG with IFN-α on HCV replication. Moreover, the sustained antiviral effect and thebenefit of combination treatments were examined
using Ava5 cells harboring HCV subgenomic replicon. We also showed herein that the combination of SSG and IFN-α could sustained the antiviral response in the subgenomic replicon system. Therefore, results from this study suggest that SSG should be considered for evaluation of its clinical efficacy.
Even though SSG has been used in the treatment of leishmaniasis for more than half a century, there is a limited knowledge of its pharmacokinetics properties in humans. SSG consists of Sb conjugated with glucose. After administration into humans, the peak concentration of Sb in plasma is approximately 10 µg/mL (~ 82 µM) (2, 32, 86).
Though such a peak Sb concentration seems relatively low compared to SSG EC50 for
HCV inhibition found in this study (0.2 to 0.3 mg/mL), several factors need to be taken into consideration. First, at ~ 40 µg/mL (equivalent to the achievable Sb levels in humans), SSG is capable of inhibiting HCV replication by ~ 30% and inducing a synergistic anti-HCV activity with IFN-α. These results are similar to a recent report by Tanabe et al. (184). They have shown that, using HCV replicon cells, the EC50 value of
ribavirin alone was126 µM, far above the concentrations achievable in plasma after
administration of standarddoses while ribavirin at clinically achievable concentration (~
10 µM) in combination with IFN showed strong synergistic inhibitory effects on HCV
replication. Second, if the peak Sb concentration (~ 82 µM) in plasma is in its free form but not in the glucose-conjugated form in plasma, this Sb concentration would be well above the effective concentration of free Sb (< 10 µM). As shown in Table 1, we showed that 5 µM of SbCl3 could inhibit the replication of HCV by 68% as revealed in
released from SSG after it is administred into humans. Finally, the concentration of Sb in the liver (one of the main target tissues for anti-HCV indication) after administration of SSG is not known. Several studies have indicated that high concentration of Sb in the liver could be achieved by modifying the delivery methods of SSG (7, 81). Nieto et al. have also shown that improved pharmacokinetic properties, including prolonged half-life and increased volume of distribution at steady state, could be achieved with a new formulation of SSG (134, 135). Thus, future investigations are warranted to develop novel formulations of SSG for the treatment of HCV infection. In conclusion, we believe that SSG, in its current dosage form or in a new formulation, is a promising candidate drug to be used in combination therapy for treating HCV infection.
Chapter 6:
Anti-HCV Activities of Selective Polyunsaturated Fatty AcidsThe inability to efficiently propagate HCV in cell culture had impeded the development of antiviral agents against this virus. This obstacle was partly overcome by the development of a bicistronic subgenomic HCV replicons in Huh-7 cells (18, 118). These subgenomic replicon systems have greatly facilitated the studies of HCV replication. With the aid of the HCV subgenomic RNA replication system, we evaluated the effect of the in-house compounds on HCV replication. We observed that acetaminophen, which is a cyclooxygenase inhibitor, could increase the HCV subgenomic RNA in the dose-dependent manner. Hence, we hypothesized that increasing the substrate for cyclooxygenase might decrease the HCV subgenomic RNA. To test this possibility, we treat the HCV subgenomic replicon-containing cells (Ava5) with arachidonic acid, which is the main substrate of cyclooxygenase. Indeed, we found that arachidonic acid could inhibit the HCV subgenomic RNA in a dose-dependent manner in Ava5 cells.
In this report, we showed that arachidonic acid (AA), docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA) are able to exert anti-HCV activity. Detailed dose-dependent studies showed that AA was effective at concentration that is achievable at normal physiological conditions. Importantly, when combined with IFN-α, AA was able to exert strong synergistic anti-HCV activity.
6.1 Results and Discussion
6.1.1 Effects of PUFAs on HCV RNA replication
To study the effects of PUFAs on HCV replication, the HCV sub-genomic replicon cells (Ava5) containing HCV subgenomic RNA were employed (19). Cells were treated with fatty acids, including polyunsaturated, monounsaturated and saturated fatty acids, for 24 hrs. As shown in Fig. 9 (lanes 2 - 4), several PUFAs, including arachidonic acid (AA), docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA) were able to exert potent anti-HCV activities at 100 µM. At the same concentration, α-linolenic acid, γ-linolenic, and linoleic acid only reduced HCV RNA levels slightly (Fig. 9, lanes 5 - 7). In contrast, saturated fatty acids including oleic acid, myristic acid, palmitic acid, and steric acid slightly enhanced HCV RNA levels (Fig. 9, lanes 8 - 11). The RNA levels of GADPH, a house-keeping gene, were not affected by drug treatments.
