國立臺灣大學生命科學院生命科學系 碩士論文
Department of Life Science College of Life Science
National Taiwan University Master Thesis
線蟲及魚蚤與盲鰻之寄生關係 Parasitism of Anisakid Nematodes and
Chondracanthid Copepod in Hagfish
駱皓元 Hao-Yuan Luo
指導教授:施秀惠 博士 Advisor: Hsiu-Hui Shih, Ph.D.
中華民國 104 年 6 月
June 2015
i
ii
誌謝
在此感謝在我研究所生涯幫助過我的所有人。首先,最要感謝的是我的指導 教授施秀惠老師,老師對學生的信任,不斷的給學生信心,同時在研究上的叮嚀、
指導,以及對學生生活、課業上的關心,讓學生能夠順利的完成碩士生涯。感謝 王蓮成老師、冉繁華老師,兩位老師細心修改學生的論文,並且給予學生許多論 文上的指點、建議,指出學生沒注意到的盲點,讓學生的論文能夠更加完整。
感謝海洋所鄭有容博士協助橈足類鑑定以及後續發表,感謝大溪漁港陳朝明 夫婦熱心幫忙,協助樣本的採集。感謝慧瑜學姊細心的修改論文,不時的與我討 論論文的內容,並擔任我的口試委員。感謝惠冠學姊及茂麟學長,從我一進實驗 室開始就不斷的帶領我,在生活、實驗上給予大量的建議與幫助,並一起分擔生 活中的各種苦樂。
最後要感謝我的父母,系上的助教、老師,研究所的同學,以及諸位朋友,
填滿了我碩士生涯的每一的部分。
iii
中文摘要
盲鳗是脊椎動物中最原始的一群,只有軟骨形成的頭骨,沒有脊椎骨。在海 洋食物鏈中扮演重要的腐食生物角色。海獸胃線蟲為海水魚中常見的寄生蟲,藉 由寄主的掠食行為進行寄主轉移,並且以橈足類、頭足類、硬骨魚類為保蟲寄主。
本研究將探討盲鰻寄生蟲相,以及盲鰻食用其他海洋動物的屍體時,是否能經由 腐食方式感染海獸胃線蟲。
盲鰻樣本於2013 年 11 月至 2014 年 6 月宜蘭大溪漁港向漁民購買,共包含四 種盲鰻:蒲氏黏盲鰻 (Eptatretus burgeri)、楊氏黏盲鰻 (E. yangi)、沈氏黏盲鰻 (E.
sheni)、臺灣黏盲鰻 (E. taiwanae),經調查後發現七種海獸胃線蟲寄生,主要寄生 於腸道,極少部分進入體腔及肌肉,經過PCR-RFLP 及 rDNA ITS 序列定序後確定 為 Anisakis pegreffii、A. simplex s.s.、A. pegreffii 與 A. simplex s.s.的重組基因型、A.
typica、Anisakis sp.、A. brevispiculata、Hysterothylacium amoyense。盲鰻為 7 種海 獸胃線蟲之寄主新紀錄,此外 Anisakis sp.及 H. amoyense 為地理新紀錄。另外,也 發現一種橈足類,盲鰻刺魚蚤 (Acanthochondria eptatreti),寄生於盲鰻的咽部,已 於2014 發表為新種。
研究首度發現海獸胃線蟲能以腐食方式傳播並寄生於盲鰻,修改了海獸胃線 蟲之生活史。統計結果顯示,蒲氏黏盲鰻之線蟲盛行率及盲鰻刺魚蚤盛行率皆顯 著高於其他三種盲鰻。蒲氏黏盲鰻與沈氏黏盲鰻之線蟲感染強度也與體長呈現正 相關,盲鰻刺魚蚤感染強度只與蒲氏黏盲鰻有正相關。也觀察到寄生蟲於寄主的 累積效應。
關鍵字:盲鰻、食腐動物、海獸胃線蟲、橈足類寄生蟲、盲鰻刺魚蚤
iv
Abstract
Hagfishes are the most primitive vertebrates known. They have a cartilaginous
skull but no vertebral column. Hagfish is marine benthic scavenger which feed on dead
organic matter, including teleosts, cetaceans, marine inverts, etc. Anisakis is transferred
by host predation, and their paratenic hosts include crustaceans, cephalopods, and
teleost fish. The aim of this study is to investigate the parasitical fauna of hagfish and
confirm that hagfish can be infected by Anisakis via host scavenging.
Hagfishes were caught from Daxi fishery harbor in northeastern Taiwanese waters
from November 2013 to June 2014. A total of 265 samples were examined and four
species of the genus Eptatretus, Eptatretus burgeri, E. yangi, E. sheni and E. taiwanae.
These four species of hagfishes were infected by Anisakidae nematodes, which
parasitized mainly in the gastrointestinal tract wall. Also, anisakid nematodes species
were identified by PCR-RFLP. Seven species of nematodes were recorded: Anisakis
pegreffii, A. simplex s.s., recombinant genotype of A. pegreffii and A. simplex s.s.,
A.typica, Anisakis sp., A. brevispiculata and Hysterothylacium amoyense. All Anisakid
nematodes are new host records, and two of them (Anisakis sp. and H. amoyense) are
new locality records. In addition, a new species of chondracanthid copepod,
v
Acanthochondria eptatreti, was identified and isolated from the pharynx of hagfish, and
has been published in 2014.
The results proved that anisakid nematodes can transfer by hagfish scavenging and
modified Anisakis life cycle. Statistical analysis showed that the prevalences of two
kinds of parasites (nematode and copepod) are significant higher in E. burgeri. The
intensity of nematodes positively correlated with host size of E. burgeri and E. sheni.
The intensity of copepod only has positive correlation with E. burgeri. The
accumulation of parasites in larger hosts can be observed in this study.
Key word: Hagfish, scavenger, Anisakis, copepod parasite, Acanthochondria eptatreti
vi
目 錄
口試委員會審定書 ... i
誌謝 ... ii
中文摘要 ... iii
Abstract ... iv
目 錄 ... vi
表目錄 ... ix
圖目錄 ... x
第 1 章 前言 ... 1
1.1 寄生關係 ... 1
1.2 魚源性寄生蟲 ... 1
1.3 寄生蟲於生物標籤之應用 ... 2
1.4 海獸胃線蟲 ... 3
