國立宜蘭大學自然資源學系碩士論文
Department of Natural Resources National Ilan University
Master Thesis
棲蘭山地區檜木林物種多樣性之研究
Study on Species Diversity in Cypress Forest of Chi-Lan Shan
指導教授:陳子英 博士
Tze-Ying Chen Ph. D.
研究生:魏瑞廷
Jui-Ting Wei
中華民國九十五年六月
摘要
棲蘭山地區清查與分化多樣性之研究,分別於四個集水區設置七條樣 帶,各樣帶沿海拔每隔100 m,有系統的設置 50 m × 20 m 樣區,合計 54 個樣區。調查上共紀錄 463 個物種,其中以台灣扁柏最為優勢,經典型對 應分析發現,樣帶之植群分布主要受海拔與地形的影響,經降趨對應分析 及雙向指標種分析並依據鑑別種群將植物社會區分為 8 型。至於物種多樣 性的趨勢大致隨海拔增加各生活型物種多樣性呈現逐漸下降的趨勢。在植 群結構上,木本植物隨海拔增加胸高斷面積也逐漸增大,物種豐富度隨海 拔增加則呈現逐漸下降的趨勢;草本植物之物種豐富度則與木本植物呈現 相反現象。在樣帶內因海拔與地形的不同,各生活型物種多樣性呈現不同 趨勢。除120-130 集水區之西向坡樣帶外,其它集水區坡向樣帶均有隨海拔 增加物種多樣性呈現下降的趨勢。樣帶內 β 多樣性的變化受海拔與地形的 影響,當海拔與地形變化愈大 β 多樣性變化愈急劇,其中以邊吉岩集水區 之北向坡樣帶變化最明顯,鴛鴦湖集水區之東向坡樣帶變化則最不明顯。
樣帶間β 多樣性顯示,物種組成主要受環境異質性之變化影響。在 γ 多樣性 方面顯示,鴛鴦湖集水區物種數明顯低於其他集水區。
經由清查多樣性與分化多樣性之比較,鴛鴦湖集水區在清查多樣性大 小與分化多樣性的變化都低於其他集水區,同時在台灣扁柏族群結構與林 分結構亦與其他集水區略有不同,此種差異也反應在第二層林分結構欠缺 台灣櫟林帶常見的優勢樟科與殼斗科物種。
關鍵字:棲蘭山檜木林、鴛鴦湖集水區、清查多樣性、分化多樣性、
族群結構、林分結構
Abstract
This study focused on investigating the inventory diversity and differentiation diversity of Cypress forest in Chilanshan. The 54 research stands (50 m × 20 m, 0.1 ha) were systematically set up with 100 m intervals along the altitudinal gradient in four watersheds. Several research stands in the same landscape slope were gathered as one transect, and therefore totally 7 transects were examined in this study. The results of this study indicated that 436 species recorded, and the Chamaecyparis obtusa var. formosana was the major and dominance species. According to the Canonical correspondence analysis, we found that the main factors controlled the vegetation distributions of 7 transects were the elevation and topographic positions. The Detrended correspondence analysis, and Two-way indicator species analysis. Based on the diagnostic species group were separated into eight vegetation types. Vegetation richness and diversity of all life forms decreased with the elevation increased. In the vegetation structure, the basal area of woody plant was increased with the elevation increased, and the species richness was decreased. However, the richness of herb was in contrast with the woody plant. The different types of α-diversity of all life forms in the same transect were showed, and which were affected by the elevation and the topographic positions.
Except for the west-facing transect in 120-130 watershed, the transect trend of the α-diversity was reduced with elevation increased. The variation of the β-diversity was affected by the elevation and topographic in the same transect, that is, significant multiple turnovers of the β-diversity were found with the elevation increased. The most significant change was found in north-facing transect of Pienjiyan watershed, but the east-facing transect of Yuan-yang Lake watershed was least. The results of the β-diversity within transects indicated that the influence on the
species composition was mainly due to the environmental heterogeneity.
The result of the γ-diversity showed that the species richness in the Yuanyang Lake watershed were lower than other watersheds.
In the Yuanyang lake watershed value of inventory diversity and variate of differentiation diversity were lower than that of other watersheds and the population structure of Chamaecyparis obtusa var.
formosana and the forest stand structure in Yuanyang lake watershed were also slightly different form other watersheds. In addition, the differences between Yuanyang lake watershed and other watersheds indicated that, the dominance species, F a g a c e a e a n d Lauraceae species, in Quercus zone in Taiwan were lack of the second layer forest stand structure.
Keyword: Chi-Lan Shan Cypress forest, Yuanyang Lake Watershed,
Inventory diversity, Differentiation diversity, Population structure, Forest structure.謝誌
當聽到自己通過碩士論文口試的一剎那,淚水就不聽使喚的在眼眶裡不 停的打轉,因為植群調查過程的辛苦是眾所皆知的,然而當研究成果完成 時,心中的喜悅一時間無法以言語道盡,因為終於完成屬於我自己的論文,
可謂是苦盡甘來。
想當初自己一個人來到宜蘭大學就讀碩士班,求學期間感謝恩師陳子 英博士在植群研究上的啟蒙,讓我從一個不懂植群研究到以植群為研究題 材之碩士論文具體呈現。論文的初稿承蒙葉慶龍博士與彭鏡毅博士費心審 稿斧正以及給予寶貴意見,在此謹致上由衷的感謝,讓我能通過碩士階段 的考驗,感謝您老師!
研究期間,特別感謝宋梧魁、程宗德、葉清旺、李智群學長、洪淑華、
