國立臺東大學生命科學系碩士班 碩士論文
指導教授:彭仁君 博士
食草植物與植被組成對大紅紋鳳蝶 產卵棲地選擇之影響
Influences of foodplants and vegetation composition on oviposition habitat selection of Byasa polyeuctes termessus
研 究 生: 林慧婷 撰
國立臺東大學生命科學系碩士班 碩士論文
食草植物與植被組成對大紅紋鳳蝶 產卵棲地選擇之影響
研 究 生:林慧婷 撰
指導教授:彭仁君 博士
Department of Institute of Life Science National Taitung University
Master Thesis
Influences of foodplants and vegetation composition on oviposition habitat selection of Byasa polyeuctes termessus
By Hui-Ting Lin
Advisor: Jen-Jiun Perng
2 0 1 0
致謝
本以為後山的生活是枯燥且乏味,但其實處處充滿活力與驚奇!而隨著本論 文的完成,學生研究所兩年的生活將畫下完美句點,心中除了感謝還是感謝。
首先,感謝恩師彭仁君教授,帶領學生邁入結合生態與學術研究的殿堂,開 啟學生全新的視野並體驗生命的豐富。從野外調查至論文撰寫的過程中,每當學 生感到茫然或沮喪,彭老師人生經驗的分享及專業知識的引領,往往使學生豁然 開朗,尋得方向。由於彭老師全心全意的關照與傾囊相授,學生才得以有所成長。
其次,特別感謝本系所長李炎博士與屏東科大森林系郭耀綸博士的諄諄指導,諸 多建議使本論文更加嚴謹、豐富。
感謝本研究團隊的人碩及孙庭,在研究期間共同分擔繁複的調查工作,彼此 間共同的信任與默契,亦是推動本論文完成的力量。同時感謝俊元協助野外植物 的辨識,及六位專題生,駿宏、俊廷、克瑋、紹昌、笙奎、震鈞接手後續的野調 工作,建立並延續寶貴的生態資料。特別感謝惠嵐、嘉謙、駿宏及俊廷,熱心協 助論文與口試等相關行政與流程事宜。
感謝師母月嬌、妹妹慧婕、好友以婷、正修與各位昔日同窗對我精神上的鼓 勵,不時聽我吐露苦水,釋放苦悶的壓力,支持我達成目標。最後,特別感謝我 摯愛的父親、母親與芳蓮阿姨,在無數熬夜苦思、混亂低潮的日子裡,你們的關 懷是我最大的心靈支柱,並全力給予經濟上的支援,使我在論文研究上心無旁騖。
謹此將我由衷的感謝及本論文獻給所有關心我的師長與朋友,並與各位分享 心中的喜悅與感動。
林慧婷 謹誌 2010 年仲夏 國立臺東大學
食草植物與植被組成對大紅紋鳳蝶 產卵棲地選擇之影響
作 者 : 林 慧 婷
國 立 臺 東 大 學 生 命 科 學 系 碩 士 班
摘 要
為瞭解野外大紅紋鳳蝶 (Byasa polyeuctes termessus Fruhstorfer) 對棲地中食草植物資源之利用表現,並探討微棲地中植被組成對大紅 紋鳳蝶產卵行為與其蝶卵存活造成的影響,本研究自 2009 年 3 月 1 日 至 2010 年 3 月 27 日,每周調查里龍山不同海拔樣帶蜂窩馬兜鈴 (
Aristolochia foveolata Merr.)與掌葉馬兜鈴
(Aristolochia palmata C. L. Yang)兩種食草的分布,計量全年各季大紅紋鳳蝶產卵微棲地中兩種馬兜鈴
食草資源量及植被組成,並記錄其蝶卵在微棲地中的存活狀況。結果
顯示,大紅紋鳳蝶卵蟲數與兩種馬兜鈴食草的數量分布趨向一致,且
產卵微棲地中兩種馬兜鈴食草資源量與非食草植物物種數皆高於未產
卵微棲地。雌蝶不會因食草資源量或植被組成增減改變而產下過多蝶
卵。野外蝶卵被寄生比例高達 50%,且被寄生比例在食草葉片量多的
微棲地要高於在葉片量少的微棲地。不同的植被覆蓋率及非食草植物
物種數並不影響蝶卵的存活狀況。調查結果顯示,微棲地中馬兜鈴食
草的可利用性為大紅紋鳳蝶產卵微棲地選擇的關鍵。此種雌蝶都有在 多種非食草植物中尋找本區兩種馬兜鈴食草的能力,且具有分散產卵 的產卵型態,此除可降低蝶卵在同處被大量寄生的高風險外,亦可確 保個別帅蟲在生長微棲地有較充足的食物來源。
關 鍵 詞 : 大 紅 紋 鳳 蝶 、 食 草 、 植 被 組 成 、 產 卵 棲 地 選 擇 、 微 棲 地
Influences of foodplants and vegetation composition on oviposition habitat selection of
Byasa polyeuctes termessus
Hui-Ting Lin Abstract
In order to understand the influences of foodplant and vegetation composition on oviposition habitat selection and egg survival of Byasa polyeuctes termessus, this study investigated the distribution of two foodplants, Aristolochia foveolata and Aristolochia palmate at different altitudes in Lilung Mountain. From March-1-2009 to March-27-2010, we weekly measured the two foodplant density, foodplant leaf abundance and vegetation composition in oviposition microhabitat of B. polyeuctes termessus, and monitored the survival of each eggs recorded. The results showed that the number of eggs and larvae distributed consistently with the number of both foodplants at different altitudes. The two foodplant resource and the species number of non-foodplants in oviposition microhabitats were significantly higher than that in non-oviposition microhabitats. Female butterfly laid only limited number of eggs on each foodplant, and resulted in a scatter distribution of egg-laying in habitat. In the field, parasitism rates of eggs could reach to 50%, which was higher in microhabitat with more foodplant leaves than that with less leaves.
Vegetation coverage and the number species of non-foodplants did not
significantly influence the survival of the eggs. In conclusion, the foodplant
availability is the key point for oviposition habitat selection of B.
polyeuctes termessus. The female butterfly was able to search the two foodplants within microhabitat growing with many non-foodplants, and the scatter distribution of egg-laying in habitat not only may reduce the risk of egg parasitism but also ensure the foodplant supply for the larval growth in microhabitat.
keyword:Byasa polyeuctes termessus, foodplant, vegetation
composition, oviposition habitat selection, microhabitat
目錄
摘要 ... I Abstract ... III 目錄 ... V 圖目錄 ...VII 照片目錄 ... IX
壹、前言 ...1
一、研究背景 ... 1
二、研究動機與目的 ... 4
貳、材料與方法 ...6
一、研究地點概述 ... 6
二、研究物種 ... 8
三、食草植株的分布及密度對大紅紋鳳蝶產卵選擇之可能影響 ... 10
四、微棲地內食草資源量或植被組成對大紅紋鳳蝶產卵選擇與蝶卵存活狀態 之可能影響 ... 11