1.4.1 海獸胃線蟲之概略 ... 3
1.4.2 海獸胃線蟲之生活史 ... 3
1.4.3 海線胃線蟲症 ... 4
1.4.4 安尼線蟲科之鑑定 ... 7
1.5 盲鰻 ... 9
1.5.1 分類地位 ... 9
1.5.2 形態特徵 ... 9
1.5.3 生態習性 ... 10
1.5.4 地理分布 ... 10
1.5.5 盲鰻利用現況 ... 11
1.5.6 相關研究 ... 12
1.6 研究目的 ... 13
vii
第 2 章 材料與方法 ... 14
2.1 樣本取得 ... 14
2.2 樣本處理 ... 15
2.3 安尼線蟲科線蟲之PCR-RFLP 鑑種 ... 16
2.3.1 Genomic DNA 萃取 ... 16
2.3.2 PCR 增幅 ... 17
2.3.3 RFLP 分析 ... 18
2.4 感染參數之分析 ... 18
第 3 章 結果 ... 20
3.1 盲鰻之寄生蟲相 ... 20
3.2 海獸胃線蟲之PCR-RFLP 鑑種結果 ... 20
3.3 感染參數之分析結果 ... 22
3.3.1 四種盲鰻感染參數之比較分析 ... 22
3.3.2 盲鰻性別分組與感染參數之比較分析 ... 23
3.3.3 盲鰻體長與線蟲感染參數分析 ... 25
3.3.4 盲鰻體長與盲鰻刺魚蚤感染參數分析 ... 26
第 4 章 討論 ... 28
4.1 盲鰻之寄生蟲相 ... 28
4.2 海獸胃線蟲物種 ... 30
4.3 感染參數結果討論 ... 33
4.3.1 四種盲鰻之感染參數之比較分析討論 ... 33
4.3.2 盲鰻體長與線蟲感染參數之分析討論 ... 34
4.3.3 盲鰻體長與盲鰻刺魚蚤感染參數之分析討論 ... 36
4.4 盲鰻與海獸胃線蟲症 ... 37
4.5 總結 ... 38
viii
參考文獻 ... 40 附錄 ... 79
ix
表目錄
表 1 盲鰻體內檢獲之安尼線蟲科線蟲經二種內切酶酵素切割後片段大小 .... 50
表 2 四種盲鰻感染線蟲之感染參數比較 ... 51
表 3 四種盲鰻感染盲鰻刺魚蚤之感染參數比較 ... 52
表 4 盲鰻性別分組與海獸胃線蟲感染參數比較 ... 53
表 5 盲鰻性別分組與盲鰻刺魚蚤感染參數比較 ... 54
x
圖目錄
圖 1 海獸胃線蟲之生活史 ... 4
圖 2 盲鰻黏液孔分佈 ... 14
圖 3 盲鰻鰓孔特徵 ... 55
圖 4 盲鰻體長測量 ... 56
圖 5 盲鰻體長分佈圖 ... 57
圖 6 寄生於盲鰻之海獸胃線蟲 ... 58
圖 7 盲鰻刺魚蚤寄生於蒲氏黏盲鰻 ... 59
圖 8 7 種海獸胃線蟲 ITS 序列經 HinfI 酵素作用之電泳分析圖 ... 60
圖 9 7 種海獸胃線蟲 ITS 序列經 HhaI 酵素作用之電泳分析圖 ... 61
圖 10 寄生於四種盲鰻之海獸胃線蟲蟲種百分比 ... 62
圖 11 沈氏黏盲鰻體長與線蟲感染率之邏輯迴歸關係 ... 63
圖 12 所有盲鰻體長與線蟲感染率之邏輯迴歸關係 ... 64
圖 13 所有盲鰻分組體長與線蟲盛行率之線性關係 ... 65
圖 14 盲鰻分組體長與線蟲盛行率之線性關係圖 ... 66
圖 15 所有盲鰻體長與線蟲感染強度之線性關係圖 ... 67
圖 16 盲鰻體長與線蟲感染強度之線性關係圖 ... 68
圖 17 所有盲鰻體長與線蟲豐富度之線性關係圖 ... 69
圖 18 盲鰻體長與線蟲豐富度之線性關係圖 ... 70
圖 19 所有盲鰻體長與盲鰻刺魚蚤感染率之邏輯迴歸關係圖 ... 71
圖 20 所有盲鰻分組體長與盲鰻刺魚蚤盛行率之線性關係圖 ... 72
圖 21 盲鰻分組體長與盲鰻刺魚蚤盛行率之線性關係圖 ... 73
圖 22 所有盲鰻體長與盲鰻刺魚蚤感染強度 ... 74
圖 23 盲鰻體長與盲鰻刺魚蚤感染強度之線性關係圖 ... 75
xi
圖 24 所有盲鰻體長與盲鰻刺魚蚤豐富度之線性關係圖 ... 76 圖 25 盲鰻體長與盲鰻刺魚蚤豐富度之線性關係圖 ... 77 圖 26 海獸胃線蟲之生活史修改 ... 78
1
第 1 章 前言
1.1 寄生關係
自然界的生物以多種方式生活在地球上,包括自由生活與共生生活。共生,
是指生物共同生活在一起的現象,包括片利共生 (Commensalism)、互利共生 (Mutualism) 與寄生 (Parasitism)。在共生生活中,當兩個物種生活在一起時,其中 一物種從另一物種獲取利益,而另一物種受害,這種關係稱之為寄生。而獲取利 益的動物稱為寄生蟲 (parasite),受害的物種稱為寄主 (host)。
以魚類為寄主的寄生蟲繁多,除了多種原生生物外,還有體型較大的後生動 物,如扁形動物門中的單殖吸蟲、複殖吸蟲及絛蟲,圓形動物門的線蟲,節肢動 物門的魚虱、甲殼動物和棘頭動物門的棘頭蟲等。這些寄生蟲會對魚體造成負面 影響,主要有四種:(一) 機械性刺激及損傷:如魚虱寄生時,利用其口器撕裂寄 主皮膚或是鰓部,使組織受損;(二) 擠壓與阻塞:如等足目的魚怪寄生於魚體內,
擠壓內部器官及影響寄主的生殖腺發育;(三) 掠奪寄主營養:以寄主血液為食的 吸蟲,造成寄主瘦弱,甚至死亡;(四) 毒素作用:寄生蟲於寄主體內時,其代謝 物或排泄物對寄主產生的負面影響 (張等,1999)。
1.2 魚源性寄生蟲
魚類寄生蟲種類繁多,其中有包含許多人魚共通的寄生蟲,在食用生魚或是 未完全煮熟的魚時,就有可能遭到感染。病例多發於中、低收入的國家,同時也 和飲食習慣、地方傳統有關,隨著國際市場的需求、飲食文化交流,國與國間的 貿易往來與交通的便利性,其感染魚源性寄生蟲的風險也隨之增加 (Chai et al.
2005)。
2
常見的人魚共通寄生蟲如寄生於淡水魚與軟體動物的肝吸蟲 (liver fluke),會 造成吸蟲症 (Trematodiasis),主要症狀為膽管炎、膽管結石、胰腺炎、膽管癌等症 狀;寄生於淡水、海水及洄游性魚類的裂頭絛蟲 (Diphyllobothrium) 引起的裂頭絛 蟲症 (Diphyllobothriasis),寄生於人類腸道時會造成疲倦、腹瀉等輕微症狀,隨著 蟲體長大可能會造成腸道阻塞及膽道阻塞的症狀;以及在海水魚常見的海獸胃線 蟲 (Anisakis),其引起海獸胃線蟲症 (Anisakiasis),隨著線蟲幼蟲入侵腸、胃壁造 成腹痛、噁心、嘔吐外,同時也可能產生強烈的過敏反應 (Chai et al. 2005)。
1.3 寄生蟲於生物標籤之應用
魚類寄生蟲為很好的生物標籤,可用來評估魚類的系群、移動、攝食習慣和 族群補充。其中,系群是漁業管理中,族群的基本單位,系群的研究可以減少錯 誤的評估、改善資源的管理。系群區分的方法包含形態差異、遺傳變異、耳石微 量元素、生活史、寄生蟲相等。如地中海的竹莢魚 (Trachurus trachurus)可依據寄 生蟲相的不同來區分系群;東部地中海的竹筴魚受到黏孢子蟲 Alataspora solomoni 及絛蟲長尾幼蟲感染,中部地中海則是有複殖吸蟲 Bathycreadium elongatum、單殖 吸蟲 Paradiplectanotrema trachuri 及橈足類 Lernanthropus trachuri 寄生,再加上形 態測量數據就可以將地中海竹筴魚分為東部、中部、西部三個系群 (Abaunza et al., 2008)。
地中海與大西洋的海獸胃線蟲物種也明顯不同,如 Anisakis pegreffii 在地中海 皆為優勢物種,A. simplex s.s.則為大西洋優勢物種,同時也可以利用海獸胃線蟲物 種來判斷寄主的食性,如底棲性魚類的歐洲狗鱈 (Merluccius merluccius),其體內 線蟲物種較大洋性小型魚類的竹筴魚 (T. trachurus) 豐富,推測為歐洲狗鱈會以頭 足類、甲殼類及其他底棲性魚類為食,接受不同寄主的海獸胃線蟲,而竹莢魚主 要以小型魚類及浮游生物為食,因此感染線蟲物種數較少。劍旗魚 (Xiphias gladius)
3
也可以利用海獸胃線蟲的物種來判斷寄主的食性,如 A. physeteris 主要發現在魷魚 體內,當劍旗魚感染高比率的 A. physeteris 時,就可以推測魷魚可能為劍旗魚主要 獵物之一 (Mattiucci et al., 2007)。
1.4 海獸胃線蟲
1.4.1 海獸胃線蟲之概略
海獸胃線蟲又稱異尖線蟲、安尼線蟲,主要是指海獸胃線蟲屬中的物種,以 及部分安尼線蟲科中物種。其最終寄主為海洋哺乳類,如:鯨魚、海豚、鰭腳目 動物,保蟲寄主為橈足類以及多種硬骨魚、頭足類,為海水魚常見之寄生蟲。
海獸胃線蟲之分類地位如下:
圓形動物門 (Phylum Nematoda) 色矛綱 (Class Chromadorea)
蛔蟲目 (Order Ascaridida)
安尼線蟲科 (Family Anisakidae) 海獸胃線蟲屬 (Genus Anisakis) 對盲囊線蟲屬 (Genus Contracaecum) 宮脂線蟲屬 (Genus Hysterothylacium) 前盲囊線蟲屬 (Genus Porrocaecum)
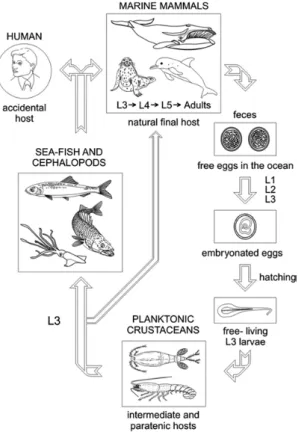
1.4.2 海獸胃線蟲之生活史
海獸胃線蟲之最終寄主為海洋哺乳類,寄生於消化道中,其蟲卵會隨著寄主 的糞便排出;排出後的蟲卵在海水中發育為第一期幼蟲 (Larval stage 1),經兩次蛻 變後,孵化為在海中自由生活的第三期幼蟲 (L3);第三期幼蟲會被橈足類及磷蝦
4
等小型甲殼類動物食入 (Klimpel et al., 2004),接著這些小型甲殼類被其他魚類、
頭足類食入後,第三期幼蟲會鑽入寄主的腸道並寄生,或是穿出腸道侵入腹腔、
其他器官或肌肉且寄生於其中,此時第三期幼蟲並不會發育,這些受感染的小型 甲殼類、魚類及頭足類稱為保蟲寄主 (paratenic hosts),最常見的硬骨魚寄主為鯖 科、鱈科、鰺科及鯡科 (Kuhn et al., 2013)。經由食物鏈關係,將幼蟲向食物鏈上 層感染,最終回到最終寄主;第三期幼蟲在最終寄主海洋哺乳類的胃中,受到溫 度、酸鹼值及胃蛋白酶促使下 (Iglesias et al., 2001),經過兩次蛻變後,發育為具有 生殖能力成蟲,此為海獸胃線蟲之生活史。
圖 1 海獸胃線蟲之生活史 (Audicana and Kenned, 2008)
1.4.3 海線胃線蟲症
海獸胃線蟲症 (Anisakiosis) 是指人類受到了安尼線蟲科 (Family Anisakidae) 幼蟲感染所產生的症狀,海獸胃線蟲症的感染往往是因為生食或食入未煮熟的魚 類、頭足類這些帶有海獸胃線蟲幼蟲的保蟲寄主而感染。病症名稱依感染物種還
5
可分為:Anisakiasis,以海獸胃線蟲屬 (Anisakis) 幼蟲感染的病症,包含 A. simplex、
A. pegreffii;Pseudoterranovosis,為前盲囊線蟲屬幼蟲 Pseudoterranova (Porrocaecum) decipiens 所感染的病症;Anisakiosis 則是泛指所有安尼線蟲科幼蟲感染的病症 (Audicana and Kenned, 2008)。全球至少有兩萬筆感染案例,主要發生於日本,部 分發生於西歐國家,在美國、加拿大、智利、紐西蘭、埃及等國也有案例,隨著 醫療技術的進步,以及對生食海鮮風險的認知,海獸胃線蟲症被診斷出來的機會 也會增高 (Chai et al. 2005)。
海獸胃線蟲症根據期病灶及位置可以分成四型:(一) 胃型海獸胃線蟲症,一 般在食入後1~12 小時內急遽發作,此時線蟲鑽入腸黏膜,造成紅斑、黏膜水腫,
會有強烈的上腹疼痛、噁心、嘔吐、低程度發燒,若是沒有治療可能會有慢性潰 瘍的症狀;(二) 腸型海獸胃線蟲症,在食入 5~7 天後,發生間續或持續性腹痛,
以及腹水或腹膜炎的跡象,發炎位置主要在迴腸末端及結腸,少部分會發生腸阻 塞、腸套疊、腸穿孔或氣腹;(三) 異位型海獸胃線蟲症,較為少見,為線蟲幼蟲 穿出腸胃道後,移動到腹膜腔、胸膜腔、肝臟、胰臟、卵巢、腹部皮下組織,造 成慢性發炎;(四) 過敏型海獸胃線蟲症,因海獸胃線蟲所以起的過敏反應,平均 在食用後 5 個小時開始產生過敏症狀,如蕁麻疹、血管性水水腫,有時會伴隨腸 胃道症狀一起發生 (Hochberg and Hamer, 2010)。根據美國食藥署建議,在食用魚 類時至少要以 -20℃冷凍 7 天,或是-35℃冷凍 15 小時才能確保線蟲死亡 (FDA/CFSAN, 2001)。
在美國、歐洲等海獸胃線蟲症病例較少的國家,因其腹痛的症狀常常被誤認 為是其他原因造成,而非海獸胃線蟲而被忽略,因此如何確實診斷為海獸胃線蟲 症則非常重要。首先要確認病患在腹痛前是否有食用生魚及距離食用的時間多久,
在美國一則回顧病例的報告中顯示,39 例胃型海獸胃線蟲症平均在食用生魚後 6 小時產生劇烈腹痛,腸型海獸胃線蟲平均在48 小時後產生腹痛。胃型海獸胃線蟲
6
症可利用內視鏡診斷胃部是否有蟲體,並且將蟲體取出。腸型海獸胃線蟲症則可 以利用電腦斷層掃描,可以看見腸壁增厚、腹水、組織發炎等徵狀,以及搭配血 液中抗海獸胃線蟲的抗體濃度 (IgG、IgE) 來進行診斷,然而確診後只能接受輸液、
止痛這種較保守性的治療,使患者自行復原;如果開刀將蟲體取出,造成的傷口 及感染等風險可能高於原本的病症。另外,腸型海獸胃線蟲症發生症狀時可能距 離食用生魚已有一段時間,病患可能遺忘是否有食用生魚,這也使海獸胃線蟲被 確診的機會降低 (Chung and Lee, 2014; Takabayashi et al., 2014)。
臺灣首度報導海獸胃線蟲症病例,病患為ㄧ名69 歲臺灣男性,在進行內視鏡 檢察時發現線蟲寄生於病患的胃部,且一端已經嵌入胃壁之中,胃中的線蟲取出 鑑定後,確定為 A. simplex s.s.,與日本海域主要感染魚類的海獸胃線蟲種類相同,
而與臺灣週遭海域主要出現的 A. pegreffii 不同。而該名男子正好在檢察前三天有 到日式餐廳食用生魚片,是否因為在運輸過程中冷凍時間不足,無法確實殺死線 蟲而感染已無法確定,但此案例也顯示漁貨間的貿易也可能增加寄生蟲傳染的可 能性 (Li et al., 2015)。
海獸胃線蟲症案例最多的國家是日本,每年約有 500 例確診病例,與其喜愛 食用生魚的文化有密切相關,感染物種則是以 A. simplex s.s.為主 (Suzuki and Murata, 2011)。在韓國每年也約有 200 件感染海獸胃線蟲症病例,從 16 名病患胃 部取出的蟲體,經過分子鑑定後,有15 名病患是由 A. pegreffii 所感染,1 名為 A.
simplex s.s.,結果與韓國海域中主要感染魚類的海獸胃線蟲物種相同 (Lim et al., 2015)。研究顯示,A. simplex s.s.在人工胃液的處理時,對酸的忍受度較 A. pegreffii 高,且在培養基培養時發現 A. simplex s.s.鑽入培養基膠體的比率明顯高於 A.
pegreffii,推測 A. simplex s.s.在進入消化道後,更容易鑽入消化道壁而造成明顯症 狀,因此推測 A. simplex s.s.是造成海獸胃線蟲症的主要物種 (Arizono et al., 2012)。
無論是 A. pegreffii 或是 A. simplex s.s.都確定會造成海獸胃線蟲症,而海獸胃線蟲的
7
地理分布,或許才是造成不同地區間感染物種不同的主因。
海獸胃線蟲過敏症,是經由輔助T 細胞 Th-2,產生細胞激素吸引嗜酸性、嗜 鹼性白血球、肥大細胞,以及活化B 細胞來產生大量 IgE 抗體,與 IgE 結合的肥 大細胞釋出包含組織胺等過敏因子的顆粒,引發過敏反應。目前已知的海獸胃線 蟲過敏抗原有13 種 (Ani s1-13),7 種為海獸胃線蟲產生的代謝產物 (Ani s1, 4-9),
以及2 種線蟲本體蛋白抗原 (Ani s2, 3),這些抗原對加熱、冷凍及胃酸環境的忍受 力高,經過煮沸及胃酸處理後仍然可以被IgE 抗體辨別而產生過敏反應 (Caballero and Moneo, 2004; Baird et al., 2014)。
1.4.4 安尼線蟲科之鑑定
安尼線蟲之第三期幼蟲,依照其消化道形態、頭端是否具有用來鑽孔的齒脊 (boring tooth)、排泄孔 (excretory pore) 位置,以及尾端是否有棘刺 (mucron) 等特 徵來進行鑑定。
海 獸 胃 線 蟲 屬 (Anisakis) 幼 蟲 , 頭 端 具 有 突 出 的 齒 脊 , 明 顯 膨 大 的 胃 (ventriculus),且沒有延伸的胃盲囊 (ventricular appendage) 及腸盲囊 (intestinal caecum) (張等,1999;Shih, 2004)。接著根據尾端形態及棘刺有無可細分為 Type I-IV 四種形態。Type I 為典型海獸胃線蟲形態,胃較 Type II-IV 長,尾端具有棘刺,包 含了 A. simplex s.s.、 A. pegreffii、A. simplex C、A. typica、A. ziphidarum 和 A. nascettii。
