Anti-oxidative and anti-inflammatory protection of three triterpenic acids in
normal human bronchial epithelial cells
Shih-ming Tsaoø,§, Mei-chin Yin†,,*
øInstitute of Medicine, Chung Shan Medical University, Taichung City, Taiwan
§Sections of Infectious Diseases and Chest Medicine, Department of Internal Medicine, Chung Shan Medical University Hospital, Taichung City, Taiwan
†Department of Health and Nutrition Biotechnology, Asia University, Taichung City, Taiwan
Department of Nutrition, China Medical University, Taichung City, Taiwan
Running title: protective effects of triterpenic acids
*To whom correspondence should be addressed: Dr. Mei-chin Yin, Professor, Department of Nutrition, China Medical University, 91, Hsueh-shih Rd., Taichung City, Taiwan TEL: 886-4-22053366 ext. 7510, FAX: 886-4-22062891
Email: [email protected] 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
ABSTRACT
Protective effects of three triterpenic acids, asiatic acid (AA), glycyrrhizic acid (GA) and oleanolic acid (OA), for two normal human bronchial epithelial cell lines, 16HBE and BEAS-2B cells, against hydrogen peroxide (H2O2) induced injury were examined. Cells were pre-treated by triterpenic acid at 4 or 8 μmol/L, and followed by H2O2 treatment. Results showed that H2O2 increased Bax and cleaved caspase-3 expression, and decreased Bcl-2 expression in two cell lines (P<0.05). AA at two doses retained Bcl-2 expression; but GA and OA only at high dose reserved Bcl-2 expression (P<0.05). Three triterpenic acids dose-dependently decreased cleaved caspase-3 expression (P<0.05). H2O2 treatment lowered mitochondrial membrane potential and Na+-K+-ATPase activity in both cell lines (P<0.05). Triterpenic acids pretreatments dose-dependently maintained mitochondrial membrane potential and Na+-K+-ATPase activity (P<0.05). H2O2 enhanced the production of reactive oxygen species (ROS), interleukin-6, tumor necrosis factor- and prostaglandin E2 in test cells (P<0.05). Three triterpenic acids treatments dose-dependently reversed these changes (P<0.05). H2O2 also up-regulated the expression of p47phox, gp91phox, cyclooxygenase (COX)-2, nuclear factor-B (NF-B) and mitogen-activated protein kinase in 16HBE and BEAS-2B cells (P<0.05). AA, GA or OA pretreatments dose-dependently down-regulated the expression of p47phox, COX-2, NF-B p65 and p-p38 (P<0.05); and only at high dose decreased gp91phox expression (P<0.05). These findings suggest that these triterpenic acids could protect normal bronchial epithelial cells to attenuate apoptotic, oxidative and inflammatory stress.
KEYWORDS: triterpenic acids; bronchial epithelial cells; ROS; NF-B; MAPK 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43
INTRODUCTION
Environmental contamination including air pollution impairs the human respiratory system. Bronchial epithelium acts as a barrier to prevent the invasion of foreign materials. Apoptosis of bronchial epithelial cells could be induced by toxins such as cigarette, particulate matter or heavy metals.1,2 It is reported that mitochondrial intrinsic apoptotic pathway, mainly mediated by BCL family, is involved in bronchial epithelial cell death.3 This BCL family consists of pro-apoptotic molecules such as Bax, and anti-apoptotic molecules such as Bcl-2. Thus, agent(s) with anti-apoptotic activity may potentially protect bronchial epithelium, and prevent the development of respiratory disorders.
Oxidative stress from excessive production of reactive oxygen species (ROS) is a crucial pathophysiological factor for the progression of chronic airway disorders including asthma and lung cancer.4,5 The activation of NADPH oxidase, an enzyme complex with several subunits such as p47phox and gp91phox, is the major ROS source in airway epithelial cells and responsible for oxidative cell death.6,7 In addition, enhanced cyclooxygenase (COX)-2 expression and prostaglandin E2 (PGE2) production in respiratory epithelial cells promote airway inflammatory stress.8,9 It has been documented that ROS could activate signaling pathways such as nuclear factor-B (NF-B) and mitogen-activated protein kinase (MAPK) in airway epithelium, which in turn elicits the transcription of genes encoded for oxidative and inflammatory factors and facilitates the progression of respiratory diseases.10,11 Thus, any agent with the capability to reduce ROS and PGE2 generation; and/or suppress NADPH oxidase, COX-2, NF-B and MAPK expression may potentially protect airway epithelial cells to attenuate oxidative and inflammatory injury.
Asiatic acid (AA), glycyrrhizic acid (GA), and oleanolic acid (OA) are pentacyclic triterpenic acids naturally occurring in many plant foods such as hawthorn fruit (Crataegi Pinnatifidae Fructus), basil (Ocimum basilicum), brown mustard (Brassica juncea) and licorice (Glycyrrhiza glabra).12-14 Yin et al.12 indicated that dietary intake of AA and OA 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69
resulted in their deposit in intact forms in organs including liver and heart. It has been documented that these triterpenic acids possess anti-oxidative and anti-inflammatory activities.15,16 Wächter et al.17 reported that some pentacyclic triterpenoids such as OA could protect respiratory system through antitubercular activity. Liu et al.18 indicated that ursolic acid, an isomer of OA, at 3.2, 6.3, 12.5 and 25 μmol/L, protected normal human bronchial epithelial cells against cigarette smoke extract induced death, and cell survival rates were in the range of 73-86%. Those previous studies suggest that OA and other triterpenic acids such as AA and GA might benefit survival and function of human bronchial epithelial cells.
Both 16HBE and BEAS-2B are normal human bronchial epithelial cell lines, and have been widely used in bronchial diseases related researches.3,18 Hydrogen peroxide, an oxidant in cigarette smoke extract, could cause apoptotic and oxidative injury in those bronchial epithelial cells.19,20 In our present study, these two cell lines were used to examine the protective effects of AA, GA and OA at two doses, 4 and 8 μmol/L, against hydrogen peroxide induced apoptotic, oxidative and inflammatory stress. The impact of these triterpenic acids upon the level, activity and/or protein expression of associated factors including NADPH oxidase, COX-2, NF-B and MAPK was evaluated.