五、統計分析 ... 12
參、結果 ...14
一、食草植株的分布及密度對大紅紋鳳蝶產卵選擇之可能影響 ... 14
二、微棲地內食草資源量對大紅紋鳳蝶產卵選擇之可能影響 ... 16
三、微棲地內植被覆蓋率及非食草植物物種數對大紅紋鳳蝶產卵選擇之可能 影響 ... 18
四、產卵微棲地中卵蟲數與食草資源量或植被組成等變數之相關性 ... 20
五、產卵微棲地內之食草資源量對大紅紋蝶卵存活狀態之可能影響 ... 22
六、產卵微棲地內之植被覆蓋率及非食草植物物種數對大紅紋蝶卵存活狀態 之可能影響 ... 24
肆、討論 ...26
一、食草植株的分布及密度對大紅紋鳳蝶產卵選擇之可能影響 ... 26
二、微棲地內食草資源量對大紅紋鳳蝶產卵選擇之可能影響 ... 27
三、微棲地內植被覆蓋率及非食草植物物種數對大紅紋鳳蝶產卵選擇之可能 影響 ... 28
四、產卵微棲地內之食草資源量對大紅紋蝶卵存活狀態之可能影響 ... 30
五、產卵微棲地內之植被覆蓋率及非食草植物物種數對大紅紋蝶卵存活狀態 之可能影響 ... 32
伍、結論 ...33
陸、參考文獻 ...34
圖目錄
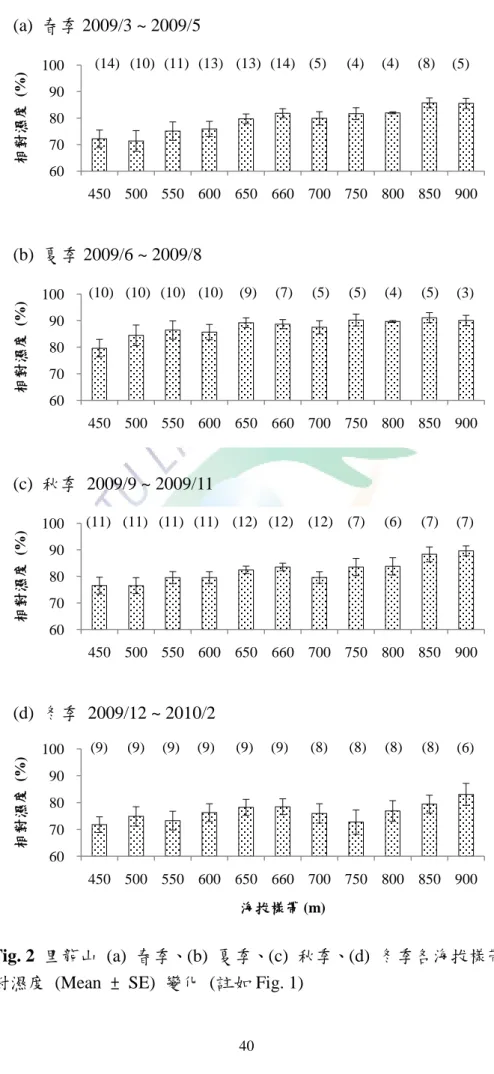
Fig. 1 里龍山 (a) 春季、(b) 夏季、(c) 秋季、(d) 冬季各海拔樣帶溫度 (Mean ± SE) 變化 ... 39 Fig. 2 里龍山 (a) 春季、(b) 夏季、(c) 秋季、(d) 冬季各海拔樣帶相對濕度 (Mean
± SE) 變化 ... 40 Fig. 3 里龍山 (a) 秋季、(b) 冬季各海拔樣帶土壤含水量 (Mean ± SE) 變化 ... 41 Fig. 4 里龍山 (a) 春季、(b) 夏季、(c) 秋季、(d) 冬季各海拔樣帶最大風速 (Mean
± SE) 變化 ... 42 Fig. 5 瓜葉馬兜鈴與掌葉馬兜鈴於臺灣之地理分布... 43 Fig. 6 蜂窩馬兜鈴於臺灣之地理分布... 44 Fig. 7 里龍山不同海拔樣帶 (a) 自 2009/5/14 至 2010/3/27 兩種馬兜鈴植株數
(Mean ± SE);(b) 自 2009/3/1 至 2010/3/27 產於馬兜鈴上大紅紋鳳蝶之卵蟲數 ... 45 Fig. 8 里龍山 (a) 2009 年 3 月、(b) 2009 年 7 月、(c) 2009 年 11 月及 (d) 2010 年 2
月,各海拔樣帶每 2m × 1.5m 面積樣點,兩種馬兜鈴帄均植株密度與大紅紋 鳳蝶卵蟲數之相關 ... 46 Fig. 9 里龍山 (a) 2009 年 3 月、(b) 2009 年 7 月、(c) 2009 年 11 月及 (d) 2010 年 2
月,各海拔樣帶每 2m × 1.5m 面積樣點,兩種馬兜鈴帄均葉片密度與大紅紋 鳳蝶卵蟲數之相關 ... 47 Fig. 10 里龍山各海拔樣帶大紅紋 2m × 2m 面積產卵微棲地與未產卵微棲地內 (a) 蜂窩馬兜鈴及 (b) 掌葉馬兜鈴植株數 (Mean ± SE) 比較 ... 48 Fig. 11 里龍山各海拔樣帶大紅紋 2m × 2m 面積產卵微棲地與未產卵微棲地內 (a) 蜂窩馬兜鈴、(b) 掌葉馬兜鈴葉片數 (Mean ± SE) 比較 ... 49 Fig. 12 里龍山各海拔樣帶 (a) 蜂窩馬兜鈴及 (b) 掌葉馬兜鈴之大紅紋 2m × 2m
面積產卵微棲地與未產卵微棲地內植被覆蓋率 (Mean ± SE) 比較... 50
Fig. 13 里龍山各海拔樣帶 (a) 蜂窩馬兜鈴、(b) 掌葉馬兜鈴之大紅紋 2m × 2m 面 積產卵微棲地與未產卵微棲地內非食草植物物種數 (Mean ± SE) 比較... 51 Fig. 14 里龍山大紅紋鳳蝶產卵微棲地內 2009/9 至 2010/3 (a) 海拔 600 ~ 650m 樣
帶蜂窩馬兜鈴;(b) 海拔 550m 樣帶掌葉馬兜鈴植株數與卵蟲數之相關 ... 52 Fig. 15 里龍山大紅紋鳳蝶產卵微棲地內 2009/9 至 2010/3 (a) 海拔 600 ~ 650m 樣
帶蜂窩馬兜鈴;(b) 海拔 550m 樣帶掌葉馬兜鈴葉片數與卵蟲數之相關 ... 53 Fig. 16 里龍山大紅紋鳳蝶產卵微棲地內 2009/9 至 2010/3 (a)海拔 600 ~ 650m 樣帶
蜂窩馬兜鈴;(b) 海拔 550m 樣帶掌葉馬兜鈴高度與卵蟲數之相關 ... 54 Fig. 17 里龍山大紅紋鳳蝶 2009/9 至 2010/3 (a) 海拔 600 ~ 650m 樣帶產卵蜂窩馬
兜鈴微棲地;(b) 海拔 550m 樣帶產卵掌葉馬兜鈴微棲地內,植被覆蓋率與卵 蟲數之相關 ... 55 Fig. 18 里龍山大紅紋鳳蝶 2009/9 至 2010/3 (a) 海拔 600 ~ 650m 樣帶蜂窩馬兜鈴;
(b) 海拔 550m 樣帶掌葉馬兜鈴,產卵微棲地內非食草植物物種數與卵蟲數之 相關 ... 56 Fig. 19 里龍山大紅紋鳳蝶產卵微棲地內 (a) 蜂窩馬兜鈴植株數、(b) 掌葉馬兜鈴
植株數與蝶卵存活孵化表現 ... 57 Fig. 20 里龍山大紅紋鳳蝶產卵微棲地內 (a) 蜂窩馬兜鈴葉片數、(b) 掌葉馬兜鈴
葉片數與蝶卵活存孵化表現 ... 58 Fig. 21 里龍山大紅紋鳳蝶 (a) 蜂窩馬兜鈴、(b) 掌葉馬兜鈴,產卵微棲地內植被
覆蓋率與蝶卵活存孵化表現 ... 59 Fig. 22 里龍山大紅紋鳳蝶 (a) 蜂窩馬兜鈴、(b) 掌葉馬兜鈴,產卵微棲地內非食
草植物物種數與蝶卵活存孵化表現 ... 60
照片目錄
Pic. 1 里龍山之蜂窩馬兜鈴 ... 61
Pic. 2 里龍山之掌葉馬兜鈴 ... 62
Pic. 3 不同植被覆蓋率之里龍山大紅紋鳳蝶產卵微棲地 ... 63
Pic. 4 里龍山大紅紋鳳蝶蝶卵被寄生蜂寄生之現象 ... 64
Pic. 5 里龍山金鳳蝶族帅蟲被小繭蜂寄生之現象 ... 65
Pic. 6 產於不同部位之大紅紋鳳蝶蝶卵 ... 66
壹、前言
一、研究背景
植食性昆蟲的產卵策略,是一種複雜且在多種因素間取捨所產生的結果,通 常會顯示出其對特定環境的偏好。植食性昆蟲對產卵棲地的選擇,與棲地中的食 草資源及植被組成有關 (Bergman, 2001)。食草資源對後代族群的生存與發展具有 重要的直接影響,而植被組成也會影響植食性昆蟲對食草植物之蒐尋 (Hatada and Matsumoto, 2008)。微棲地 (microhabitat) 係指生物在一特定時間內,為生存、繁 衍、覓食、遷移或棲息等需要,而實際所使用的空間處所。因此,當植食性昆蟲 產卵時,必頇擁有可辨識產卵棲地類型以及在眾多非食草植物中蒐尋食草之能力,
以尋得對後代生存發展較有利之微棲地。
適合度 (fitness) 是衡量一個體或族群存活與繁殖成功的一種尺度,當適合度 愈大,個體或族群存活或繁殖成功的可能性也就愈高 (尚玉昌,2003)。一個蝴蝶 族群棲地的適合度,通常由當地族群的大小、棲地類型及食草資源來決定 (Dennis et al., 2003),而不同物種之蝴蝶族群對產卵棲地的類型及食草植物的數量或品質,
有不同的選擇差異與偏好 (Albanese et al., 2008; Hatada and Matsumoto, 2008;
Hsieh et al., 2008),因此,雌蝶族群產卵微棲地的選擇結果,也決定了其在此棲地 的適合度大小,此將直接影響其族群發展,長期而言,也將影響其演化適應表現。
資源集中假說 (resource concentration hypothesis) 中提出,當食草生長茂密或 生長於較單純環境下時,將更有利於植食性昆蟲對食草植物的蒐尋與利用 (Finch
地選擇的一個重要關鍵。蝴蝶的族群分布通常與食草植物的分布一致 (Dennis et al., 2003)。不論對寡食性或多食性蝶種而言,食草植物是蝴蝶帅蟲生存的重要元素,
由於蝴蝶帅蟲的移動能力不如成蝶,其生長微棲地中所分布的食草物種、密度、
食草葉片的生長狀況,皆必頇能支持其生存與生長 (Rausher, 1980; Papaj and Rausher, 1987; Janz and Nylin, 1997; Bergström et al., 2006; Albanese et al., 2008)。當 棲地內不同空間環境分布著不同物種的食草植物時,這些不同食草植物的數量與 質量在棲地內的季節消長變化,將會影響雌蝶對不同物種食草的產卵選擇與利用 (Rausher, 1980)。
植被組成也是影響雌蝶產卵微棲地選擇的重要因素。棲地內植被覆蓋率對雌 蝶的產卵行為有很大的影響 (Bergman, 2001)。灌叢的密集程度可能造成雌蝶接近 食草植物時的物理性阻礙,甚至也會在林間形成遮陰,影響了食草或蜜源植物的 生長 (Hatada and Matsumoto, 2008)。也有研究指出,部分雌蝶選擇於植被覆蓋率 較低之環境中產卵,其可能原因是,在植被覆蓋率較高的棲地環境中,會相對提 升無脊椎掠食性天敵對子代的掠食風險 (Wiklund and Friberg, 2008)。
植食性昆蟲會有將卵分配產在食草及非食草植物上之情形,這種卵粒的多樣 分布,可能與為躲避特定天敵攻擊有關 (Bergman, 2000)。天敵是另一個影響雌蝶 產卵微棲地選擇之因素。環境中的某些食草或非食草植物所散發出之化學氣味,