楊暄慧學姊、卓子右、邱宗儀、戴炎明、何正輝、李書銘、鄭婉蒨、鄭茜 如、鄧若菁、林哲榮、陳建忠、李孝勤、林秀蓉等學弟妹,不論在野外調 查或是資料處理上都給予幫忙、協助及建議,因為有你們的相挺陪同出生 入死,才能在兩年內完成54 個 0.1 ha 的樣區。在校期間感謝蔡呈奇老師在 學業及生活上給予實質的幫忙。張瑀芳、楊涵馨、李思佳、許佳雯、蘇柏 翰、邱孟瑋、劉啟斌、陳建帆、陳雅華、謝光普、羅琨評、廖家宏及陳至 瑩等好友在日常生活上多方面給予協助與照顧。助理陳小玲及吳欣玲小姐 感謝妳們在論文排版上的協助,要不然肯定手忙腳亂。另外感謝森林保育 處棲蘭山工作站林進龍主任在調查期間給予幫忙及協助,使調查研究得以 順利完成。此外更感謝遠在屏東科技大學的女友徐惠君,因為有妳的關懷、
體諒及精神上的鼓勵,讓我更放心努力在宜蘭大學完成碩士學位。
最後將此論文獻給我的家人,在我研究期間給予我支持,讓我無後顧 之憂的得以順利完成學業。對於曾給予幫忙及協助,在此卻未能提及者,
於此一併致上最深的謝意。
研究生
目次
摘要...I Abstract...II 謝誌...IV 圖目次...VII 表目次...IX
壹、前言... 1
貳、前人研究... 3
ㄧ、棲蘭山地區之植群研究概況... 3
二、生物多樣性的基本概念... 11
三、物種多樣性指標之含意... 14
四、國內外多樣性研究的發展... 16
參、研究地區環境概述... 20
一、研究地區... 20
二、氣候... 20
三、地質與土壤... 20
肆、研究項目與研究方法... 23
一、資料收集... 23
二、植群多樣性之取樣... 23
三、環境因子觀測與評估... 23
四、原始資料計算... 26
五、植群分析方法... 28
六、多樣性分析方法... 29
伍、結果... 35
一、物種組成... 35
二、優勢度... 35
三、植群多樣性... 39
四、清查與分化多樣性... 61
五、各集水區台灣扁柏之族群結構... 104
六、各集水區之林分結構... 107
陸、討論... 113
一、棲蘭山地區植群型分類群之比對... 113
二、各集水區之植群型分類群之差異... 114
三、棲蘭山地區之植群與環境的關係... 117
四、不同尺度下各物種多樣性差異... 118
五、鴦湖集水區與台灣扁柏族群結構變化與森林永續經營... 125
六、鴛鴦湖集水區林分結構之探討... 126
七、鴛鴦湖集水區之獨特性... 126
柒、結論... 129
捌、參考文獻... 131
附錄一、棲蘭山地區各樣區之植物名錄... 137
附錄二、棲蘭山地區植群動態樣區之優勢度指數(D 值)計算一覽表... 159
圖目次
圖1 棲蘭山地區生態氣候圖... 21
圖2 研究區域範圍位置... 22
圖3 棲蘭山地區多樣性研究流程圖... 27
圖4 棲蘭山地區木本植物株數比例圖... 36
圖5 棲蘭山地區優勢樹種胸高斷面積分布比例圖... 36
圖6 棲蘭山地區優勢樹種重要值分布比例圖... 37
圖7 棲蘭山地區樣區在CCA 分布序列第一軸與第二軸之分布圖... 40
圖8 棲蘭山地區樣區在DCA 分布序列第一軸與第二軸之分布圖 ... 43
圖9 棲蘭山地區之植群結構示意圖... 70
圖10120-130 集水區西向坡-A 樣帶與東向坡-B 樣帶各生活型之物種多樣性 趨勢... 72
圖11 鴛鴦湖集水區東向坡-C 樣帶與西向坡-D 樣帶各生活型之物種多樣性 趨勢... 74
圖12160-170 集水區北向坡-E 樣帶與南向坡-F 樣帶各生活型之物種多樣性 趨勢... 76
圖13 邊吉岩集水區北向坡-G 樣帶各生活型之物種多樣性趨勢... 78
圖14120-130 集水區西向坡-A 樣帶與東向坡-B 樣帶各生活型在不同海拔之 物種轉換率... 83
圖15120-130 集水區西向坡-A 樣帶與東向坡-B 樣帶不同各生活型之物種社 會係數的轉換程度... 84
圖16 鴛鴦湖集水區東向坡-C 樣帶與西向坡-D 樣帶各生活型在不同海拔之 物種轉換率... 86
圖17 鴛鴦湖集水區東向坡-C 樣帶與西向坡-D 樣帶不同生活型之物種社會 ... 87
圖18160-170 集水區北向坡-E 樣帶與南坡-F 樣帶各生活型在不同海拔之物
... 89
圖19160-170 集水區北向坡-E 樣帶與南坡-F 樣帶不同生活型之物種社會 90 圖20 邊吉岩集水區北向坡-G 樣帶各生活型在不同海拔之物種轉換率... 92
圖21 邊吉岩集水區北向坡-G 樣帶不同生活型之物種社會性係數的轉換程 度... 93
圖22120-130 集水區樣帶木本物種分布情形... 98
圖23 鴛鴦湖集水區樣帶木本物種分布情形... 99
圖24160-170 集水區樣帶木本物種分布情形... 100
圖25 邊吉岩集水區樣帶木本物種分布情形... 100
圖26120-130 集水區樣帶草本物種分布情形... 101
圖27 鴛鴦湖集水區樣帶草本物種分布情形... 102
圖28160-170 集水區樣帶草本物種分布情形... 103
圖29 邊吉岩集水區樣帶草本物種分布情形... 104
圖30120-130 集水區中坡台灣扁柏之徑級... 105
圖31 鴛鴦湖集水區中坡台灣扁柏之徑級... 105
圖32160-170 集水區中坡台灣扁柏之徑級... 105
圖33 邊吉岩集水區中坡台灣扁柏之徑級... 106
圖34120-130 集水區林分結構示意圖... 109
圖35 鴛鴦湖集水區林分結構示意圖... 110
圖36160-170 集水區林分結構示意圖... 111
圖37 邊吉岩集水區林分結構示意圖... 112
圖38 棲蘭山地區各集水區之植群多樣性示意圖... 116
表目次
表1 棲蘭山地區植群型一覽表... 8
表2 清查多樣性與分化多樣性之量測形式與尺度... 12
表3 國內外與本研究相關之物種多樣性研究報告一覽表... 19
表4 棲蘭山地區之氣候表... 21
表5 調查樣區植物物種科屬種統計表... 35
表6 棲蘭山地區植物社會組成... 38
表7 棲蘭山地區CCA 特徵值與環境相關性... 39
表8 棲蘭山地區DCA 三軸與環境因子之相關係數... 42
表9 棲蘭山地區植群分表... 45
表10 棲蘭山地區植群型環境與物種特徵表... 46
表11 棲蘭山地區長梗紫麻-猪腳楠型之植群結構... 50
表12 棲蘭山地區大葉木犀-木荷型之植群結構... 51
表13 棲蘭山地區長葉木薑子-紅檜型之植群結構... 53
表14 棲蘭山地區小葉赤楠-台灣扁柏型之植群結構... 54
表15 棲蘭山地區木荷-台灣扁柏型之植群結構... 55
表16 棲蘭山地區昆欄樹-台灣扁柏型之植群結構... 56
表17 棲蘭山地區台灣杜鵑-台灣扁柏型之植群結構... 57
表18 棲蘭山地區台灣鐵杉-台灣扁柏型之植群結構... 60
表19 棲蘭山地區植群型及亞型之喬木多樣性... 64
表20 棲蘭山地區植群型及亞型之灌木多樣性... 65
表21 棲蘭山地區植群型及亞型之地被多樣性... 67
表22 棲蘭山地區植群型及亞型之附生植物多樣性... 68
表23 棲蘭山地區植群型及亞型之藤本多樣性... 69
表24 棲蘭山地區各集水區各生活型物種數之一覽表... 79
表25 棲蘭山地區各樣帶內之物種轉換率... 82
表26 樣帶間喬木層物種轉換率... 95
表27 樣帶間灌木層物種轉換率... 95
表28 樣帶間地被層物種轉換率... 96
表29 樣帶間附生層物種轉換率... 96
表30 樣帶間藤本層物種轉換率... 97
表31 棲蘭山地區各集水區之主要木本植物組成一覽表... 108
表32 棲蘭山地區植群分類群結果與前人研究之植群分類群對照表... 114
表33 櫟林帶與楠櫧林帶之分化多樣性比較表... 123
表34 相關研究區域物種轉換率之比較表... 124
表35 棲蘭山地區各集水區之比較... 128
壹、前言
台灣地處大陸邊緣,第四紀冰河時期,曾連接南洋地區與大陸之間,
為冰河時期物種南北遷移必經重要的橋樑。在過去五百萬年前的造山運 動,形成全島高山林立巒亙蜿蜒與複雜的山地環境,如此複雜的環境條件,
造就台灣地區物種多樣性極高的現象。依台灣中部山地植群垂直海拔梯度 變化,可將台灣山地植群多樣化區分為:高山植群帶(Alpine vegetation)、冷 杉林帶(Abies zone)、鐵杉林帶(Tsuga zone)、櫟林帶(Quercus zone)、楠櫧林 帶(Machilus-Castanopsis zone)、榕楠林帶(Ficus-Machilus zone) (蘇鴻傑,
1992)。其中以櫟林帶之環境條件較為特殊,其環境屬恆濕型氣候,濕度大,
全年無乾季,又可稱為山地盛行雲霧帶。由於環境適宜因而匯聚相當多的 物種於此生長,故雲霧帶亦有台灣森林命脈之美稱(蘇鴻傑,1992、2002;
Su, 1984b)。
依據山地植群垂直海拔梯度棲蘭山地區屬櫟林帶,植物組成大致以檜 木(Chamaecyparis spp.)主要優勢樹種,生長環境終年雲霧繚繞,亦可稱檜木 林或雲霧林。櫟林帶於台灣山地森林植群中扮演著明顯的分界線(Su, 1984b ; 1985),林帶上以針葉林為主,林帶下以闊葉林為主;依據前人研究報告得 知,雲霧林由於氣候恆定性高,環境適合各物種生長,而造就生物多樣性 極高之現象(邵彬,1999;Vazquez and Givnish, 1998 ; Itow, 1991)。以台灣來 看,棲蘭山地區雖然不是生物多樣性最高的區域,但其所包含之特有物種 的比率是最高的。如近年來所發現的鴛鴦湖細辛(Asarum crassusepalum)、
棲蘭山杜鵑(Rhododendron chilanshanense)等即可印證(王震哲,2000),此外 棲蘭山地區在林分組成結構也特別多樣化(陳子英等,2002)。故基於生物多 樣 性 保 育 的 觀 念 亟 應 積 極 調 查 與 探 討 , 並 利 用 清 查 多 樣 性(Inventory diversity)與分化多樣性(Differentiation diversity)之各種指標特性,顯示植群 結構與物種組成,並依據各種不同特性之指標間接顯示植群與物種本身所