MATERIALS AND METHODS
Materials. AA, GA and OA (95%) were purchased from Aldrich Chemical Co. (Milwaukee, WI, USA). 16HBE and BEAS-2B were obtained from American Type Culture Collection (ATCC, Rockville, MD, USA). Medium, plates, antibiotics and chemicals used for cell culture were purchased from Difco Laboratory (Detroit, MI, USA). Experimental Design and Cell Culture. Our preliminary data indicated that 12, 24, 36 and 48 hr incubation resulted in approximately 21.7, 48.6, 72.9 and 96.3% 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94
incorporation of test compound into test cells. Thus, 48 hr incubation was used for present study. In addition, the improvement upon cell viability of test compounds at 0.5, 1, 2, 4 and 8 mol/L against H2O2 was also examined in preliminary test, and results showed that these compounds at 0.5, 1 or 2 mol/L did not significantly improve cell viability. Thus, these compounds at 4 and 8 mol/L were further used. Stock solution of three triterpenic acids was prepared in dimethyl sulfoxide (DMSO) and diluted with bronchial epithelial growth medium (BEGM). The final DMSO concentration in cell culture was <0.5%. At this concentration, DMSO did not affect any measurements (data not shown). 16HBE and BEAS-2B cells were routinely cultured in BEGM, supplemented with 10% fetal calf serum, 2.5 mM L-glutamine, 100 units/mL of penicillin and 100 units/mL of streptomycin (pH 7.4) at 37°C under 95% air and 5% CO2. The culture medium was changed every three days, and cells were subcultured once a week. A phosphate buffer saline (PBS, pH 7.2) was added to adjust the cell number to 105/mL for various experiments and analyses. After aspiration, cells were treated with test compound at 4 or 8 mol/L at 37°C for 48 hr and followed by washing twice with PBS. PBS was collected and the concentration of test compound was determined by a HPLC method described in Yin et al.12 The compound left in cells was considered as incorporated one. Results showed that 48 hr incubation resulted in 97.11.3% and 95.80.9% incorporation into 16HBE and BEAS-2B cells. Then, cells were further treated by 100 M H2O2 at 37°C for 4 h. Control groups contained no test compound and without H2O2 treatment.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Assay. MTT assay was performed to examine cell viability.21 Cells (105/mL) were treated by test compound for 48 hr, then H2O2 treatment for 4 hr. MTT at 0.25 mg MTT/mL was added into medium and followed by incubating for 3 hr at 37°C. The amount of MTT formazan product was determined by measuring absorbance at 570 nm (630 nm as a reference) using 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119
a microplate reader (Bio-Rad, Hercules, CA, USA). Cell viability was expressed as a percentage of control groups.
Measurement of DNA Fragmentation. Cell death detection ELISA plus kit (Roche Molecular Biochemicals, Mannheim, Germany) was used to quantify DNA fragmentation. Cells were lysed in 5 mL cold lysis buffer containing 10 mM HEPES (pH 7.9), 1.5 mM MgCl2, 10 mM KCl, 0.1 mM EDTA, 1 mM DTT, 1 mM PMSF and 0.6% Nonidet P-40 for 30 min and followed by centrifugation at 200 xg for 10 min. Then, 20 L supernatant was transferred onto the streptavidin-coated plate, and 80 L freshly prepared immunoreagent was added to each well and incubated for 2 hr at room temperature. After washing with PBS, the substrate solution was added and incubated for 15 min. The absorbance at 405 nm (reference wavelength 490 nm) was measured using a microplate reader. DNA fragmentation was expressed as the enrichment factor using the following equation: enrichment factor = (absorbance of the sample) / (absorbance of the control), sample: cells treated with test compound or/and H2O2; control: cells without H2O2 and test compound.
Measurement of Mitochondrial Membrane Potential (MMP). MMP was monitored using a flow cytometry (Beckman-FC500, Beckman Coulter, Fullerton, CA, USA) and the fluorescent dye Rhodamine123 (Rh123) was purchased from Sigma Chemical Co. (St. Louis, MO, USA).22 Cells (105/mL) were centrifuged at 1200 xg for 5 min and re-suspended in BEGM medium. Rh123 (100 g/L) was added to medium. After 45 min incubation at 37°C, cells were collected and washed twice with PBS. The mean fluorescence intensity (MFI) in cells was analyzed with flow cytometry.
Preparation of Mitochondrial Fractions. Control or treated cells were lysed and centrifuged at 200 xg for 10 min at 4°C to obtain nuclei pellet. The supernatant from this spin was further centrifuged at 10,000 xg for 20 min at 4°C to obtain mitochondria pellet. This pellet was re-suspended by PBS. Protein concentration of mitochondrial fraction 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144
was determined by a commercial assay kit (Pierce Biotechnology Inc., Rockford, IL, USA) with bovine serum albumin as a standard.
Na+-K+-ATPase Activity Assay. Na+-K+-ATPase activity was determined by measuring the amount of inorganic phosphate (Pi) released from ATP.23 The reaction mixture contained 100 mM NaCl, 20 mM KCl, 2 mM ATP, 30 mM Tris-HCl buffer (pH 7.4) and the freshly isolated cellular mitochondria. The assay was initiated by adding ATP, and terminated by adding 15% trichloroacetic acid after 15 min incubation at 37°C. The released Pi was determined by measuring the absorbance at 640 nm. The values of the treated cells were normalized against the values of control, and expressed as a percentage of control groups.
Measurement of Caspase Activity. Activity of caspase-3 and -8 was detected by using fluorometric assay kits (Upstate, Lake Placid, NY, USA) according to the manufacturer’s protocol. The intra-assay CV was 4.2-4.9%, and the inter-assay CV was 4.5-6.1%. In brief, control or treated cells were lysed in 25 mL cold lysis buffer and incubated in ice for 10 min. Fifty μL cell lysates was mixed with 25 mL reaction buffer and 5 mL fluorogenic substrates specific for caspase-3 or -8 in a 96-well microplate. After incubation at 37°C for 1 hr, fluorescent activity was measured using a Hitachi F-4500 fluorophotometer (Tokyo, Japan) with excitation at 400 nm and emission at 505 nm. Data were expressed as a percentage of control groups.