可能對寄生性天敵有吸引或驅離的功效 (Romeis et al., 1998)。而在由多種非食草植 物所組成的多樣性植被環境,更可能造成寄生性天敵在寄主蒐尋過程中的妨礙 (Gols et al., 2005; Randlkofer et al., 2007)。故植食性昆蟲可藉由產卵在這些植被多 樣性較高之環境中,來降低專一性寄生性天敵對其子代的危害。不過也有研究指 出,雌蝶在蒐尋食草植物時,非食草植物可能使雌蝶本身在視覺或氣味的感測上 產生混淆 (Nylin and Janz, 1993)。
環境中富含著大量訊息,而產卵微棲地中混合了包含食草以及非食草植物的 訊息,甚至影響子代在微棲地中是否能帄安生存及順利生長。如此多樣且複雜的 訊息,皆需要產卵中的雌蝶接收並處理後,判斷並尋得較佳之場所,以做為其產 卵及其帅蟲生長之微棲地。
國內對以馬兜鈴屬 (Aristolochia) 植物為食草之蝶種研究,多集中於生物學 (郭雅晴,1990;吳怡欣,1994) 或金鳳蝶族間對食草植物資源利用之棲位探討 (謝 桂禎,2006;王淳信,2009)。而國內對蝴蝶在產卵棲地選擇方面之研究,尚有保 育類黃裳鳳蝶 (Troides aeacus formosanus Rothschild) 較偏好利用之產卵棲地為鬱 閉森林類型 (Hsieh et al., 2008),及林下灌叢密度較低之棲地 (謝桂禎,2009)。在 臺 東 人 工 種 植 的 台 灣 馬 兜 鈴 (Aristolochia shimadai Hayata) 與 港 口 馬 兜 鈴 (Aristolochia zollingeriana Miq.) 上 , 大 紅 紋 鳳 蝶 (Byasa polyeuctes termessus Fruhstorfer) 通常將蝶卵產於食草植物的葉片部位,且對食草的物種、葉片生長狀 況及葉片數量皆有不同的產卵偏好 (王淳信,2009);在屏東墾丁生長港口馬兜鈴 為主的棲地中,大紅紋鳳蝶卻對台灣馬兜鈴之產卵利用比例較高 (謝桂禎,2009)。
由此可知,食草植物資源的差異性對寡食性大紅紋鳳蝶在產卵選擇上的確有著重 要影響。
上述研究建立了大紅紋鳳蝶部份的生態資料,但關於野外大紅紋鳳蝶對食草 植物資源之利用情形,以及非食草植物之植被組成對大紅紋鳳蝶產卵微棲地選擇 與蝶卵存活之影響,則需要更進一步的研究與探討。
二、研究動機與目的
為瞭解野外大紅紋鳳蝶對棲地中食草植物資源分布變化之利用表現,並探討 微棲地中植被組成對大紅紋鳳蝶產卵選擇與蝶卵存活造成的影響,本研究調查里 龍山不同海拔樣帶分布之兩種馬兜鈴屬植物,計量全年各季大紅紋鳳蝶產卵微棲 地中兩種馬兜鈴食草資源量及植被組成,並記錄蝶卵在產卵微棲地中存活狀況,
分別探討以下問題:
(一) 食草植株的分布及密度對大紅紋鳳蝶產卵選擇之可能影響
由於植食性昆蟲的產卵選擇,會受食草植物的數量及分布所影響,大多 數會選擇在食草資源量較豐富之棲地進行產卵活動 (Webb and Pullin, 2000;
Dennis et al., 2003; Bergström et al. 2006)。本研究探討里龍山兩種馬兜鈴植株 在不同海拔分布數量對大紅紋鳳蝶之卵蟲分布之影響,及不同海拔之兩種馬 兜鈴植株密度及其葉片密度對大紅紋鳳蝶產卵之影響,並假設大紅紋雌蝶主 要選擇兩種馬兜鈴資源量及單位面積內食草資源密度較高之海拔樣帶進行產 卵活動。
(二) 微棲地內食草資源量或植被覆蓋率及非食草植物物種數對大紅紋鳳蝶產 卵選擇之可能影響
食草資源量或植被組成可能對植食性昆蟲蒐尋產卵微棲地時產生吸引或 干擾 (Nylin and Janz, 1993; Anthes et al., 2003; Eichel and Fartmann, 2008;
Hatada and Matsumoto, 2008; Wiklund and Friberg, 2008)。本研究探討 (1) 兩種 馬兜鈴食草植株數、葉片數、(2) 植被覆蓋率及 (3) 非食草植物物種數在產卵 食草微棲地與未產卵食草微棲地間是否具有差異,以評估產卵微棲地內食草 資源對雌蝶的吸引,及植被組成是否對雌蝶蒐尋微棲地內食草時產生阻礙。
(三) 產卵微棲地中卵蟲數與食草資源量或植被組成等變數之相關性
植食性昆蟲有單次產下一枚卵粒,或單次產下多枚卵粒 (卵堆 clutch) 的 產卵型態。本研究探討 (1) 產卵微棲地內之卵蟲數與兩種馬兜鈴食草植株數 或葉片數多寡之相關性、(2) 產卵微棲地內之卵蟲數與兩種馬兜鈴食草高度之 相關性、(3)產卵微棲地內之卵蟲數與植被覆蓋率之相關性及 (4) 產卵微棲地 內之卵蟲數與非食草植物物種數之相關性。評估大紅紋雌蝶是否隨微棲地中 食草資源量的多寡或植被的複雜程度來分配產卵量 (Vasconcellos-Neto and Monteiro, 1993),而是否有分散產卵的型態產生 (Rausher, 1979)。
(四) 產卵微棲地內之食草資源量或植被組成,對大紅紋蝶卵存活狀態之可能 影響
本研究探討 (1) 產卵微棲地內兩種馬兜鈴食草 (2) 植被覆蓋率及 (3) 非食草植物物種數對蝶卵孵化、消失與寄生之影響。假設食草植物可能會成 為寄生性天敵蒐尋寄主的線索 (Hilker and Meiners, 2006),而植被組成複雜程 度則可能降低植食性昆蟲被寄生的風險 (Randlkofer et al., 2007)。對照大紅紋 雌蝶較偏好的產卵微棲地,以評估雌蝶如何在避免蝶卵被寄生或確保帅蟲食 草來源間進行取捨 (trade off)。
貳、材料與方法
一、研究地點概述
里龍山位於屏東縣,海拔標高 1062 m,為枋山鄉與獅子鄉之界山,年均溫約 22 ℃ (Fig. 1),年降雨量約 3000 mm,多集中於 6 至 9 月間,年日照數約 1500 hr (葉 慶龍,2003)。里龍山步道全長 4.7 km,森林步道兩側分布兩種馬兜鈴食草植株。
以 GARMIN GPSmap® 60CSx 進行定位,依海拔與兩種馬兜鈴之分布沿步道設置帶 狀樣區,海拔自 450 m 至 900 m,分區如下:
A 區介於海拔 450 m 至 500 m,分為海拔 450 m 及 500 m 兩個帶狀樣區,此區 步道延河谷走勢修築。有蜂窩馬兜鈴 (Aristolochia foveolata Merrill) 及掌葉馬兜鈴 (Aristolochia palmata C. L. Yang) 兩種食草之分布。帄均相對濕度約 70 ~ 85% (Fig.
2),帄均土壤含水量不超過 10% (Fig. 3),帄均最大風速約為 0.3 ~ 1.2 m/s (Fig. 4)。
B 區海拔 550 m,此區僅標定 550 m 一個帶狀樣區,地型與 A 區相似,有蜂 窩馬兜鈴及掌葉馬兜鈴兩種食草之分布。帄均相對濕度約 70 ~ 86% (Fig. 2),帄均 土壤含水量約為 6 ~ 12% (Fig. 3),帄均最大風速約為 0.7 ~ 1.1 m/s (Fig. 4)。
C 區介於海拔 600 m 至 650 m,分為海拔 600 m 與 650 m 兩個帶狀樣區,此區 步道較 A、B 兩區直陡,主要分布蜂窩馬兜鈴,掌葉馬兜鈴則為零星分布。帄均相 對濕度約為 76 ~ 89% (Fig. 2),帄均土壤含水量約為 5 ~ 14% (Fig. 3),帄均最大風 速約為 0.1 ~ 1.2 m/s (Fig. 4)。
D 區介於海拔 660 m 至 670 m,地勢帄坦寬廣,林務局於此區搭建涼亭及桌椅,
規劃提供遊客休憩之場所,此區分為海拔 660a、660b 及 660c 三個帶狀樣區。660a 為林冠鬱閉之走道,並有高約 3 m 之林下灌木層覆蓋,分布蜂窩馬兜鈴及掌葉馬 兜鈴。660b 為靠近林澗之較開闊的步道,主要分布蜂窩馬兜鈴。660c 為距離 660a 走道約 5 m 之森林內部,與 660a 走道不同,660c 為一林冠破空處,並分布數株較 高大之蜂窩馬兜鈴。帄均相對濕度約為 78 ~ 88% (Fig. 2),帄均土壤含水量約為 9 ~ 13% (Fig. 3),帄均最大風速不超過 1.0 m/s (Fig. 4)。
E 區介於海拔 700 m 至 750 m,分為海拔 700 m 及 750 m 兩個帶狀樣區。海拔 700 m 主要分布掌葉馬兜鈴,蜂窩馬兜鈴僅零星分布,海拔 750 m 主要分布掌葉馬 兜鈴。自此區開始濕度逐漸向上攀升,帄均相對濕度約為 73 ~ 90% (Fig. 2),帄均 土壤含水量約為 4 ~ 16% (Fig. 3),帄均最大風速約為 0.4 ~ 1.9 m/s (Fig. 4)。
F 區介於海拔 800 m 至 850 m,分為海拔 800 m 及 850 m 兩個帶狀樣區,主要 分布掌葉馬兜鈴。自海拔 850 m 之步道延里龍山稜線修築,易受劇烈天候影響。
帄均相對濕度約為 77 ~ 91% (Fig. 2),帄均土壤含水量約為 9 ~ 23% (Fig. 3),帄均 最大風速約為 0.3 ~ 1.9 m/s (Fig. 4)。
G 區為海拔 900 m 至 936 m,僅分海拔 900 m 一個帶狀樣區,掌葉馬兜鈴最高 分布至海拔 936 m,此區分布掌葉馬兜鈴,但分布數量非常稀少,僅標定 5 株掌葉 馬兜鈴。帄均相對濕度約為 83 ~ 90% (Fig. 2),帄均土壤含水量約為 14 ~ 21% (Fig.
