誌謝
時光匆匆流過,我的碩士生涯就要正式落幕了,兩年的碩士班 看似漫長,實則如過眼雲煙轉瞬即逝。人生的美好不在於延續生命 的永恆不滅,而在於瞬間璀璨所散發出來的繽紛光芒,雖然兩年時 間匆匆而逝,但已在我的人生之中留下許多不可磨滅的奪目光采。
首先最要感謝的是施美份老師,從入學前的暑假一直到畢業的 暑假,整整兩年寒暑的辛勤教導,雖然學生資質愚鈍,學如逆水行 舟,時常未能如老師所期盼中的成長,但老師依然帶我一路勇往直 前奮力不倦,不僅於實驗上給予指導,更教曉我做人做事的態度,
如今輕舟已過萬重山,老師對我的照顧及付出,點滴在心,無限感 激!
另外,要感謝實驗室已畢業的敏華學姊,對我在實驗技術上的 細心及不吝指導。還有藥學系的紹彥學弟和老師的專任助理佳蓉,
感謝你們給予我在實驗上的協助和提供寶貴的意見,俾使論文得以 順利完成。
接著還要感謝在 F 棟、Q 棟 7 樓及 9 樓實驗室的同學們,阿 邦、阿立、秀珍、佳蓉、學益、建安、敏修及碩士班各位同學等 人,大家有緣聚首,謝謝你們平時的陪伴與分享實驗上的經驗和一 些喜怒哀樂的瑣事,使我在碩士班兩年不感孤獨,並能於良好的心 情全力以赴完成我的論文。
此外,還要特別感謝我的爸爸、媽媽及我的老友聖岡,如不是 你們一直以來的鼓勵與支持,我也不可能有今天的小小成就。當然 還要感謝這兩年對我不離不棄的伊廷,妳就像是我手心的太陽,總
在我忙碌無助時,適時給予我安定的力量,支持我一路走下去,謝 謝妳!
最後,要感謝的人實在太多,如有被遺忘的朋友在此亦一併感 謝,感謝大家於這兩年來對我的照顧。有些人的碩士生涯過的如黑 白像片般愁雲慘淡,但我的碩士生涯用資訊豐富的高光譜影像來描 述亦不為過,在此也祝福所有畢業的同學們都能夠在未來的路上綻 放出各式各樣的光芒!
僅將誌謝獻給每一個曾經在我的人生路上給我鼓勵的你們。
中文摘要
長期性細菌感染以及常見性感染是糖尿病患者主要的併發症之 一,這兩者皆與發炎細胞之活化具有相關聯性。然而,長期高濃度 葡萄糖對於巨噬細胞活化作用,卻無太多相關性的研究。
因此,本研究以 LPS 刺激 RAW 264.7 巨噬細胞,作為一個實驗 的模型,再以 15 mM 高濃度葡萄糖培養液去探討不同的培養時間,
與正常 DMEM 培養下的 RAW 264.7 巨噬細胞,分別對於 LPS 刺激 後的情形。實驗結果發現,RAW 264.7 巨噬細胞在15 mM 高濃度葡 萄糖培養 1 天(急性), 7 天(中慢性)和 14 天(慢性) 之後,
basal NO 產生量會比正常細胞的多,而 LPS 刺激的 NO 產量卻明顯 不如正常細胞。然而,在 iNOS protein 與 iNOS mRNA 的表現上,
經長期慢性培養後,卻有反彈性大量增加的情形。
添加 15 mM 葡萄糖培養液培養 1 天及 14 天的 RAW 264.7 巨噬 細 胞 , 於 LPS 刺 激 後 所 產 的 促 發 炎 細 胞 激 素 TNF-α,與正常 DMEM 培養的細胞相比較是沒有明顯差異。不過,LPS 刺激的 TNF-α 產量在經過 15 mM 葡萄糖培養液培養培養 7 天後的巨噬細胞 卻明顯比正常 DMEM 培養的細胞的產量少。15 mM 葡萄糖培養後 的 RAW 264.7 巨噬細胞,所產生的 IL-1 β 產量在 basal 值與正常 DMEM 培養的細胞相比,均有明顯大量誘發的情形。培養 1 天及 14 天 15 mM 高濃度葡萄糖,在添加 LPS 刺激後,所產生的 IL-1 β 濃 度與正常 DMEM 培養的細胞相比,有較為明顯上升的情形。然而,
在 IL-6 的表現量方面,經 LPS 刺激 15 mM 高濃度葡萄糖培養 1 天 後的 RAW 264.7 巨噬細胞,與正常 DMEM 培養的細胞相比,有著
明顯上升的情形。有趣的是,在培養 7 天後,IL-6 的產量有被抑制 的情況,而 14 天高葡萄糖培養後,卻又有一個回升的現象。
然而造成這些結果的原因,可能與細胞隨著高濃度葡萄糖長期 培養後,而產生適應性和一個生理功能上正向調節 ( up-regulation ) 的情形。隨著葡萄糖培養之 RAW 264.7 巨噬細胞,經由 LPS 活化細 胞後,所誘發的促發炎細胞激素 ( TNF-α, IL-6, IL-1 β ) 皆與 iNOS protein 和 iNOS mRNA 的表現有著正相關性。
關鍵字:糖尿病、一氧化氮、發炎反應、高血糖症、細胞激素。
Abstract
One of major complications of diabetes is the frequency of infection and long-term duration of bacterial infections. Both conditions have been shown to be related to activation of inflammatory cells. However, the influences of macrophage activation by long-term exposure to high glucose, a situation that mimics the hyperglycemia of diabetics, have not been fully investigated.
We used a lipopolysaccharide ( LPS ) activated macrophage RAW 264.7 model to investigate effects of acute ( 1 day ), sub-chronic ( 7 days ), and chronic ( 14 days ) 15 mM glucose treatment on activation of inflammatory cells. Basal NO production was higher in all glucose treated groups than that produced in normal cells. In contrast, LPS- induced NO production was sustained lower in the glucose treated cells than in the normal group. Interestingly, LPS-stimulated iNOS protein or iNOS mRNA expression in chronic glucose treated cells showed a more condensed bend than acute, sub-chronic or normal cells.
Treatment of RAW264.7 macrophages with 15 mM glucose for 1 and 14 days did not affect LPS-stimulated tumour necrosis factor-alpha ( TNF-α ) production. However, the same treatment for 7 days suppressed TNF-α production. Basal interleukin-1 beta (IL-1β) was increased significantly in high glucose treatment groups than that in normal control group. LPS-stimulated IL-1 productions were also increased except for 7-day treated group. Acute glucose challenge induced both basal and LPS-stimulated interleukin-6 ( IL-6 ) production.
These increases were both suppressed in sub-acute groups. Interestingly, chronic glucose treatment did not affect basal IL-6 production but the production was raised the LPS-stimulated condition.
These results shown that may be due to an up-regulation or adaptation change after cells exposed to glucose chronically. Pro- inflammatory cytokines, TNF-α, IL-1β, and IL-6 that mediate the activation of macrophage via LPS stimulation, are also shown a similar adaptive pattern as iNOS protein and mRNA expression in responding to glucose treatment.
Key words: Diabetes, Nitric Oxide ( NO ), inflammation, Hyperglycemia, cytokines ( TNF-α, IL-1 β, IL-6 ).