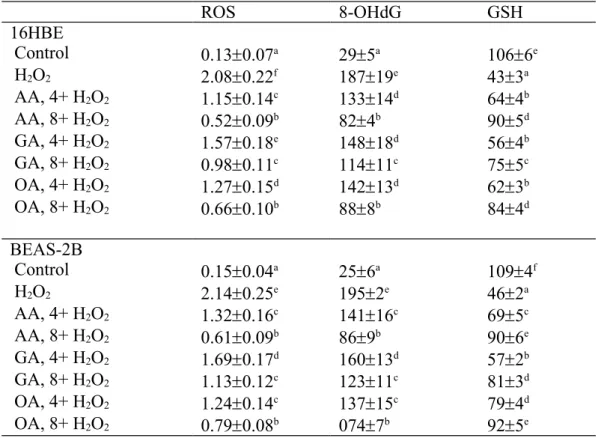
Analyses for ROS, 8-Hydroxydeoxyguanosine (8-OHdG) and Glutathione (GSH) Levels. The dye DCFH2-DA was used to measure ROS level. Briefly, cells were washed and suspended in cell medium. After incubating with 50 mol/L dye for 30 min and washing with PBS, cell suspension was centrifuged at 412 xg for 10 min. Cells were collected and dissolved with 1% Triton X-100. Fluorescence changes were measured at an excitation wavelength of 485 nm and an emission wavelength of 530 nm using a fluorescence microplate reader. Relative fluorescence unit (RFU) was the difference in 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170
fluorescence values obtained at time 0 and 5 min. Results are expressed as RFU/mg protein. 8-OHdG is an indicator of DNA oxidative stress. DNA fractions were obtained using a DNA Extractor WB kit (Wako Pure Chemical Industries Ltd., Tokyo, Japan). Extracted DNA was dissolved in water (1 mg/mL), and oxidative damage was determined using an ELISA kit for 8-OHdG (OXIS Health Products Inc., Portland, OR, USA). GSH concentration (ng/mg protein) was determined by a commercial colorimetric GSH kit (OxisResearch, Portland, OR, USA). The detection limit was 10 pg/mg protein and 10 ng/mg protein for 8-OHdG and GSH, respectively.
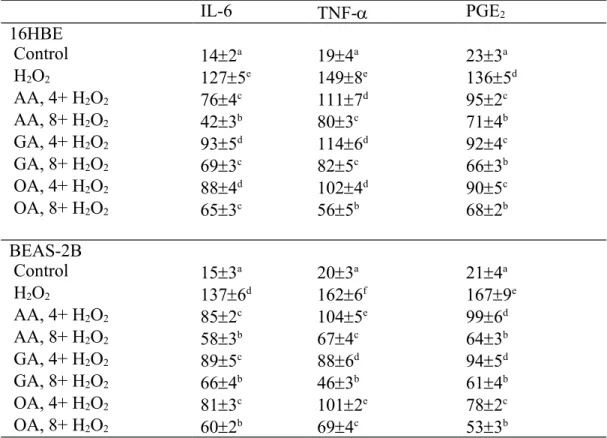
Measurements for Interleukin (IL)-6, Tumor Necrosis Factor (TNF)- and PGE2. Cells were washed and homogenized, and the released level (pg/mg protein) of IL-6
and TNF- in supernatant was measured by ELISA methods using cytoscreen immunoassay kits (BioSource International, Camarillo, CA, USA). The level (pg/mg protein) of PGE2 was determined using a PGE2 EIA kit (Cayman Chemical Co., Ann Arbor, MI, USA) according to the manufacturer's instructions. The detection limit was 5 pg/mg protein for IL-6, and 10 pg/mg protein for TNF- and PGE2.
Preparation of Cytosolic and Nuclear Fractions. Cells were suspended in ice-cold buffer containing 10 mM HEPES, pH 7.9, 1.5 mM MgCl2, 10 mM KCl, 0.5 mM DTT, 0.5 mM PMSF, 1 mg/mL of leupeptin, and 1 mg/mL of aprotinin for 15 min. Cytosolic fractions were collected after centrifugation at 14,000 xg for 1 min at 4°C. The remaining nuclear pellets were re-suspended in buffer containing 20 mM HEPES, pH 7.9, 1.5 mM MgCl2, 450 mM NaCl, 25% glycerol, 0.2 mM EDTA, 0.5 mM DTT, 0.5 mM PMSF, 1 mg/mL of leupeptin and 1 mg/mL of aprotinin for 30 min. Nuclear fractions were collected after centrifugation at 14,000 xg for 15 min at 4°C.
Western Blot Analyses. Plate was washed twice by PBS. Cells were scraped from plates, and suspended in above lysis buffer. Protein content was determined by Bio-Rad 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195
protein assay reagent (Bio-Rad Laboratories Inc. Hercules, CA, USA). Protein sample at 40 μg was applied to 10% SDS-polyacrylamide gel electrophoresis, and transferred to a nitrocellulose membrane (Millipore, Bedford, MA, USA) for 1 hr. After blocking with a solution containing 5% nonfat milk for 1 hr to prevent non-specific binding of antibody, membrane was incubated with cleaved caspase-3, Bcl-2, Bax (1:1000), anti-p47phox, anti-gp91phox, anti-COX-2, anti-NF-B p50, anti-NF-B p65 and anti-MAPK (1:2000) monoclonal antibody (Boehringer-Mannheim, Indianapolis, IN, USA) at 4ºC overnight, and followed by reacting with horseradish peroxidase-conjugated antibody for 3.5 hr at room temperature. The detected bands were quantified by an image analyzer (ATTO, Tokyo, Japan) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a loading control. The blot was quantified by densitometric analysis. Results were normalized to GAPDH.
Statistical Analysis. Shapiro-Wilk test was used to assess normality of data. The effect of each treatment was analyzed from nine different preparations (n=9). Statistical analysis was undertaken by using one-way analysis of variance, and post-hoc comparisons were carried out using Dunnett's t-test. P<0.05 level was considered to be significantly different.
RESULTS
Effects of AA, GA or OA upon cell viability.