Type II-IV 尾端皆沒有棘刺,胃相對較短且形狀似梯形,三種 Type 的尾端形態也 有差異;Type II 尾端較長 (0.16-0.38 mm),呈圓錐形,沒有棘刺,經分子鑑定後 確認為 A. physeteris。Type III 尾端形狀短圓 (0.11-0.15 mm),大部分沒有棘刺,極 少部分具有極小的棘刺,其物種為 A. brevispiculata。Type IV 尾端短 (0.07-0.17 mm),
為圓錐形,其物種為 A. paggiae (Murata et al., 2011)。
8
對盲囊線蟲屬 (Contracaecum) 幼蟲,不具有齒脊,具有向前延伸的腸盲囊及 向後延伸的胃盲囊,尾端具有突出的小棘 (spine),排泄孔位於前端,靠近唇的基 部 (張等,1999;施,2001)。
宮脂線蟲屬 (Hysterothylacium) 幼蟲,與對盲囊線蟲屬相似,同樣不具有齒脊,
具有腸盲囊及胃盲囊,尾端具有突出的小棘,其排泄孔位置靠近神經環 (nerve ring),
或在神經環稍前處,與對盲囊線蟲屬相異 (張等,1999;施,2001;Shih, 2004)。
前盲囊線蟲屬 (Porrocaecum) 幼蟲,具有齒脊,具有向前延伸之腸盲囊,無 胃盲囊,尾端無棘刺 (張等,1999;Shih, 2004)。
然而,形態鑑定法在蟲種間形態極為相似、無法取得良好視角、或是蟲體有 破損時,往往無法準確判定海獸胃線蟲之蟲種,因此利用分子生物學的方法來鑑 定安尼線蟲科之蟲種越來越普遍。
在分子鑑定時,最常被使用的是DNA 片段中的 rDNA ITS 片段,包含 ITS 1、
5.8S rRNA 及 ITS 2,以及 mitochondrial DNA cytochrome c oxidase subunit II (cox2) 序列;先利用Polymerase chain reaction (PCR) 擴增 DNA 序列,直接進行定序比較 核苷酸序列,或是使用不同方法處理DNA 片段,如 PCR-Single strand conformation polymorphism (SSCP),將 PCR 擴增的產物經變性後形成單股 DNA,再因其核酸序 列的差別而形成不同的構形,影響電泳時的泳動速率 (Zhu et al, 2007);或是 PCR-Restriction fragment length polymorphism (RFLP),利用核酸限制酶針對特定序 列的切位,將PCR 增幅後的核酸片段切成不同長度的片段 (Abollo et al., 2003)。
也可以直接將DNA 萃取出來後,加入多個引子進行 PCR,稱為 Random amplified polymorphic DNA (RAPD)-PCR,利用核酸序列差異導致引子結合位不同而增幅出
不同長度的核苷酸片段 (Martin-Sanchez et al., 2005)。
9
1.5 盲鰻
1.5.1 分類地位
盲鰻有著古老脊椎動物的特徵,與現生的脊椎動物有著共同的祖先,在脊椎 動物的演化上佔有重要的地位。全世界已知的盲鰻共有六屬七十八種,而臺灣地 區盲鰻科總計有三屬十三種,其中包括黏盲鰻屬 (Genus Eptatretus) 9 種,副盲鰻 屬(Genus Paramyxine) 2 種,及盲鰻屬 (Genus Myxine) 2 種 (邵,2013)。
本研究之盲鰻分類如下:
脊索動物門 (Phylum Chordata) 盲鰻綱 (Class Myxini)
盲鰻目 (Order Myxiniformes) 盲鰻科 (Family Myxinidae)
黏盲鰻屬 (Genus Eptatretus) 盲鰻屬 (Genus Myxine) 線盲鰻屬 (Genus Nemamyxine) 新盲鰻屬 (Genus Neomyxine) 南盲鰻屬 (Genus Notomyxine) 副盲鰻屬 (Genus Paramyxine)
1.5.2 形態特徵
盲鰻有著似鰻魚般的圓筒狀身軀,沒有鰭及硬骨,只有由軟骨形成的頭骨。
頭部有一對具感光功能的眼點,無視覺影像;口部為漏斗狀且沒有上下頷,口內 有兩對角質化的舌齒。吻端有兩對鬚,口側有一到二對鬚;身體腹面有成對的鰓 孔,體側各有一列黏液孔,黏液孔由眼睛後方延伸至尾部,依照分布區域可分為
10
鰓孔前黏液孔、鰓孔上黏液孔、軀幹黏液孔,以及尾部黏液孔,其鰓孔形態與黏 液孔數量是辨別盲鰻物種重要的特徵。
1.5.3 生態習性
盲鰻主要棲息在海洋底層,屬於底棲性魚類,是海洋底棲魚類中數量最多的。
因為視覺的退化,盲鰻主要依賴靈敏的嗅覺及吻部的觸鬚覓食,利用舌齒的外翻 內收撕取食物,也會利用黏液使獵物窒息死亡後再進食。盲鰻為海裡主要的腐食 者,一般以死亡的底棲生物或是死亡的魚類、海洋哺乳類為主食,扮演著海洋清 道夫的角色,而盲鰻在覓食、挖洞的同時,也可以促進底部的物質翻轉。而盲鰻 及盲鰻的卵也是其他魚類、海洋哺乳類或是大型無脊椎動物的食物,在受到掠食 者攻擊時,身上黏液孔可以排出大量黏液來保護自己,進而從掠食者口中逃脫。
目前已知盲鰻的掠食者有鱈魚、鎧鯊、貓鯊、糯鰻、鼬鳚、美洲多鋸鱸、南極櫛 鯧、白斑角鯊、港海豹、鼠海豚、海獅、海象、章魚等 (Jorgensen et al., 1998; Zintzen et al., 2011)。
盲鰻沒有耳石及硬骨組織,目前仍沒有辦法得知盲鰻生活史長度及實際年齡。
同時盲鰻並無特定的產卵季節,在雌性體內全年都有發現卵,可能為分批成熟,
一年中可產多次卵 (陳,2004;Barss, 1993),已知臺灣西南海域的紐氏黏盲鰻可能 在六月底至七月有一產卵季 (洪,2003)。產下的卵根據物種不同,大約為 14~25mm 長,卵的兩端有成叢的錨鉤,可使卵互相鉤連並固定於海底。
1.5.4 地理分布
盲鰻在全球分布範圍包含了太平洋、大西洋、印度洋、北極海和南極海域,
在地中海、白令海和加勒比海也有盲鰻分布,而限制盲鰻分布的主要因素有鹽度、
溫度、深度及底質。盲鰻主要棲息於一般鹽度的海水 (34 ppt),以及沙、泥等可挖
11
掘的底質,也有部分會盤曲於岩石之間,以利於躲避掠食者。根據物種不同,其 最適棲息溫度可從最低0℃至20℃,而深度可從20公尺至5000公尺 (Jorgensen et al., 1998)。
臺灣的盲鰻主要分布在臺灣北部、東北部海域,以及高雄西南海域。根據文 獻紀錄 (Kuo et al., 1994; Mok and Chen, 2001),臺灣北部及東北部海域有捕獲過楊 氏黏盲鰻、蒲氏黏盲鰻、沈氏黏盲鰻、臺灣黏盲鰻、紫黏盲鰻、紅尾黏盲鰻 (Kuo et al., 2010);西部及西南海域有楊氏黏盲鰻、陳氏黏盲鰻、中華黏盲鰻 (Kuo and Mok, 1994)、紐氏黏盲鰻、紫黏盲鰻、沈氏黏盲鰻、臺灣盲鰻 (Mok and Kuo, 2001)、
郭氏盲鰻 (Mok, 2002)、費氏副盲鰻;以及台東富岡海域捕獲的懷氏副盲鰻。
1.5.5 盲鰻利用現況
盲鰻在臺灣過去被視為無經濟價值的魚種,主要以底拖網方式混獲,捕獲地 區以宜蘭大溪漁港及屏東東港漁港為主。但隨著漁業資源下降,越來越多人開始 食用盲鰻,因此在屏東東港也開始利用盲鰻筒專門捕撈盲鰻。盲鰻筒有著易進難 出的漏斗狀開口的長筒,筒內置入鰹魚或花腹鯖等餌料,吸引盲鰻進入 (吳,2005)。
捕撈的盲鰻在經過剝皮、去除內臟後,餘下的盲鰻肉則直接販售至海產店,俗稱
「龍筋」(賴,2008),因盲鰻並非主要經濟魚種,其捕撈量量、產值亦沒有相關記 錄。在中國、日本、韓國也都有食用盲鰻,而盲鰻在韓國更是著名的料理,具有 非常多樣化的料理方式,甚至成為許多觀光客必吃的美食。此外,大量的盲鰻被 捕捉後送往南韓,這些盲鰻的皮取下後經加工處理製成「鰻皮」,被大量製成皮 夾、皮帶等飾品 (Barss, 1993),然而韓國因為過度捕撈盲鰻,而使韓國本地的盲鰻 幾乎消失殆盡,現在則是大量從美國 (奧勒岡州) 進口盲鰻 (Dillman, 2013)。
另外,底拖網漁法會造成盲鰻的混獲及棲地的破壞。在國際自然保護聯盟 (IUCN) 紅色名錄中 (Mincarone and Mok, 2013),臺灣黏盲鰻被列為瀕危物種
12
(Endangered),因臺灣黏盲鰻只分佈於臺灣東北海域,分佈深度淺,同時受到底拖 網漁法造成嚴重的混獲及海底棲地的破壞;蒲氏黏盲鰻在東中國海屬於常見的盲 鰻物種,但也被列為近危物種 (Near Threatened),因其體型相對較大,加上食用及 鰻魚皮的需求而過度的漁撈,造成蒲氏黏盲鰻的數量驟減;楊氏黏盲鰻與沈氏黏 盲鰻雖然列為無危物種 (Least Concern),但其分佈範圍同樣限於臺灣週遭海域,
也同樣受到底拖網造成的混獲及棲地破壞。近年對盲鰻的需求越來越高,不論是 食用安全方面或是資源管理都應該受到重視。