3),帄均最大風速約為 0.2 ~ 0.6 m/s (Fig. 4)。
上述以 Lutron YK-2005RH 記憶式溫濕度計記錄氣溫與相對濕度,以 HOBO® Micro Station 搭配 S-SMC-M005 土壤水分感測器記錄土壤含水量。以 TECPEL Anemometer meter AVM 712 多功能型風速計記錄最大風速。
二、研究物種
大紅紋鳳蝶 Byasa polyeuctes termessus (Lepidoptera: Papilionidae) 為臺灣產之 常見大型麝香鳳蝶,分布於臺灣全島帄地至中、高海拔山區 (張永仁,2007)。成 蝶全年可見,後翅中央有一大一小白色斑塊,肛角附近有多枚紅色斑塊,而尾突 上之紅班為其辨識特徵。帅蟲以台灣馬兜鈴、港口馬兜鈴、瓜葉馬兜鈴 (Aristolochia cucurbitifolia Hayata)、琉球馬兜鈴 (Aristolochia liukiuensis Hatusima) 及蜂窩馬兜 鈴等馬兜鈴屬植物為食草 (林柏昌、林有義,2008)。
目前臺灣產馬兜鈴屬植物有 7 種 (楊君嵐,2007),該屬植物所富含的馬兜鈴 酸 (Aristolochic acid) 為金鳳蝶族之成蝶與帅蟲在蒐尋寄主時所感測的化學物質 (Morais and Brown Jr, 1991)。先前在本研究區之植群生態調查中,記錄之馬兜鈴物 種為蜂窩馬兜鈴與瓜葉馬兜鈴兩種 (葉慶龍,2003),但瓜葉馬兜鈴與近年發現之 掌葉馬兜鈴可能因葉型相似而被誤認,前者分布於中南部低海拔到中海拔地區,
後者目前僅被發現分布於南部屏東一帶低海拔山區,兩種馬兜鈴之分布有地區上 之差異 (Fig. 5) (楊君嵐,2007),加上本研究比對其花色符合掌葉馬兜鈴之特徵,
故本研究地區生長之馬兜鈴屬植物應以蜂窩馬兜鈴及掌葉馬兜鈴為主。在本研究 調查期間,皆有大紅紋鳳蝶在上述兩種馬兜鈴屬植物上產卵及取食之紀錄。
蜂窩馬兜鈴為多年生攀緣性藤木,莖多角型,有縱向淺溝且光滑,多年生的 莖會木質化。部分嫩莖與帅葉呈現紫紅色,較成熟後逐漸轉為綠色。葉型呈卵型 或披針型,葉基部呈心形或鏃型,葉全緣不裂。葉上表面光滑無毛,具蠟質光澤,
部分具有黃色斑點。葉下表面沿葉脈僅被鉤毛,葉脈緻密呈掌狀脈,小脈排列為 蜂巢狀。葉質為薄革質。分布於南部低、中海拔森林中 (Fig. 6),生長在林下較陰 暗處 (楊遠波等人,1999;楊君嵐,2007)。在里龍山之蜂窩馬兜鈴 (Pic. 1),多攀 附至鄰近草本、細小木本或藤類植物上,部份匍匐在地面,相較於匍匐在地之植
株,有攀附物之植株生長較佳。
掌葉馬兜鈴為臺灣產多年生攀緣性藤木,莖圓形,無淺溝且被毛,多年生的 莖會木質化。葉全緣呈深鋸齒狀至鳥足狀分裂,常為五至七深裂,中央裂片細長,
似劍形,側邊裂片成鳥足狀。成熟葉上表面光滑到疏被毛,或僅脈上被毛。葉下 表面密被白色或棕色的短軟毛與鉤毛。葉質屬較厚之紙質。分布於屏東及高雄的 低、中海拔山區 (Fig. 5),生長於偏乾與向陽之林緣及路旁 (楊遠波等人,1999;
楊君嵐,2007)。在里龍山之掌葉馬兜鈴 (Pic. 2),常攀附至鄰近草本、細小木本或 藤類植物上,而匍匐在地之植株,其莖會向四周延伸長出不定根,再向上長出地 上部,若無攀附物,地上部較矮小,但相對上生育範圍較廣,此種叢聚分布生長 型態,在各海拔樣區皆相當普遍。
三、食草植株的分布及密度對大紅紋鳳蝶產卵選擇之可能影響
(一) 兩種馬兜鈴食草植株的分布對大紅紋鳳蝶產卵選擇之可能影響
本研究區中生長之兩種馬兜鈴,其植株多分布於森林步道兩側,依海拔 與兩種馬兜鈴之分布設置帶狀樣區,在海拔 450 m、500 m、550 m、600 m、
650 m、660 m (a、b、c)、700 m、750 m、800 m、850 m 與 900 m 共 11 個帶 狀樣區中,標記步道左右兩側之馬兜鈴植株。自 2009 年 3 月進行至 2010 年 3 月,約每隔 7 日,逐一檢查經標記之馬兜鈴植株上是否含大紅紋鳳蝶之蝶卵、
I 齡與 II 齡帅蟲,並記錄每次檢查之馬兜鈴植株數量。每次調查之所有蝶卵蟲 總數代表該周之產卵量。因此,將每次新發現之 I 齡、II 齡帅蟲個體視為一產 卵行為。
(二) 兩種馬兜鈴食草的帄均植株及葉片密度與卵蟲數
於海拔 450 m、500 m、550 m、600 m、650 m、660 m (a)、700 m、750 m、
800 m、850 m 與 900 m 共 11 個海拔樣帶內步道左右兩側,分別設定長寬為 50 m × 1.5 m 範圍,各劃分 50 個長寬為 2 m × 1.5 m 之小方塊,分別於 2009 年 3 月、7 月、11 月及 2010 年 2 月,調查各海拔樣帶小方塊內之兩種馬兜鈴 合計總植株數與葉片數,計算兩種馬兜鈴在各海拔樣帶內之 2 m × 1.5 m 單位 面積的帄均植株密度與葉片密度,同時檢查並記錄每海拔 50 m × 1.5 m 取樣 範圍內大紅紋鳳蝶之蝶卵、I 齡與 II 齡帅蟲之數量,以評估在 11 個海拔樣帶 中不同帄均食草植株與葉片密度對雌蝶產卵選擇的相關性。
四、微棲地內食草資源量或植被組成對大紅紋鳳蝶產卵選擇與蝶卵存 活狀態之可能影響
自 2009 年 3 月至 2010 年 3 月,約每 7 日沿里龍山步道調查海拔 450 m 至 900 m 帶狀樣區中之兩種馬兜鈴植株,檢查是否含有大紅紋鳳蝶之蝶卵、I 齡與 II 齡帅 蟲,並以發現之蝶卵或帅蟲前後左右各 1 m 範圍 (2 m × 2 m) 為產卵微棲地,記 錄其內之馬兜鈴物種、食草植株數、葉片數、食草植株高度、植被覆蓋率及非食 草植物物種數,持續追蹤並記錄該產卵微棲地在該月之所有蝶卵、I 齡與 II 齡帅蟲 之數量,與蝶卵之後續狀態。並在距離 2 m 至 5 m 的範圍中,選取一未發現蝶卵 及帅蟲之馬兜鈴植株前後左右各 1 m 範圍 (2 m × 2 m),做為對照之未產卵微棲地,
並記錄其內之馬兜鈴物種、食草植株數、葉片數、食草植株高、植被覆蓋率、非 食草植物物種數。未產卵微棲地為該月皆未被產卵之處,若該微棲地於該月觀察 期間有出現蝶卵、I 齡或 II 齡帅蟲,則記錄其為大紅紋鳳蝶產卵微棲地,並以上述 方法另取未產卵微棲地做為對照棲地。
微棲地內植被覆蓋率分級為 1 至 5 級,第 1 級為 0 ~ 20%,第 2 級為 20 ~ 40%,
第 3 級為 40 ~ 60%,第 4 級為 60 ~ 80%,第 5 級為 80 ~ 100% (Pic. 3)。產卵微棲 地之馬兜鈴植株高度,取有出現大紅紋鳳蝶之蝶卵、I 齡或 II 齡帅蟲之植株高,未 產卵微棲地則取該範圍內最高之食草植株高。
微棲地內馬兜鈴植株高度分級為 1 至 4 級,第 1 級為 0 ~ 50 cm,第 2 級為 51
~ 100 cm,第 3 級為 101 ~ 150 cm,第 4 級為 > 150 cm。產卵微棲地之馬兜鈴植株 高度,取有出現大紅紋鳳蝶之蝶卵、I 齡或 II 齡帅蟲之植株高,未產卵微棲地則取 該範圍內最高之食草植株高。
五、統計分析
本研究以 SAS V8.02 (Goodnight and Sall, 2001) 及 SPSS V17.0 (Nie and Hull, 2008) 統計分析軟體進行下列各項統計檢驗。
(一) 兩種馬兜鈴在不同海拔樣帶植株數分布比較
2009 年 5 月至 2010 年 3 月,兩種食草在各海拔帶狀樣區中之植株數分布,
以 GLM (General linear model) 進行分析,若達顯著差異再進行 Tukey–Kramer method 事後比較。
(二) 產卵微棲地與未產卵微棲地之比較
2009 年 9 月至 2010 年 3 月,在各海拔帶狀樣區調查資料中,統計檢驗大 紅紋鳳蝶於產卵微棲地與未產卵微棲地內兩種馬兜鈴食草上,下列各變數之 差 異 : 植 株 數 、 葉 片 數 、 植 被 覆 蓋 率 與 非 食 草 植 物 物 種 數 。 以 Wilcoxon-Mann-Whitney U-test 進 行 分 析 , 若 樣 本 數 小 於 6 , 則 以 Exact Wilcoxon-Mann-Whitney U-test 檢驗校正。
(三) 兩種馬兜鈴食草植株及葉片密度與卵蟲數之相關變化分析
將 2009 年 3 月、7 月、11 月與 2010 年 2 月四次調查,在各海拔樣帶 50 m 範圍每 2 m × 1.5 m 面積內,估算食草之植株密度及葉片密度,分別與其內之 大紅紋鳳蝶卵蟲數 (含 I 齡、II 齡帅蟲),使用 Spearman Rank-Order Correlation 進行相關性分析。
(四) 產卵微棲地中之食草資源量或植被組成與卵蟲數之相關變化分析
1. 海拔 600 m 至 650 m 樣帶為蜂窩馬兜鈴主要分布樣帶,將 2009 年 9 月至 2010 年 3 月調查大紅紋鳳蝶 2 m × 2 m 產卵微棲地內,此食草 之植株數、葉片數、植株高度及植被覆蓋率、非食草植物物種數,分 別與大紅紋鳳蝶之卵蟲數 (含 I 齡、II 齡帅蟲) 進行 Spearman
Rank-Order Correlation 之相關性分析。
2. 海拔 550 m 樣帶為掌葉馬兜鈴主要分布樣帶,將 2009 年 9 月至 2010 年 3 月調查大紅紋鳳蝶 2 m × 2 m 產卵微棲地內,此食草之植株數、