具有之功能的特徵,奠定此區域在生物多樣性保育上的重要性(馬克平,
1994;蘇鴻傑,2002;Su, 1994)。
近年來生態學者常應用的多樣性指標大致區分清查與分化多樣性二 類。清查多樣性係指α 多樣性和 γ 多樣性;分化多樣性則係指 β 多樣性(馬 克平,1994;邱祈榮等,1999;蘇鴻傑,2002;蘇鴻傑,2004;Kent and Coker, 1992 ; Su, 1994;Magurran, 2004)。α 多樣性係指在某一均質的生育地或植物 社會中物種的總數;γ 多樣性即為某一區域中所有生育地所出現之物種總 數;分化多樣性;β 多樣性為沿著某一環境梯度上的生育地至另一個生育地 上物種轉換。
棲蘭山地區不論在清查或分化多樣性上均具有其研究價值。近年來有 對棲蘭山地區植群種類組成分別進行研究,但對於棲蘭山地區清查與分化 多樣性,一直未有相關研究的報導(王震哲,2000;陳子英,2002;朱恩良,
2004)。本文主要目的有下列三項:(1) 清查棲蘭山地區植群與物種多樣性 及探討物種和社會間的分化多樣性,並利用清查與分化多樣性之結果與前 人研究作一比較;(2) 探討中台灣地區在中低海拔林帶物種多樣的變化(蘇 鴻傑,2002);(3) 同時以棲蘭山地區各集水區為單位,探討鴛鴦湖集水區 與其他集水區,在物種多樣性、台灣扁柏族群結構及林分結構之差異(陳子 英等,2002),以提供未來林業相關單位作為育林工作擬定實施前之參考方 針。
貳、前人研究
ㄧ、棲蘭山地區之植群研究概況
棲蘭山地區海拔約1100-2900 m,120 及 130 林道、鴛鴦湖集水區、160 及 170 林道與邊吉岩集水區均位於大漢溪之上游處。本區域由於氣候極具 特異,且境內之動植物相極為豐富,故長久以來頗受植物學家與其他相關 研究人員之重視。依據地理氣候區的劃分,棲蘭山地區範圍介於東北氣候 區與西北區兩處氣候交會轉換帶(蘇鴻傑,1985),且氣候變化明顯,造成分 布於兩區域之植物種類有明顯的差異。一般生態交會帶上通常具有明顯且 較高的生物多樣性,故棲蘭山地區在台灣生物多樣性有其獨特的地理位置 (王震哲,2000)。
綜合棲蘭山地區植物資源調查研究(王震哲,2000);棲蘭野生動物重要 棲息環境棲地植群調查(陳子英,2002);棲蘭山 170 林道檜木之植群調查(陳 子英等,2002);蘭陽溪的植群分類系統之研究(陳子英,2004);棲蘭野生 動物重要棲息環境植群生態之研究(朱恩良,2004);台灣北部溫帶針葉林之 植物組成及檜木的更新(廖啟政,2005),這些研究報告將棲蘭山植群區分成 以下各個林帶(表 1):
(一)鐵杉林帶
此林帶分布於海拔1500 m以上之山頂稜線,氣候乾燥,土壤腐植 質 高 之 地 區 。 包 含 的 植 群 型 為 台 灣 鐵 杉 林 型(Tsuga chinensis var.
formosana type),此林帶以台灣鐵杉(Tsuga chinensis var. formosana)為優 勢種並有玉山箭竹(Yushania niitakayamensis)伴隨著,而其他草本則比 較少,種類組成較為單純。鐵杉林帶於高海拔處較易形成純林,於海 拔較低處之山頂稜線則常與檜木林相混形成山地上層針葉林型(Upper montane coniferous forest type)。
此林型主要分布於稜線ㄧ帶,海拔1900-2721 m,包含的植群型為
台灣鐵杉-玉山杜鵑型(Tsuga chinensis var. formosana-Rhododendron pseudochrysanthum type) ; 台 灣 二 葉 松 - 台 灣 鐵 杉 型 (Pinus taiwanensis-Tsuga chinensis var. formosana type)。此林帶以巒大花楸 (Sorbus randaiensis)、台灣扁柏(Chamaecyparis obtusa var. formosana)、
玉 山 杜 鵑 (Rhododendron pseudochrysanthum) 、 厚 葉 柃 木 (Eurya glaberrima) 、 假 繡 球 (Viburnum sympodiale) 、 台 灣 八 角 金 盤 (Fatsia polycarpa) 、 台 灣 鐵 杉 、 台 灣 扁 柏 、 高 山 新 木 薑 子 (Neolitsea acuminatissima)及毽子櫟(Cyclobalanopsis sessilifolia)為優勢種;台灣鐵 杉、台灣二葉松(Pinus taiwanensis)則為特徵種。
(二)櫟林帶
此林帶相當於台灣盛行之雲霧帶,出現海拔約700-2500 m;本林 帶依分布的海拔高度不同,可分為:分布於海拔1000-2700 m之櫟林帶 上層(Quercus Upper Zone),700-1600 m之櫟林帶下層(Quercus Lower Zone)。
1.櫟林帶上層
櫟林帶上層的森林帶為:山地針闊葉混淆林型(Montane mixed coniferous forest type)、台灣扁柏型(Chamaecyparis obtusa var. formosana type)、紅檜型(Chamaecyparis formosensis type)。
(1)山地針闊葉混淆林型
主 要 為 中 坡 植 物 社 會 西 施 花 - 台 灣 二 葉 松 型 (Rhododendron leptosanthum-Pinus taiwanensis type);中下坡植 物社會台灣二葉松-西施花型(Pinus taiwanensis-Rhododendron leptosanthum type) 及 木 荷 - 西 施 花 型 (Schima superba var.
superba-Rhododendron leptosanthum type)。此林帶以台灣鐵 杉、台灣扁柏、西施花(Rhododendron leptosanthum)、台灣二
葉松、木荷(Schima superba var. superba)、杜英(Elaeocarpus sylvestris)、蘭邯千金榆(Carpinus rankanensis)、台灣鐵杉、昆 欄樹(Trochodendron aralioides)、小花鼠刺(Itea parviflora)、台 灣 紅 榨 槭 (Acer morrisonense) 、 錐 果 櫟 (Cyclobalanopsis longinux)、台灣樹參(Dendropanax dentiger)、薯豆(Elaeocarpus japonicus)、香桂(Cinnamomum subavenium)、烏心石(Michelia compressa var formosana)及薯豆為優勢種;並以台灣二葉松、
木荷及烏皮茶為特徵種。
(2)台灣扁柏型
主要分布於中坡至稜線ㄧ帶,海拔1333-2435 m,植群型 為 台 灣 扁 柏- 台 灣 鐵 杉 型 ; 台 灣 杜 鵑 - 台 灣 扁 柏 型 (Rhododendron formosanum-Chamaecyparis obtusa var.
formosana type) ; 台 灣 扁 柏 - 巒 大 杉 - 台 灣 杉 林 型 (Chamaecyparis obtusa var. formosana-Cunninghamia konishii-Taiwania cryptomerioides type) ; 木 荷 - 台 灣 扁 柏 型 (Schima superba var. superba - Chamaecyparis obtusa var.
formosana type)。以台灣扁柏、台灣鐵杉、赤柯(Cyclobalanopsis morii)、高山新木薑子、台灣樹參、台灣杜鵑(Rhododendron formosanum) 、 毽 子 櫟 、 霧 社 木 薑 子 (Litsea elongata var.
mushaensis)、白花八角(Illicium anisatum)、假長葉楠(Machilus japonica)及長葉木薑子(Litsea acuminata)等為優勢種;木荷、
台灣扁柏、高山新木薑子、霧社木薑子、烏皮茶及烏心石等 為特徵種。
(3) 紅檜型
植物社會海拔963-2249 m,主要分布於下坡、溪谷等生 育地,但偶亦有分布於高海拔上坡之乾溝生育地上。主要植
群型為紅檜-假長葉楠型(Chamaecyparis formosensis-Machilus japonica type) ; 紅 檜 - 長 葉 木 薑 子 型 (Chamaecyparis formosensis-Litsea acuminatetype);紅檜群叢。以紅檜、錐果 櫟、早田氏冬青(Ilex hayataiana)、長葉木薑子、假長葉楠、
銳葉柃木(Eurya acuminata)、薯豆、昆欄樹、台灣八角金盤、
墨點櫻桃(Prunus phaeosticta)、小花鼠刺及西施花等為優勢 種;紅檜特徵種。
2. 櫟林帶下層
櫟 林 帶 下 層 的 森 林 依 其 林 型 可 分 為 ; 山 地 常 綠 闊 葉 林 型 (Montane evergreen broad-leaved forest type)、山地落葉闊葉林型 (Montane deciduous broad-leaved forest type)。
(1)山地常綠闊葉林型
主要的植物社會分布於海拔700-1500 m,其植群型包含 長 尾 尖 葉 櫧 - 香 楠 型 (Castanopsis cuspidata var.