1.5.6 相關研究
在過去盲鰻寄生蟲研究中,以盲鰻為寄主的寄生蟲有四種單殖吸蟲及兩種橈 足類。四種單殖吸蟲皆屬於單殖吸蟲綱 (Class Monogenea),Acanthocotylidae 科,
Myxinidocotylinae 亞科,三種 Myxinidocotyle 屬和一種 Lophocotyle 屬。單殖吸蟲 寄生於盲鰻體表,呈橢圓扁平狀,寄生蟲長度約2 mm,尾端有一個卵圓形的偽固 著器。M. eptatreti 被發現寄生於南非一處水族館飼養的六鰓黏盲鰻 (E. hexatrema) 體表(Vaughan and Christion, 2010);M. californica 則是於美國加州一處水族館中,
一尾新捕獲到太平洋黏盲鰻 (Eptatretus stoutii)身上發現寄生,且隨後快速感染該 水族缸的其他盲鰻;M. japonica 被發現寄生於蒲氏黏盲鰻 (E. burgeri),且在水族 箱水溫升高至17-18℃時出現大量感染;L. novaezeelandica 被發現寄生於新西蘭黏 盲鰻 (E. cirrhatus),同樣好發於水族箱水溫升高時 (Malmberg and Fernholm, 1989)。
Izawa (1996) 在日本熊野海域捕獲的紫黏盲鰻 (E. okinoseanus) 及紫盲鰻 (Myxine garmani) 發現一種橈足類寄生,該物種寄生於盲鰻之鰓囊,其屬顎足綱 (Class Maxillopoda) , 橈 足 亞 綱 (Subclass Copepoda) , 管 口 目 (Order Siphonostomatoida),Archidactylinidae 科,Archidactylina myxinicola,該科只有一
13
屬一種,同時也是首次於發現寄生於盲鰻的橈足類。
1.6 研究目的
2013 年首次發現無頷總綱魚類,海七鰓鰻感染海獸胃線蟲,推測是七鰓鰻寄 生在鮭魚等寄主時,藉由寄生時造成的傷口感染海獸胃線蟲,而七鰓鰻在歐洲被 視為珍饈,具有很高的經濟價值,此篇報導也提出食用七鰓鰻而感染海獸胃蟲的 風險,同時海獸胃線蟲也因七鰓鰻溯河洄游的行為而有進入淡水水域的可能性 (Bao et al., 2013)。與七鰓鰻同樣為無頷總綱的盲鰻,是否能透過腐食的方式感染 海獸胃線蟲?至今,全世界對於盲鰻及盲鰻寄生蟲相關的研究非常貧乏。本篇研 究目的擬查明盲鰻是否感染海獸胃線蟲,及調查臺灣東北海域盲鰻的後生動物寄 生蟲相。分析 2013 至 2014 年間之採樣資料,以形態和分子技術鑑別寄生蟲種類 和寄生蟲感染情形,包括盛行率 (prevalence)、平均感染強度 (mean intensity)、平 均豐富度 (mean abundance) 等參數變動,以統計分析各項參數於不同盲鰻物種間、
性別及體長的關係。藉此了解海洋底棲盲鰻與寄生蟲感染間之交互作用,查明腐 生魚類盲鰻是否為海獸胃線蟲的保蟲寄主,並提供食用盲鰻食品安全衛生相關資 訊。
14
第 2 章 材料與方法
2.1 樣本取得
本實驗樣本來源為臺灣東北部宜蘭大溪漁港,漁法為底拖網,作業範圍於龜 山島附近海域 (121˚ 94'E; 24˚ 84'N),捕撈深度於 100-300 m。委託當地漁民向有捕 獲盲鰻的船家收集後,以碎冰冷藏魚隻運送至實驗室。採樣時間為 2013 年 11 月 至2014 年 6 月,共採樣 6 次,樣本總數 265 尾盲鰻,其中包含 66 尾蒲氏黏盲鰻 (Eptatretus burgeri)、96 尾楊氏黏盲鰻 (E. yangi)、81 尾沈氏黏盲鰻 (E. sheni)和 22 尾臺灣黏盲鰻 (E. taiwanae)。
盲鰻在物種分辨上,主要依據體色、體長,鰓孔的數量及特徵,以及身體兩 側的黏液孔數量來辨別盲鰻物種,黏液孔又可以依部位分為鰓孔前黏液孔、鰓孔 上黏液孔、軀幹黏液孔、尾部黏液孔 (沈、吳,2011;Kuo et al., 1994)。
圖 2 盲鰻黏液孔分佈 (Kuo et al., 1994)
本次研究共採集到四種盲鰻。蒲氏黏盲鰻,體色為淡褐色,體型最大可達60 cm,
背部中央有一條白色帶,鰓孔每側 6 個,成縱線排列,左側最後一個鰓孔大於其 餘鰓孔 (圖 3)。鰓孔前黏液孔數量為 18-21 個,鰓孔上黏液孔為 4-6 個,軀幹黏液 孔為46-51 個,尾部黏液孔為 11-12 個,黏液孔總數為 79-90 個 (沈、吳,2011;
邵,2013)。
楊氏黏盲鰻,體色為淡紫灰色,最大體長約30 cm,鰓孔每側 5 個,聚集成群,
鰓孔前黏液孔 鰓孔及
鰓孔上黏液孔 軀幹黏液孔 尾部黏液孔
15
呈不規則分布,左側最後一個鰓孔大於其餘鰓孔 (圖 3)。鰓孔前黏液孔數量為 16-23 個,無鰓孔上黏液孔,軀幹黏液孔為42-47 個,尾部黏液孔為 8-11 個,黏液孔總 數為66-78 個 (沈、吳,2011;邵,2013)。
沈氏黏盲鰻,體色為灰褐色,最大體長46 cm,背部無白線,鰓孔每側 6 個,
呈縱線排列,排列間距較小,且為鰓孔周圍為明顯白色,左側最後一個鰓孔大於 其餘鰓孔 (圖 3),且背部無白色帶。鰓孔前黏液孔數量為 13-18 個,鰓孔上黏液孔 為 0-2 個,軀幹黏液孔為 39-46 個,尾部黏液孔為 8-12 個,黏液孔總數為 64-74 個 (沈、吳,2011;邵,2013)。
臺灣黏盲鰻,體色為淡紫灰色,腹部偏白,鰓孔每側 6 個,聚集成群,呈不 規則分布,左側最後一個鰓孔大於其餘鰓孔 (圖 3)。鰓孔前黏液孔數量為 16-19 個,
無鰓孔上黏液孔,軀幹黏液孔為36-42 個,尾部黏液孔為 6-9 個,黏液孔總數為 62-67 個 (沈、吳,2011;邵,2013)。
2.2 樣本處理
樣本解凍後,進行編號並測量其全長、體重 (圖 4),以肉眼及解剖顯微鏡輔助 觀察是否有體外寄生蟲。先從口部注水洗出消化道內容物於燒杯中,觀察是否寄 生蟲被洗出,再以解剖剪從泄殖孔直線向頭端剪開體腔,露出肌肉、消化道及內 臟,觀察是否有寄生蟲存在並記錄性別。雌性盲鰻的卵巢分佈於整個腸繫膜右側;
雄性盲鰻的精巢分佈於體腔右側後三分之一部分。最後,將消化道從體腔分離取 下並置於燒杯中漂洗,剪開消化道並置於燈箱上觀察消化道內壁。檢獲之寄生蟲 則利用解剖顯微鏡根據其形態特徵初步鑑定寄生蟲種類,接著保存於70%酒精中,
進行後續種類鑑定。
16
2.3 安尼線蟲科線蟲之 PCR-RFLP 鑑種
經初步鑑定後確定為安尼線蟲科之線蟲,從 70%酒精取出並進行 PCR-RFLP 分子鑑定,包含Genomic DNA 萃取、PCR 增幅及 RFLP 分析。
2.3.1 Genomic DNA 萃取
材料與儀器:
液態氮、Geno Plus™ Genomic DNA Extraction Miniprep System (VIOGENE)、60℃
與70℃水浴槽、離心機
方法:
(1) 將保存於 70%酒精之線蟲樣本置於純水清洗。
(2) 蟲體移入 1.5 ml 微量離心管,加入液態氮磨碎。
(3) 加入 180 µl LYS buffer 均質。
(4) 加入 Proteinase K 20 µl,快速震盪。
(5) 置於 60℃水浴槽 30 分鐘,每 10 分鐘上下翻轉 10 次。
(6) 加入 300 µl Fx buffer,快速震盪。
(7) 置於 70℃水浴槽 20 分鐘,同時預熱 Elution buffer。
(8) 6000×g 離心 2 分鐘,取上清液於裝有 200 µl 之微量離心管,快速震盪。
(9) 將混合液全數移入 Mini column 中,並裝入收集管,6000×g 離心 2 分鐘。
(10) 更換新的收集管,加入500 µl WS buffer,6000×g 離心 2 分鐘。
(11) 移除收集管中液體,加入500 µl WS buffer,6000×g 離心 2 分鐘。
(12) 移除收集管中液體,10000×g 離心 2 分鐘。
(13) 將Mini column 裝入 1.5 ml 微量離心管,加入 100 µl Elution buffer,靜置 5 分鐘。
17
(14) 10000×g 離心 2 分鐘,得到含 Genomic DNA 之溶液,保存於-20℃。
2.3.2 PCR 增幅
以Forward premer NC5、Reverse premer NC2 進行 PCR,增幅 rDNA 中 ITS 1、
5.8S rDNA、ITS 2 片段,約 1000 bp。
(1)
將下表所列加入200 µl 微量離心管中。名稱 濃度 體積 (µl)
10X Ex Taq Buffer 10× 5