目 錄
頁次
中文摘要………I 英文摘要………...III 本文目錄.………...V 縮寫表……….VIII
第一章、緒論……….1
1-1. 糖尿病相關研究………1
1-2. 糖尿病的定義………2
1-2-1. 糖尿病的診斷………..………...2
1-2-2. 糖尿病的分類………..……….……..3
1-2-3. 糖尿病之流行病學………...…..4
1-3. 糖尿病與感染………...5
1-3-1. 糖尿病與其細胞免疫調節………..………....6
1-3-2. 免疫反應與巨噬細胞………..………7
1-3-3. 細菌內毒素與巨噬細胞…………..………7
1-3-4. 發炎反應與巨噬細胞………..………8
1-3-5. 一氧化氮 ( Nitric Oxide )…………..………..8
1-3-6. 腫瘤壞死因子 ( TNF-α )………….………9
1-3-7. 介白素-1 ( IL-1 )…………..…..……….10
1-3-8. 介白素-6 ( IL-6 )………..………...10
1-3-9. 一氧化氮合成酶 ( iNOS )…..………..…………...11
1-4. 研究動機…………..………...………12
第二章、材料與方法………...14
2-1. RAW 264.7 巨噬細胞之培養………...14
2-1-1. RAW 264.7 巨噬細胞之繼代培養……….15
2-1-2. RAW 264.7 巨噬細胞之冷凍..………...16
2-1-3. RAW 264.7 巨噬細胞之解凍..……...17
2-2. 15 mM 高葡萄糖 DMEM 培養 RAW 264.7 巨噬細胞...…….17
2-3. 細胞計數 / 種植細胞..……..……….….18
2-4. 亞硝酸鹽產物測定………....…….19
2-5. TNF-α 濃度測定………...……….20
2-6. IL-1 β 濃度測定…………...……….……….23
2-7. IL-6 濃度測定………...……….………...25
2-8. 蛋白質抽取……….……….…...27
2-9. 蛋白質濃度測定…… …………..……….…….28
2-10. 西方墨點法 ( Western Blot )………..………..……….……...29
2-11. RNA 抽取………..……..……….………31
2-12. 反轉錄-聚合酶鏈反應 ( RT-PCR )……..………33
2-13. DNA 瓊膠製備/ DNA 電泳…………..……….……..34
2-14. 統計方法………...35
第三章、結果………...36
3-1. NO 產量試驗……..………..…...36
3-2. iNOS protein assay by Western Blot...……….…...37
3-3. iNOS mRNA expression by RT-PCR.………..……….……...37
3-4. TNF-α 產量試驗.…………..………...38
3-5. IL-6 產量試驗..…...………...39
3-6. IL-1 β 產量試驗……..………….………...39
第四章、討論………..41
第五章、結論………..45
參考文獻………..46
附錄………..60
圖1.a 1 day glucose treatment on basal NO production………...60
圖1.b 1 day glucose LPS stimulated NO production………...61
圖1.c 7 day glucose treatment on basal NO production………...62
圖1.d 7 day glucose LPS stimulated NO production…………...63
圖1.e 14 day glucose treatment on basal NO production………64
圖1.f 14 day glucose LPS stimulated NO production………...65
圖2.a iNOS protein on basal expression by Western Blot………..66
圖2.b iNOS protein on LPS stimulated expression by Western Blot…..67
圖3.a iNOS mRNA expression by RT-PCR………68
圖4.a Effect glucose treatment on basal TNF-α production…………...69
圖4.b Effect glucose treatment on LPS stimulated TNF-α production...70
圖5.a effects of glucose treatment on basal IL-6 production……...71
圖5.b effects of glucose treatment on LPS-stimulated IL-6 production.72 圖6.a effects of glucose treatment on basal IL-1 β production…...73 圖6.a effects of glucose treatment on LPS stimulated IL-1 β production74
縮寫表
IFG; impaired fasting glucose IGT; impaired glucose tolerance NDDG; National Diabetes Data Group WHO; World Health Organisation
MRDM; Malnutrition-related diabetes mellitus HLA; human leukocyte antigen
LPS; lipoplysaccharide NO; Nitric Oxide
TNF-α; tumour necrosis factor α IL-1; interleukin-1
IL-6; interleukin-6
PDGF; platelet-derived growth factor PAF; platelet-activating factor
NOS; Nitric Oxide Synthase
iNOS; inducible Nitric Oxide Synthase
第一章、緒論
1-1. 糖尿病之相關研究
隨著國人生活品質日漸的提高,醫療科技的進步,全人類壽命 的延長,高齡人口快速增加,慢性病漸漸取代傳染性的疾病,而在 慢性病之中的糖尿病,一直位居國人十大死因之中。 根 據 衛 生 署 2005 年統計,糖尿病死亡率近二十年來有不斷成長的趨勢,從 1982 年每十萬人口 10.82 人,到 2003 年的 44.38 人,增加了 4.1 倍 。 過 去 十 年 來 , 糖 尿 病 在 十 大 死 亡 原 因 中 都 維 持 在 第 五 位,但 2002 年上升至第四位,2003 年亦維持不墜。年齡 45 歲 以上者糖尿病盛行率達到 11 %,也就是每 10 個人之中就有一個 人罹患糖尿病 (1)。
過去的十年之中,有許多文獻報導指出肥胖與糖尿病及慢性發 炎反應之間有著密不可分的正相關性 (2) (3)。肥胖是因為過度的內臟 脂肪堆積所造成的現象, 從預估身體腹部的內臟脂肪,可以來推算 罹患糖尿病的機率 (4),故肥胖是造成糖尿病的一個重要因素之一。
當腰臀圍比增加時,腹部內的脂肪也就會相對增加,並促使血中游 離脂肪酸也隨之增加,進而阻礙肝臟與胰島素的攝取及代謝,最後 造成高胰島素血症 (5),引起高血壓 (6),導致糖尿病的產生。肥胖的 人發生糖尿病的機率是非肥胖者的 2.9 倍 (7),男性與女性在脂肪堆 積的程度上是有所差異的 (8) ,男性方面會隨著年齡的增加而增加,
以 50 歲做分隔,為好發年齡 (9),女性則是在停經的前後做為分隔 點。
1-2. 糖尿病的定義
根據美國糖尿病學會專家委員會 1997 年報告定義,糖尿病是由
於胰臟 β 細胞所分泌胰島素的功能異常或是胰島素分泌和作用缺
陷,以致生理功能代謝無法發揮降血糖的作用時,而引起的高血糖 症狀,進而衍生出來的一種新陳代謝疾病。人體所攝取的食物,經 過消化、吸收、轉化成葡萄糖之後,血液中的葡萄糖含量就會累積 超出了正常範圍。病人臨床症狀為高葡萄糖血症,伴隨其他的臨床 症狀為多尿、多喝、多吃、體重減輕、視力模糊、疲勞及容易感染 等。
糖尿病的患者沒辦法像正常人一樣,他們會有代謝異常及慢性 血糖升高的情況。是由於長期處在高血糖之中,使得組織蛋白和大 分子物質受到糖化,或是由於長期的高血糖,產生過多的聚合化合 物,均會使一些器官受損,亦時常伴隨著許多長期的併發症 (10),例 如:眼睛、腎臟、神經、心血管,都會導致機能障礙甚至衰竭。
1-2-1. 糖尿病的診斷
糖尿病是一種多重原因的慢性疾病,雖然臨床表徵均以高血糖 為主,但致病原因各異。目前糖尿病的診斷仍以血糖濃度測定為依 據。依美國糖尿病學會對於糖尿病的診斷標準,就是空腹血糖偏高 ( impaired fasting glucose; IFG ) (11) 及葡萄糖耐受性障礙 ( impaired glucose tolerance; IGT ) (12) 兩種情形。
前者是需整夜空腹 8 小時後抽血檢查,血糖介於 110 mg/ dL 與 126 mg/ dL 之間(達到 126 mg/ dL 以上就屬於糖尿病),後者則有 賴整夜空腹 8 小時後接受口服葡萄糖耐量檢查兩小時血糖值,介於 140 mg/ dL 與 200 mg/ dL 之間(達到 200 mg/ dL 以上就屬於糖尿
病)。
1-2-2. 糖尿病的分類
世界衛生組織於 1980 年發佈了糖尿病的分類方法,完全採用美 國國家糖尿病資料組 ( National Diabetes Data Group; NDDG ) 的分類 方法,但到了 1985 年世界衛生組織 ( World Health Organisation;
WHO) 另 外 添 增 一 個 新 的 糖 尿 病 類 別 , 及 營 養 失 調 糖 尿 病 ( Malnutrition-related diabetes mellitus; MRDM )。
第一型糖尿病 ( type 1 diabetes ):由於胰臟 β 細胞的破壞,導致 胰島素缺乏,通常在 40 歲以前發生。其第一型糖尿病並患者的特徵
是逐漸失去胰臟蘭氏小島的 β 細胞,最後導致循環中無胰島素存
在,病因被認為與人類白血球抗原 ( human leukocyte antigen; HLA ) DQ、DR 最有關聯 (13),故第一型糖尿病的病因被認為和抗原表現有 關的遺傳性自體免疫疾病。然而這類的病人因有抗胰島組織抗體,
所引起 T 淋巴球浸潤,最後摧毀胰臟蘭小島。目前第一型糖尿病的 所有病人皆須依賴補充胰島素來控制血糖。