6.1.2 Arachidonic acid reduced HCV RNA level dose-dependently
To further confirm the results, we chose to analyze the effect of AA for its anti-HCV activity in more details. Ava5 cells were treated with AA at various concentrations for 24 hrs. In Fig. 10A, it is clear that AA was able to suppress HCV RNA levels in a dose-dependent manner. When cells were treated with 100 µM of AA, there was only 7.5% of HCV RNA left compared to untreated cells while treatment with 500 IU/mL of IFN-α reduced HCV RNA level to 13.4 % of the control (Fig. 10B). The EC50 (effective
concentration required to inhibit 50% of HCV RNA level) of AA is approximate 4 µM which is physiologically relevant. The plasma concentration for AA varied from 5.8 to 49.3 µM (34). As a comparison, the anti-HCV activity of IFN-α was measured in parallel and the EC50 of IFN-α was around 3.1 IU/ml (Fig. 9B).
There was no cellular toxicity for cells treated with AA at this range as revealed by MTS assay (Fig. 10C) and the IC50 (concentration required to inhibit 50% of cell viability) was
measured to be around 350 µM and 380 after 24 and 72 hrs, respectively, of drug treatment. The cellular morphology did not change after treatment with AA for 24 hrs at
100 µM (not show). Thus, AA might exert its anti-HCV effects through a specific
pathway but not because of its cellular toxicity.
6.1.3 Synergistic antiviral activity of AA combined with IFN-α
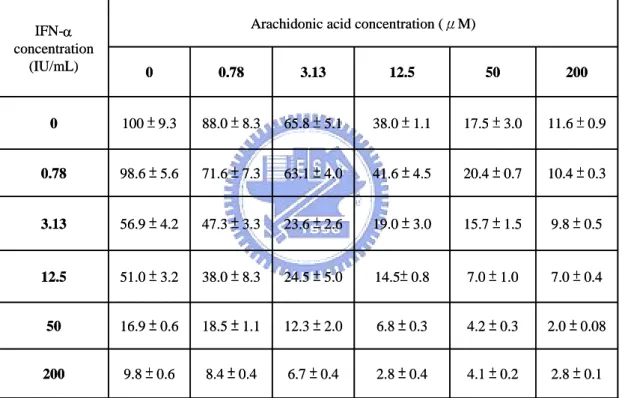
Whether AA and IFN-α combination exert synergistic, additive, or antagonistic effects was assessed by an isobologram method (31, 104). In general, representation of an isobologram to measure drug-drug interaction is shown in Fig. 11A. It was proposed that synergy, additivity, and antagonism would be represented by concave, linear, and convex isoeffective curves (isoboles), respectively. The anti-HCV effects of AA and IFN-α in combination were evaluated. Ava5 cells were treated with these two drugs in combination in a checkerboard titration manner. HCV subgenomic RNA levels in cells were then measured. Dose-response inhibition of HCV RNA replication was evaluated for varying AA concentrations (0, 0.78, 3.13, 12.5, 50, 200 µM) in the presence of various doses of
IFN-α (0, 0.78, 3.13, 12.5, 50, 200 IU/mL) (Table 1). The data in Table 4 were used to generate isoboles of 50% and 90% inhibition of HCV replication (Fig. 11B). AA and IFN-α exerted strong synergistic anti-HCV activities as revealed by the curvy concave plots of 50% and 90% isoboles.
Current IFN-based therapy for treating HCV infection is not satisfactory and development of more effective drugs has not been very fruitful over the past few years. In fact, many HCV patients often seek other complement and alternative medicine (CAM) and some may even avoid or abandon standard IFN-based therapy and seek other types of therapy (170, 177). Many forms of CAM have shown scientific evidence as cytoprotective agents. However, few were known to possess specific antiviral activity. PUFAs such as AA, DHA, and EPA were all recognized as essential nutrients in human diet. The metabolites of PUFAs also play numerous important roles in normal physiological conditions and progression of diseases (59). In this study, AA was found to be able to inhibit HCV replication at physiologically relevant concentration. Further research is needed to evaluate the therapeutic role of PUFAs in the clinical management for HCV-infected patients. It may be possible to that these fatty acids can be both as adjunctive or complementary treatment to benefit HCV patients through dietary control. In this study, we also found that antiviral activity of IFN-α can be accentuated by AA and probably also by other PUFAs. Thus, further studies are warranted if management of AA or other PUFAs through dietary control could increase the effectiveness of current IFN-based treatment as antiviral therapy.
Polyunsaturated fatty acids (PUFAs) are important for many physiologic functions (64, 98, 187). AA, the precursor of eicosanoids, can be catalyzed by at least three types of enzymes in cells, cyclooxygenases (COXs), lipoxygenases (LOXs), and P450 epoxygenase (CYPs), to generate numerous metabolites that can mediate diverse physiological and pathological responses such as blood pressure, inflammation, phagocyte activation, pain, and fever (25, 63, 151, 187). The mechanism of action of PUFAs in inhibition of HCV replication is not clear. Nevertheless, this study provides a potentially favorable observation of drug-food interactions. A human trial is mandatory to understand the clinical value of PUFAs in HCV therapy. It is also important to elucidate of the exact anti-HCV mechanism caused by the PUFAs identified this study. Such understanding may lead to the development of agents with potent activity against HCV or related viruses.