dNTP 2.5 mM 4
Ex Taq 5 U/µl 0.25
Forward primer 10 µM 1
Reverse primer 10 µM 1
DNA template 100 ng/ml 15
ddH2O 23.75
Total 50
Forward primer NC5: 5’-GTAGGTGAACCTGCGGAAGGATCATT-3’
Reverse primer NC2: 5’-TTAGTTTCTTTTCCTCCGCT-3’
(2)
在 95℃下進行 denaturation 10 分鐘;接著進行 35 個循環包括:95℃ 30 秒(denaturation)、55℃ 30 秒 (annealing)、72℃ 75 秒 (extension);最後在 72℃
下進行extension 10 分鐘。
(3)
於加入GelRed™ Nucleic Acid Stain 之 2% agarose gel 進行電泳。(4)
以紫外線燈箱照射觀察,並照相記錄。18
2.3.3 RFLP 分析
利用 HinfI、HhaI 兩種核酸限制酶酵素進行 RFLP 分析。
(1)
將下表所列加入200 µl 微量離心管中。名稱 體積 (ul)
10X Digest buffer 2
PCR 產物 10
Enzyme (HinfI / HhaI) 1
ddH2O 17
Total 30
(2)
37℃乾浴 1 小時後。(3)
於加入GelRed™ Nucleic Acid Stain 之 1.5% agarose gel 進行電泳。(4)
以紫外線燈照射觀察,並照相記錄。2.4 感染參數之分析
根據Bush et al. (1997) 之定義計算盛行率 (Prevalence)、平均感染強度 (Mean intensity) 與平均豐富度 (Mean abundance):
(1)
盛行率 (Prevalence):所有採樣寄主中,感染特定寄生蟲之數量百分比。盛行率 = (感染特定寄生蟲之寄主數 ÷ 所有採樣之寄主數) × 100%
(2)
平均感染強度 (Mean intensity):所有感染特定寄生蟲的寄主中,特定寄生蟲之平均感染數。
平均感染強度 = 特定寄生蟲之總數 ÷ 受特定寄生蟲感染之寄主數
(3)
平均豐富度 (Mean abundance):所有採樣寄主中,特定寄生蟲之平均感染數。19
平均豐富度 = 特定寄生蟲之總數 ÷ 所有採樣寄主數
使用軟體R 與 Microsoft Excel 進行統計分析,因無法得知野外盲鰻之寄生蟲 感染參數是否為常態分配,且樣本數量小,故選擇Chi-Square test 來檢測盲鰻物種 與 盛 行 率 之 相 關 性 , 以 及 盲 鰻 在 不 同 性 別 間 盛 行 率 相 關 性 。 以 無 母 數 分 析 Wilcoxon-Mann-Whitney rank sum test 檢測感染強度與豐富度在不同盲鰻物種間,
以及在盲鰻性別間的差異。另外,四種盲鰻之海獸胃線蟲蟲種百分比也利用 Chi-Square test 檢測盲鳗體內之海獸胃線蟲蟲種是否有差異。
盲鰻體長與寄生蟲盛行率分析中,以體長為自變數,感染參數為依變數,進 行邏輯迴歸分析;體長進行分組計算後,使用皮爾森相關係數 (Pearson correlation coefficient)及簡單直線迴歸分析。體長與豐富度及感染強度同樣以體長為自變數,
感染參數為依變數,使用皮爾森相關係數及簡單直線迴歸分析。
在皮爾森相關係數中,定義R 值於 0 ~ 0.1 為無相關 (0 ≤ R2 < 0.01) 、R 值於 0.1 ~ 0.4 為低度相關 (0.01 ≤ R2 < 0.16)、R 值於 0.4 ~ 0.7 為中度相關 (0.16 ≤ R2 <
0.49)、R 值於 0.7 ~ 1 為高度相關 (0.49 ≤ R2 < 1)、R 值於 1 為完全相關 (R2 = 1)。
20
第 3 章 結果
3.1 盲鰻之寄生蟲相
265 尾盲鰻體長範圍為 16.2-55.9 cm;體重範圍為 9.70-274.95 g。分別為蒲氏 黏盲鰻66 尾,平均體長 37.8 ± 7.0 cm,平均體重 101.67 ± 56.16 g;楊氏黏盲鰻 96 尾,平均體長26.1 ± 2.9 cm,平均體重 32.71 ± 8.92g;沈氏黏盲鰻 81 尾,平均體 長32.4 ± 6.0 cm,平均體重 85.74 ± 42.75g;臺灣黏盲鰻 22 尾,平均體長為 27.1 ± 3.7 cm,平均體重為 51.89 ± 21.05g (圖 5)。
此次研究首次發現海獸胃線蟲屬線蟲和盲鰻刺魚蚤寄生於盲鰻。海獸胃線蟲 主要發現於消化道內壁,從消化道表面可以觀察到不規則凸起或是紅斑的現象,
用鑷子撕開後,則可以看見線蟲嵌入其中;而極少部分的線蟲可以鑽出消化道,
移動到腹腔之中,或是寄生於腹腔兩側之肌肉 (圖 6)。在實驗期間,發現一種寄生 於 盲 鰻 咽 喉 處 的 橈 足 類 , 經 過 鑑 定 後 , 確 認 為 橈 足 類 「 盲 鰻 刺 魚 蚤 」 (Acanthochondria eptatreti),屬於劍水蚤目 (Order Cyclopoida),軟刺魚蚤科 (Family Chondracanthidae),刺魚蚤屬 (Genus Acanthochondria)。盲鰻刺魚蚤雌蟲體長約 1 cm,尾端帶著兩條與約與體長相等的卵囊,而雄蟲體型極小,約 1 mm,附著於雌 蟲的尾端。被盲鰻刺魚蚤寄生的盲鰻,也可以從咽喉部外觀看見紅腫突起,剪開 咽喉部位則可看到盲鰻刺魚蚤寄生咽喉內側 (圖 7)。而消化道洗出物中可以觀察到 魚骨、魚鱗、蝦殼及附肢,未完全消化的小魚、小蝦及小管等頭足類,以及部分 線蟲。
3.2 海獸胃線蟲之 PCR-RFLP 鑑種結果
本實驗取 106 隻線蟲樣本進行 PCR-RFLP 實驗,藉此鑑定其物種。經過兩種 限制酶切割後得到7 種圖譜 (圖 8、9),根據過去文獻 (Abollo et al., 2003; D'Amelio
21
et al., 2000; Quiazon et al., 2013) 得知:(1)為 A. pegreffii,以 HinfI 作用後得到 370 bp、
300 bp 及 250 bp 三個片段,HhaI 作用後得到 550 bp 及 430 bp 兩個片段;(2)為 A.
simplex s.s.,以 HinfI 作用後得到 620 bp 及 250 bp 兩個片段,HhaI 作用後得到 550 bp 及 430 bp 兩個片段;(3)為 A. pegreffii 與 A. simplex s.s.的重組種,以 HinfI 作用 後得到620 bp、370 bp、300 bp 及 250 bp 四個片段,HhaI 作用後得到 550 bp 及 430 bp 兩個片段;(4)為 A. typica,以 HinfI 作用後得到 620 bp 及 350 bp 兩個片段,HhaI 作用後得到310 bp、210 bp、180 bp、150 bp 及 100 bp 五個片段 (表 1)。
(5)、(6)、(7) 三種同時利用 PCR 產物定序,並與資料庫 (NCBI) 核酸序列比 對確認物種。(5)為 Anisakis sp. (KC342894),以 HinfI 作用後得到 900 bp 一個片段,
HhaI 作用後得到 360 bp、210 bp、160 bp 及 110 bp 四個片段;(6)為 A. brevispiculata (KC342887),以 HinfI 作用後得到 900 bp 一個片段,HhaI 作用後得到 390 bp、330 bp 及 190 bp 三個片段;(7)為 H. amoyense (KP252133),以 HinfI 作用後得到 660 bp 及350 bp 兩個片段,HhaI 作用後得到 360 bp、330 bp、130 bp 及 80 bp 四個片段 (表 1)。
106 隻線蟲樣本中,檢獲寄生於蒲氏黏盲鰻的線蟲 57 隻,包含 51 隻 A. pegreffii、
1 隻 A. typica、1 隻 A. brevispiculata、3 隻 H. amoyense、及 1 隻 A. pegreffii 與 A. simplex s.s.的重組基因型。檢獲寄生於楊氏黏盲鰻的線蟲 15 隻,包含 12 隻 A. pegreffii 及 3 隻 Anisakis sp.。檢獲寄生於沈氏黏盲鰻的線蟲 31 隻,包含 5 隻 A. pegreffii、1 隻 A. simplex s.s.及 25 隻 Anisakis sp.。檢獲寄生於臺灣黏盲鰻的線蟲 3 隻,皆為 A.