第二型糖尿病 ( type 2 diabetes ):初期為胰島素有阻抗性,後期 為 胰 島 素 分 泌 不 足 , 大 多 由 於 胰 島 素 接 受 器 的 下 調 節 ( down regulation ) 造成胰島素接受器濃度降低,或是在細胞表面接受器之 後,細胞內的葡萄糖代謝有缺陷。以上種種現象造成患者血中有足 夠的胰島素,卻無法對於使細胞正常攝取葡萄糖。此類型糖尿病通 常在 40 歲以後發生,患者多為肥胖者,血漿胰島素多為正常或偏 高 。
其他特異型:成人糖尿病患中,至多 2 % 是由胰臟疾病 (急 性、慢性胰臟炎、胰臟癌等)及藥物(如利尿劑、避孕丸、類固醇
等)等引發的高血糖狀態,但符合現行糖尿病診斷標準,於是被歸 類為其他類糖尿病。
1-2-3. 糖尿病之流行病學
隨著國人生活品質的提高,飲食及生活形態的改變,糖尿病已 成為普遍分布全世界的慢性疾病,並且有逐漸增加的趨勢 (10)。世界 上約有兩億人口罹患糖尿病,再以開發國家中均排名主要十大死因 之內 (14) 。在台灣地區 2005 調查顯示,患有糖尿病的人數約在 30 ~ 40 萬人口左右,位居國人十大死因中的第四名,而且逐年以 ( +/ - ) 13.8 % 的年增率增加 (1),顯示了對於糖尿病的控制及延緩其併發症 的發展上仍有很大的改善空間。
糖尿病是一種慢性疾病,目前僅可控制而無法根治,其最擾人 的是慢性併發症的問題。在第一型糖尿病中,主要微血管病變居 多,例如:腎病變 (15) 、視網膜病變。而第二型糖尿病則是以心臟血 管與腦中風方面居多 (16)。
腎病變方面,糖尿病所引起的腎病變是造成尿毒症,亦是患者 需要洗腎的主要原因。最先出現的臨床表現是蛋白尿,其後肌酸酐 也逐漸上升,大約有三、四成的第一型糖尿病人最後演變為尿毒 症,需要洗腎。在蛋白尿出現之前,其實腎臟已有變化,例如:血 流增加,腎絲球過濾率增加,體積變大等。糖尿病與腎病變之間的 關係在免疫學上是有著互相影響的情況,但在病理機轉上仍然不清 楚 (15)。
糖尿病視網膜病變在歐美等已開發國家中是造成失明的主要原 因之一 (17)。而視網膜病變在台灣地區現況而言,研究發現糖尿病併 發視網膜病變盛行率在 25 % ~ 30 % 之間 。所謂糖尿病視網膜病
變,是因為視網膜上佈滿纖細的微小血管,非常容易受到高血糖的 破壞,糖尿病患中約有 10 % 到 12 % 的人,因此而失明 (18)。其發生 率與患者年齡及罹患糖尿病時間的長短有密切的關係 (19)。有 25 ~ 30
% 胰島素依賴型(第 1 型)糖尿病患者發病 10 ~ 15 年後會有視網膜 病變的徵象,而 23 % 的非胰島素依賴型(第 2 型 )糖尿病患者發 病 11 ~ 13 年後會有視網膜病變的徵象 (20)。糖尿病視網膜病變,初 期稱為基底型,視網膜上出現血門管的小阻塞,造成局部的缺氧導 致出血點及血管滲出物。此時病人多半無明顯症狀,若不及時檢查 並加以控制,一旦隨著疾病進行演變成後期的增殖期,視網膜及視 神經盤增殖出易破裂的新生血管,嚴重時便可能引起玻璃體大出 血,甚至牽引視網膜剝離,導致視力大量減退而致失 (18)。
1-3. 糖尿病與感染
感染一直以來為糖尿病患者最大的問題之一,也因此常使糖尿 病患者受到足部截肢的命運 (21),而感染後所伴隨的現象就是發炎反 應,在過去一些研究報導中 (16) (22) (23) 指出糖尿病與發炎反應之間有 著相正關聯性。發炎反應是人體第一道免疫反應,當人體受到感 染、刺激或是受傷時,就會引發免疫性的發炎反應。 臨床觀察發現 糖尿病病人易罹患感染,而且一旦遭受感染,其病情常較嚴重,使 得感染症成為糖尿病病人的重要致死原因。糖尿病併發感染的成因 有二大類,其一為病人身體的抵抗力之缺陷,如血管病變造成血流 供應不足、周邊神經病變和免疫力異常;其二為病人體內環境受到 糖化關係而適合某些致病原之滋長。
細菌性感染方面以足部感染為糖尿病患者最常見的細菌感染之 一,糖尿病合併周邊神經病變者容易造成足部潰瘍,這些潰瘍常合
併深部組織及骨頭之感染,如果未妥為處理,易導致截肢之命運。
糖尿病患者在罹患糖尿病 25 年內,有 10 % 會遭受截肢。糖尿病神 經病變患者因對疼痛感覺差,而容易受創傷並產生局部合併症。另 者,指溝炎也很容易造成感染擴散,皮膚黴菌感染造成皮膚傷口是 細菌侵入的管道。穿刺傷造成感染也很常見。糖尿病足部感染常由 多種細菌造成 (24) (25),平均約為 2 ~ 5 種細菌,包括需氧菌和厭氧 菌。需氧菌方面以革蘭氏陰性菌 ( Gram-Negative Bacillus; G(-) ) 最為 常見,而單一細菌感染中以革蘭氏陽性菌 ( Gram-Postive Bacillus;
G(+) ) 的金黃色葡萄球菌感染最常見,腸球菌次之。其他如厭氧菌之 類桿菌 ( Bacteroides )、綠膿桿菌 (26) 及白色念珠菌亦可見。
其次為黴菌感染,主要發生於宿主免疫防衛系統不健全者,造 成伺機性感染,也可能與一再重複使用抗生素有關 (24)。糖尿病病人 最常見的致病性黴菌是白色念珠菌、平滑念珠菌 ( Candidaglabrata ) 和接合黴菌 ( Zygomycetes ) 等。糖尿病病人易遭受感染的原因很 多,除了血管病變所致的局部血流不足和周邊神經病變外,免疫防 禦功能之缺陷亦扮演一部分角色 (27)。
1-3-1. 糖尿病與其細胞調節免疫
研究顯示糖尿病患者體外實驗的細胞調節免疫功能受到損害,
例如:血糖控制不良的病患,其淋巴球對於植物血球凝集素刺激 後,所產生的轉型能力 (指由 monocyte 轉變為 macrophage) 較正 常人微弱 (28)。儘管血糖已受到控制的病患,其淋巴球遭受葡萄球菌 抗原入侵時,轉型能力也較常人低下 (29)。
1-3-2. 免疫反應與巨噬細胞
免疫反應是身體抵禦外來感染的防禦機制。當病源體入侵時,
會先引起自體的免疫發炎反應,其中巨噬細胞 ( macrophage ) 可藉由 吞噬作用除去外來的入侵者。 巨噬細胞是由骨髓所製造出來的單核 球所成熟分化而成。他們的生命力比起一些多核細胞長,且具有吞 噬細菌、病毒的能力,亦是在免疫系統上扮演一個重要的防禦角色
(30)。
巨噬細胞在吞噬作用發生之前,這些微小的外來物必須先附著 在多形核球或巨噬細胞的表面上。這和一種可能包括碳水化合物的 原始辨識機制有關,一個微粒附著到表面上後可能會啟動肌凝-肌動 收縮系統 ( actin-myosin contractile system ) 而引起吞噬作用。首先形 成偽足 ( pseudopods ),然後當鄰接的受器接觸到微生物後,整個細 胞膜就像拉上拉鍊般地包微粒,直到完全將微粒包起,形成食泡為 止。大約在一分鐘內,細胞質內的容菌體便和食泡融合,釋出水 解,如此便可使外來物致死。有研究報告指出,當巨噬細胞活化 時,不僅會進行吞噬作用,同時也會釋放許多自由基物質(例如:
nitric oxide)來毒殺入侵的外來物 (31) (32)。
1-3-3. 細菌內毒素 ( endotoxin ) 與巨噬細胞
巨噬細胞在一般生理下是未被活化的,它的生理功能是受限制 的,須受到特定的刺激如:脂多醣體 ( lipoplysaccharide; LPS ) 或一 些細胞激素 ( cytokines ) 刺激下,才會被活化作用 (33)。脂多醣體為 革蘭氏陰性菌細胞璧的組成份之一 (革蘭氏陽性菌則無此成分),
巨噬細胞在 LPS 的誘發下會產生許多細胞激素,例如:interleukin-1
( IL-1 ) ,interleukin-6 ( IL-6 ) 及 tumour necrosis factor ( TNF ) 等,這 些細胞激素在生理上,均有重要的角色 (34)。
已 知 這 些 細 胞 激 素 可 誘 導 巨 噬 細 胞 、 平 滑 肌 細 胞 ( smooth muscle cells )、上皮細胞 ( epithelia ) 等細胞 iNOS ( inducble nitric oxide synthase ) 的表現。但 LPS 的刺激只對巨噬細胞可誘發出 NO ( nitric oxide ) (35), 其 他 如 上 皮 細 胞 、 平 滑 肌 細 胞 及 中 性 球 ( neutrophils ) 則無任何相關文獻提及其 LPS 對細胞的誘發作用。
1-3-4. 發炎反應與巨噬細胞
發炎反應是人體受傷時所引發的免疫反應之一,許多研究證 實,在慢性發炎及感染之細胞組織會有一系列的細胞激素產生(例 如:IL-1, TNF-α) (36) 、酵素及訊息蛋白生成。iNOS 及 COX-2 分別 可催化 NO及 PGs ( Prostaglandins ) 大量產生時,不但造成組織發炎 及併發敗血休克,亦使得慢性傳染病(例如:肺結核)和自體免疫 疾病(例如:風濕性關節炎)等疾病症惡化 (37)。
1-3-5. 一氧化氮 ( NO )
NO可在許多細胞內產生,例如:巨噬細胞 (38) (39) (40)、內皮細胞
(41)、平滑肌細胞 (42) 。當細胞受刺激後,會釋放出 NO 並賦予巨噬細 胞具有細胞毒殺活性來對抗外來物的入侵,例如:病毒、細菌、黴 菌、寄生蟲和腫瘤細胞 (43)。
NO 在生理上扮演的角色相當重要也相當多樣,例如:神經、
免疫、新血管系統等調節均可見其參與,過多的 NO 也會導致敗血 症休克、中風、血壓大量下降等疾病的產生 (44)。有研究報告指出,
活化後的巨噬細胞其殺菌及吞噬能力與 NO 的產生有關 (45),而在嚴 重的細菌感染情況下,巨噬細胞會產生大量的 NO 而導致敗血性休
克 (32) (46) 。此外在臨床上曾有研究指出抑制過量 NO 的產生,可治
療敗血症 (47)。
1-3-6. 腫瘤壞死因子 ( TNF-α )
在過去的二十年裡,肥胖的人持續增加。高卡路里的飲食、少 量的運動,往往導致許多糖尿病 (第二型)的產生 。肥胖的人對游 離脂肪酸的利用減少,易造成血脂中的游離脂肪酸堆積,血脂容量 升高,進而促使巨噬細胞中一系列的促發炎反應 ( pro-inflammatio- n ),引發細胞激素大量的釋放,例如:腫瘤壞死因子 ( TNF-α ) (48)。 在生理或病理因子(如 LPS, IL-1 等)刺激的情況下,人類血液單核
球 (49) (50) 及老鼠的巨噬細胞 (51) 中可引起腫瘤壞死因子的釋放。
TNF-α 是一種促發炎細胞激素 ( pro-inflammatory cytokine ),在人體 內也扮演為與細胞增生 ( proliferation )、分化 ( differentiation )、細胞 凋亡 ( apoptosis ) 有關的重要角色 (52) (53)。
TNF-α 亦 屬 於 一 種 多 元 作 用 的 細 胞 激 素 ( pleiotropic cytokines ),可影響許多發炎相關性的蛋白質表現,例如:發燒,加 強 IL-1 β 產生等。TNF-α 作用在發炎細胞,主要藉由細胞膜表面之 特殊接受體 TNFR1 ( p55 ) 或是 TNFR2 ( p75 ) (54) (55) 二者,再經由不 同 之 細 胞 內 訊 息 傳 遞 , 例 如 : mitogen-activated protein kinases ( MAPKs ) 或 是 NFκB-IκB 等 路 徑(56), 進 而 造 成 轉 錄 因 子 ( transcription factor ) 的活化及表現。