carlesii-Machilus zuihoensis type);錐果櫟型(Cyclobalanopsis longinux type)。此林型的植物組成為長尾尖葉櫧(Castanopsis cuspidata var. carlesii)、香楠(Machilus zuihoensis)、猪腳楠 (Machilus thunbergii)、山紅柿(Diospyros morrisiana)、杜英、
薯豆、小花鼠刺、猴歡喜(Sloanea formosana)、錐果櫟、黃杞 (Engelhardia roxburghiana)及西施花。
(2)山地落葉闊葉林型
主要分布於下坡近溪谷地區或經崩塌之生育地,海拔 1339-1600 m , 主 要 植 群 型 合 歡 - 長 梗 紫 麻 型 (Albizia julibrissink-Oreocnide pedunculata type);台灣赤楊-芒型(Alnus formosana-Miscanthus sinensis type)。植物組成以長梗紫麻
(Oreocnide pedunculata) 、 台 灣 赤 楊 (Alnus formosana) 及 芒 (Miscanthus sinensis)為優勢種;合歡(Albizia julibrissin)及台灣 蘆竹(Arundo formosana)為特徵種。
表1 棲蘭山地區植群型一覽表
植群帶 生育地環境 研究區域植物社會 特徵種或優勢種 作者
鐵杉林帶
海拔1500 m 以上之山頂稜線 棲蘭山檜木林區
台灣鐵杉林型 台灣鐵杉* 王震哲,2000
海拔2355-2721 m,稜線,主稜,邊吉 岩山至喀拉業山ㄧ帶及發徒山至南湖 大山中途
蘭陽溪上游 台灣鐵杉-玉山杜鵑型
巒大花楸、台灣扁柏、假繡球、
玉山杜鵑*、厚葉柃木、台灣八 角金盤、台灣鐵杉*
陳子英,2002
山地上層針葉林型
海拔1900-2111 m,稜線 棲蘭山170 林道 台灣二葉松-台灣鐵杉型
台灣二葉松*、台灣鐵杉、台灣
扁柏、高山新木薑子、毽子櫟 陳子英等,2002 海拔1680 m,中坡生育地環境
乾燥,司馬庫斯ㄧ帶
棲蘭山地區
西施花-台灣二葉松型 西施花*、台灣二葉松、木荷 朱恩良,2004
海拔1455 m,下坡 蘭陽溪上游 台灣二葉松-西施花型
杜英、台灣二葉松*、蘭邯千金 榆、台灣鐵杉、昆欄樹、小花鼠 刺、台灣紅榨槭、西施花*
陳子英,2002
海拔 1325-1975 m,下坡,中坡小支 稜,南山村以上
蘭陽溪上游 木荷-西施花型
木荷*、錐果櫟、香桂、烏心石、
薯豆 陳子英,2002
地針闊葉混淆林型
海拔1405-1655 m,支稜,南山村至邊 吉岩山右側與思源啞口附近之支稜
蘭陽溪上游 台灣杜鵑-薯豆型
台灣杜鵑*、烏皮茶*、錐果櫟、
薯豆、台灣樹參、台灣扁柏 陳子英,2002
8
續表 1 棲蘭山地區植群型一覽表
植群帶 生育地環境 研究區域植物社會 特徵種或優勢種 作者
海拔1500-2700 m 棲蘭山地區 台灣扁柏-台灣鐵杉型
台灣扁柏*、台灣鐵杉、赤柯、高山新木薑
子 朱恩良,2004
海拔1017-2150 m 棲蘭山地區 台灣扁柏-台灣杜鵑林型
台灣扁柏*、台灣鐵杉、白花八角、毽子櫟、
西施花、台灣杜鵑* 廖啟政,2005
海拔1800-1900 m,上坡 棲蘭山170 林道 台灣杜鵑-台灣扁柏型
台灣杜鵑*、台灣扁柏、台灣鐵杉、高山新
木薑子、毽子櫟、白花八角 陳子英等,2002 海拔1000 m 以上,160-170
鄰道沿線之坡面
棲蘭山檜木林區 台灣扁柏-巒大杉-台灣杉
林型
台灣扁柏*、巒大杉、台灣杉、木荷、毽子
櫟、霧社木薑子 王震哲,2000
台灣扁柏型
海拔1700-1800 m,中坡 棲蘭山170 林道 木荷-台灣扁柏型
木荷*、台灣扁柏、烏心石*、霧社木薑子*、
烏皮茶*、白花八角 陳子英等,2002
海拔1700 m,下坡或溪谷 棲蘭山170 林道 紅檜-假長葉楠型
紅檜*、假長葉楠、錐果櫟、長葉木薑子、
早田氏冬青、銳葉柃木
陳子英等,2002
海拔1450-1760 m,棲蘭山歷代 神木園區、四季南山一帶之中 坡乾溝
蘭陽溪上游 紅檜群叢
紅檜*、薯豆、假長葉楠、西施花、台灣八 角金盤、長葉木薑子、昆欄樹、墨點櫻桃、
小花鼠刺
陳子英,2004 櫟林帶上層 山地針闊葉混淆林型 紅檜型
海拔963-1630 m 棲蘭山地區 紅檜-長葉木薑子林型
紅檜*、台灣扁柏、長葉木薑子、猪腳楠、
假長葉楠、錐果櫟
廖啟政,2005
9
續表 1 棲蘭山地區植群型一覽表
植群帶 生育地環境 研究區域植物社會 特徵種或優勢種 作者 海拔700-1500 m。 棲蘭山檜木林區
長尾尖葉櫧-香楠林型
長尾尖葉櫧*、香楠、猪腳楠、山紅柿、杜
英、薯豆、小花鼠刺、猴歡喜 王震哲,2000 海拔1325-1725 m,下坡、
中坡及上坡,為過南山村後 蘭陽溪右側
蘭陽溪上游 木荷-大葉石櫟群叢
假長葉楠、錐果櫟、薯豆、墨點櫻桃,香
桂、木荷*、烏心石,西施花、長葉木薑子 陳子英,2004
山地常綠闊葉林型
海拔1065 m,近溪谷兩側,
生育地由崩落之土石堆積 形成,北橫公路沿線
棲蘭山地區
錐果櫟型 錐果櫟*、台灣黃杞、西施花 朱恩良,2004 海拔1339-1405 m,下坡近
溪谷,南山村
蘭陽溪上游
合歡-長梗紫麻型 合歡*、台灣蘆竹*、長梗紫麻 陳子英,2002 櫟林帶下層 山地落葉闊葉林型
海拔1500-1600 m,東北向 山坡母岩裸露、土壤化育不 佳之崩塌地區
棲蘭山檜木林區
台灣赤楊-芒型群型 台灣赤楊*,芒 王震哲,2000
註:*為特徵種
10
二、生物多樣性的基本概念
生物多樣性係指各種生命形式的資源,它包含數以萬計的植物、動物、
微生物及各物種所擁有的基因,此外,更包含了物種與環境間相互影響所 形成的生態系統(陳靈芝,1994),由於生物多樣性的研究範圍廣泛且複雜,
對其的理解也不盡一致。過去生物多樣性研究學者公認生物多樣性涵蓋三 層級即為:遺傳多樣性、物種多樣性、生態系多樣性(Magurran, 2004)。近 年來由於大眾逐漸重視景觀多樣性,因此將其認定為第 4 層級,並與前三 個層級之組成、結構、功能與動態等方面作綜合性的研究討論。
在生態系多樣性的研究層級中,物種的多樣性是最早被研究且也是最 廣泛研究的一個層級,不論是概念或量測方法上相對都比較成熟。物種多 樣性的研究內容多以植群的組成與結構為主要對象(馬克平,1994;蘇鴻傑,
2002;Su, 1994)。探討物種的多樣性時,通常會依據不同對象採用適合之形 式及尺度加以定義,且由於研究者理念並非相一致,因此在物種的多樣性 定義上極為分歧,並經常導致混淆(蘇鴻傑,2004)。因此有生態學者將物種 的多樣性區分為清查與分化兩種形式(表 2)。所謂清查多樣性即為清查一生 態單元內之物種豐富度及植物社會數目,分化多樣性則量測物種在生態單 位內形成局部植群或小單元之方式,此目的不在於計算物種數,而在於探 討生態單元或植群間之變化差異程度。由於本研究限於區域與面積,因此 無法針對1.區塊、2.格局樣性、3.同生群多樣性、4.聚落群多樣性、5. ε 多樣 性與6.δ 多樣性進行討論,於此僅針對 α、β 及 γ 與植群型多樣性進行論述(蘇 鴻傑,2004)。
表2 清查多樣性與分化多樣性之量測形式與尺度
形式 清查多樣性 分化多樣性
層級 植群多樣性
Vegetation diversity
物種多樣性
Species diversity