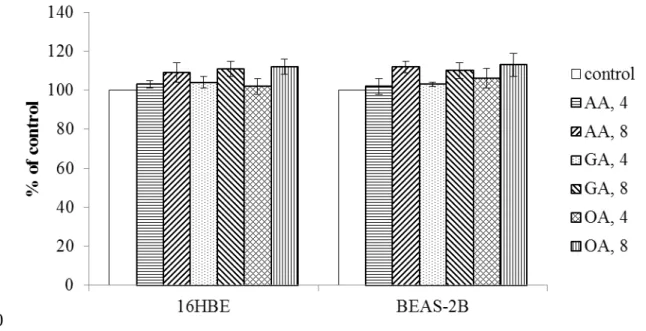
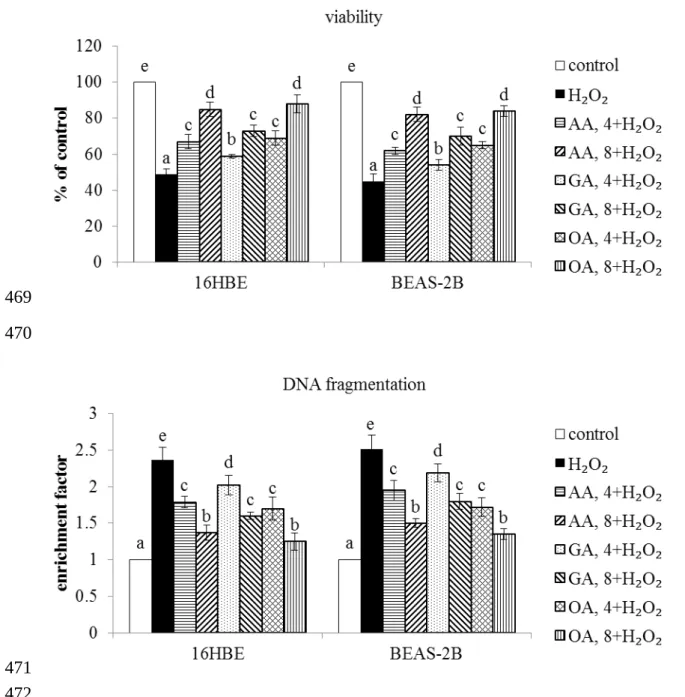
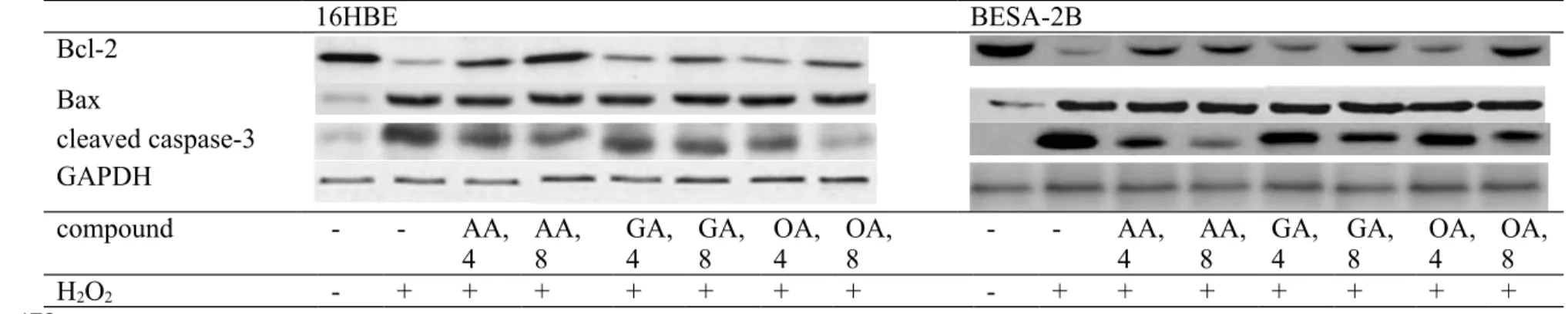
As shown in Figure 1, without H2O2 treatment, three triterpenic acids at two doses did not affect the viability of 16HBE and BEAS-2B cells (P>0.05). H2O2 treatment decreased viability and increased DNA fragmentation in test cells (Figure 2, P<0.05). However, AA, GA or OA pre-treatments enhanced cell survival and lowered DNA fragmentation in those cells (P<0.05). H2O2 treatment enhanced cleaved caspase-3 and Bax expression, and declined Bcl-2 expression in test two cell lines (Figure 3, P<0.05). AA at two doses 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221
retained Bcl-2 expression; but GA and OA only at high dose preserved Bcl-2 expression (P<0.05). AA, GA or OA dose-dependently lowered cleaved caspase-3 expression (P<0.05). Three test compounds did not affect Bax expression in test cells (P>0.05). Effects of AA, GA or OA upon mitochondrial stability.
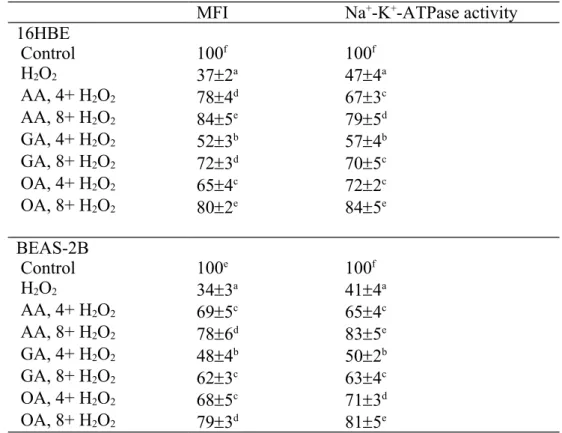
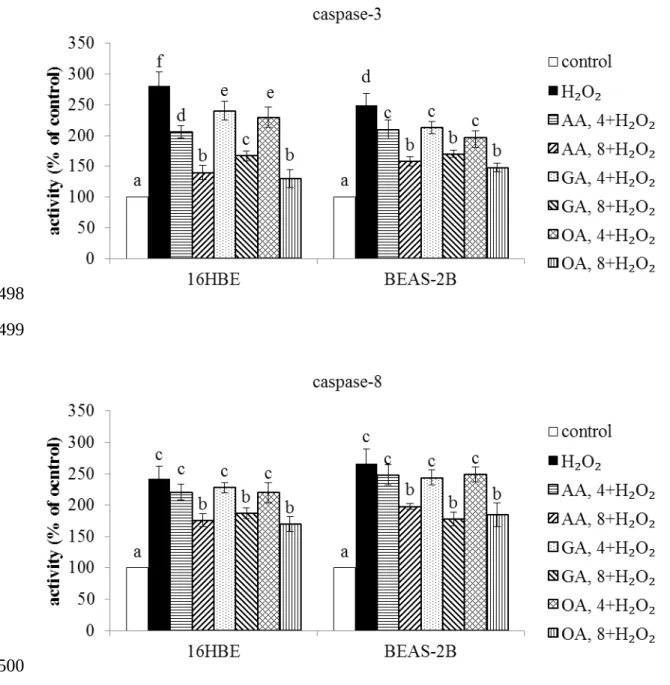
H2O2 treatment lowered MMP and Na+-K+-ATPase activity in two cell lines (Table 1, P<0.05). Three triterpenic acids dose-dependently maintained MMP and Na+-K+-ATPase activity (P<0.05). H2O2 treatment raised caspase-3 and caspase-8 activities in 16HBE and BEAS-2B cells (Figure 4, P<0.05). Three triterpenic acids dose-dependently decreased caspase-3 activity in both cell lines; but only at high dose reduced caspase-8 activity in test cells (P<0.05).
Effects of AA, GA or OA upon oxidative and inflammatory factors.