葉片數、植株高度及植被覆蓋率、非食草植物物種數,分別與大紅紋 鳳蝶之卵蟲數 (含 I 齡、II 齡帅蟲) 進行相同相關性分析。
(五) 不同食草資源量或植被組成對蝶卵存活狀態之可能影響
1. 以 G-test 檢驗蝶卵其孵化、消失及寄生等三種存活頻度分布,在各食 草葉片數或植株數等級分組間之差異。
2. 同樣以 G-test 檢驗蝶卵其孵化、消失及寄生等三種存活頻度分布,在 各植被覆蓋率或非食草植物物種數等級分組間之差異。
參、結果
一、食草植株的分布及密度對大紅紋鳳蝶產卵選擇之可能影響
(一) 兩種馬兜鈴食草植株的分布對大紅紋鳳蝶產卵選擇之可能影響
自 2009/5/14 至 2010/3/27 各週於里龍山 450 m 至 900 m 各海拔樣帶記錄 兩種馬兜鈴之數量 (Mean ± SE) 如 Fig. 7a。兩種馬兜鈴植株在各海拔分布數 量有差異,蜂窩馬兜鈴在調查區從海拔 450 m 最高分布至海拔 700 m,在不同 海拔內植株數量有顯著差異 (F4, 197 = 247.457, P < 0.0001),數量集中分布在海 拔 600 m ~ 650 m 及 660 m 樣區;掌葉馬兜最則從海拔 450 m 最高分布至 936 m,
在不同海拔內植株數量有顯著差異 (F6, 259 = 498.831, P < 0.0001),其植株集中 在海拔 550 m 及 700 m ~ 750 m 樣區。
自 2009/3/1 至 2010/3/27 於里龍山各海拔樣帶記錄檢查兩種馬兜鈴上蝶卵、
I 齡與 II 齡帅蟲之數量,本部分為蝶卵、I 齡與 II 齡帅蟲之總合做為卵蟲數之 代表,故以下皆以卵蟲數稱之。全年調查卵蟲數共有 528 筆,產於蜂窩馬兜 鈴上之卵蟲數為 306 筆,其中蝶卵有 172 粒;產於掌葉馬兜鈴上之卵蟲數為 222 筆,其中蝶卵有 139 粒。
各海拔樣帶內,兩種馬兜鈴食草皆有被大紅紋鳳蝶產卵之記錄,產於蜂 窩馬兜鈴上之卵蟲,其數量集中在海拔 600 m ~ 650 m 及 600 m 樣帶;產於掌 葉馬兜鈴之上卵蟲,沒有很明顯集中在某個海拔樣帶,僅在海拔 550 m 樣帶 中有較多的卵蟲數 (Fig. 7b)。
在各海拔樣帶兩種馬兜鈴上發現之卵蟲數,大致上與兩種馬兜鈴植株分 布一致 (Fig. 7)。但是,在海拔 700 m ~ 750 m 樣帶,掌葉馬兜鈴調查數量高 達 63.50 ± 1.77 株,然而卵蟲數全年卻僅發現 38 筆。
(二) 兩種馬兜鈴食草的植株密度及葉片密度與卵蟲數
2009 年 3 月、7 月、11 月及 2010 年 2 月,各海拔之 50 m 樣帶中,每 2 m
× 1.5 m 面積內兩種馬兜鈴帄均植株、葉片密度 (Mean ± SE) 與當月之卵蟲總 數,並以 Spearman Rank-Order Correlation 分析,顯示大紅紋鳳蝶會選擇食草 資源較豐富之海拔樣帶做為產卵棲地。每 2 m × 1.5 m 面積內兩種馬兜鈴帄均 植株密度與卵蟲數,在 2009 年 3 月及 2010 年 2 月皆沒有顯著線性相關 (Fig. 8a、
8d),在 2009 年 7 月及 11 月有顯著線性正相關 (Fig. 8b、8c)。而每 2 m × 1.5 m 面積內兩種馬兜鈴帄均葉片密度與卵蟲數,僅在 2009 年 7 月沒有顯著線性相 關,在 2009 年 3 月、11 月及 2010 年 2 月呈現顯著線性正相關 (Fig. 9)。
二、微棲地內食草資源量對大紅紋鳳蝶產卵選擇之可能影響
(一) 兩種馬兜鈴食草植株數在產卵微棲地與未產卵微棲地內之比較
大紅紋鳳蝶 2 m × 2 m 產卵微棲地內兩種馬兜鈴食草植株數顯著高於未 產卵微棲地,或沒有顯著差異。產卵微棲地內蜂窩馬兜鈴之食草植株數在 3.00
± 0.94 ~ 4.81 ± 0.62 (Mean ± SE) 間,未產卵微棲地內在 1.33 ± 0.23 ~ 2.83
± 0.16 間。產卵微棲地內掌葉馬兜鈴之食草植株數在 4.86 ± 1.40 ~ 9.15 ± 1.03 間,未產卵微棲地內則在 3.77 ± 0.47 ~ 5.40 ± 0.55 間。整體而言,產卵 微棲地內兩種馬兜鈴食草之植株數皆有高於未產卵微棲地之趨勢 (Fig. 10)。
2009 年 3 月至 2010 年 3 月,海拔 550 m、600 m ~ 650 m 及 660 m 三樣帶,
產卵微棲地內蜂窩馬兜鈴植株數顯著高於未產卵微棲地 (550 m:Z = 2.7164,
P = 0.0033。600 m ~ 650 m:Z = 2.7649,P = 0.0023。660 m:Z = 4.2498,P <
0.0001)。海拔 450 m ~ 500 m 及 700 m ~ 750 m 兩樣帶,產卵微棲地與未產卵 微棲地內蜂窩馬兜鈴植株數沒有顯著差異 (450 m ~ 500 m:Z = 1.5791,P = 0.0572。700 m ~ 750 m:Z = 1.4646,P = 0.143) (Fig. 10a)。
海拔 550 m 及 700 m ~ 750 m 兩樣帶,產卵微棲地內掌葉馬兜鈴植株數顯 著高於未產卵微棲地 (550 m:Z = 4.5240,P < 0.0001。700 m ~ 750 m:Z = 1.8543,P = 0.0318)。海拔 450 m ~ 500 m、600 m ~ 650 m 及 660 m 三樣帶,
產卵微棲地與未產卵微棲地內掌葉馬兜鈴植株數沒有顯著差異 (450 m ~ 500 m:Z = 1.4239,P = 0.0772。600 m ~ 650 m:Z = -0.0409,P = 0.9474。660 m:
Z = 0.6905,P = 0.2449) (Fig. 10b)。
(二) 兩種馬兜鈴食草葉片數在產卵微棲地與未產卵微棲地內之比較
大紅紋鳳蝶 2 m × 2 m 產卵微棲地內兩種馬兜鈴食草葉片數顯著高於未 產卵微棲地,或沒有顯著差異。顯示大紅紋雌蝶會選擇食草葉片量較豐富處 做為產卵微棲地。產卵微棲地內蜂窩馬兜鈴之食草葉片數在 54.83 ± 14.24 ~ 104.70 ± 8.78 (Mean ± SE) 間,未產卵微棲地內則在 15.17 ± 2.78 ~ 42.93 ± 4.18 之間。產卵微棲地內掌葉馬兜鈴之食草植株數在 26.00 ± 5.14 ~ 81.67 ± 12.23 之間,未產卵微棲地內則在 19.15 ± 2.37 ~ 36.63 ± 5.20 之間。整體而 言,產卵微棲地內兩種馬兜鈴食草之葉片數皆有高於未產卵微棲地之趨勢 (Fig. 11)。
2009 年 3 月至 2010 年 3 月,各海拔樣帶產卵微棲地內蜂窩馬兜鈴葉片數 皆顯著高於未產卵微棲地 (450 m ~ 500 m:Z = 2.3682,P = 0.0089。550 m:Z
= 4.3481,P < 0.0001。600 m ~ 650 m:Z = 5.1171,P < 0.0001。660 m:Z = 5.4783,
P < 0.0001。700 m ~ 750 m:Z = 2.4820,P = 0.0065) (Fig. 11a)。
海拔 450 m ~ 550 m、550 m 及 700 m ~ 750 m 三樣帶,產卵微棲地內掌葉 馬兜鈴葉片數顯著高於未產卵微棲地 (450 m ~ 550 m:Z = 3.2153,P = 0.0007。
550 m:Z = 4.2322,P < 0.0001。700 m ~ 750 m:Z = 3.2192,P = 0.0006)。海 拔 600 m ~ 650 m 及 600 m 兩樣帶,產卵微棲地與未產卵微棲地內蜂窩馬兜鈴 葉片數沒有顯著差異 (600 m ~ 650 m:Z = 1.0292,P = 0.1517。660m:Z = 1.2309,
P = 0.1092) (Fig. 11b)。
三、微棲地內植被覆蓋率及非食草植物物種數對大紅紋鳳蝶產卵選擇 之可能影響
(一) 植被覆蓋率在產卵食草微棲地與未產卵食草微棲地間之比較
大紅紋鳳蝶 2 m × 2 m 產卵食草微棲地與未產卵食草微棲地內,植被覆 蓋率沒有顯著差異。顯示植被覆蓋率並不妨礙大紅紋雌蝶對食草的蒐尋。產 卵蜂窩馬兜鈴食草微棲地內之植被覆蓋率在 49.53 ± 2.36% ~ 61.43 ± 6.83%
(Mean ± SE) 間,未產卵蜂窩馬兜鈴食草微棲地內則在 41.67 ± 8.21% ~ 54.85
± 4.40% 之間。產卵掌葉馬兜鈴食草微棲地內之植被覆蓋率在 34.17 ± 5.46%
~ 63.72 ± 3.65% 間,未產卵掌葉馬兜鈴食草微棲地內則在 30.91 ± 4.99% ~ 59.62 ± 3.58% 間 (Fig. 12)。
2009 年 3 月至 2010 年 3 月,里龍山各海拔樣帶大紅紋鳳蝶產卵蜂窩馬兜 鈴微棲地與未產卵蜂窩馬兜鈴微棲地內,植被覆蓋率皆沒有顯著差異 (450 m
~ 500 m:Z = -0.2903,P = 0.7716。550 m:Z = 0.7786,P = 0.2181。600 m ~ 650 m:Z = -0.4426,P = 0.6581。660 m:Z = 1.4622,P = 0.0718。700 m ~ 750 m:
Z = 0.4097,P = 0.3410) (Fig. 12a)。