物種與社會間
Between sp. and Community
單位符號 α、γ 多樣性 β 多樣性
區塊
內部α 多樣性、點多樣 性:均質社會內為棲地 樣本內之種數
林分
同生群多樣性:社會 樣本內微環境分化之 同生群數目
α 多樣性、棲地內多樣 性:均質社會樣本之物 種數
內部 β 多樣性、格局多 樣性:社會內部分化微 環境間之物種組成變化 程度
地景、區域
植群型多樣性:區域 地景內異質植物社會 基本單位之數目,如 群叢、群系、優勢型 或其他單位之數目
γ 多樣性、地景或區域 多樣性:區域地景內異 質社會樣本之總計物 種數
β 多樣性、棲地間多樣 性:地景或區域內沿環 境梯度或不同社會樣本 間之物種組成變化程度 量
測 尺 度
地理區域 聚落群多樣性:大地 理區域內異質植物社 會層級分類系統之單 位數目
ε 多樣性、地理區域多 樣性:廣大地理區域內 涵蓋不同地景之總計 物種數
δ 多樣性、地理分化多樣 性:大氣候梯度或地理 區域間之物種組成變化 程度
(參考自蘇鴻傑,2004)
註:本研究針對粗體部分進行論述
(一)清查多樣性
1.植群多樣性
植群多樣性一詞,在過去較少被台灣生態研究學者使用,而 是以物種多樣性的使用較為廣泛。事實上,在過去生態研究進行 之植群分類或植群分型,所區分之生育地型,植群型或聚落群 (Syntaxa)也都是植群多樣性的一部份,主要都以清查地域性植物 社會單元及數目為目的。然而在植群多樣性與物種多樣性之差別 僅是將物種層級提升至植物社會層級,兩者間的差異於植群多樣 性之單位是採用植物社會,而物種多樣性的單位則採用物種。因 為量測尺度上的差異,植群多樣性的量測對象依尺度單位不同可 分為:林分尺度,即表現於同生群多樣性上;地景區域尺度,即
表現於植群型多樣性上(蘇鴻傑,2004)。
2.物種多樣性
目前對於物種多樣性的量測,以清查多樣性之α和γ多樣性及 分化多樣性之β多樣性三個層級為主。清查多樣性之α多樣性即係 林分尺度下探討社會的物種多樣性、γ多樣性則以地景區域尺度下 探討區域內的物種多樣性,分化多樣性之β多樣性在林分尺度下即 探討社會內部微環境間物種組成差異的程度,在地景區域尺度下 則探討棲地間或區域間物種沿環境梯度物種組成轉換程度(馬克 平,1994;邱祈榮等,1999;蘇鴻傑,2002;蘇鴻傑,2004;Kent and Coker, 1992;Su, 1994;McCune, 2002;Magurran, 2004),茲 分述如下:
(1) α 多樣性
α多樣性的概念最早是由Williams於1943首先提出來的,
隨後,Fisher等在研究Rothamasted鱗翅目昆蟲物種-豐富度的 關係首次使用「多樣性指數」這個概念。Simpson(1949)提出 Simpson多樣性指數,此指數必須考慮樣區大小、物種的豐富 程 度 以 及 物 種 分 配 的 均 勻 度 。 同 年Shannon與Wiener提出 Shannon-Wiener多樣性指數,該指數提出後一直受到植群生態 學界的青睞並廣為應用,且被認為是量測多樣性最有效的指 數之ㄧ。上述各種多樣性量測概念均只針對一個生育地內的 多樣性進行探討,而並未探討物種與環境間的關係即稱為α多 樣性(劉棠瑞、蘇鴻傑,1983;馬克平,1994;Magurran,2004)。
(2) γ 多樣性
即探討景觀或一個以上均質植物社會的多樣性,而此多 樣性會隨著同一生育地型,不同地區的地理變換,遭遇額外
物種或喪失某些物種的速率即為某一區域中所有生育地所出 現之物種總數。因此γ多樣性隨著不同地景單元或地理氣候區 的擴大而產生不同的物種轉換率或生育地之適應 (趙榮台,
1996)。γ多樣性亦稱為景觀多樣性係指一定區域內α多樣性與β 多樣性的總和,故又可稱為總體多樣性(中國科學院多樣性委 員會,1994)。
(二)分化多樣性
1.物種與社會間的差異 (1) β 多樣性
β多樣性主要是探討植群間結構組成的相似性以及差異 性,並且可反應植群與其生態隙區(Niche)的異質性或多樣 性,而每個物種在不同的環境梯度上具有一定的分布範圍 (Austin, 1985),且各物種的生存空間會隨環境的改變而有所變 化,由此得之,環境因子對於物種的分布以及生存有著極重 要相關性,植群間之變異現象又稱為物種轉換率(Species turnover rate),主要是受海拔、土壤、水分、地形、坡度及日 照量等環境梯度所影響(蘇鴻傑,1987a),在這些環境因子中 又以海拔及地形因子影響最大。
三、物種多樣性指標之含意
在物種多樣性的研究上常應用各種多樣性指數作為評估植群組成與結 構變化的依據,因此在使用多樣性指數時必須先熟悉與暸解各種指數的含 意。
(一) α 多樣性
α多樣性雖然僅探討植群內物種之總數,但在量測取樣時必須注意 面積的大小及數目對於物種數的影響。除特別注意物種數外,物種豐
富度之分配,宜採用各種多樣性指數加以計算,以利於比較。此外,
在選擇α多樣性指數時必須符合下列各個標準,避免量測時指數使用不 當的錯誤出現,以減少研究上誤差增加研究結果的可信度(馬克平,
1994;Magurran, 2004)。
1.判別差異的能力:若某一生育地表面上差異不大時,可採用物種 豐富度指數、Shannon-Wiener 多樣性指數來作為判斷依據,因為 此兩種指數的判斷能力較強。
2.對於樣區大小的敏感度:在眾多多樣性指數種以物種豐富度指數 最容易受到樣區大小的影響。
3.強調哪一種多樣性成分(稀有種或優勢種):主要在說明 Simpson 多樣性指數對優勢種較為敏感,Shannon-Wiener 多樣性指數則對 稀有種較敏感。
4.使用較為廣泛:目前在植群生態上多樣性指數使用較廣泛的分別 為物種豐富度、Shannon-Wiener 多樣性指數、Simpson 多樣性指 數及均勻度指數等。
(二) β 多樣性
植群多樣性研究中α多樣性指所能傳達的生態意義,僅侷限於植群 內物種的組成的變化,而無法探討不同植群間物種組成上的差異,因 此必需藉由β多樣性的研究揭示各植群間物種轉換與相似的程度,並藉 此瞭解各區域內微環境的變化(馬克平,1994、Magurran, 2004)。
1.β 多樣性可反映棲地的變化程度或指示棲地被物種分隔的程度:
棲地間結構組成上的相似,可以反應植群與其生態棲位的異質性 或多樣性(Austin, 1985)。在不同的環境梯度下各物種的生存空間 會隨環境的改變而有所變化,這些變化均可藉由β 多樣性的量測 來瞭解。
2.β 多樣性的高低可以用來比較不同地點的棲地多樣性:利用相似 性指數來量測棲地間的差異程度,若各棲地間相似性高則代表該 區域棲地多樣性低,若相似性低則表示該區域的棲地多樣性高。
3.β 多樣性與 α 多樣性一起討論可構成景觀多樣性或即可瞭解ㄧ定 地段內的生物多樣性:β 多樣性與 α 多樣性兩者相互綜合討論即 可瞭解該區域景觀多樣性,景觀多樣性愈複雜即代表某ㄧ區域內 棲地異質性高,相對的其生物多樣性也隨之較高。
(三)γ 多樣性
γ多樣性與α多樣性探討的意涵相同,其之間的差異為尺度上的不 同。α多樣性的尺度僅侷限於某ㄧ個區域上某ㄧ個植物社會內物種豐富 度;γ多樣性的尺度則明顯大於α多樣性的尺度,其尺度為探討某ㄧ個 區域內所有植物社會的物種豐富度,即為組合某ㄧ區域內所有樣區做 探討(蘇鴻傑,2002;Su, 1994)。由於二種多樣性僅為尺度上的不同,
因此在量測上所需注意的事項也大致相同。