As shown in Table 2, H2O2 treatment increased the production of ROS and 8-OHdG, and decreased GSH level in 16HBE and BEAS-2B cells (P<0.05). AA, GA or OA treatments dose-dependently lowered ROS and 8-OHdG formation; and reserved GSH content in those cells (P<0.05). H2O2 treatment also increased the release of IL-6, TNF- and PGE2 (Table 3, P<0.05). AA, GA or OA treatments dose-dependently decreased the generation of IL-6, TNF- and PGE2 (P<0.05).
Effects of AA, GA or OA upon NADPH oxidase, COX-2, NF-B and MAPK Expression.
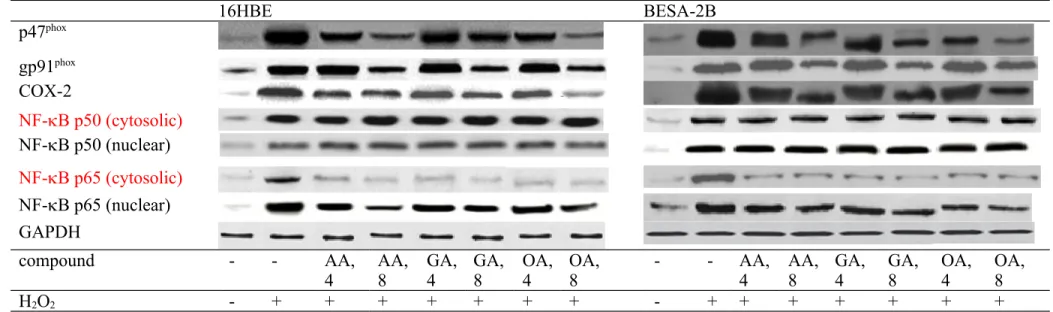
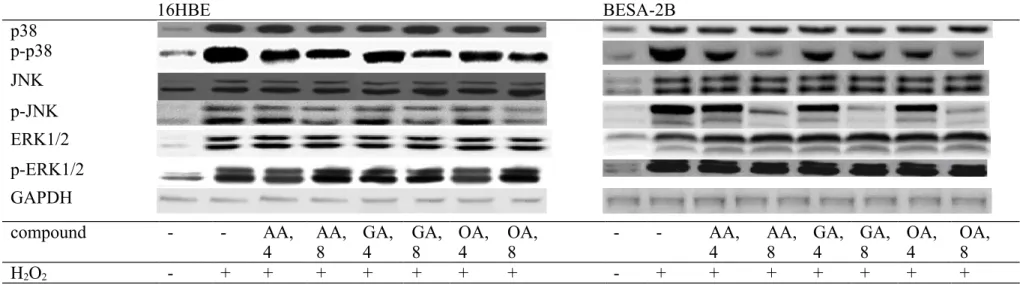
H2O2 treatment up-regulated the expression of p47phox, gp91phox, COX-2, NF-B p50 and NF-B p65 in 16HBE and BEAS-2B cells (Figure 5, P<0.05). AA, GA or OA treatments down-regulated the expression of p47phox, COX-2, cytosolic and nuclear NF-B p65 (P<0.05); but only at high dose decreased gp91phox expression (P<0.05). Three triterpenic acids failed to affect NF-B p50 expression in test cell lines (P>0.05). As shown in Figure 6, H2O2 treatment up-regulated MAPK expression in 16HBE and BEAS-2B cells (P<0.05). 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246
AA, GA or OA treatments suppressed p-p38 expression, and only at high dose decreased p-JNK expression in test cell lines (P<0.05).
DISCUSSION
Cigarette smoke and particulate matter could stimulate the release of hydrogen peroxide in airway epithelial cells, which contributes to apoptotic, oxidative and inflammatory stress in those cells.24,25 In our present study, pretreatments from three triterpenic acids, AA, GA and OA, markedly protected 16HBE and BEAS-2B cells, two human bronchial epithelial cell lines, against hydrogen peroxide induced oxidative and inflammatory injury, which in turn enhanced cell survival. Furthermore, we found that these compounds were able to mediate protein expression of NADPH oxidase, COX-2, NF-B and MAPK in these bronchial epithelial cells. In addition, these triterpenic acids treatments alone, without hydrogen peroxide, did not affect the growth of these test cells. These findings implied that these triterpenic acids might be safe and benefit respiratory epithelium stability and functions.
BCL family molecules are involved in mitochondria-mediated apoptotic pathway, in which Bcl-2 is an anti-apoptotic molecule, and Bax and cleaved caspase-3 are pro-apoptotic molecules. Hydrogen peroxide triggered mitochondria-mediated pro-apoptotic pathway via decreasing Bcl-2 expression, and increasing Bax and cleaved caspase-3 expression. However, triterpenic acids pretreatments effectively down-regulated cleaved caspase-3 expression and mildly retained Bcl-2 expression in two cell lines, which consequently mitigated hydrogen peroxide elicited mitochondria-mediated apoptotic stress. Na+-K+-ATPase, a key enzyme of TCA cycle, is critical for the maintenance of mitochondrial membrane ion homeostasis. Abnormality in this enzyme disturbs ion homeostasis and causes mitochondrial malfunctions.26 Thus, the reduction in Na+-K+
-ATPase activity and mitochondrial membrane potential in hydrogen peroxide treated cells 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271 272
indicated that mitochondrial structure and functions in those cells had been impaired, which in turn contributed to apoptotic insult. However, three triterpenic acids pretreatments markedly maintained Na+-K+-ATPase activity and mitochondrial membrane
potential in 16HBE and BEAS-2B cells. These results suggested that these compounds could enhance mitochondrial stability for those cells. Furthermore, our data revealed that these triterpenic acids alleviated hydrogen peroxide induced DNA fragmentation and 8-OHdG formation, indicators of DNA oxidative stress. These findings implied that these compounds could penetrate these bronchial epithelial cells, protect DNA and counteract apoptotic stress from hydrogen peroxide. On the other hand, loss of mitochondrial membrane potential activates caspases including caspase-3 and caspase-8.27 Both caspase-3 and caspase-8 are apoptotic executors because they directly affect cell morphological changes and the cleavage of nuclear proteins.21 Thus, the increased 3 and caspase-8 activities in hydrogen peroxide treated 16HBE and BEAS-2B cells could be partially ascribed to mitochondrial membrane potential change. Since test triterpenic acids already maintained mitochondrial membrane potential in both cell lines, the lower activity of both caspases could be explained. These results support that three triterpenic acids could protect 16HBE and BEAS-2B cells against apoptosis by stabilizing mitochondrial membrane and caspases.