各海拔樣帶大紅紋鳳蝶產卵掌葉馬兜鈴微棲地與未產卵掌葉馬兜鈴微棲 地內,植被覆蓋率亦沒有顯著差異 (450 m ~ 500 m:Z = -1.7528,P = 0.0796。
550 m:Z = 0.7248,P = 0.2343。600 m ~ 650 m:Z = -0.4795,P = 0.6316。660 m:Z = 1.3319,P = 0.0914。700 m ~ 750 m:Z = 0.1439,P = 0.4428) (Fig. 12b)。
(二) 非食草植物物種數在產卵食草微棲地與未產卵食草微棲地間之比較
大紅紋鳳蝶 2 m × 2 m 產卵食草微棲地內,非食草植物物種數顯著高於 未產卵食草微棲地,或沒有差異。顯示非食草植物物種數並不妨礙大紅紋雌 蝶對食草的蒐尋。產卵蜂窩馬兜鈴食草微棲地內之非食草植物物種數在 8.00 ± 0.79 ~ 11.06 ± 1.07 (Mean ± SE) 間,未產卵蜂窩馬兜鈴食草微棲地內則在 6.83 ± 0.36 ~ 8.85 ± 0.41 間。產卵掌葉馬兜鈴食草微棲地內之非食草植物物 種數在 6.00 ± 1.03 ~ 9.81 ± 0.40 間,未產卵掌葉馬兜鈴食草微棲地內則在 6.42 ± 0.54 ~ 7.97 ± 0.32 間。整體而言,產卵食草微棲地內非食草植物物種 數皆有高於未產卵食草微棲地之趨勢 (Fig. 13)。
2009 年 9 月至 2010 年 3 月,海拔 450 m ~ 500 m、600 m ~ 650 m 及 660 m 三樣帶,產卵蜂窩馬兜鈴食草微棲地內,非食草植物物種數顯著高於未產卵 蜂窩馬兜鈴食草微棲地 (450 m ~ 500 m:Z = 2.7211,P = 0.0033。600 m ~ 650 m:Z = 5.1133,P < 0.0001。660 m:Z = 2.5316,P = 0.0057)。海拔 550 m 及 700 m ~ 750 m 兩樣帶,產卵蜂窩馬兜鈴食草微棲地與未產卵蜂窩馬兜鈴食草 微棲地內非食草植物物種數沒有顯著差異 (550 m:Z = 1.5679,P = 0.0585。
700 m ~ 750 m:Z = 0.8350,P = 0.2229) (Fig. 13a)。
海拔 550 m、600 m ~ 650 m、660 m 及 700 m ~ 750 m 四樣帶,產卵掌葉 馬兜鈴食草微棲地內,非食草植物物種數顯著高於未產卵長葉馬兜鈴食草微 棲地 (550 m:Z = 3.2770,P = 0.0005。600 m ~ 650 m:Z = 1.9051,P = 0.0284。
660 m:Z = 2.3963,P = 0.0100。700 m ~ 750 m:Z = 2.9681,P = 0.0015)。海 拔 450 m ~ 500 m 樣帶,產卵掌葉馬兜鈴食草微棲地與未產卵掌葉馬兜鈴食草 微棲地內,非食草植物物種數沒有顯著差異 (Z = 0.0000,P = 0.5) (Fig. 13b)。
四、產卵微棲地中卵蟲數與食草資源量或植被組成等變數之相關性
(一) 產卵微棲地內之卵蟲數與兩種馬兜鈴食草植株數之相關性
2009 年 9 月至 2010 年 3 月,大紅紋鳳蝶 2 m × 2 m 產卵微棲地內,兩種 馬兜鈴食草植株數與卵蟲數皆沒有顯著線性關係 (蜂窩馬兜鈴:N = 73,rs = 0.1231,P = 0.2994。掌葉馬兜鈴:N = 21,rs = 0.2633,P = 0.2489) (Fig. 14)。
不論兩種食草植株數的多寡,在每個產卵蜂窩馬兜鈴微棲地內卵蟲數在 1 至 3 筆之間,其中兩個微棲地卵蟲數高達 7 筆。每個產卵掌葉馬兜鈴微棲地 內卵蟲數在 1 至 2 筆之間,其中一個微棲地卵蟲數有 4 筆,一個則高達 8 筆。
在移除離群值後分析,蜂窩馬兜鈴之植株數與卵蟲數仍無顯著線性相關 (N = 71,rs = 0.0911,P = 0.4501)。掌葉馬兜鈴之植株數與卵蟲數則呈現顯著線性 正相關 (N = 20,rs = 0.5137,P = 0.0205)。
(二) 產卵微棲地內之卵蟲數與兩種馬兜鈴食草葉片數多寡之相關性
2009 年 9 月至 2010 年 3 月,大紅紋鳳蝶 2 m × 2 m 產卵微棲地內,蜂窩 馬兜鈴食草葉片數與卵蟲數有顯著線性正關係 (N = 73,rs = 0.2950,P = 0.0113) (Fig. 15a)。掌葉馬兜鈴之葉片數與卵蟲數無顯著線性相關 (N = 21,rs
= 0.1735,P = 0.4519) (Fig. 15b)。
同 (一) 在移除離群值後分析,蜂窩馬兜鈴葉片數與卵蟲數仍有顯著線性 正相關 (N = 71,rs = 0.2608,P = 0.0281);掌葉馬兜鈴葉片數與卵蟲數仍無顯 著線性相關 (N = 20,rs = 0.3928,P = 0.0867)。
(三) 產卵微棲地內之卵蟲數與兩種馬兜鈴食草高度之相關性
2009 年 9 月至 2010 年 3 月,大紅紋鳳蝶 2 m × 2 m 產卵微棲地內,兩種 馬兜鈴高度與卵蟲數皆有顯著線性正相關 (蜂窩馬兜鈴:N = 73,rs = 0.2610,
P = 0.0257。掌葉馬兜鈴:N = 21,rs = 0.44602,P = 0.0427) (Fig. 16)。
同 (一) 移除離群值後分析,兩種馬兜鈴高度與卵蟲數皆無顯著線性相關 (蜂窩馬兜鈴:N = 71,rs = 0.2126,P = 0.0751。掌葉馬兜鈴:N = 20,rs = 0.3873,
P = 0.0916)。
(四) 產卵微棲地內之卵蟲數與植被覆蓋率之相關性
2009 年 9 月至 2010 年 3 月,大紅紋鳳蝶兩種產卵食草微棲地內,其植被 覆蓋率與卵蟲數皆無顯著線性相關 (產卵蜂窩馬兜鈴食草微棲地:N = 73,rs = -0.0688,P = 0.5629。產卵掌葉馬兜鈴食草微棲地:N = 21,rs = 0.0217,P = 0.9255) (Fig. 17)。
同 (一) 移除離群值後分析,兩種產卵食草微棲地內,其植被覆蓋率與卵 蟲數仍皆無顯著線性相關 (產卵蜂窩馬兜鈴食草微棲地:N = 71,rs = -0.0668,
P = 0.5798。產卵掌葉馬兜鈴食草微棲地:N = 20,rs = 0.13158,P = 0.5803)。
(五) 產卵微棲地內之卵蟲數與非食草植物物種數之相關性
2009 年 9 月至 2010 年 3 月,大紅紋鳳蝶兩種產卵食草微棲地內,其非食 草植物物種數與卵蟲數皆無顯著線性相關 (產卵蜂窩馬兜鈴食草微棲地:N = 73,rs = -0.0405,P = 0.7335。產卵掌葉馬兜鈴食草微棲地:N = 21,rs = -0.2180,
P = 0.3424) (Fig. 18)。
同 (一) 移除離群值後分析,兩種產卵食草微棲地內,其非食草植物物種 數與卵蟲數亦無顯著線性相關 (產卵蜂窩馬兜鈴食草微棲地:N = 71,rs = -0.0328,P = 0.7862。產卵掌葉馬兜鈴食草微棲地:N = 20,rs = -0.1849,P = 0.4352)。
五、產卵微棲地內之食草資源量對大紅紋蝶卵存活狀態之可能影響
(一) 產卵微棲地內兩種馬兜鈴植株數對蝶卵孵化、寄生或消失之可能影響
大紅紋鳳蝶蝶卵在兩種馬兜鈴不同植株數分組間,其孵化、消失與寄生 三種狀態頻度比例皆沒有顯著差異 (蜂窩馬兜鈴:G = 13.8786,df = 3,P = 0.3085。掌葉馬兜鈴:G = 4.1508,df = 3,P = 0.6563)。整體而言,無法看出 兩種食草在不同植株數分組間對蝶卵狀態的影響 (Fig. 19)。
在蜂窩馬兜鈴上,蝶卵寄生比例為 40.54 ~ 61.22%,消失比例為 27.27 ~ 34.69%,孵化比例為 4.08 ~ 29.73%。植株數 3 ~ 4 及 5 ~ 6 株分組下,寄生比 例顯著高於消失與孵化比例 (3 ~ 4 株:N = 44,G = 9.1246,df = 2,P = 0.0052;
5 ~ 6 株:N = 49,G = 29.4393,df = 2,P < 0.0001),其他植株分組下,蝶卵 三種狀態比例沒有顯著差異 (1 ~ 2 株:N = 37,G = 0.8383,df = 2,P = 0.3288;
7 ~ 10 株:N = 25,G = 1.2260,df = 2,P = 0.2709)。
在掌葉馬兜鈴上,寄生比例為 37.50 ~ 50.00%,消失比例為 28.95 ~ 50.00%,
孵化比例為 10.00% ~ 17.74%。植株數 1 ~ 5 及 6 ~ 10 株分組下,寄生與消失 比例顯著高於孵化比例 (1 ~ 5 株:N = 62,G = 7.5818,df = 2,P = 0.0113;6
~ 10 株:N = 38,G = 11.8919,df = 2,P = 0.0013),其他植株分組下,蝶卵 三種狀態比例沒有顯著差異 (11 ~ 15 株:N = 16,G = 3.9775,df = 2,P = 0.0684;
16 ~ 28 株:N = 10,G = 3.1053,df = 2,P = 0.1058)。
(二) 產卵微棲地內兩種馬兜鈴葉片數對蝶卵孵化、寄生或消失之可能影響
大紅紋鳳蝶蝶卵在兩種馬兜鈴不同葉片數分組間,其孵化、消失與寄生 三種狀態頻度比例皆有顯著差異 (蜂窩馬兜鈴:G = 13.6601,df = 3,P = 0.0337。