有關多樣性指數的量測與定義至今仍有許多研究學者不斷地提出 新的見解,雖然目前多樣性的定義與量測方法很多,但在進行研究時 必須掌握其概念與技術層面上的問題,這樣所量測獲得的多樣性指數 才具有生態上的意義。
四、國內外多樣性研究的發展
葉清旺(2004)里龍山植群多樣性之研究;于德永等(2003)長白山北坡植 物物種群落相異性及其海拔梯度變化;Bhattarai 和 Vetaas(2003)沿尼泊爾 東部喜馬拉雅山區之亞熱帶海拔梯度上,不同生活型之種類豐富度變異;
Patricia et al.(2002)墨西哥熱帶乾燥林 β 多樣性類型;王國宏(2002)祁連山北 坡中段植物群落多樣性的垂直分布格局;Kessler(2000)玻利維亞中部安地斯 山沿海拔梯度物種豐富度與特有性之分布;邵彬(1999)長白山北坡亞高山雲
冷杉林的物種多樣性;Vazquez 和 Givnish(1998)墨西哥 Cerro Grand 山區植 群多樣性調查之研究;廖啟政(1995)南仁山區亞熱帶雨林海拔梯度與植被組 成、結構、多樣性及分布類型的關係;Su(1994)台灣森林植物的物種多樣性 研究;Itow(1991)沿著梯度變化-探討常綠闊葉林之物種轉換及多樣性類型 (表 3)。回顧上述各篇研究報告可得知清查與分化多樣性在不同環境下有著 各種不同的趨勢。
(一)清查多樣性 1. α 多樣性
物種多樣性(α多樣性)隨著海拔的變化是沒有固定的趨勢。相 關文獻可歸納下述3點:(1).隨著海拔的上升物種多樣性及豐富度 隨之下降(物種數量多但物種種類少)(Su, 1994;Vazquez and Givnish, 1998;Kessler, 2000);(2).隨著海拔的上升物種多樣性呈 現單峰(Unimodal)的現象(環境兩端物種種類少,中間物種種類 多)(王國宏,2002;葉清旺,2004; Kessler, 2000;Bhattarai and Vetaas, 2003);(3).隨著隨著海拔的上升物種多樣性呈現上升的趨 勢(物種種類多)(Itow, 1991)。一般來說,物種多樣性的趨勢以單 峰變化現象最常見,主要是受雲霧帶的影響,因為雲霧帶終年維 持穩定的氣候以及濕度適合各種物種的生長(邵彬,1999;Itow, 1991; Vazquez & Givnish, 1998)。此外隨海拔上升物種的增加或減 少也受調查尺度及均質性等問題影響,在取樣時必須特別注意,
以求得資料之正確性。
2. γ 多樣性
γ多樣性的高低主要受β多樣性的影響,當區域受β多樣性影響 越大即代表該區域內γ多樣性越高;反之,受β多樣性影響越小即 代表該區域內γ多樣性越低。以台灣全島來看γ多樣性最高的區域
位於東南地區的楠櫧林帶,主要因為該地區屬南北植物區系之過 度渡帶,以及該區域受環境急劇變化所影響,因此造成區域內有 許多微環境的發生進而促使該地區物種多樣性特別高而反映於γ 多樣性上(Su, 1994)。
(二)分化多樣性 1. β 多樣性
物種對於環境的適應有著不同的耐性範圍,隨著耐性範圍的 不同各物種在空間分布上有著多樣化的類型,故由此得知,β多樣 性的變化會依據物種的耐性範圍而有所變化,因此並不會有一定 變化的趨勢,以海拔地型位置變化來看可區分為二類(1).隨海拔地 形變化急劇物種轉換次數增加及轉換數率較快(葉清旺,2004;廖 啟政,1995);(2).當海拔與地形位置變化較小時,物種轉換次數 降低及轉換數率變慢(葉清旺,2004;Itow, 1991)。
表3 國內外與本研究相關之物種多樣性研究報告一覽表
作者 年代 α-多樣性 β-多樣性 γ-多樣性 國家/地區
葉清旺 2004 ○ ○ 里龍山
于永德等 2003 ○ ○ 中國長白山
王國宏 2002 ○ ○ 中國祁連山
邵彬 1999 ○ ○ 中國長白山
廖啟政 1995 ○ ○ 南仁山
Su 1994 ○ ○ ○ 台灣
Bhattarai 和
Vetaas. 2003 ○ 尼泊爾
Patricia et al. 2002 ○ ○ 墨西哥
Kessler 2000 ○ 波利維亞
Vazquez 和
Givnish 1998 ○ ○ 墨西哥
Itow 1991 ○ ○ 日本
註:○符號代表進行過的研究
參、研究地區環境概述
一、研究地區
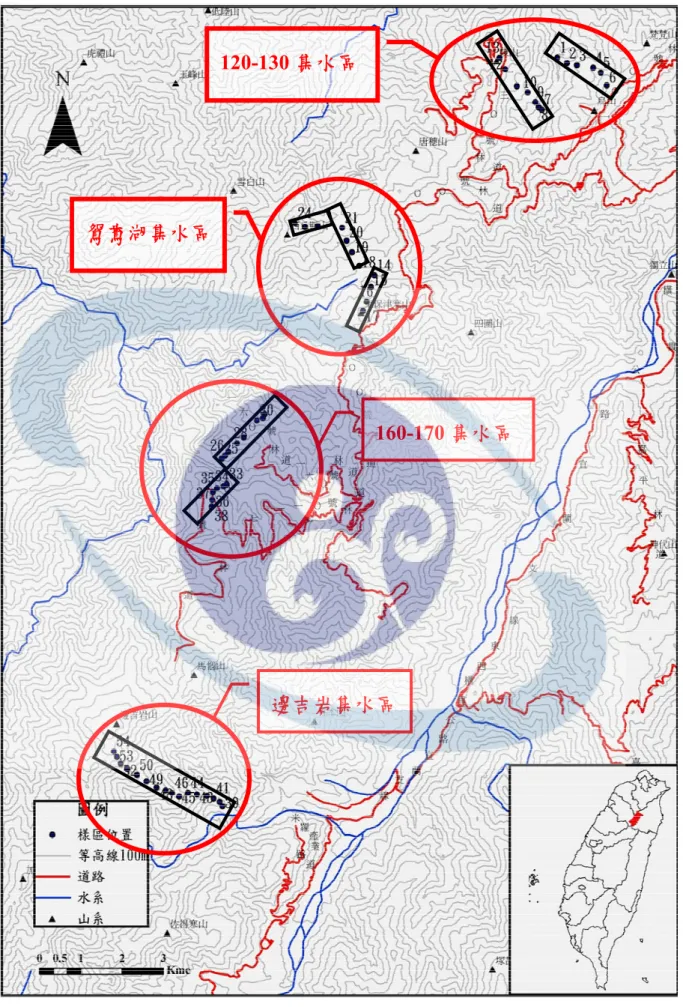
棲蘭山地區位於宜蘭縣、新竹縣、桃園縣等三縣市之交界處,地理位 置位於中央山脈西翼地質區內的雪山山脈帶北段,以喀拉業山主脊稜線向 東北延伸,經馬惱山、眉有岩山、唐穗山、棲蘭山至拳頭母山之雪山山脈 主脊稜線兩側大片山區皆屬其範圍,平均高度為海拔約2,000 m,本區域分 屬蘭陽溪水系及大漢溪水系源頭(王震哲,2000)。在本研究區域內共有四個 集水區,各集水區之名稱以兩側林道及區域名稱作為集水區之簡稱。各集 水區分別為120 林道與 130 林道之間的 120-130 集水區,鴛鴦湖地區之鴛鴦 湖集水區,160 林道與 170 林道之間的 160-170 集水區,邊吉岩地區之邊吉 岩集水區。各集水區皆有多條林道可到達交通頗為方便,而各林道均需經 由北部橫貫公路與外連接;此外,經由新竹縣尖石鄉的新興、司馬庫斯與 鎮西堡等地區之步道亦可達研本究區域內(呂金誠,2002;2003)(圖 2)。
二、氣候
本研究以棲蘭山地區氣象站(2004)所測得之氣象資料為依據(表 4),年 平均降雨量約3070 mm,且多集中於 7 月至 10 月,其中以 7 月單月降雨量 1252 mm 為全年之最;年平均溫度為 11.9 ℃,最高溫為 8 月的 16.8 ℃,最 低溫為1 月的 5.1 ℃,於冬季偶有降雪。依台灣地理氣候區之劃分本研究區 域屬於東北以及西北內陸區,以山地植群帶的區分本研究區域屬於櫟林 帶,亦是一般所稱之山地盛行雲霧帶,氣候屬恆濕型氣候,濕度大,全年 無乾季(蘇鴻傑,1992;Su, 1984a)。
三、地質與土壤
棲蘭山地區地質大部分都經過變硬或變質的第三紀巨厚泥質沈積岩組 成,包括深灰或灰黑色劈理良好的硬頁岩(Argillite)、板岩(Slate)以及千 枚岩(Phyllite)等(王鑫,2000);土壤質地多屬坋質壤土或坋質黏壤土,少
部分地區為砂質黏壤土,土壤多呈強酸性,pH 值常在 3.5-4.5。
表4 棲蘭山地區之氣候表 月份
項目
1 月
2 月
3 月
4 月
5 月
6 月
7 月
8 月
9 月
10 月
11 月
12 月 降雨量(mm) 37.0 100.0 113.5 82.5 143.0 76.5 266.5 1252.0 361.5 337.0 23.5 277.5 平均氣溫(℃) 5.1 6.8 10.2 12.8 15.3 16.1 16.6 16.8 15.6 10.0 10.1 7.0 相對濕度(%) 94.2 86.4 89.7 88.3 92.5 91.7 93.9 93.6 98.1 92.5 93.4 95.2
註:本研究以棲蘭山氣象站所測得之氣象資料為依據(2004)
0 50 100 150 200 250 300 350 400 450 500 550 600 650
1 2 3 4 5 6 7 8 9 10 11 12 月份
溫度
0 100 200 300 400 500 600 700 800 900 1000 1100 1200 1300
雨量
圖1 棲蘭山地區生態氣候圖(摘自棲蘭山氣候測站,2004) 溫
度
℃
降 雨 量 mm
月份
圖2 研究區域範圍位置
邊吉岩集水區