Hydrogen peroxide evoked oxidative and inflammatory stress in two cell lines in our present study, which was evidenced by overproduction of ROS, 8-OHdG, IL-6 and TNF-. It has been reported that AA, GA and OA possessed anti-oxidative and anti-inflammatory activities.28-30 Thus, it seems reasonable to observe lower formation of oxidative and inflammatory factors in triterpenic acid-treated cells. NADPH oxidase complex is a key regulator for ROS generation and involved in the progression of respiratory system disorders.31,32 We found that triterpenic acids treatments substantially down-regulated protein expression of p47phox and gp91phox, cytosolic and membrane components of NADPH 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298
oxidase, which subsequently decreased ROS production in hydrogen peroxide treated cells. Apparently, these triterpenic acids could execute their anti-oxidative action through mediating NADPH oxidase. It is known that ROS could activate NF-B and MAPK pathways, which further exacerbates the formation of pro-inflammatory mediators.33,34 In our present study, triterpenic acids pretreatments dose-dependently down-regulated nuclear NF-B p65 and p-p38 expression; and at high dose limited JNK phosphorylation. These results could be partially ascribed to these agents already reduce ROS production. The other possibility was that these triterpenic acids might directly regulate NF-B and MAPK expression. Since both signalling pathways have been declined, the lower release of inflammatory cytokines such as IL-6 and TNF- in triterpenic acids treated cells could be explained. In addition, COX-2 is a critical enzyme responsible for the synthesis of PGE2, another key pro-inflammatory mediator. It has been indicated that COX-2 and its derived prostanoids, mainly including PGE2, thromboxane A2 and prostacyclin, are involved in cigarette smoking associated lung cancer progression.35 In our present study, three triterpenic acids markedly suppressed COX-2 protein expression, which in turn decreased PGE2 formation, and finally diminished inflammatory stress in those cells. These findings revealed that these compounds could protect 16HBE and BEAS-2B cells against hydrogen peroxide induced oxidative and inflammatory injury via declining NF-B, MAPK and COX-2 pathways.
It is interesting to find that OA was greater than other test compounds in suppressing NF-B p65 expression in 16HBE cells; but AA was more effective than others in lowering cleaved caspase-3 expression in BEAS-2B cells. Although AA, GA and OA have similar structures, their protective effects for these two bronchial epithelial cell lines were not identical. Three triterpenic acids are naturally occurring in many plant foods or botanicals used as food supplement ingredients.36 Consumers could obtain these natural triterpenic 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323
acids through dietary intake of certain plant foods. Thus, these agents seem safe for application. Our previous animal study indicated that dietary AA and OA could be absorbed and deposited in their intact forms in organs.12 The bioavailability of these agents certainly benefits their application. Thus, using these compounds for respiratory protection seems feasible. However, further in vivo study is necessary in order to verify the efficiency of these agents for airway diseases prevention or alleviation.
In conclusion, asiatic acid, glycyrrhizic acid and oleanolic acid at 4 or 8 mol/L protected human bronchial 16HBE and BEAS-2B cells against hydrogen peroxide induced oxidative and inflammatory injury via enhancing mitochondrial stability, decreasing ROS and PGE2 production and suppressing protein expression of NAPDH oxidase, COX-2, NF-B p65 and p-p38. These findings suggest that these agents or foods rich in these compounds could benefit bronchial epithelial cells; and might prevent the development of airway disorders.
Conflicts of interest statement
The authors declare that there are no conflicts of interest.
Acknowledgement
This study was partially supported by a grant from Ministry of Science and Technology, Taipei City, Taiwan (NSC 102-2313-B-039 -002 -MY3).
LITERATURE CITED
1. Goldkorn, T.; Filosto, S. Lung injury and cancer : Mechanistic insights into ceramide and EGFR signaling under cigarette smoke. Am. J. Respir. Cell Mol. Biol. 2010, 43, 259-268.
2. Tesfaigzi, Y. Roles of apoptosis in airway epithelia. Am. J. Respir. Cell Mol. Biol. 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349
2006, 34, 537-547.
3. Shi, Y.; Wang, F.; He, J.; Yadav, S.; Wang, H. Titanium dioxide nanoparticles cause apoptosis in BEAS-2B cells through the caspase 8/t-Bid-independent mitochondrial pathway. Toxicol. Lett. 2010, 196, 21-27.
4. Domej, W.; Oettl, K.; Renner, W. Oxidative stress and free radicals in COPD -implications and relevance for treatment. Int. J. Chron. Obstruct. Pulmon. Dis. 2014, 9, 1207-1224.
5. Mukherjee, B.; Bindhani, B.; Saha, H.; Ray, M.R. Increased oxidative DNA damage and decreased expression of base excision repair proteins in airway epithelial cells of women who cook with biomass fuels. Environ. Toxicol. Pharmacol. 2014, 38, 341-352.
6. Cho, I.H.; Gong, J.H.; Kang, M.K.; Lee, E.J.; Park, J.H.; Park, S.J.; Kang, Y.H.
Astragalin inhibits airway eotaxin-1 induction and epithelial apoptosis through modulating oxidative stress-responsive MAPK signaling. BMC Pulm. Med. 2014, 14, 122.
7. Geiszt, M.; Witta, J.; Baffi, J.; Lekstrom, K.; Leto, T.L. Dual oxidases represent novel hydrogen peroxide sources supporting mucosal surface host defense. FASEB J. 2003, 17, 1502-1504.
8. Ahn, E.K.; Yoon, H.K.; Jee, B.K.; Ko, H.J.; Lee, K.H.; Kim, H.J.; Lim, Y. COX -2 expression and inflammatory effects by diesel exhaust particles in vitro and in vivo.
Toxicol. Lett. 2008, 176, 178-187.
9. Park, G.Y.; Hu, N.; Wang, X.; Sadikot, R.T.; Yull, F.E.; Joo, M.; Peebles, R.S. Jr.; Blackwell, T.S.; Christman, J.W. Conditional regulation of cyclooxygenase-2 in tracheobronchial epithelial cells modulates pulmonary immunity. Clin. Exp. Immunol. 2007, 150, 245-254. 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372 373 374
10. Yang, J.; Li, Q.; Zhou, X.D.; Kolosov, V.P.; Perelman, J.M. Naringenin attenuates mucous hypersecretion by modulating reactive oxygen species production and inhibiting NF -κB activity via EGFR-PI3K-Akt/ERK MAPKinase signaling in human airway epithelial cells. Mol. Cell. Biochem. 2011, 351, 29-40.
11. Antognelli, C.; Gambelunghe, A.; Talesa, V.N.; Muzi, G. Reactive oxygen species induce apoptosis in bronchial epithelial BEAS-2B cells by inhibiting the antiglycation glyoxalase I defence: involvement of superoxide anion, hydrogen peroxide and NF - κB. Apoptosis 2014, 19, 102-116.