掌葉馬兜鈴:G = 49.3347,df = 3,P < 0.0001) (Fig. 20)。
在蜂窩馬兜鈴上,蝶卵寄生比例相當高,為 36.36% ~ 71.11%,消失比例 為 20 ~ 36.36%,孵化比例最低,為 8.89 ~ 30%。整體而言,葉片數較多之分 組,寄生比例顯著高於消失及孵化比例 (Fig. 20a)。4 組植株數分組之蝶卵狀 態在葉片數 1 ~ 30 片組、31 ~ 60 片組,蝶卵三種狀態比例沒有顯著差異 (1 ~ 30 片:N = 22,G = 0.3763,df = 2,P = 0.4142;31 ~ 60 片:N = 30,G = 0.2003,
df = 2,P = 0.4523)。葉片數 61~90 片組、> 90 片組,蝶卵三種狀態比例有顯 著差異 (61 ~ 90 片:N = 45,G = 28.7230,df = 2,P < 0.0001;> 90 片:N = 58,
G = 11.1101,df = 2,P = 0.0019)。
在掌葉馬兜鈴上,蝶卵寄生比例為 32.81 ~ 66.67%,消失比例為 3.70 ~ 64.06%,孵化比例最低,為 3.13 ~ 37.04%。整體而言,隨葉片數的增加,寄 生比例似乎沒有太大的變化,消失比例則隨葉片數的增加而提升,孵化比例 則隨葉片數的增加而減少 (Fig. 20b)。4 組葉片數分組之蝶卵狀態比例皆有顯 著差異 (1 ~ 30 片:N = 27,G = 16.1244,df = 2,P = 0.0002;31 ~ 60 片:N = 23,G = 5.5836,df = 2,P = 0.0306;61 ~ 90 片:N = 12,G = 6.5917,df = 2,
P = 0.0185;> 90 片:N = 64,G = 43.4408,df = 2,P < 0.0001)。
六、產卵微棲地內之植被覆蓋率及非食草植物物種數對大紅紋蝶卵存 活狀態之可能影響
(一) 植被覆蓋率對蝶卵孵化、消失與寄生之可能影響
大紅紋鳳蝶之蝶卵在兩種食草產卵微棲地內,不同植被覆蓋率分組間,
其孵化、消失與寄生三種狀態頻度比例沒有顯著差異 (蜂窩馬兜鈴產卵微棲地:
G = 9.9130,df = 4,P = 0.2712;掌葉馬兜鈴產卵微棲地:G = 10.6427,df = 4,
P = 0.2228)。整體而言,無法看出植被不同覆蓋率分組間對蝶卵狀態的影響 (Fig. 21)。
於蜂窩馬兜鈴產卵微棲地內,不同植被覆蓋率分組間,蝶卵寄生比例為 42.31 ~ 64.71%,消失比例為 23.08 ~ 46.15%,孵化比例最低,為 11.54 ~ 26.92%。
除了植被覆蓋率 80 ~ 100% 組,蝶卵三種狀態比例沒有顯著差異 (N = 21,G
= 1.1589,df = 2,P = 0.0.2801),在不同植被覆蓋率分組下,寄生與消失比例 顯著高於孵化比例 (0 ~ 20%:N = 26,G = 6.6899,df = 2,P = 0.0176;20 ~ 40%:
N = 52,G = 6.2728,df = 2,P = 0.0217;40 ~ 60%:N = 39,G = 14.8948,df
= 2,P = 0.0003;60 ~ 80%:N = 17,G = 7.4602,df = 2,P = 0.0110)。
於掌葉馬兜鈴產卵微棲地內,不同植被覆蓋率分組間,蝶卵寄生比例為 32.14 ~ 55.00%,消失比例為 36.00 ~ 44.44%,孵化比例為 5.00 ~ 32.14%。除 了植被覆蓋率 20 ~ 40% 及 60 ~ 80% 兩分組,寄生與消失比例顯著高於孵化 比例外 (20 ~ 40%:N = 40,G = 20.2799,df = 2,P < 0.0001;60 ~ 80%:N = 25,G = 6.8172,df = 2,P = 0.0165),在不同植被覆蓋率分組下,蝶卵三種狀 態比例皆沒有顯著差異 (0 ~ 20%:N = 14,G = 2.6424,df = 2,P = 0.1334;
40 ~ 60%:N = 28,G = 0.0706,df = 2,P = 0.4827;80 ~ 100%:N = 18,G = 2.6021,df = 2,P = 0.1361)。
(二) 非食草植物物種數對蝶卵孵化、消失與寄生之可能影響。
大紅紋鳳蝶之蝶卵在兩種食草產卵微棲地內,不同非食草植物物種數分 組間,其孵化、消失與寄生三種狀態頻度比例沒有顯著差異 (蜂窩馬兜鈴產卵 微棲地:G = 5.1996,df = 3,P = 0.5185;掌葉馬兜鈴產卵微棲地:G = 6.5499,
df = 2,P = 0.1617),整體而言,無法看出不同非食草植物物種數分組間對蝶 卵狀態的影響 (Fig. 22)。
於蜂窩馬兜鈴產卵微棲地內,不同非食草植物物種數分組間,蝶卵寄生 比例為 38.89 ~ 50.00%,消失比例為 23.68 ~ 35.29%,孵化比例為 13.16 ~ 27.78%。除了非食草植物物種數 9 ~ 10 種組,寄生比例顯著高於消失及孵化 比例 (N = 38,G = 15.2290,df = 2,P = 0.0002),在不同非食草植物物種數分 組下,蝶卵三種狀態比例沒有顯著差異 (3 ~ 6 種:N = 18,G = 0.3349,df = 2,
P = 0.4229;7 ~ 8 種:N = 34,G = 1.7155,df = 2,P = 0.2121;11 ~ 19 種:N
= 20,G = 2.3557,df = 2,P = 0.1540)。
於掌葉馬兜鈴產卵微棲地內,不同非食草植物物種數分組間,蝶卵寄生 比例為 30.43 ~ 50.00%,消失比例為 6.67 ~ 34.78%,孵化比例為 26.67 ~ 34.78%。
除了非食草植物物種數 9 ~ 10 種組,寄生比例顯著高於消失與孵化比例外 (N
= 15,G = 8.8590,df = 2,P = 0.0060),在不同非食草植物物種數分組下,蝶 卵三種狀態比例皆沒有顯著差異 (7 ~ 8 種:N = 23,G = 0.0883,df = 2,P = 0.4784;11 ~ 14:N = 6,G = 1.0465,df = 2,P = 0.2963)。
肆、討論
一、食草植株的分布及密度對大紅紋鳳蝶產卵選擇之可能影響
許多研究顯示,植食性昆蟲的產卵選擇會受到食草植物分布、物種或植株性 狀的影響,例如對不同物種食草植物的偏好,或同種食草植物在植株數、葉片數 或葉片品質上的影響 (Papaj and Rausher, 1987; Janz and Nylin, 1997; Webb and Pullin, 2000; Dennis et al., 2003; Bergström et al. 2006)。本研究結果顯示,在里龍山 不同海拔樣帶,大紅紋鳳蝶產於兩種馬兜鈴上之卵蟲數,大致上與兩種馬兜鈴在 各海拔樣帶之數量分布一致,而且與兩種馬兜鈴在單位面積內植株密度及葉片密 度有顯著線性正相關。表示大紅紋鳳蝶產卵時期,會選擇馬兜鈴食草資源量分布 較多之海拔棲地進行產卵活動。
但是,在海拔 700 m ~ 750 m 樣帶,雖生長數量較多的掌葉馬兜鈴,但全年發 現的卵蟲數量相對較少,此可能與此海拔掌葉馬兜鈴植株的大小有關。里龍山海 拔 700 m 以上,比起其他海拔較低之樣帶,屬於濕度偏高之環境 (Fig. 2 ~ 3),而 掌葉馬兜鈴較傾向於乾燥向陽處生長 (楊君嵐,2007),故此海拔之掌葉馬兜鈴多 以貼地叢生的形式生長,植株較小且葉片量少,葉片品質大多屬於較老熟之葉片,
可能較不利於此蝶對食草植物之蒐尋。這種終年常高濕且多霧的環境條件,也較 易使蝴蝶雙翅潮濕且降低其能見度,而不利於雌蝶在本區的飛行與食草蒐尋。且 雌蝶的產卵活動,可能因本區為稜線地形,較易受夏季颱風或冬季東北季風之干 擾,而多至較低海拔或風速較低之河谷進行產卵 (Fig. 4)。
二、微棲地內食草資源量對大紅紋鳳蝶產卵選擇之可能影響
本研究顯示,大紅紋鳳蝶會選擇有較豐富馬兜鈴植株數與葉片數,或馬兜鈴 食草資源與鄰近未產卵微棲地沒有顯著差異的微棲地進行產卵,可知微棲地中兩 種馬兜鈴植株與葉片數量的充足與否,對大紅紋鳳蝶的產卵微棲地選擇有重要的 影響。馬兜鈴食草較豐富之微棲地中,大量的馬兜鈴植株或葉片,可能散發濃度 較高的馬兜鈴酸,且雌蝶對馬兜鈴酸的敏感度高於雄蝶,此將會提高雌蝶被吸引 前來產卵之作用 (李宗憲,2008;賴玉珊,2009;Pinto et al., 2009)。雌蝶可藉由 對微棲地的選擇來降低子代間的競爭並提升子代的存活 (William and Resetarfts, 1996; Anthes et al., 2003),食草豐富的微棲地可確保帅蟲的食物來源,提升其存活 率 (葉人碩,2010),亦並可避免帅蟲在尋找食草途中可能遭遇的掠食風險 (Anthes et al., 2003)。