160-170 集水區 鴛鴦湖集水區
120-130 集水區
肆、研究項目與研究方法
一、資料收集
本研究自 2005 年 6 月開始,首先著手蒐集與本研究區域相關之基本資 料,包括二萬五千分之一比例尺之經建版地圖與五千分之一比例尺之正射 投影圖,以及棲蘭山地區與鄰近地區之植群類型、氣候、土壤質地和地形 等基本環境資料。在研究之前現場實地勘查的工作是很重要,可藉由實地 勘查確定研究區域內的交通路徑系統,並藉著沿途的觀察、拍照與植物採 集來暸解當地植群的類型、組成結構、分布概況以及環境的變化情形(圖 3)。
二、植群多樣性之取樣
本研究之取樣法採用計數樣區法(Count plot method)(劉棠瑞、蘇鴻傑,
1983),同時亦考慮研究區域內之地形與林木在空間上的分布型式,選擇地 形與林木組成較為均質(Homogeneous)的林分,並設置 10 個 10 × 10 m
2
之小 區(Plot)集合為一個合成樣區,若樣區位於斜坡上則需將樣區長軸與坡向垂 直以橫向方式設置(Golden, 1979)。量測樣區內胸高直徑(DBH)大於 1 cm 以上之木本植物記錄其種類名稱 與胸高直徑,而樣區內之小灌木、地被、附生及藤本植物則以記錄覆蓋度 及種類名稱的方式進行,另外,在調查時所採集之植物標本鑑定後製作為 證據標本存放於宜蘭大學標本館,以便於後續研究複查之用。此外,樣區 的調查除對植物種類及胸高直徑及覆蓋度進行記錄外,在樣區環境因子同 時觀測海拔高度(Altitude)、坡度(Slop)、方位(Aspect)、含石率(Stoniness)、
全天光空域(Whole Light sky Space, WLS)、直射光空域(Direct Light sky Space, DLS)、地形位置(Topographic position)(蘇鴻傑,1987a)。本研究共調 查54 個樣區,各樣區位置的分布如圖 2。
三、環境因子觀測與評估
植群生態(Vegetation ecology)之研究對象,主要為野生植物在不同生育
地之分布,以及各種天然植物社會或林型(Forest type)之識別與分類,然亦 須以環境因子之評估及其與林型之相關性,作為研判及解釋之基礎(蘇鴻 傑,1987a)。
在植群研究調查時,經常將眾多的環境變數整合成幾個較重要的環境 指數,並利用這些指數找尋出與植群變異之相關性,給予適度及合理的解 釋,若未能將眾多的環境變數整合來探討環境的變異相關性時,在植群的 變異上則無法得到一個完善的解釋與結果(蘇鴻傑,1987b)。本研究採用的 環境因子評估項目與方法如下:
(一)海拔高度
海拔為高層次影響植群之環境因子,可作為局部氣候之評估值,
觀測時以全球衛星定位系統(Global Positioning System, GPS)配合地圖 位置直接讀取為評估值。
(二)坡度
坡度又稱為傾斜度,即為樣區之傾斜角度,其影響土壤含水量之 保存及排水之狀況。坡度還控制著太陽之入射角度,影響生育地太陽 輻射的強度及局部的氣候變化。觀測時利用傾斜儀於樣區內量測若干 次,取其平均值為代表,並以角度表示之。
(三)方位
方位係指樣區或生育地最大之坡度所面臨的方向,方位的差異將 會造成對溫度、日照、濕度與土壤含水量之影響,通常方位可由指北 針直接觀測之,但其數值大小與其產生的效應並無相關性。若要探討 與植物間的關係,通常將角度轉化為效應之相對值來代表其所影響的 環境因子,此一影響梯度常以水分機制稱之。依Whittaker的方法採用 16個方位加以評估,於山坡下側及溪谷者不計方位,深谷且面臨溪水 者,其值給予1;淺谷無明顯溪水者,其值給予2;隱蔽之山坡下側,
其值給予3;其餘的地形位置則依方位,由東北至西南給予4-10之值,
值越高表示越乾燥(蘇鴻傑,1987a)。
(四)含石率
對於野外植群研究調查者而言,若要以現場實測一定體積的土體 含石率是相當的困難,大多以現場直接評估的方式,粗略的評定土壤 裸露岩石塊的程度,給予觀測之數值(蘇鴻傑,1987a)。
(五)全天光空域
樣區所在位置上方之空域經扣除受周圍地形地物遮蔽的部份,所 得之天空比例,稱為全天光空域。量測的方式是先將256點平均分配於 圓面積,並實地量測出樣區周圍8個方位遮蔽物各高低轉折點之方位角 及高度角後,再將這些數值分別標於圓面積上,經連結各轉折點,以 未遮蔽區內之點數為分子,圓面積上之256點數為分母,所得之百分率 即為樣區的全天光空域值(蘇鴻傑,1987a)。
(六)直射光空域
ㄧ年之中,太陽在樣區上空運行軌跡線所夾之天空面積,扣除周 圍地形遮蔽部份,所得之比例稱為直射光空域。此二軌跡線即為夏至 及冬至兩天之太陽軌跡。同樣地,此二線中未受遮蔽之點數為分子,
全部點數100為分母,所得之百分率為樣區之直射光空域值(蘇鴻傑,
1987a)。
(七)地形位置
地形位置係指生育地之位置與當地地形形成起伏的相對關係,通 常山地地形位置可分為山頂、稜線、上坡、中坡、下坡、溪谷旁、溪 谷等項目作為描述,而地形位置主要影響生育地的土壤含水量及乾性 至濕性之局部氣候。本研究採用蘇鴻傑(1987a)的觀念並將指數轉化為 1-7個評估值,其代表由濕至亁之土壤水分的情形,各指數所代表的位
置為:1:溪谷;2:溪谷旁;3:下坡;4:中坡;5:上坡;6 :稜線;
7:山頂。
四、原始資料計算
重要值指數(Important Value Index, IVI)係指樣區內植物社會基本介量 之綜合表示,即為相對密度、相對頻度、相對優勢度三者綜合而來,每一 介量之最大值為100%,因此三種植物重要值之總和最高可達 300%,此值 代表某植物在該林分中之相對重要性,及兼顧密度、頻度與優勢度之綜合 特性(劉棠瑞、蘇鴻傑,1983)。本研究將所獲得之樣區資料區分為喬木、灌 木、地被、附生、藤本植物 5 種生活型,並分別計算各層級之重要值指數。
在計算各樣區的喬木與灌木植物時則分別計算其密度、頻度、優勢度後轉 化為相對值;在計算地被、附生及藤本植物時則僅計算其相對覆蓋度與相 對頻度並將兩者相加之總和除2,各層級之重要值指數最高僅為 100%。在 完成各物種重要值指數之計算後,並將其計算之值依據Gauch (1982)八分級 制(Octave scale)將各物種重要值指數轉化為 0-9,共十級。上述樣區資料之 計算是以Excel 編輯程式輸入後在個人電腦上執行,在環境因子的資料部分 是不做任何的轉化,直接以觀測或評估值輸入於電腦備以將來做為分析 用。
圖3 棲蘭山地區多樣性研究流程圖
確定研究動機與目的
界定研究方向內容範圍
蒐集研究區域 環境相關資料 蒐集前人
相關研究文獻
研究地區勘察 初步區分植物社會
設定調查路線 初步植物鑑定
文獻整理回顧 樣區選擇與取樣方法
野外調查:植物資料觀測 環境因子估測
資料統計:製成原始矩陣表
植群資料分析
清查多樣性:植群多樣性、
物種多樣性(α、γ-多樣性) 分化多樣性:β-多樣性
綜合分析、討論 初步結果
結 果 參考陳子英, 2004
植群分型探討植群與環境間 之 相 關 性 (DCA 、 CCA 、 TWINSPAN)
參考蘇鴻傑,2004
五、植群分析方法