12. Yin, M.C.; Lin, M.C.; Mong, M.C.; Lin, C.Y. Bioavailability, distribution, and antioxidative effects of selected triterpenes in mice. J. Agric. Food Chem. 2012, 60, 7697-7701.
13. Kao, T.C.; Wu, C.H.; Yen, G.C. Bioactivity and potential health benefits of licorice .
J. Agric. Food Chem. 2014, 62, 542-553.
14. Raphael, T.J.; Kuttan, G. Effect of naturally occurring triterpenoids glycyrrhizic acid , ursolic acid , oleanolic acid and nomilin on the immune system. Phytomedicine 2003, 10, 483-489.
15. Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709-724.
16. Ramachandran, V.; Saravanan, R. Efficacy of asiatic acid , a pentacyclic triterpene on attenuating the key enzymes activities of carbohydrate metabolism in streptozotocin-induced diabetic rats. Phytomedicine 2013, 20, 230-236.
17. Wächter, G.A.; Valcic, S.; Flagg, M.L.; Franzblau, S.G.; Montenegro, G.; Suarez, E.; Timmermann, B.N. Antitubercular activity of pentacyclic triterpenoids from plants of Argentina and Chile. Phytomedicine 1999, 6, 341-345.
18. Liu, W.; Tan, X.; Shu, L.; Sun, H.; Song, J.; Jin, P.; Yu, S.; Sun, M.; Jia, X. Ursolic acid inhibits cigarette smoke extract-induced human bronchial epithelial cell injury
375 376 377 378 379 380 381 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396 397 398 399 400
and prevents development of lung cancer. Molecules 2012, 17, 9104-9115.
19. Kastner, P.E.; Casset, A.; Pons, F. Formaldehyde interferes with airway epithelium integrity and functions in a dose- and time-dependent manner. Toxicol. Lett. 2011, 200, 109-116.
20. Deng, Z.; Zhou, J.J.; Sun, S.Y.; Zhao, X.; Sun, Y.; Pu, X.P. Procaterol but not dexamethasone protects 16HBE cells from H ₂ O ₂ -induced oxidative stress. J. Pharmacol. Sci. 2014, 125, 39-50.
21. Sharifi, A.M.; Eslami, H.; Larijani, B.; Davoodi, J. Involvement of caspase -8, -9, and -3 in high glucose-induced apoptosis in PC12 cells. Neurosci. Lett. 2009, 459, 47-51. 22. Musa, M.A.; Badisa, V.L.; Latinwo, L.M. Cytotoxic activity of N, N'-Bis
(2-hydroxybenzyl) ethylenediamine derivatives in human cancer cell lines. Anticancer
Res. 2014, 34, 1601-1607.
23. Torlinska, T.; Grochowalska, A. Age-related changes of Na+, K+-ATPase, Ca2+
-ATPase and Mg2+-ATPase activities in rat brain synaptosomes. J. Physiol. Pharmacol. 2004, 55, 457-465.
24. Yi, S.; Zhang, F.; Qu, F.; Ding, W. Water-insoluble fraction of airborne particulate matter (PM10) induces oxidative stress in human lung epithelial A549 cells . Environ. Toxicol. 2014, 29, 226-233.
25. Downs, C.A.; Montgomery, D.W.; Merkle, C.J. Age-related differences in cigarette smoke extract-induced H2O2 production by lung endothelial cells . Microvasc. Res. 2011, 82, 311-317.
26. Liu, N.; Li, Y.; Su, S.; Wang, N.; Wang, H.; Li, J. Inhibition of cell migration by ouabain in the A549 human lung cancer cell line. Oncol. Lett. 2013, 6, 475-479. 27. Xiao, H.; Rawal, M.; Hahm, E.R.; Singh, S.V. Benzo[a]pyrene-7,8-diol-9,10-epoxide
causes caspase -mediated apoptosis in H460 human lung cancer cell line. Cell Cycle 2007, 6, 2826-2834. 401 402 403 404 405 406 407 408 409 410 411 412 413 414 415 416 417 418 419 420 421 422 423 424 425 426
28. Chan, C.Y.; Mong, M.C.; Liu, W.H.; Huang, C.Y.; Yin, M.C. Three pentacyclic triterpenes protect H9c2 cardiomyoblast cells against high-glucose-induced injury.
Free Radic. Res. 2014, 48, 402-411.
29. Kao, T.C.; Wu, C.H.; Yen, G.C. Glycyrrhizic acid and 18β-glycyrrhetinic acid recover glucocorticoid resistance via PI3K-induced AP1, CRE and NFAT activation.
Phytomedicine 2013, 20, 295-302.
30. Wang, X.; Liu, R.; Zhang, W.; Zhang, X.; Liao, N.; Wang, Z.; Li, W.; Qin, X.; Hai, C. Oleanolic acid improves hepatic insulin resistance via antioxidant, hypolipidemic and anti- inflammatory effects. Mol. Cell. Endocrinol. 2013, 376, 70-80.
31. Griffith, B.; Pendyala, S.; Hecker, L.; Lee, P.J.; Natarajan, V.; Thannickal, V.J. NOX enzymes and pulmonary disease. Antioxid. Redox. Signal. 2009, 11, 2505-2516. 32. Dergham, M.; Lepers, C.; Verdin, A.; Billet, S.; Cazier, F.; Courcot, D.; Shirali, P.;
Garçon, G. Prooxidant and proinflammatory potency of air pollution particulate matter (PM ₂ . ₅₋₀ . ₃ ) produced in rural, urban, or industrial surroundings in human bronchial epithelial cells ( BEAS-2B ). Chem. Res. Toxicol. 2012, 25, 904-919.
33. Yao, H.; Yang, S.R.; Kode, A.; Rajendrasozhan, S.; Caito, S.; Adenuga, D.; Henry, R.; Edirisinghe, I.; Rahman, I. Redox regulation of lung inflammation: role of NADPH oxidase and NF-kappaB signalling. Biochem. Soc. Trans. 2007, 35, 1151-1155.
34. Park, W.H. Effects of antioxidants and MAPK inhibitors on cell death and reactive oxygen species levels in H2O2-treated human pulmonary fibroblasts. Oncol. Lett. 2013, 5, 1633-1638.
35. Huang, R.Y.; Chen, G.G. Cigarette smoking, cyclooxygenase-2 pathway and cancer .
Biochim. Biophys. Acta 2011, 1815, 158-169.
36. Caligiani, A.; Malavasi, G.; Palla, G.; Marseglia, A.; Tognolini, M.; Bruni, R. A simple GC-MS method for the screening of betulinic, corosolic, maslinic, oleanolic 427 428 429 430 431 432 433 434 435 436 437 438 439 440 441 442 443 444 445 446 447 448 449 450 451 452
and ursolic acid contents in commercial botanicals used as food supplement ingredients. Food Chem. 2013, 136, 735-741.