1-3-7. 介白素-1 ( IL-1 )
細胞激素 ( cytokines ) 或 interleukins 在各種不同組織與細胞中,
扮演著調節細胞增生與分化的重要角色。其擁有多樣性與豐富性的 特性,例如:一種細胞激素在不同的組織或是細胞中,有不同的機 能與功能,或是數種不同的細胞激素在同一個組織或細胞裡,有相 似的功能並且相互作用著。
IL-1 是由周邊血液中的活化單核球/巨噬細胞或組織中之單核 球 (57),巨噬細胞 (58) (59),淋巴球 (60) (61),嗜中性白血球 (62),及纖維
母細胞 (58) (63) 所製造。IL-1 是最早被發現細胞激素之一,依其分子
構造可分為兩種形態,IL-1 α 和 IL-1 β。兩者之間具有 48 % 的構造 同質性,生物活性上也互相類似 (64)。在一些刺激物質,例如:細菌 抗原、脂多醣內毒素、淋巴動素 ( lymphokine )、發炎物質等,均能 刺激細胞產生 IL-1 的分泌 (65) (66) (67) (68)。
Duarte 等人在 2007,年對於第二型糖尿病患者因牙周病所引起 的發炎反應中,檢測到IL-1 β、IL-6 等細胞激素皆有大量表現的情 形,意味著 IL-1 β 與發炎反應有著正相關性 (69)。而在第一型糖尿病 患者中,有很多多功能免疫的媒介者,例如: TNF-α 、IL-1 β 等都 會造成胰臟 β 細胞自體免疫功能的破壞 (70)。
1-3-8. 介白素-6 ( IL-6 )
IL-6 是一種免疫蛋白,亦是一個多功能的細胞激素。最初在誘 導 B 細胞成熟,在成為可分泌抗體的 B 細胞中被發現,因此稱為 B- cell differentiation factor ( BCDF or BSFp-2 ) (71) (72)。但是後來在人工 合成之重組蛋白 IL-6 及 IL-6 的抗體研究發現,IL-6 不僅僅作用於 B
細胞,另外也作用於 T 細胞、hepatocytes、hematopoietuc progenitor cell 和 neuronal cell (71) (72) (73)。
IL-6 其功能包括調節人體免疫反應、造血作用、急性期蛋白的 合成及發炎反應 (74)。 IL-6 也能夠增生如血小板的產生、血小板的活 化及促進凝固作用 (75)。活化後的吞噬細胞、 T 細胞、B 細胞、角質 細胞、纖維母細胞、內皮細胞及神經膠細胞等都可分泌 IL-6 (76)。淋 巴毒素 ( lymphotoxin ) 、 IL-1、TNF、血小板生長因子 ( platelet- derived growth factor; PDGF )、血小板活化因子 ( platelet-activating factor; PAF ) 和顆粒性球抑制蛋白 ( franulocyte-inhibitory protein ) 等 分裂原 ( mitogens ) 也會誘導 IL-6 的合成 (77) (78) (79) (80)。
IL-6 與IL-1 β、TNF-α 在第二型糖尿病患者中也扮演著一個調 節發炎反應的角色 (81)。文獻報導指出糖尿病合併視網膜病變的患者 有明顯的 IL-6 上升表現 (82)。由第一型糖尿病之周邊血液中單核球分 化而來的巨噬細胞受 LPS 刺激後所產生的 TNF-α、 IL-6 和 IL-1 β 均會迅速地明顯的表現 (83)。
1-3-9. 一氧化氮合成酶 ( Nitric Oxide Synthase, NOS )
一氧化氮為生理代謝 L-arginine 為 L-citrulline 時所產生的附加 產物。而催化這個反應所需要的酵素之唯一氧化氮合成酶 ( Nitric oxide synthase, NOS )。現在已知的 NOS 共有三種異構形態,分別為 組成性的 neuronal NOS ( nNOS ) 和 endothelial NOS ( eNOS ) 及誘發 性的inducible NOS ( iNOS ) 兩大類 (84)。
這兩大類的酵素調節機制和特性各不相同,組成性的 NOS 在 生理方面扮演維持一般生理功能的角色,合成時需要鈣離子和調鈣 蛋白 ( calmodulin ) 的存在 (85),且生成一氧化蛋的量很少。組成性的
NOS在作用時不會受到皮質固醇的影響,且受激發後生成一氧化氮 的時間較快,作用維持時間相對也較短。誘發性 NOS ( iNOS ) 蛋白 質分子量大約 130 KDa ,已知巨噬細胞、內皮細胞在 TNF-α、LPS 及 INF-γ 刺激時,均可誘發出 iNOS 基因表現進而使細胞合成大量的 NO。在細胞表現 iNOS 時,NO 產生量的變化時常與 iNOS mRNA 量的改變相似,表示調控 iNOS 的表現,大部分是位於轉錄過程 ( transcription )。
1-4. 研究動機
長期處於高血糖的糖尿病患者常伴隨著許多併發症和感染性問 題,現今糖尿病也慢慢被歸類唯一種慢性發炎的症狀。過去的許多 文獻中也提及到在高糖的環境下,會促使內皮細胞、巨噬細胞、白 血球本身的一些活化現象,進而產生 interleukins 或 cytokines,例 如: IL-1β、IL-6 和 TNF-α 。而 NO 的釋放則是賦予巨噬細胞作為 毒殺外來物的手段之一。在文獻中大多利用外加葡萄糖液的方式來 培養細胞,或是以糖尿病患者體內的發炎細胞(例如:血管內皮細 胞、腎絲球、上皮細胞等)去觀察研究細胞受刺激後,所產生的 interleukins 或 cytokines 分泌情形,然而對於巨噬細胞類似的研究文 獻卻是極少。
本實驗之主要目的是想藉由體外的模式去探討經由高濃度葡萄 糖培養於不同的時間,對於巨噬細胞 RAW264.7 的活性影響,以及 探討其所產生的 NO 與 iNOS 蛋白質及基因表現關係。已知巨噬細 胞經由 LPS 活化後會釋出 TNF-α、IL-1 β 及 IL-6 等促發炎的細胞激 素,而這些細胞激素亦可作為一種發炎性的指標。因此,本研究的
另一目的是想藉由模擬糖尿病之高糖環境的體外試驗模式,去探討 高濃度葡萄糖對於巨噬細胞在釋放這些細胞激素能力的影響。
第二章 材料與方法
2-1. RAW 264.7 巨噬細胞之培養 ( RAW 264.7 cell culture )
化學製劑、儀器設備:
1. 細胞株 RAW 264.7(巨噬細胞,BCRC 60001):由食品工業研究 發展所 ( Food Industry Research and Development Institute ) 購得。
2. FBS( HyClone, SH30071-03 胎牛血清)
3. Non-essential amino acid ( Biosera, BIOK0293 ):購自莫比爾 ,5 mL/ 15 mL 離心管分裝並凍至 – 20 °C。
4. Sodium pyruvate 100 mM ( Gibical ) :購自莫比爾,5 mL/ 15 mL 離心管分裝 並凍至 – 20 °C。
5. Penicillin ( 100 U/ mL ) / streptomycin:購自伯昂 ( Gibco, 15140- 122 ) ,5 mL/ 15 mL 離心管分裝 並凍至 – 20 °C。
6. Hepes buffer ( 1 M ) 配製:Hepes powder 購自友和 ( JT Baker-4153- 01 ) , 取 Hepes powder 59.58 g 溶於 150 mL Mini Q 中,調至 pH 7.2,之後再用 Mini Q 補足至 500 mL,滅菌爐滅菌後並放置 4
°C。
7. ( 5 L ) DMEM 基礎培養基配製 : DMEM powder 購自莫比爾 ( 12800-017-1 ) , 將 DMEM powder 倒入 4000 mL 的 Mini Q 中溶 解均勻,取 18.5 g NaHCO3 + Mini Q 溶解均勻,再將二者混合並 補足體積至 5000 mL 於無菌操作台中過濾分裝 (500 mL 一 瓶)。
* DMEM 培養基配製:10 % 胎牛血清 ( FBS ) + 1 % non-essential amino acid + 1 M Hepes buffer + 1mM sodium pyruvate + penicillin
( 100 U/ mL ) / streptomycin( 100 μg/ mL ) + DMEM solution(500 mL 一瓶)。
8. 酒精 ( 70 % ) 9. 無菌操作台
10. 水浴槽(設定: 37 °C)
11. T25 Flask 12. 培養箱
實驗方法:
將所購買的細胞株 RAW 264.7,第一時間迅速放入水浴槽中回 溫後,再以 ( 70 % ) 的酒精噴灑、擦拭管壁及管口並移入無菌操作 台內,並將冷凍管中的細胞液全數吸出並均勻注入已事先回溫的 DMEM 培養基中並培養在 T25 Flask,Flask 上面標上日期,代數之 後並放置培養箱中培養( 培養條件:37 °C,5 % CO2)。
2-1-1. RAW 264.7 巨噬細胞之繼代培養 ( RAW 264.7 cell subculture )
化學製劑、儀器設備:
1. DMEM 培養基 2. 酒精 ( 70 % )
3. T75 Flask:購自莫比爾代理商。
4. 15 mL 離心管:購自莫比爾代理商。
5. 無菌操作台 6. 培養箱
實驗方法:
將滿盤的細胞,更換新鮮 DMEM 培養基後,拍打至細胞懸浮於 培養基中,再全數吸出到 15 mL 離心管裡,上下抽吸約 20 次至均勻 後,注入已先回溫的 DMEM 培養基中培養,Flask上面標上日期,
代數之後並放置到培養箱中培養( 培養條件:37 °C,5 % CO2)。
2-1-2. RAW 264.7 巨噬細胞之冷凍 ( RAW 264.7 cell froze)
化學製劑、儀器設備:
1. DMEM 培養基
2. DMSO ( BIOCHROM KG ) 3. 酒精 (70 %)
4. Trypan blue ( BIOCHROM KG ) 5. 15 mL 離心管
6. 培養箱 1.5 mL 冷凍管
7. 1.5 mL 微量離心管 ( eppendorff ) 8. 倒立式顯微鏡
9. 血球計數器 ( chamber ) :含蓋玻片。
10. 無菌操作台
實驗方法:
將滿盤的細胞 ,更換新鮮 DMEM 培養基後,拍打至細胞懸浮於 培養基中, 再全數吸出到 15 mL 離心管中,上下抽吸約 20 次混合 均勻後,從 15 mL 離心管中取 50 μL 細胞液與等量的 Trypan blue 50 μL ( 1 : 1 ) 在微量離心管中混合均勻,注入至血球計數器 ( chamber )
上的凹槽處 (上下兩個凹槽都要注入) 並蓋上蓋玻片後,於倒立式 顯微鏡下觀看計數。 計算後細胞以 2 × 10 5/ mL(含 10 % DMSO)
注入到 1.5 mL 冷凍管中並放置到 – 20 °C,次日再以乾冰移置 – 80
°C 存放。