目前在植群分析上均採用多變數分析(Multivariate analysis)方法,此方 法可區分為三大類:1.直接梯度分析法(Direct gradient analysis)、2.分布序列 法(Ordination)、3.分類法(Classification)。直接梯度分析法:即為直接測試 物種與環境間的關係;分布序列法:必須先分析樣區資料(原始矩陣)中之資 料結構求得物種之變異方向後,再進行變異方向與環境因子間的測試,以 求出兩者之相關性,進而推測影響植物分布之環境梯度,並尋求植群與環 境間的關係,此種分析法係將樣區在環境梯度中依某種次序加以排列,然 測試前對於環境因素的影響是未知的,因此又稱為間接梯度分析(Indirect gradient analysis);分類法:即係將樣區資料加以比較,把相似之樣區加以 合併,組成某種林型或植群型(Vegetation type),然後再由環境因子之資料 推測各型之生育地環境,並將各型予以命名。
本研究所採用之植群分析法與其原理之詳述如下:
( 一 ) 直 接 梯 度 分 析 法 之 典 型 對 應 分 析 法 (Canonical Correspondence Analysis, CCA)。CCA 是由對應分析法(Correspondence Analysis, CA)與交互 平均法(Reciprocal Averaging, RA)修改後所產生的新方法,它是把 CA/RA 與 多元回歸結合起來,每ㄧ步驟之計算結果都與環境因子進行回歸,並詳細 研究植群與環境間的關係。CCA 進行分析時需要兩個資料矩陣,一個為植 群資料矩陣,另一個為環境資料矩陣,且其仍可結合更多的環境因子一起 分析,並更加能反映出植群與環境間的關係。CCA 係以 CA/RA 為修改基 礎,同時亦存在著與 CA/RA 相同之拱形效應(Arch effect)等缺點(張金屯,
1995;2004),但此效應可藉由移除過多的環境因子而消失,所謂過多的環 境因子即是指第二軸有極高的相關因子而言。過去研究專家進行 CCA 分析 前皆先採用 DCA 進行軸長的測試後在進行 CCA 的分析(張金屯,1995;
2004)。
( 二 ) 間 接 梯 度 分 析 法 之 降 趨 對 應 分 析 法 (Detrended Correspondence Aanalysis, DCA)。DCA 之基本運算原理與 CA/RA 相同亦是利用加權平均法 反覆計算至前後兩次之計算值無任何差異為止,但由於CA/RA 所計算之序 列 軸 會 因 為 運 算 方 法 的 缺 陷 , 導 致 所 計 算 之 序 列 軸 產 生 軸 端 壓 縮 (Compression of axis ends)及拱形效應,因此 DCA 是針對兩個效應進行修 正,其修正之方法即係將 CR/RA 每一軸所求得之序列分數給予重新刻劃 (Rescaling),以消除軸端壓縮,而在第二軸之後的運算中每次反覆 WA 之計 算時,均進行降趨(Detrendeding)之步驟以降低拱形效應(蘇鴻傑,1987b;
張金屯,1995、2004)。
(三)分類法之雙向指標種分析法(Two-Way Indicator SPecies ANalysis, TWINSPAN)。目前雙向指標分析被廣泛運用於植群分類上,此為兼顧定性 與定量之多元切分法(Polythetic dividive classification)的一種,其運算原理係 先將樣區資料先以交互平均法做排序,同時做兩極化之區分,並接著於排 序軸接近終點處作一切分,將其切分為二群,以此類推,在對各群進行切 分,切分的次數通常以所含之樣區數少於某一預設值時該群及不再作細 分,最後則會產生一個安置矩陣(Arranged matrix),樣區與樹種之層級與分 類即可由此結果表示(蘇鴻傑,1996)。
上述三種植群分析法主要用於探討植物社會的分化與環境間的相互關 係,在環境方面一般常採用 CCA 釐清影響植群分化的因子。DCA 則配合 TWINSPAN 清查地域內植物社會的數目及分類。並由 PC-ORD 套裝軟體於 個人電腦上執行(張金屯,1995;2004;McCune, 2002)。
六、多樣性分析方法
在進行清查與分化多樣性的研究時,大多數以物種的多寡來判斷該區 域之多樣性豐富與變化的程度,若研究區域內之優勢物種多集中於少數幾 個物種時,還採用物種種類之多寡的觀念進行多樣性的量測時則無法詳細
的瞭解該區域內物種多樣性變化的程度。由上述觀念得知,本研究在進行 多樣性的量測時,若未將調查所獲得之物種資料依各物種之生活型做區分 則無法瞭解棲蘭山檜木林各集水區內物種多樣性的變化程度。
本研究對於各生活型的分類係參照下述研究文獻所定義,如台灣恆春 半島里龍山(葉清旺,2004)、尼泊爾喜馬拉雅山(Bhattarai and Vetaas, 2003)、
中國祁連山(Wang et al., 2002)墨西哥熱帶乾燥林(Patricia, 2002)、中國長白山 (邵彬,1999)及日本 Tatera 保護區(Itow, 1991)的喬木及灌木植物,玻利維亞 安底斯山(Kessler, 2000)、墨西哥 Cerro Grand 山區(Vazquez and Givnish, 1998) 地被植物、藤本植物及附生植物。將本研究區域內所調查之物種依其生活 型區分為喬木、灌木、草本、藤本、附生植物等五個層級分別利用各種指 數計算各層級中物種多樣性與變化程度。以便瞭解各物種對於生存之多元 環境空間的需求即稱為生態棲位(Austin, 1985),並依不同的生態棲位來找尋 各物種在生態上之意義。並藉由生活型的區分,充分瞭解棲蘭山地區內之 物種多樣性豐富的程度。
(一)清查多樣性 1. α 多樣性
本研究在 α 多樣性的測計上採用的指數分為物種豐富度指 數即為物種的數目、物種多樣性指數和物種均勻度指數。
(1)物種豐富度
係指在一定範圍內所量測的物種數量,即可以代表該地區物 種之豐富程度,但此方法容易受到樣區面積大小的影響,進而影 響樣區物種的豐富度,因此樣區面積大小的確定對於物種豐富度 是很重要的。本研究在物種豐富度上即為採用0.1 ha 樣區面積內 之物種數表示之。
(2)物種多樣性指數
物種多樣性係指物種種數與均勻度結合的單一統計量,在生 物社會中,若其物種極多,物種豐富度非常均勻,則該生物社會 則具有高的多樣性;相對地,若生物社會物種較少,物種豐富度 不均勻,則多樣性也隨之較低。近年來物種多樣性得到廣泛的應 用,由於不同學者不同研究對象,因此對於量測方法也有所差 異,故本研究採用目前較最常被應用的兩種多樣性指數進行測 計。
a. Simpson 指數
Simpson 指數又稱為優勢度指數,優勢度本身可指示生物社 會中物種多樣性,惟優勢度越高,多樣性則越低,優勢度越低,
則多樣性越高,兩者為逆關係。Simpson 指數介於 0-1 之間,若 D 值越大則多樣性越高;反之,D 值越小則多樣性越小(劉棠瑞、
蘇鴻傑,1983;Simpson, 1949)。
( )
∑
= −∑
−
=
−
=1 C 1 ni/N 2 1 Pi2
D ……….…式 1
式中ni:某一樹種之株數;N:樣區內所有樹種之株數總和
b. Shannon-Wiener 指數
Shannon-Wiener 可能或然率之自乘,而係乘以本身之自然對 數。Shannon-Wiener 指數也是群體生態學中應用較為廣泛,為植 物社會的特徵之一。
(
ni N) (
ni N)
Pi PiH
= − ∑ / ln / = − ∑ ln
………...……...式 2 式中ni:某一樹種之斷面積;N:樣區內所有樹種之斷面積總和。由式 2 得之,可能率之自然對數為負值,因此在公式前再多 加一個負號,將其負值轉變成為正值。此值最大者並無固定,物 種數越多,個體數分布越均勻,則 Shannon-Wiener 多樣性指數