pegreffii。
因四種盲鰻皆有 A. pegreffii 感染,因此利用卡方檢定比較 A. pegreffii 在四種盲 鰻體內之海獸胃線蟲感染百分比,A. pegreffii 於蒲氏黏盲鳗之海獸胃線蟲百分比為 89% (51/57)、於楊氏黏盲鳗為 80% (12/15)、於沈氏黏盲鰻為 16% (5/31)、於臺灣 黏盲鳗100% (3/3);結果顯示 A. pegreffii 於沈氏黏盲鰻之海獸胃線蟲百分比明顯低
22
於其他三種盲鰻 (p < 0.05) (圖 10)。
3.3 感染參數之分析結果
3.3.1 四種盲鰻感染參數之比較分析
265 尾盲鰻經解剖檢查後,共檢出 307 隻線蟲,57 尾盲鰻遭感染,主要寄生 於消化道壁,部分於消化道洗出物中檢獲,極少部分會進入到體腔或是進入肌肉 中;2 尾蒲氏黏盲鰻各在肌肉表面檢獲 1 隻線蟲包囊,1 尾楊氏黏盲鰻同樣在肌肉 表面檢出1 隻線蟲包囊,1 尾沈氏黏盲鰻於肌肉檢出 1 隻線蟲包囊及 1 隻線蟲游離 於體腔。
整體線蟲盛行率為 21.51%,平均感染強度為 5.39 隻蟲/尾魚,平均豐富度為 1.16 隻蟲/尾魚。各別盲鰻物種中,66 尾蒲氏黏盲鰻共檢出線蟲 252 隻,盛行率為 48.48%,平均感染強度 7.88 隻蟲/尾魚,平均豐富度 3.82 隻蟲/尾魚,其中一尾蒲 氏黏盲鰻感染134 隻線蟲;96 尾楊氏黏盲鰻共檢出線蟲 16 隻,盛行率 9.38%,平 均感染強度1.78 隻蟲/尾魚,平均豐富度 0.17 隻蟲/尾魚;81 尾沈氏黏盲鰻共檢出 線蟲33 隻,盛行率 14.81%,平均感染強度 2.75 隻蟲/尾魚,平均豐富度 0.41 隻蟲 /尾魚;22 尾臺灣黏盲鰻檢出線蟲 6 隻,盛行率 18.18%,平均感染強度 1.5 隻蟲/
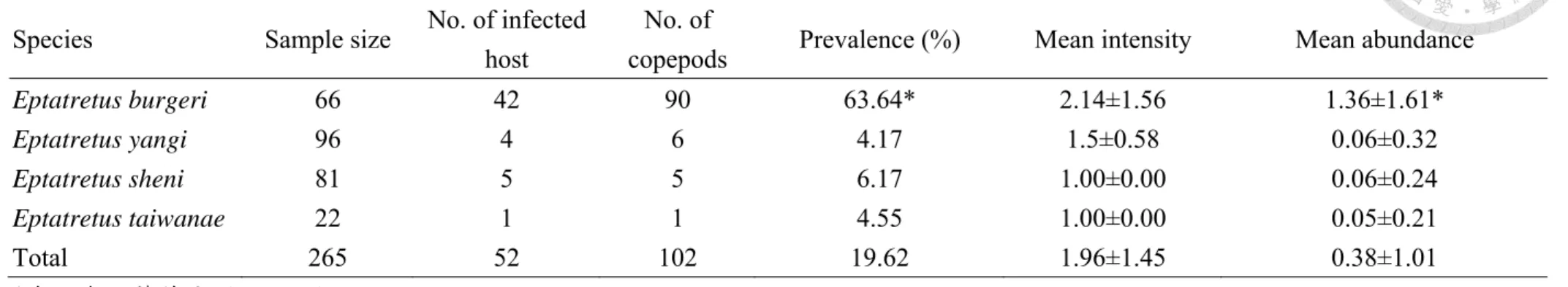
尾魚,平均豐富度0.27 隻蟲/尾魚 (表 2)。
蒲氏黏盲鰻之線蟲盛行率 (48.48%) 顯著高於楊氏黏盲鳗 (9.38%)、沈氏黏盲 鰻 (14.81%) 及臺灣黏盲鳗 (18.18%)(p < 0.05)。線蟲感染強度中,四種盲鰻之間 並無顯著差異,而蒲氏黏盲鰻之線蟲豐富度 (3.82 隻蟲/尾魚) 顯著高於楊氏黏盲 鰻 (0.17 隻蟲/尾魚)、沈氏黏盲鰻 (0.41 隻蟲/尾魚)、臺灣黏盲鰻 (0.27 隻蟲/尾魚)(p
< 0.05),其餘盲鰻物種之線蟲感染參數均無顯著差異 (表 2)。
解剖檢查265 尾盲鰻其中 52 尾盲鰻遭感染,共檢獲 102 隻盲鰻刺魚蚤,盛行
23
率為19.62%,平均感染強度為 1.96 隻蟲/尾魚,平均豐富度為 0.38 隻蟲/尾魚。各 別盲鰻物種中,蒲氏黏盲鰻檢出盲鰻刺魚蚤數90 隻,盛行率 63.64%,平均感染強 度2.14 隻蟲/尾魚,平均豐富度 1.36 隻蟲/尾魚;楊氏黏盲鰻檢出盲鰻刺魚蚤 6 隻,
盛行率4.17%,平均感染強度 1.50 隻蟲/尾魚,平均豐富度 0.06 隻蟲/尾魚;沈氏黏 盲鰻檢出盲鰻刺魚蚤數5 隻,盛行率 6.17%,平均感染強度 1.00 隻蟲/尾魚,平均 豐富度0.06 隻蟲/尾魚;臺灣黏盲鰻檢出盲鰻刺魚蚤 1 隻,盛行率 4.55%,平均感 染強度1.00 隻蟲/尾魚,平均豐富度 0.05 隻蟲/尾魚 (表 3)。
盲鰻刺魚蚤之感染參數分析中,蒲氏黏盲鰻之盲鰻刺魚蚤盛行率 (63.64%) 顯 著高於其他三種盲鰻 (p < 0.05),盲鰻刺魚蚤之感染強度在各盲鰻物種間皆無顯著 差異,其中沈氏黏盲鰻與臺灣黏盲鰻隻感染強度皆為1,故無法比較;在盲鰻刺魚 蚤之豐富度檢定中,蒲氏黏盲鰻之盲鰻刺魚蚤豐富度 (1.36 隻蟲/尾魚) 顯著高於 其他三種盲鰻 (p < 0.05),其餘物種之間並無顯著差異 (表 3)。
3.3.2 盲鰻性別分組與感染參數之比較分析
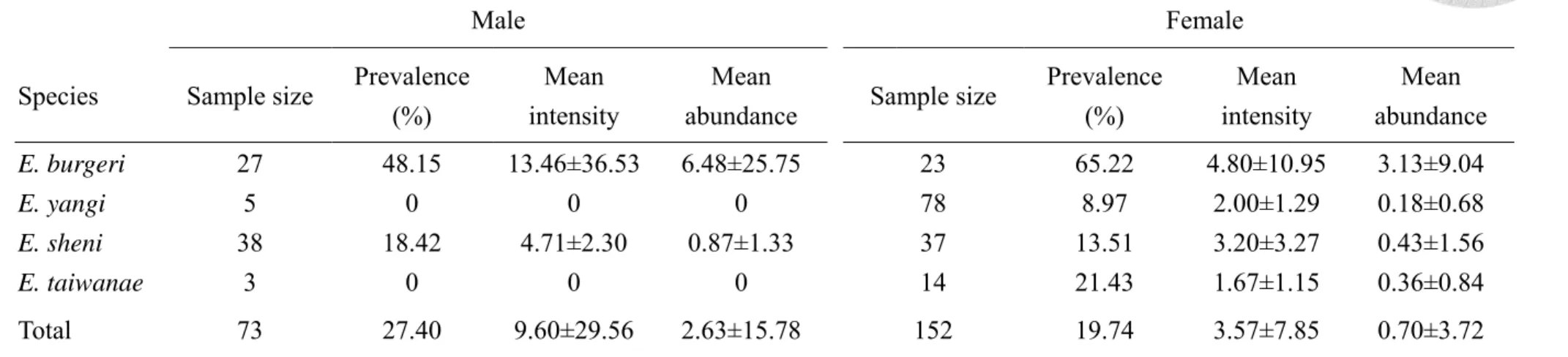
盲鰻以性別分組後計算各項感染參數,其中40 尾無法經生殖線判斷性別之盲 鰻則不列入計算。雄性盲鰻 (Male) 共 73 尾,其線蟲盛行率為 27.4%,線蟲平均 感染強度為9.6 隻蟲/尾魚,線蟲平均豐富度為 2.63 隻蟲/尾魚;雌性盲鰻 (Female) 共152 尾,線蟲盛行率為 19.74%,線蟲平均感染強度為 3.57 隻蟲/尾魚,線蟲平均 豐富度為0.7 隻蟲/尾魚。各項感染參數於盲鰻性別間均無顯著差異 (表 4)。
在個別盲鰻物種中,27 尾雄性蒲氏黏盲鰻,線蟲盛行率為 48.15%,平均感染 強度13.46 隻蟲/尾魚,平均豐富度 6.48 隻蟲/尾魚;23 尾雌性蒲氏黏盲鰻,線蟲盛 行率為65.22%,平均感染強度 4.80 隻蟲/尾魚,平均豐富度 3.13 隻蟲/尾魚。各項 感染參數於蒲氏黏盲鰻性別間均無顯著差異。
24
5 尾雄性楊氏黏盲鰻,線蟲盛行率為 0%,平均感染強度和平均豐富度皆為 0 隻蟲/尾魚;78 尾雌性楊氏黏盲鰻,線蟲盛行率為 8.97%,平均感染強度 2.00 隻蟲 /尾魚,平均豐富度 0.18 隻蟲/尾魚。雄性楊氏黏盲鰻感染參數皆為 0,無法進行分 析。
38 尾雄性沈氏黏盲鰻,線蟲盛行率為 18.42%,平均感染強度 4.71 隻蟲/尾魚,
平均豐富度 0.87 隻蟲/尾魚;37 尾雌性沈氏黏盲鰻,線蟲盛行率為 13.51%,平均 感染強度 3.20 隻蟲/尾魚,平均豐富度 0.43 隻蟲/尾魚。各項感染參數於沈氏黏盲 鰻性別間均無顯著差異。
3 尾雄性臺灣黏盲鰻,線蟲盛行率為 0%,平均感染強度和平均豐富度皆為 0 隻蟲/尾魚;14 尾雌性臺灣黏盲鰻,線蟲盛行率為 21.43%,平均感染強度 1.67 隻 蟲/尾魚,平均豐富度 0.36 隻蟲/尾魚。雄性臺灣黏盲鰻感染參數皆為 0,無法進行 分析。
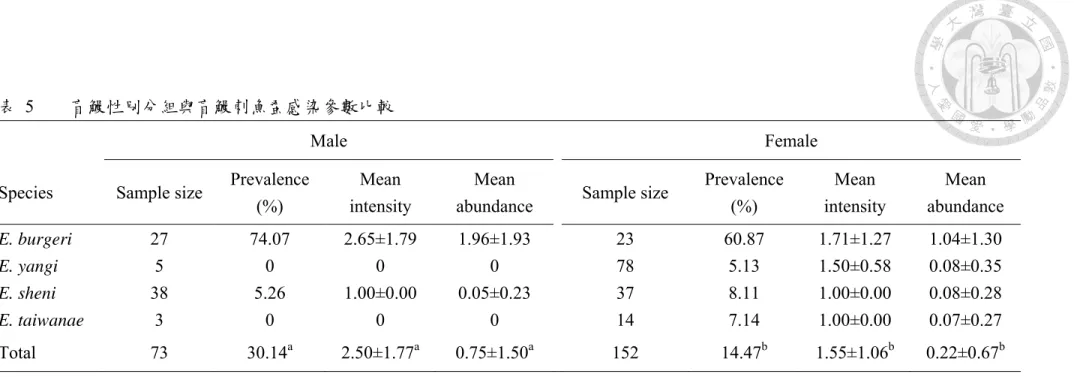
盲鰻刺魚蚤於盲鰻性別感染參數分析中 (表 5),雄性盲鰻共 73 尾,其盲鰻刺 魚蚤盛行率為30.14%,盲鰻刺魚蚤平均感染強度為 2.50 隻蟲/尾魚,盲鰻刺魚蚤平 均豐富度為0.75 隻蟲/尾魚;雌性共 152 尾,盲鰻刺魚蚤盛行率為 14.47%,盲鰻刺 魚蚤平均感染強度為 1.55 隻蟲/尾魚,盲鰻刺魚蚤平均豐富度為 0.22 隻蟲/尾魚,