2-1-3. RAW 264.7 巨噬細胞之解凍 ( RAW 264.7 cell unfreeze )
化學製劑、儀器設備:
1. DMEM 培養基 2. 酒精 (70 %) 3. T25 Flask 4. 15 mL 離心管 5. 無菌操作台 6. 培養箱
7. 水浴槽(設定: 37 °C)
實驗方法:
從 – 80 °C 拿出的細胞液,第一時間迅速放入水浴槽中回溫後,
再以 ( 70 % ) 的酒精噴灑,擦拭管壁及管口並移入無菌操作台內,
將冷凍管中的細胞液全數吸出並均勻注入已事先回溫的 DMEM 培養 基中培養,Flask上面標上日期,代數之後並放置培養箱中培養( 培 養條件:37 °C,5 % CO2)。
2-2. 15 mM 高葡萄糖 DMEM 培養基培養 RAW 264.7 巨噬細胞
化學製劑、儀器設備:
1. 葡萄糖 ( 150 mM ) 溶液配製 :取 1.35 g 葡萄糖粉末溶 50 mL Mini Q 均勻溶解後過濾備用。
* 使用時,取 4 mL 葡萄糖液與正常 DMEM 培養基 46 mL 混合均勻 後 ( 15 mM ),再以每一個 T 75 flask/ 10 mL 培養細胞。
2. DMEM 培養基 3. 培養箱
4. 50 mL 離心管
實驗方法:
將計算後配置好的 glucose solution ( 150 mM ) 與 DMEM 培養基 混合,更換到 T75 Flask 裡並放置培養箱中進行培養( 培養條件:
37 °C,5 % CO2)。RAW 264.7 須每天更換新鮮 含 15 mM 高葡萄糖 DMEM 培養基,分別培養 7 天,14 天 。
2-3. 細胞計數/ 種植細胞 ( Cell Count/ Cell grow in the 96 well plate )
化學製劑、儀器設備:
1. Trypan blue 2. 酒精 ( 70 % ) 3. 白槽(滅菌過)
4. 八爪 pipette ( 300 μL )
5. 血球計數器 ( chamber ) :含蓋玻片。
6. 倒立式顯微鏡
7. 1.5 mL 微量離心管 ( eppendorff )
實驗方法:
以 DMEM 培養基培養的 RAW 264.7 細胞,更換新鮮 DMEM 培 養基後拍打至細胞懸浮,再用無菌更換型 pipette 全數抽出至 50 mL 離心管中並上下抽吸約 20 次混合均勻,細胞液與 Trypan Blue ( 1 : 1) 混合後,注入至血球計數器 ( chamber ) 上的凹槽處 (上下兩個凹 槽都要注入)並蓋上蓋玻片後,於倒立式顯微鏡下觀看計數(細胞 計算方法:計數血球計數器上下兩處的各兩個大方格數的細胞量;
細胞計數公式:(將上下兩處的細胞量相加總和)/ 2 × 未知數 ( X )
× 2 × 10 4 ﹦總培養基的體積量(含細胞數 × 2 × 10 5)。
計數及計算後,取細胞液與新鮮 DMEM 培養基混合均勻 ,再以 八爪 pipette(調至 100 μL)均勻的將細胞種至 96 well plate 中(2.0
× 10 4 / well ) , 並 放 置 培 養 箱 培 養 ( 培 養 條 件 : 37 °C,5 % CO2)。隔天更換 15 mM 高葡萄糖 DMEM 培養基 1 天(仿急性高 糖環境)。
15 mM DMEM 培養基培養的 RAW 264.7 細胞,分別培養 6 天、
13 天後, 更換新鮮 15 mM DMEM 培養基後拍打至細胞懸浮,再用 無菌更換型 pipette 全數抽出至 50 mL 離心管中並上下抽吸約 20 次 混合均勻,細胞液與 Trypan Blue ( 1 : 1 ) 混合後,注入至血球計數 器 ( chamber ) 上的凹槽處 (上下兩個凹槽都要注入)並蓋上蓋玻片 後,於倒立式顯微鏡下觀看計數。均勻種植在 96 well plate 中,並 放置培養箱培養( 培養條件:37 °C,5 % CO2)。隔天更換 15 mM 高葡萄糖 DMEM 培養基至隔天 (仿慢性 7 天、 14 天高糖環境)。
2-4. 亞硝酸鹽產物測定 ( NO Production test )
化學製劑、儀器設備:
1. LPS ( LipoPolySaccharide ) Stock ( Fluka, FL-62326 ):購自友和 , 1 mg/ mL 分裝並凍至 – 20 °C。
* LPS stock 裡取所需的 μL 數,經 10 X 稀釋後,以 1 μL/ well 的體 與 99 μL/ well 的 DMEM 培養基/ 15 mM 高葡萄糖 DMEM 培養基 混合,並注入至 96 well plate ( 100 μL/ well ) 中進行培養。
2. Griess reagent ( Sigma, SI-G4410 ) :購自友和。
3. DMEM 培養基/ 15 mM 高葡萄糖 DMEM 培養基 4. 培養箱
5. 白槽(滅菌過)
6. 八爪 pipette ( 300 μL )
7. 96well plate ( coating/ un-coating )
實驗方法:
將培養 1 天 ( 24 hr )、7 天、或 14 天高葡萄糖 15 mM DMEM 培 養基的 RAW 264.7 加入含 LPS ( 1 mg/ mL ) 新鮮 15 mM 高葡萄糖 DMEM 培 養 基 後 , 放 置 培 養 箱 培 養 ( 培 養 條 件 : 37 °C , 5 % CO2)。培養於 6 hr, 12 hr, 18 hr, 24 hr 分別收集其上層液,加入等量 Griess reagent 反應 10 min 之後,用 ELISA reader 550 nm 測其 NO 值。
2-5. TNF-α 濃度測定 ( TNF-α assay ) 化學製劑、儀器設備:
TNF-α assay kits ( CytoLab ELISA, 900K54 ):
1. Capture Antibody:分裝 500 μL/ 1.5 mL 微量離心管 並凍至 – 20 °C。
* 使用時以 PBS ( 1 : 20 ) 稀釋,並以 100 μL/ well 加入至 96 well plate 即可。
2. Standard stock:3 ng/ ml
3. 1X-PBS 配製:KH2PO4, 0.245 g + Na2HPO4*2H2O, 0.57 g +
NaCl, 4.25 g 溶於 Mini Q, 400 mL,調至 pH 值 7.2 後再補足 Mini Q 500 mL。
4. Washing buffer 配製:Tween 20 ( 2 % )+ 1X- PBS, 500 mL。使用時 以 300 μL/ well 做清洗。
5. Block buffer 配製:BSA powder 0.4 g 溶於 PBS 40 mL 後,用 0.2 孔徑的 filtre 過濾並放置 4 °C。
6. Diluent solution 配製:BSA powder 0.5 g 溶於 PBST(即 Washing buffer)50 mL 後,用 0.2 孔徑的 filtre 過濾並放置 4 °C。
7. ABTs Liquid substrate:購自友和 ( SI-A3219 ) 。
8. Detection solution stock:使用時以 Diluent ( 1 : 20 ) 稀釋,並以 100 μL/ well 加入至 96 well plate 即可。
9. Conjugate solution:使用時以 Diluent 稀釋 ( 1 : 2000 ) 並以 100 μL/
well 加入至 96 well plate 即可。
10. LPS
11. 96 well plate ( un-coating ) 12. 八爪 pipette ( 300 μL )
13. ELISA reader:設定 450 nm/ 630 nm(雙波長)
實驗方法:
Capture Ab ( 100 μL/ well )
Over-night at room temperature
Wash 4 times ( 300 μL/ well )
Dry and add Block buffer ( 300 μL /well ) 1 hr
Wash 4 times ( 300 μL/ well ) 1 hr
Series Standard (100 μL/ well) / samples ( 100 μL/ well) 2 hr
Wash 4 times ( 300 μL/ well )
Detection solution ( 100 μL/ well ) 2 hr
Wash 4 times ( 300 μL/ well )
Conjugate solution ( 100 μL/ well )
30 min
Wash 4 times ( 300 μL/ well )
ABTs Liquid substrate / ELISA reader ( 450 nm, 630 nm )
2-6. IL-1 β 濃度測定 ( IL-1 β assay)
化學製劑、儀器設備:
IL-1 β assay kit ( Biosource, CMC0813 ) :
1. Coat Antibody ( IL-1 β, 0.125 mg/ 0.125 mL ):分裝 12.5 μL/
eppendorff 1.5 mL(微量離心管)並凍至 – 20 °C。
* 使用時以 coating buffer A ( 1 : 800 ) 稀釋,並以 100 μL/ well 加入 至 96 well plate 即可。
2. Dection antibody ( IL-1 β, 0.025 mg/ 0.125 mL )
* 使用時以 assay buffer (含 5 % 血清) 稀釋,並以 100 μL/ well 加 入至 96 well plate 即可。
3. Standard IL-1 β: 放至 – 4 °C。
* 使用時以 ( 1 : 10 ) 稀釋至 1000 pg/ mL,並以 300 μL/ well 加入至 96 well plate 即可。
4. Streptavidin-HRP ( 0.025 mg/ 0.125 mL )
* 使用時以 assay buffer 稀釋至 0.05 μg/ mL,並以 100 μL/ well 加入 至 96 well plate 即可。
5. Coating buffer A ( Biosurce, CB07100 ) 配製:NaCl, 8 g + NaHPO4, 11.3 g + KH2PO4, 0.2 g + KCl 0.2 g 溶於 1 L Mini Q,調 pH 值至 7.4。
6. Assay buffer ( Biosurce, DS98200 ) 配製:NaCl, 8 g + NaHPO4, 11.3 g+ KH2PO4, 0.2 g + KCl, 0.2 g + Bovine serum albumin ( sigma, 9048-46-8 ), 5 g + Tween 20, 1 mL 溶於 1 L Mini Q,調 pH 值至 7.4。