453 454 455
Figure 1. Effects of AA, GA and OA upon the growth of 16HBE and BEAS-2B cells, without H2O2 treatment. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr. Control groups were cells contained no test compound. Data are meanSD (n=9). 456 457 458 459 460 461
Figure 2. Effects of AA, GA and OA upon viability and DNA fragmentation determined as enrichment factor in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr, and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-eMeans values among bars without a common letter differ, P<0.05. 462 463 464 465 466 467 468 469 470 471 472
Figure 3. Effects of AA, GA or OA upon Bcl-2, Bax and cleaved caspase-3 expression in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-fMeans values among bars without a common letter differ, P<0.05. 16HBE BESA-2B Bcl-2 Bax cleaved caspase-3 GAPDH compound - - AA,
4 AA, 8 GA,4 GA,8 OA,4 OA, 8 - - AA,4 AA,8 GA,4 GA, 8 OA,4 OA,8
H2O2 - + + + + + + + - + + + + + + + 473 474 475 476 478 479
480 481
482
Table 1. Effects of AA, GA or OA upon level of mitochondrial membrane potential, determined as MFI, and Na+-K+-ATPase activity in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-fMeans values in a column without a common letter differ, P<0.05.
MFI Na+-K+-ATPase activity 16HBE Control 100f 100f H2O2 372a 474a AA, 4+ H2O2 784d 673c AA, 8+ H2O2 845e 795d GA, 4+ H2O2 523b 574b GA, 8+ H2O2 723d 705c OA, 4+ H2O2 654c 722c OA, 8+ H2O2 802e 845e BEAS-2B Control 100e 100f H2O2 343a 414a AA, 4+ H2O2 695c 654c AA, 8+ H2O2 786d 835e GA, 4+ H2O2 484b 502b GA, 8+ H2O2 623c 634c OA, 4+ H2O2 685c 713d OA, 8+ H2O2 793d 815e 484 485 486 487 488 489 490 491
Figure 4. Effects of AA, GA and OA upon activity of caspase-3 and caspase-8 in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-fMeans values among bars without a common letter differ, P<0.05.
492 493 494 495 496 497 498 499 500
Table 2. Effects of AA, GA or OA upon level of ROS (RFU/mg protein), 8-OHdG (pg/mg protein) and GSH (ng/mg protein) in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-fMeans values in a column without a common letter differ, P<0.05. ROS 8-OHdG GSH 16HBE Control 0.130.07a 295a 1066e H2O2 2.080.22f 18719e 433a AA, 4+ H2O2 1.150.14c 13314d 644b AA, 8+ H2O2 0.520.09b 824b 905d GA, 4+ H2O2 1.570.18e 14818d 564b GA, 8+ H2O2 0.980.11c 11411c 755c OA, 4+ H2O2 1.270.15d 14213d 623b OA, 8+ H2O2 0.660.10b 888b 844d BEAS-2B Control 0.150.04a 256a 1094f H2O2 2.140.25e 1952e 462a AA, 4+ H2O2 1.320.16c 14116c 695c AA, 8+ H2O2 0.610.09b 869b 906e GA, 4+ H2O2 1.690.17d 16013d 572b GA, 8+ H2O2 1.130.12c 12311c 813d OA, 4+ H2O2 1.240.14c 13715c 794d OA, 8+ H2O2 0.790.08b 0747b 925e 501 502 503 504 505 506 507 508 509
Table 3. Effects of AA, GA or OA upon level (pg/mg protein) of IL-6, TNF- and PGE2 in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-fMeans values in a column without a common letter differ, P<0.05.
IL-6 TNF- PGE2 16HBE Control 142a 194a 233a H2O2 1275e 1498e 1365d AA, 4+ H2O2 764c 1117d 952c AA, 8+ H2O2 423b 803c 714b GA, 4+ H2O2 935d 1146d 924c GA, 8+ H2O2 693c 825c 663b OA, 4+ H2O2 884d 1024d 905c OA, 8+ H2O2 653c 565b 682b BEAS-2B Control 153a 203a 214a H2O2 1376d 1626f 1679e AA, 4+ H2O2 852c 1045e 996d AA, 8+ H2O2 583b 674c 643b GA, 4+ H2O2 895c 886d 945d GA, 8+ H2O2 664b 463b 614b OA, 4+ H2O2 813c 1012e 782c OA, 8+ H2O2 602b 694c 533b 510 511 512 513 514
Figure 5. Effects of AA, GA or OA upon protein expression of p47phox, gp91phox, COX-2, and NF-B in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-fMeans values among bars without a common letter differ, P<0.05.
16HBE BESA-2B p47phox gp91phox COX-2 NF-B p50 (cytosolic) NF-B p50 (nuclear) NF-B p65 (cytosolic) NF-B p65 (nuclear) GAPDH compound - - AA,
4 AA,8 GA,4 GA,8 OA,4 OA, 8 - - AA,4 AA,8 GA, 4 GA, 8 OA, 4 OA, 8
H2O2 - + + + + + + + - + + + + + + + 515 516 517 518 519
521
522
524
Figure 6. Effects of AA, GA or OA upon protein expression of MAPK in 16HBE and BEAS-2B cells. Cells were pre-treated with test compound at 4 or 8 mol/L for 48 hr and followed by adding 100 M H2O2 to induce cell damage. Control groups were cells contained no test compound, and without H2O2 treatment. Data are meanSD (n=9). a-eMeans values among bars without a common letter differ, P<0.05.
16HBE BESA-2B p38 p-p38 JNK p-JNK ERK1/2 p-ERK1/2 GAPDH compound - - AA,
4 AA,8 GA,4 GA,8 OA,4 OA, 8 - - AA, 4 AA,8 GA,4 GA,8 OA, 4 OA, 8
H2O2 - + + + + + + + - + + + + + + +
526 527 528
530 531
TOC Graphic
MS title: Anti-oxidative and anti-inflammatory protection of three triterpenic acids in normal human bronchial epithelial cells
16HBE BESA-2B p47phox gp91phox COX-2 NF-B p50 (cytosolic) NF-B p50 (nuclear) NF-B p65 (cytosolic) NF-B p65 (nuclear) GAPDH compound - - AA,
4 AA,8 GA,4 GA,8 OA,4 OA, 8 - - AA,4 AA,8 GA, 4 GA, 8 OA, 4 OA, 8
H2O2 - + + + + + + + - + + + + + + +
533 534 535