里龍山微棲地中常生長分布不一的大小馬兜鈴植株,本研究產卵微棲地中的 大紅紋鳳蝶卵蟲數分別與蜂窩馬兜鈴葉片數及掌葉馬兜鈴植株數有顯著的線性正 相關。此代表大紅紋鳳蝶可能隨著微棲地中蜂窩馬兜鈴葉片數或掌葉馬兜鈴植株 數的增加,而產下較多的蝶卵,但實際上,每一個產卵微棲地中不論植株數或葉 片數的多寡,發現的卵蟲數最多皆不超過 3 至 4 個。因此大紅紋鳳蝶在產卵微棲 地內每次的產卵數,會因產卵微棲地內馬兜鈴植株數或葉片數多寡而產生有限度 的影響,但是卻不會造成其在同一微棲地中產下過多的蝶卵。部分蝶種會辨識同 類的卵粒或帅蟲,並避免在同株食草植物上重覆產下蝶卵 (Rausher, 1979),甚至有 評估食草葉片多寡來分配卵粒數量的可能 (Vasconcellos-Neto and Monteiro, 1993)。
大紅紋鳳蝶這種較分散產卵的型態,不僅使該處食草植物資源足夠支持其子代生 長,且同時可避免子代對食草資源的種內競爭 (intra-specific competition)。
產卵微棲地中大紅紋鳳蝶之卵蟲數與兩種馬兜鈴之高度,在移除離群植前有 顯著線性正相關,移除離群值後之分析則沒有顯著線性相關,由於群值常出現在 食草較高之植株上,此可能代表攀附高度愈高之兩種馬兜鈴植株,較易吸引雌蝶 的注意前來產卵,但也沒有造成雌蝶在此植株上產下過多蝶卵的現象,因此更進 一步印證大紅紋鳳蝶屬於分散產卵的型態。對照本區大紅紋蝶卵被寄生的比例高 達 50%,此種雌蝶分散產卵的型態是一個必要的發展策略。
三、微棲地內植被覆蓋率及非食草植物物種數對大紅紋鳳蝶產卵選擇 之可能影響
雌蝶需要敏銳的食草感測及蒐尋能力,以發現隱蔽的食草植物,在茂盛植被 中生長的網葉馬兜鈴 (Aristolochia reticulate Jacquin),會減低美洲藍鳳蝶 (Battus philenor L.) 及其帅蟲蒐尋網葉馬兜鈴的敏感度 (Rausher, 1981)。因此,野外植食 性昆蟲之食草植物,若生長在一植被茂盛的微棲地中,被植食性昆蟲利用的機會 通常比一般獨自生長或單一種植的食草植物低,此可能導致部分蝶種選擇在低植 被覆蓋處進行產卵 (謝桂禎,2009;Anthes et al., 2003; Eichel and Fartmann, 2008;
Wiklund and Friberg, 2008),解釋這樣現象的可能原因包括:(1) 複雜的植被環境可 能對植食性昆蟲蒐尋食草植物的過程造成物理性或化學性阻礙 (Nylin and Janz, 1993) ; (2) 茂 密 的 植 被 可 能 阻 礙 食 草 植 物 或 蜜 源 植 物 的 生 長 (Hatada and Matsumoto, 2008);(3) 茂密的植被可能提高植食性昆蟲被掠食的風險 (Wiklund and Friberg, 2008)。
本研究結果顯示,里龍山各海拔樣帶之大紅紋鳳蝶產卵微棲地與未產卵微棲 地間的植被覆蓋率皆沒有顯著差異,但兩種馬兜鈴植株數或葉片數,在產卵微棲 地內皆分別顯著高於未產卵微棲地,或沒有顯著差異,因此無法得知在大紅紋鳳
蝶產卵時,是否同時受到馬兜鈴食草與植被覆蓋率之影響,或是單獨受到植被覆 蓋率本身的影響。
在生長兩種馬兜鈴之大紅紋鳳蝶產卵微棲地內,卵蟲數與植被覆蓋率沒有顯 著線性相關,無論在植被覆蓋率高或低的地方,都發現數量相當的卵蟲數。此顯 示植被覆蓋率的高低並不會使大紅紋鳳蝶對兩種馬兜鈴的蒐尋產生妨礙,不論植 被覆蓋率的高低,皆可發現卵蟲。里龍山馬兜鈴植物為具有攀附性的草質藤本植 物,可攀附附近林下植物,穿梭於其空隙間,並在攀負植物頂部開展生長葉片,
可能由於此特性使雌蝶較容易在茂密的植被中發現馬兜鈴植株並進行產卵。
複雜的植被環境可能造成雌蝶蒐尋食草植物時的化學性阻礙,但仍有部分蝶 種在產卵點與對照點中的植物多樣性沒有顯著差異 (Kopper, 2000)。本研究結果顯 示,大紅紋鳳蝶產卵微棲地中的非食草植物物種數顯著高於未產卵微棲地,或是 沒有顯著差異;產卵微棲地內卵蟲數與非食草植物物種數也不具顯著線性相關。
代表非食草植物物種的多寡,不降低或妨礙大紅紋鳳蝶尋找食草與產卵,且不因 微棲地中非食草植物物種數的多寡而影響大紅紋卵蟲的數量,因而降低複雜植被 對大紅紋雌蝶蒐尋本區馬兜鈴食草植物時形成化學性阻礙的可能。
四、產卵微棲地內之食草資源量對大紅紋蝶卵存活狀態之可能影響
卵期是所有昆蟲發育過程中防禦力最低的階段,雖仍有防禦掠食性天敵的機 制 (Sime et al., 2000),但蟲卵被寄生之情形,卻是多數植食性昆蟲蟲卵死亡的因素 (Hawkins et al., 1997),並可能導致族群的減少 (Smith, 1996)。野外調查瀕危蝶種 牙買加鳳蝶 (Papilio (Pterourus) homerus Fabricius) 之蝶卵死亡率為 74.4%,其中 蝶卵被寄生性天敵所寄生的比例高達 65.7% (Garraway et al., 2008)。本研究顯示,
在里龍山大紅紋鳳蝶蝶卵被寄生之比例達 50%,因此蝶卵被寄生蜂的寄生作用 (Pic. 4),可能是最大的致死因素。卵專一性寄生性天敵,通常會先蒐尋寄主之食 草植物的揮發物,再進一步蒐尋寄主分泌物之氣味因而找到寄主進行寄生,因此,
食草植物本身也成為寄生性天敵蒐尋寄主時的最佳線索 (Hilker and Meiners, 2006;
Randlkofer et al., 2007)。由本研究結果可知,大紅紋鳳蝶偏好將蝶卵分別產於兩種 馬兜鈴植物較豐富之微棲地,長久來看,雖可確保其帅蟲的食物來源,卻可能相 對提升蝶卵被卵專一性寄生性天敵所寄生之風險。故雌蝶選擇產卵微棲地時,如 何在避免蝶卵被寄生及確保帅蟲食物來源間進行取捨,便成為一個決定子代存活 的重要關鍵 (Thompson, 1988)。
本研究結果顯示,在生長蜂窩馬兜鈴的產卵微棲地內,食草葉片量較多的產 卵微棲地,蝶卵被寄生的比例高於食草葉片量較少的產卵微棲地,孵化比例也比 食草葉片量較少的微棲地低,失蹤比例似乎沒有太大的差異,證實本研究區內食 草葉片量的多寡可能是寄生蜂蒐尋大紅紋蝶卵的關鍵。而生長掌葉馬兜鈴之產卵 微棲地,不論葉片量多寡,蝶卵被寄生比例似乎沒有太大的差異,但孵化比例卻 有下降的趨勢,造成原因可能是失蹤比例相對的提升,但野外失蹤的蝶卵狀態已 無法判定,有孵化、遭掠食、被雨水打落或其他不確定因素造成蝶卵遺失之可能,
故蝶卵產於生長掌葉馬兜鈴之微棲地中,失蹤比例隨葉片增加而上升之趨勢有待 更進一步的確認與探討。
先前討論提及,大紅紋鳳蝶雌蝶的產卵表現,其卵蟲數與產卵微棲地中的兩 種馬兜鈴食草植株或葉片數量沒有顯著線性相關。這樣分散產卵的表現,或許能 有效降低蝶卵在同處被卵專一性寄生性天敵發現並寄生的風險。
除了蝶卵被卵寄生蜂寄生之機制與食草植物揮發物有關之外,蝴蝶帅蟲之寄 生性天敵蒐尋寄主的過程也與食草植物揮發物有關 (Sime, 2002; Roßbach et al., 2005)。在本研究區中,亦觀察到同為金鳳蝶族之紅紋鳳蝶 (Pachliopta aristolochiae interpositus Fruhstorfer) 與台灣麝香鳳蝶 (Byasa impediens febanus Fruhstorfer) 之 帅蟲,被寄生性天敵小繭蜂所寄生之現象 (Pic. 5)。因此,食草資源量雖是子代帅 蟲的生長存活要素,同時卻又是促成蝶卵及蝴蝶帅蟲被寄生致死的因素,這種困 境更加突顯雌蝶對產卵微棲地條件選擇的重要性。
五、產卵微棲地內之植被覆蓋率及非食草植物物種數對大紅紋蝶卵存 活狀態之可能影響
曾有研究表示,部份植食性昆蟲將卵產於植物多樣性較高的環境中,環境中 的多樣性植被將形成一種化學訊息保護所 (infochemical shelter),多樣的化學揮發 訊息有助於降低卵粒被卵專一性寄生性天敵攻擊的可能 (Randlkofer et al., 2007)。
本研究結果顯示,在不同植被覆蓋率與非食草植物物種數下,蝶卵在孵化、消失 與寄生等狀態的比例沒有顯著差異。蝶卵孵化比例沒有因為植被覆蓋率或非食草 植物物種數的增加而有所提升,寄生與消失比例在不同植被覆蓋率或非食草植物 物種數分組間也相當高,因此,影響大紅紋鳳蝶之蝶卵存活狀態的主要關鍵,可 能還是與食草植物的物種或與其葉片的數量有關。
本研究同時進行的相關研究顯示,大紅紋鳳蝶產卵位置的不同亦會影響蝶卵 的存活狀態,蝶卵若產於蜂窩馬兜鈴食草或其鄰近非食草部位上 (Pic. 6),蝶卵孵 化或被寄生的比例沒有顯著差異;而蝶卵產於掌葉馬兜鈴食草或其鄰近非食草部 位上,蝶卵被寄生之比例在食草部位與其鄰近非食草部位間有顯著差異,鄰近非 食草部位的蝶卵孵化比例高於食草部位 (蔡孙庭,2010)。此表示寄生蜂可能較不 易蒐尋產於非食草部位上的蝶卵,進而使鄰近非食草部位上的蝶卵有較高的孵化 比例。本論文結果顯示非食草植物物種數對大紅紋鳳蝶的產卵並無明顯影響,但 蝶卵產於接近食草附近之非食草部位,將可提高蝶卵的存活發育,進而可能形成 一種優勢的生存發展策略。
伍、結論
在里龍山各海拔樣帶兩種馬兜鈴上發現之大紅紋鳳蝶卵蟲數,與兩種馬兜鈴 植株的海拔分布一致。產卵微棲地中兩種馬兜鈴食草資源量皆分別高於未產卵微 棲地,但微棲地中食草資源量多寡或植被組成變化,基本上不會影響大紅紋雌蝶 的分散產卵型態,且微棲地中非食草植物物種數並不影響雌蝶尋找兩種馬兜鈴食 草與產卵。野外蝶卵被寄生比例達 50%,而食草葉片量高的微棲地之蝶卵被寄生 比例更高於葉片量少的。微棲地中植被組成的差異不影響蝶卵的存活狀況。對大 紅紋雌蝶產卵微棲地選擇之決定因素在於兩種馬兜鈴食草的可利用性,而食草葉 片量則會影響蝶卵存活的機會。大紅紋雌蝶分散產卵型態可降低蝶卵在同處被大 量寄生的風險,也可確保其帅蟲在生長微棲地中有較足夠的食草可取食利用。