7. Wash buffer ( Biosurce, WB01 ) 配製:NaCl, 9 g + Tween 20, 1 mL 溶於 1 L Mini Q 調 pH 值至 7.4。
8. Subdtrate solution ( Biosurce, SB01 ) :Tetramethylbenzidine ( TMB ) 和 Hydrogen Peroxide。
9. Stop solution ( BioSurce, SS01100 ):H2SO4 ( 1.8 N ) 10. 96 well plate ( un-coating )
11. 八爪 pipette ( 300 μL )
12. ELISA reader:設定 450 nm
實驗方法:
Coating Ab IL-1 β ( 100 μL/ well ) Over-night at 4 °C
Wash 4 times ( 300 μL/ well )
Block with assay buffer ( 300 μL /well ) 1 hr
Wash 4 times ( 300 μL/ well ) 1 hr
Series Standard (100 μL/ well) / samples or controls ( 100 μL/ well) Immediately
Detection antibody IL-1 β solution ( 50 μL/ well )
Gently tap the plate on the side 10 times to mix 2 hr at room temperture Wash 5 times ( 300 μL/ well )
TMB substrate ( 100 μL/ well )
30 min in the dark at room temperture Stop solution ( 100 μL/ well )
30 min ELISA reader ( 450 nm )
2-7. IL-6 濃度測定 ( IL-6 assay)
化學製劑、儀器設備:
IL-6 assay kit ( Biosource, CMC0063 ) :
1. Coat Antibody ( IL-6, 0.125 mg/ 0.125 mL ):凍至 – 20 °C。
* 使用時以 coating buffer B ( 1 : 800 ) 稀釋,並以 100 μL/ well 加入 至 96 well plate 即可。
2. Dection antibody ( IL-6, 0.05 mg/ mL )
* 使用時以 assay buffer (含 5 % 血清) 稀釋,並以 100 μL/ well 加 入至 96 well plate 即可。
3. Standard IL-6: 放至 – 4 °C。
* 使用時以 ( 1 : 10 ) 稀釋至 1000 pg/ mL,並以 300 μL/ well 加入至 96 well plate 即可。
4. Streptavidin-HRP ( 0.025 mg/ 0.125 mL )
* 使用時以 assay buffer 稀釋至 0.05 μg/ mL,並以 100 μL/ well 加入 至 96 well plate 即可。
5. Coating buffer B ( Biosurce, CB01100 ) 配製:NaHCO3, 4.3 g + NaCO3, 5.3 g 溶於 1 L Mini Q,調 pH 值至 7.4。
6. Assay buffer ( Biosurce, DS98200 ) 配製:NaCl, 8 g + NaHPO4, 11.3 g+ KH2PO4, 0.2 g + KCl, 0.2 g + Bovine serum albumin ( sigma, 9048-46-8 ), 5 g + Tween 20, 1 mL 溶於 1 L Mini Q,調 pH 值至 7.4。
7. Wash buffer ( Biosurce, WB01 ) 配製:NaCl, 9 g + Tween 20, 1 mL 溶於 1 L Mini Q 調 pH 值至 7.4。
8. Subdtrate solution ( Biosurce, SB01 ) :Tetramethylbenzidine ( TMB ) 和 Hydrogen Peroxide。
9. Stop solution ( BioSurce, SS01100 ):H2SO4 ( 1.8 N ) 10. 96 well plate ( un-coating )
11. 八爪 pipette ( 300 μL )
12. ELISA reader:設定 450 nm
實驗方法:
Coating Ab IL-6 ( 100 μL/ well ) Over-night at 4 °C
Wash 4 times ( 300 μL/ well )
Block with assay buffer ( 300 μL /well ) 1 hr
Wash 4 times ( 300 μL/ well ) 1 hr
Series Standard (100 μL/ well) / samples or controls ( 100 μL/ well) Immediately
Detection antibody IL-6 solution ( 50 μL/ well )
Gently tap the plate on the side 10 times to mix 2 hr at room temperture Wash 5 times ( 300 μL/ well )
TMB substrate ( 100 μL/ well )
30 min in the dark at room temperture Stop solution ( 100 μL/ well )
30 min ELISA reader ( 450 nm )
2-8. 蛋白質抽取 ( Protein Extraction )
化學製劑、儀器設備:
1. 冰塊
2. 液態氮:購自雲山行。
3. 1X- PBS(細胞用)配製:KH2PO4, 0.245 g + Na2HPO4*2H2O, 0.57 g + NaCl, 4.25 g 溶於Mini Q 400 mL,調至 pH 值 7.2,再補足Mini Q至 500 mL。
4. Incubation buffer:40 mM Tris-HCl ( pH 7.4 ) + 1 mM EDTA + 150 mM NaCl。
5. 0.25 M Tris-HCl ( pH 7.8 ) 6. 刮勺/ 酒精 ( 70 % )
7. 微量管離心機(設定: 4 °C)
8. 水浴槽(設定: 37 °C)
9. 無菌操作台 10. – 20 °C 冰箱
11. 1.5 mL 微量離心管 ( eppendorff )
實驗方法:
RAW 264.7 細胞以 1X- PBS 清洗 3 次並於去除所有的 BPS 後,
加入 Incubation buffer 1 mL 在室溫下反應 5 min ,並將細胞刮下到 1.5 mL 微量離心管放置冰上。爾後以 13000~14000 rmp 高速離心 ( 30 sec ),去除上清液,再加入 50 μL 4 °C 的 0.25 M Tris-HCl ( pH 7.8 ),打散底部的細胞團塊再 spin down。細胞以液態氮冷凍 ( 5 min ) 及水浴鍋解凍 ( 5 min ) 的方式, 如此反覆 3 次循環(約 30 min)後 打破細胞膜抽取其蛋白質 ,於 4 °C 下離心 13000 ~ 14000 rpm ( 5 min ) , 並 收 集 其 上 清 液 ( 即 蛋 白 質 液 ) 至 乾 淨 的 微 量 離 心 管 ( eppendorff ),存放於 – 20 °C 備用。
2-9. 蛋白質濃度測定 ( Protein Assay )
化學製劑、儀器設備:
1. Protein assay reagent ( Bio-rad ) 2. ELISA Reader: 波長 595 nm 3. Standard Curve
實驗方法:
將存放於 – 20 °C 蛋白質液 ( Stock ) 先取 3 μL 加入 447 μL Mini Q 混合成 500 μL 蛋白質水溶液 ,再從中取 10 μL 與 200 μL 的 Protein assay reagent ( 1 : 20 ) 混合成蛋白質試劑混合液,並反應 15 min 後,將蛋白質試劑混合液以 ELISA Reader 波長 595 nm 測其吸 光值,所得吸光值代入以血清白蛋白 ( BSA ) 所作成的 Standard Curve ( 0.05 mg/ mL ~ 0.5 mg/ mL ) 來換算其濃度。
2-10. 西方墨點法 ( Western blot )
化學製劑、儀器設備:
1. Tris-base ( Amresco, 0826-1KG ) 2. Glycine ( Amresco, 0167-1KG )
3. ( 1.5M ) Tris-HCl ( pH 8.8 ) 配製:18.171 g Tris-base 溶於 100 mL Mini Q 中,調至 pH 值 8.8。
4. ( 0.5 M ) Tris-HCl ( pH 6.8 ) 配製:9.0855 g Tris-base 溶於 100 mL Mini Q 中,調至 pH 值 6.8。
5. APS solution 配製:APS powder ( J.T.Baker, C04466 ) ,1 g APS powder 溶於 1 mL Mini Q 中混合均勻。
6. 10X- SDS-PAGE running buffer 配製:Tris-base 15 g + Glycine 72 g + SDS ( J.T.Baker, 151-21-3 ) 5 g 溶於 500 μL Mini Q,使用時稀釋 成 1 X。
7. Acrylamide ( Bio-rad, 161-0156 )
* 製膠:Separating Gel ( 10 % ): Mini Q 4.1 mL + 1.5M Tris-HCl ( pH 8.8 ) 2.5 mL+ SDS 10 % 100 μL + Acrylamide 3.3 mL + APS 100 μL + TEMED 4 μL。
Stacking Gel ( 4 % ):Mini Q 3.2 mL + 0.5 M Tris-HCl ( pH 6.8 ) 2.5 mL+ SDS 10 % 100 μL + Acrylamide 1.33 mL + APS 50 μL+
TEMED 10 μL。
8. 4X- Sample buffer loading dye : ( 1 M ) Tris 2 ml + 10 % 2- mercaptoethanol ( Sigma, 200-464-6 ) 1mL+ SDS 0.8 g + Glycerol 5 g + Bromophenoblue 0.04 g。
9. Block buffer 配製:脫脂奶粉 2 g 溶於 1X- PBS 40 mL。
10. TEMED ( Sigma, BP161-0800 ) :購自汎泰。
* Blotting Transfer buffer 配製:Tris-base 3.03 g + Glycine 14.4 g 溶於 Methanol 100 mL + Mini Q 900 mL。
11. Primary Anti-body ( Sigma ):購自友和 ( SI-N9657 ) 。
* Anti mouse-iNOS 配製 :Anti mouse-iNOS 10 μL + 1X-PBS 10 mL ( 1 : 10000 )。
12. Secondary Anti-body
* HRP-Anti-mouse IgG Antibody ( Jackson, 115-035-003-100 ) 配製 : Secondary Anti-body 10 μL + 1X - PBS 10 mL ( 1 : 10000 )。
13. α -Tubulin Anti-body ( Sigma ) 配製:購自友和 ( SI-T5168 ) ,α- Tubulin Anti-body 3 μL + 1X - PBS 12 mL ( 1:40000 )。
14. Washing buffer 配製:Tween 20 ( 2 % ) + 1X- PBS 500 mL。
16. TMB membrane 呈色劑 ( KPL, 50-77-03 ) :購自岑祥。
17. 冰塊
18. 加熱板,振盪板
19. PVDE membrane: 購自伯昂 ( IPVH 00010 ) 。
20. Bio-read 製膠套組:厚片玻璃、薄片玻璃、製膠架、comb、
( Bio-read ) 電泳(槽)套組
21. 轉膜套組:轉膜槽,冰槽,轉模板(黑板/白板),海綿,轉膜 用厚紙片。
實驗方法:
Protein extract + Sample buffer ( 3 : 1 )
Boiling 5 min and kept on ice 5 min SDS PAGE electrophoresis
Transfer electrophoresis
Blocking ( 1 hr ) / Washing 1 time ( 3 min )
Primary Ab ( over-night ) / Washing 2 times ( each 5 min )
Secondary Ab ( 2 hr ) / Washing 3 times ( each 5 min )
Scanned by TME reagent
2-11. RNA 抽取 ( RNA extraction )
化學製劑、儀器設備:
1. 1.5 mL 微量離心管 ( eppendorff )
2. RNA isolation system Kit:購自騰達行代理商 ( GS16780 )(內附:
Lysis buffer,DNA、蛋白質沈澱劑, RNA 溶解劑)。
3. Incubation buffer:4 °C 存放。
4. Isopropanol:4 °C 存放。
5. nuclearase free water ( Promega ) 6. 冰塊
7. 酒精 ( 70 % ) 8. 刮勺
9. 微量離心機 10. 分光光度計
實驗方法:
將所培養的細胞 ( 6 well plate ),去除培養基並加入 500 μL 的 Incubation buffer,刮下細胞至微量離心管 ( eppendorff ) ,以 1400 ~ 1600 rpm 10 min,去除上清液,加入 300 μL Lysis buffer 並 vortex 30 sec ,再以 13000 rpm 30 sec 後,取其上清液 300 μL 至另一 eppendorff,再加入 100 μL DNA / protein 沈澱劑,輕柔翻轉 20 次放 置冰上 5 min ,再次離心 ( 13000 rpm 8 min ),取上清液至新的微量 離心管( eppendorff ) 再以相同的條件,離心 8 min。將上清液取至新 的微量離心管 ( eppendorff ) 後加入 300 μL Isopropanol 並輕柔翻轉 60 次,以 13000 rpm 8 min,所得之沈澱物為 total RNA (呈半透明 狀)。去除上清液,加入酒精 ( 70 % ) 清洗並輕柔翻轉 10 次,以 13000 rpm 5 min 後,去除上清液,spin down 去除剩餘殘液,放置無 菌操作台裡風乾(約 15 min)。此時可看見微量離心管底部的沈澱 物由白色轉為透明,此時再加入 30 μL RNA 溶解劑並放置冰上 30 min 後。輕柔混合均勻後,以分光光度計 260 nm / 280 nm 測其 RNA 濃度。測 RNA 濃度時,先將抽取後的 RNA ( 20 μL ) 中取 1 μL +
999 μL nuclearase free water ( 1 : 1000 ),並注入石英管中放入分光光 度計機裡,測其濃度。
2-12. 反 轉 錄 - 聚 合 酶 鏈 反 應 ( Reverse transcription-Polymerase Chain Reaction )
化學製劑、儀器設備:
1. Random primer, 5X - RT buffer, 5 mM dNTP, RNase ingibitor, M- MLV-RT enzyme ( Promega ):購自勁因 ( RA1250 )(– 20 °C 存 放)。
2. 10X - PCR buffer, 5 mM dNTP, DMSO, primer ( F ), primer ( R ) , Taq ( Promega ):購自勁因 ( RA1250 ) 。
3. 分光光度計(設波長為 260 nm, 280 nm)
4. 冰塊 5. 200 μL PCR 反應管
6. Nuclearase free water ( Promega )(滅菌過)
7. PCR 反應器
實驗方法:
5X-RT buffer 4 μL + 5 mM dNTP 2 μL + RNase inhibitor 0.5 μL + RNA ( 2 μg / X μL ) + 1 μL random primer 混合後再加入 M-MLV-RT 酵素 1 μL,最後再用滅菌後的 nuclearase free water 補足體積至 20 μL。
mRNA + random primer + M-MLV-RT
1 μL cDNA + 9 μL nuclearase free
42°C, 60 min,95°C, 5 min ,4°C ∝
10X-PCR buffer 5 μL + 5 mM dNTP 2 μL + DMSO 1 μL + primer ( F ) 1.5 μL + primer ( R ) 1.5 μL + Taq 0.25 μL ( add nuclearase free
water 50 μL )
94°C, 45 sec ,65°C, 45 sec ,72°C, 2 min ,15 times cycles
94°C, 45 sec ,67°C, 45 sec ,72°C, 2 min ,10 times cycles ,4°C ∝
cDNA storage at – 20 °C
2-13. DNA 瓊膠製備/ DNA 電泳 ( DNA Agarose gel preparing / DNA electrophoresis )
化學製劑、儀器設備:
1. SYBR Safe DNA gel stain ( Invitrogen USA, 44747A ) 2. Agarose powder ( Invitrogen, 15510019 ) :購自萊富。
3. 10X - TAE ( Tris-Borate EDTA ) buffer 4. 0.5X - TAE buffer
5. 6X-loading dye 6. 微波爐
7. DNA 電泳套組
8. DNA Marker ( Biotech ) :購自友和。
9. Agarose 製膠板模組
實驗方法:
DNA Agrose gel ( 1 % ) 配製:0.4 g Agrose,溶於 1X-TAE buffer 40 mL 中並加入 4 μL DNA gel stain 混合均勻, 放入微波爐中微波 後,倒入模組中,放置室溫下冷卻。
DNA 電泳:將 Samples 從 – 20 °C 拿出,以 1 : 5 與 6X - loading dye 做混合並注入 DNA Agarose gel 中,放入 DNA 電泳槽,倒入 0.5X - TAE buffer 開始進行 DNA 電泳分析(由負極向正極泳動),
約 20 min 後即可將 Agarose gel 用 UV 拍照存檔。
2-14. 統計分析
本實驗對於細胞分析中,各組數據均以平均值加減一個正負標 準差 ( mean ± SD ) 表示,並以 Student's t-test 作統計分析,( p) 值以
* p< 0.05,** p< 0.01 和 *** p< 0.001 表示顯著性差異。圖表以 Prism4.Document 和 Sigma Plot 9.0 繪製。
第三章、結果
3-1. NO 產量試驗
正常 DMEM 培養基的 RAW 264.7 巨噬細胞所釋放在 medium 裡 的 NO 產 量 ( basal level ) 並 不 會 隨 著 時 間 而 有 所 改 變 ( 圖 1.a.) 。反之,NO 產量在有添加 15 mM glucose DMEM 培養基的 RAW 264.7 培養 18 hr 後,與正常 DMEM 培養基的 RAW 264.7 相 比,則開始有明顯增加的傾向 ( p < 0.01 )。
正常 DMEM 培養基所培養的 RAW 264.7 在 LPS 刺激之下,
NO 產量隨培養時間延長而增加。這樣的情形同樣也在添加 15 mM glucose DMEM 培養基的 RAW 264.7 觀察到,只是上升的濃度在每 個時間點,所測得的 NO 值都比前者為低 ( p < 0.01 )(圖 1.b. )。
隨著 15 mM glucose DMEM 培養基的 RAW 264.7 培養時間延長 至 7 天(中慢性),發現 6 hr 後所測得的 NO basal 值,與正常 DMEM 培養基的 RAW 264.7 相比,在統計學上卻有顯著差異 ( p <
0.01 )(圖 1.c.)。同樣的,在 LPS 刺激下,有添加 15 mM glucose DMEM 培養基的 RAW 264.7 從 6 hr 到 24 hr,可以看到所釋放出的 NO 值也有著明顯偏低的情形 ( p < 0.01 ),與正常 DMEM 培養基的 RAW 264.7 相比(圖 1.d.)。
將添加 15 mM glucose DMEM 培養基時間延長至 14 天(慢 性),從 medium 中所測得的 NO basal 值,在各時間點與正常 DMEM 培養基的 RAW 264.7 相比,有慢慢回升的情形 ( p < 0.01 )
(圖 1.e)。經由 LPS 刺激後,同樣的發現在長期受 15 mM glucose DMEM 培養基的 RAW 264.7 ,在時間點 24 hr 時,所測得的 NO 釋
放濃度值,與正常 DMEM 培養基的 RAW 264.7 相比,有明顯下降 的趨勢 ( p < 0.01 )(圖 1.f)。
3-2. iNOS protein assay by Western blot
已知細胞所釋放出來的 NO 與合成它的 iNOS 蛋白質有直接的 相關性。在無添加 LPS 刺激之下,正常 DMEM 培養基與添加 15 mM glucose DMEM 培養基,所培養(1 天,7 天,14 天)的 RAW 264.7 並沒有明顯觀察到 iNOS 蛋白質(分子量 130 KDa)的表現
(圖 2.a.)。
經由 LPS 所誘發出來的 iNOS 蛋白質表現中,15 mM glucose DMEM 培養基,培養 1 天(急性)和 7 天(中慢性)的 RAW 264.7 之 iNOS 蛋白質表現比正常 DMEM 培養基的 iNOS 表現來的少量,
其結果與 NO production test 1 天、7 天的結果相符。但在 14 天(慢 性)15 mM glucose DMEM 培養基之 RAW 264.7 的 iNOS 蛋白質濃 度(分子量 130 KDa)表現卻出乎意料地,有相對明顯升高的情形
(圖 2.b.)。
3-3. iNOS mRNA expression by RT-PCR
RAW 264.7 由 15 mM glucose DMEM 培養 1 天,7 天,14 天 後,實驗結果中發現隨著 15 mM glucose DMEM 培養基培養的時間 越久,basal iNOS mRNA 的濃度有慢慢增加的趨勢,而至 14 天(慢 性)後卻有一個消失的現象。而經 LPS 刺激的 iNOS mRNA 濃度,
在 15 mM glucose DMEM 培養 1 天(急性)後,有上升的情形。隨 著 15mM glucose DMEM 培養 7 天(中慢性)的 iNOS mRNA 濃度
卻明顯轉淡至消失的情況,在 14 天(慢性) 15mM glucose DMEM 培養後的 iNOS mRNA 濃度(分子量 607 bp)又有著明顯倍增的現 象(圖 3.a)。
3-4. TNF-α 產量試驗
TNF-α 為細胞發炎時所釋放出的一種發炎激素,不論是 basal 或是 LPS 刺激的 TNF-α 值會隨時間延長而同步增加,所以實驗中的 time courses 設為 0.5 hr、1 hr、1.5 hr ,因為這是 basal 值及 LPS 刺 激後,差異最大的時間點範圍。
在 basal TNF-α 濃度測定方面,培養含 15 mM glucose DMEM 培養基的 RAW 264.7 巨噬細胞 1 天(急性),測得的 TNF-α 濃度值 在 1.5 hr,有明顯上升的情形。經 15 mM glucose DMEM 培養基培 養 7 天(中慢性)後的 RAW 264.7 巨噬細胞,隨著時間的增加在 1 hr 及 1.5 hr 時有明顯的減少的情形。而長達 14 天(慢性)15 mM glucose DMEM 培養基培養後的 RAW 264.7 巨噬細胞,測得的 TNF- α 與正常 DMEM 培養基的 RAW 264.7 相比,則有大量提升的現象
(圖 4.a.)。
添加 LPS 刺激後,在急性 ( acute ) 與 慢性 ( chronic ) 15 mM glucose DMEM 培養基培養後的 RAW 264.7 巨噬細胞,所測得的 TNF-α 濃度,與正常 DMEM 培養基的 RAW 264.7 巨噬細胞之間並 無明顯變化。反之,中慢性 ( sub-chronic ) 15 mM glucose DMEM 培 養基培養後的 RAW 264.7 巨噬細胞,與正常 DMEM 培養基的 RAW 264.7 巨噬細胞相比,卻有一個明顯抑制的情形(圖 4. b.)。
3-5. IL-6 產量試驗
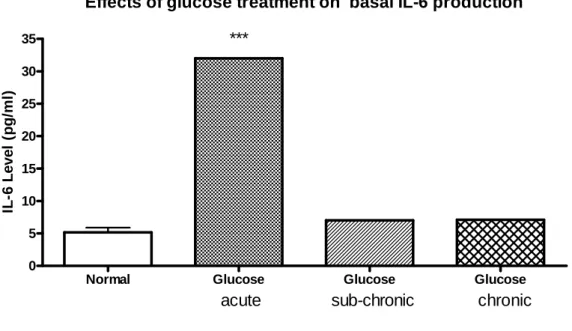
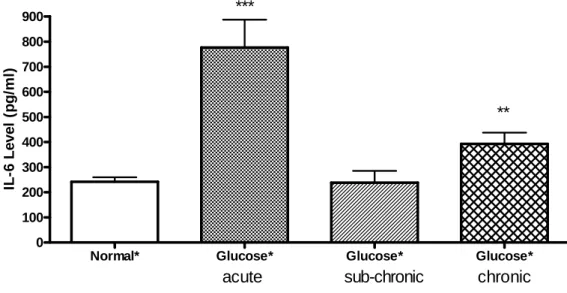
IL-6 為人體內調節免疫及發炎反應的一種細胞激素。檢測 IL-6 釋放量時,因半衰期非常短,所以以 3 小時作為一個檢測的時間 點。在 basal 1 天(急性)含 15 mM glucose DMEM 培養基所培養的 RAW 264.7 ,有略微高於正常 DMEM 培養 RAW 264.7 的情況。經 由 LPS 刺激後 15 mM glucose group,與正常 DMEM 培養基的 RAW 264.7 相比,有明顯增加的趨勢 ( p < 0.001 )(圖 5.a)。
在培養 7 天(中慢性)含 15 mM glucose DMEM 培養基所培養 的 RAW 264.7 經 LPS 添加後,所釋放出的 IL-6 濃度,觀察到含 15 mM glucose 與正常 DMEM 所培養的 RAW 264.7 比較,並無差異性
(圖 5.b)。
將培養時間延長到 14 天(慢性)15 mM glucose DMEM 培養後 發現,LPS 刺激後的 15 mM glucose group 之 IL-6 釋放量,與正常 DMEM 培養基的 RAW 264.7 相比,有明顯回升的現象 ( p < 0.01 )
(圖 5.c),但 14 天 15 mM glucose group 的 IL-6 釋放量,仍沒有比 1 天(急性)15 mM glucose group 多。
3-6. IL-1 β 產量試驗
IL-1 β 為受發炎物質刺激後,所分泌出的一種細胞激素。檢測 IL-1 β 釋放量時,因半衰期非常短,所以以 3 小時作為一個檢測的 時間點。分別在培養於 15 mM glucose DMEM 培養基之 RAW 264. 7 於 1 天(急性),7 天(中慢性),14 天(慢性)後, 在 basal IL-1 β 的表現量,與正常 DMEM 培養基的 RAW 264.7 相比,皆有明顯 增加的趨勢。( p < 0.001 ) (圖 6.a)。
正常 DMEM 培養基與 15 mM glucose DMEM 培養基培養之 RAW 264.7 於 1 天,7 天,14 天後添加 LPS 刺激,結果發現 15 mM glucose DMEM 培養 1 天(急性),與其 basal 1 天(急性)和正常 DMEM 培養基的 RAW 264.7 所釋放出的 IL-1 β 表現量相比,並沒 有太大的變化 ( p < 0.001 ) 。而 7 天(中慢性)IL-1 β 的表現量,則 有略微抑制的情形。反之,14 天(慢性)與正常 DMEM 培養基的 RAW 264.7 相比,卻又有明顯回升增加的現象 ( p < 0.001 )(圖 6.b)。
第四章、討論
高血糖的併發症是一種與發炎相關的疾病 (86)。 在過去的文獻中 指出 NO 的產生對生物體是具有保護作用,因為它參與了生物的急 性免疫發炎反應。低濃度的 NO 可以對抗外來的病源體 (87),但 NO 對細胞的傷害是無選擇性的 (88)。所以若當 NO 濃度過高,則會對本 體的細胞產生毒性,進而導致細胞突變,甚至死亡。可能原因是由 於大量的 NO 抑制了 DNA 的合成,進而對粒腺體造成了傷害,使細 胞膜失去完整性,最後走向細胞凋亡 ( apoptosis ) (89)。在局部或全身 的發炎反應中,例如:糖尿病、動脈硬化症等,也會產生高濃度的
NO (90),這也說明了高濃度葡萄糖與細胞產生 NO 釋放量和其機制有
著正相關性。
曾有文獻提及當細胞在正常濃度的葡萄糖 ( 5.5 mM ) 與高濃度 葡萄糖 ( 25 mM ) 所培養的環境中,隨著葡萄糖濃度越高,培養的時 間越久,其 NO 釋放量則有明顯上升的現象 (91)。然而在 LPS 急性的 誘發之下,巨噬細胞所產生 NO 及 iNOS 蛋白質濃度也都有著上升 的表現,但隨著高葡萄糖培養 48 hr 過後的巨噬細胞,其釋放細胞激 素的能力有著降低的情形 (91) 。亦曾有其他相關研究指出,在高濃度 葡萄糖培養下的巨噬細胞 ( J774 macrophage cell line ),經 LPS 刺激 後, NO 釋放量有減少的情況 (92)。這與本文對 NO 產生濃度測試,
所得結果有雷同。在 NO production test 的圖中 (圖1.b),我們證 實了在添加 LPS 的刺激下,高濃度葡萄糖培養 1 天的 RAW 264.7 與 正常 DMEM 培養基 ( Normal ) 的 NO 釋放濃度相比,並沒有太大的 差異性。但隨著培養時間的延長至 14 天,其 NO 釋放的濃度有比
Normal 還要低的情形 (圖1.d, 圖1.f)。
有研究報導顯示,飼養帶有糖尿病之大鼠 4 至 8 週後,未經任 何刺激之情況下,取其腎絲球細胞,經 Western bolt 後並未偵測到任 何的 iNOS protein 表現 (93)。此外有報導曾指出將血管平滑肌細胞培 養在高葡萄糖的狀態下 ( 72 hr ) 並以 IL-1 β 刺激 ( 24 hr ) 後,活化 protein Kinase C β II isoform,發現 iNOS 及 NO prodcution 皆有明顯 的表現量產生 (94)。相同地在我們的 Western blot test 研究結果中顯 示:隨著高葡萄糖濃度培養的時間越久,經由 LPS 刺激後 RAW 264.7 所產生出的 iNOS protein 表現有先下降(1 天)再上升(7 天 及 14 天)的傾向(圖2.b)。
RT-PCR test ( 圖 3.a )方面,在 basal 條件下經葡萄糖培養 RAW 264.7 後的 iNOS gene 表現有比 Normal 明顯的表現情形,14 天的結果除外。這可能的解釋是因長期葡萄糖培養細胞後,影響細 胞內部的一些途徑改變(例如:NF-kappaB binding activity 的 down- regulation),與抑制了 iNOS 轉錄過程 ( transcription ) 和其本身的活 性能力 (95)。另外在葡萄糖 1 天(急性)培養後,添加 LPS 刺激所誘 發 iNOS gene 表現有明顯增加。隨著高葡萄糖培養的時間延長至 7 天 iNOS gene 濃度卻有一個抑制的表現,這與其他相關文獻所得結
果相似 (93) (95)。然而我們的結果發現 14 天後 iNOS gene 的表現量又
有一個明顯大量表現的情況。可能的原因是巨噬細胞的 iNOS gene 隨著長期高葡萄糖培養下,短暫抑制了其轉錄的表現 然而在培養 14 天後恢復其轉錄 ( translation ) 機轉的運作,進而大量表現出 iNOS 蛋白質,與 Nakai 等人在 2004 年的研究指出人類角質細胞在長時間 高濃度葡萄糖培養下,對高糖環境有著不同的適應性及恢復性 (95)。 當糖尿病患者中的巨噬細胞受外來物,如:LPS,刺激下所產