雄性盲鰻之盲鰻刺魚蚤盛行率、感染強度、豐富度皆顯著高於雌性盲鰻 (p <
0.05)。
在個別盲鰻物種中,27 尾雄性蒲氏黏盲鰻,盲鰻刺魚蚤盛行率為 74.07%,平 均感染強度為 2.65 隻蟲/尾魚,平均豐富度為 1.96 隻蟲/尾魚;雌性性蒲氏黏盲鰻 (Female) 共 23 尾,盲鰻刺魚蚤盛行率為 60.87%,平均感染強度為 1.71 隻蟲/尾魚,
平均豐富度為1.04 隻蟲/尾魚。各項感染參數於蒲氏黏盲鰻性別間均無顯著差異。
5 尾雄性楊氏黏盲鰻,盲鰻刺魚蚤盛行率為 0%,平均感染強度和平均豐富度
25
為0 隻蟲/尾魚;78 尾雌性楊氏黏盲鰻,盲鰻刺魚蚤盛行率為 5.13%,平均感染強 度為 1.50 隻蟲/尾魚,平均豐富度為 0.08 隻蟲/尾魚。雄性楊氏黏盲鰻感染參數皆 為0,無法進行分析。
38 尾雄性沈氏黏盲鰻,盲鰻刺魚蚤盛行率為 5.26%,平均感染強度為 1.00 隻 蟲/尾魚,平均豐富度為 0.05 隻蟲/尾魚;37 尾雌性沈氏黏盲鰻,盲鰻刺魚蚤盛行 率為8.11%,平均感染強度為 1.00 隻蟲/尾魚,平均豐富度為 0.08 隻蟲/尾魚。各項 感染參數於沈氏黏盲鰻性別間均無顯著差異。
3 尾雄性臺灣黏盲鰻,盲鰻刺魚蚤盛行率為 0%,平均感染強度為 0 隻蟲/尾魚,
平均豐富度為0 隻蟲/尾魚;14 尾雌性臺灣黏盲鰻,盲鰻刺魚蚤盛行率為 7.14%,
平均感染強度為 1.00 隻蟲/尾魚,平均豐富度為 0.07 隻蟲/尾魚。雄性臺灣黏盲鰻 感染參數皆為0,無法進行分析。
3.3.3 盲鰻體長與線蟲感染參數分析
線蟲盛行率與盲鰻體長以兩種方式進行分析。分別為邏輯迴歸,以及將體長 進行分組後進行直線迴歸。而線蟲感染強度及線蟲豐富度與盲鰻體長之關係則以 直線迴歸進行分析。
邏輯迴歸中,蒲氏黏盲鰻、楊氏黏盲鰻、臺灣黏盲鰻之體長與線蟲盛行率均 未達顯著性 (p > 0.05),無法使用邏輯迴歸進行預測。沈氏黏盲鰻之體長與線蟲盛 行率於邏輯迴歸測試中達顯著性 (p < 0.01),其勝算比 (odds ratio)為 1.17,意指當 體長增加1 cm,感染機率會增加原本的 17 % (圖 11)。所有盲鰻之體長與線蟲盛行 率進行邏輯迴歸分析,顯示盲鰻體長與線蟲盛行率達顯著正相關 (p < 0.001),勝 算比為1.15,當體長增加 1 cm,感染機率會增加原本的 15 % (圖 12)。
26
直線迴歸分析中,為確保各組樣本數皆大於1,因此將體長以 4 cm 進行分組,
計算各組線蟲盛行率,以組中點與盛行率進行直線迴歸分析。所有盲鰻體長分組,
分析結果顯示盲鰻體長與線蟲盛行率有顯著高度正相關 (p < 0.05, R2 = 0.9475)(圖 13)。四種盲鰻分組體長與與盛行率分析中,蒲氏黏盲鰻體長與線蟲之盛行率呈顯 著高度正相關 (p < 0.05, R2 = 0.6097);楊氏黏盲鰻體長與線蟲之盛行率為無相關 (p = 0.05, R2 = 0.8998)。沈氏黏盲鰻體長與線蟲盛行率呈現無相關 (p = 0.07, R2 = 0.4434)。臺灣黏盲鰻體長與線蟲盛行率呈顯著高度正相關 (p < 0.05, R2 = 0.96) (圖 14)。
盲鰻體長與線蟲感染強度以及線蟲豐富度以直線迴歸進行分析。分析所有盲 鰻體長和線蟲感染強度,結果為顯著中度正相關 (p < 0.01, R2 = 0.1416) (圖 15)。個 別盲鰻中,蒲氏黏盲鰻體長與線蟲感染強度為顯著中度正相關 (p < 0.05, R2 = 0.157);楊氏黏盲鰻體長與線蟲感染強度為無顯著相關 (p = 0.65, R2 = 0.0317);沈 氏黏盲鰻體長與線蟲感染強度為顯著中度正相關 (p < 0.05, R2 = 0.4209);臺灣黏盲 鰻體長與線蟲感染強度為無顯著相關 (p = 0.19, R2 = 0.6497) (圖 16)。
盲鰻體長與線蟲豐富度之相關性測試結果。所有盲鰻體長與線蟲豐富度,結 果為顯著低度正相關 (p < 0.001, R² = 0.075) (圖 17)。蒲氏黏盲鰻體長與線蟲豐富度 為顯著低度正相關 (p < 0.01, R² = 0.1023);楊氏黏盲鰻體長與線蟲豐富度為無顯著 相關 (p = 0.15, R² = 0.0217);沈氏黏盲鰻體長與線蟲豐富度為顯著低度正相關 (p <
0.001, R² = 0.1292);臺灣黏盲鰻體長與線蟲豐富度為無顯著相關 (p = 0.46, R² = 0.0278) (圖 18)。
3.3.4 盲鰻體長與盲鰻刺魚蚤感染參數分析
盲鰻刺魚蚤盛行率與盲鰻體長同樣以兩種方式進行分析。分別為邏輯迴歸,
以及將體長分組後進行直線迴歸。而盲鰻刺魚蚤感染強度及盲鰻刺魚蚤豐富度與
27
盲鰻體長之關係則以直線迴歸進行分析。
個別盲鰻物種體長與盲鰻刺魚蚤盛行率之邏輯迴歸分析,結果皆未達顯著性,
無法使用邏輯迴歸進行預測;然而,所有盲鰻體長與盲鰻刺魚蚤盛行率分析則為 顯著正相關 (p < 0.001),勝算比為 1.13,當體長增加 1 cm,感染機率會增加原本 的13 % (圖 19)。
在分組體長與盲鰻刺魚蚤盛行率之直線迴歸分析中,所有盲鰻體長以級距 4 cm 分組並進行分析,結果顯示盲鰻體長與盲鰻刺魚蚤盛行率有顯著高度正相關 (p
< 0.001, R2 = 0.9069) (圖 20)。蒲氏黏盲鰻體長與盲鰻刺魚蚤盛行率呈現高度負相關 (p < 0.05, R² = 0.5531);其餘三種盲鰻體長與盲鰻刺魚蚤盛行率皆無顯著相關,楊 氏黏盲鰻 (p = 0.88, R² = 0.0137)、沈氏黏盲鰻 (p = 0.05, R² = 0.4762)、臺灣黏盲鰻 (p = 0.74, R² = 0.0667) (圖 21)。
盲鰻體長與盲鰻刺魚蚤感染強度之相關性測試結果。所有盲鰻體長和盲鰻刺 魚蚤感染強度分析,結果為顯著中度正相關 (p < 0.001, R² = 0.2287) (圖 22)。蒲氏 黏盲鰻為顯著中度正相關 (p < 0.01, R² = 0.2542);楊氏黏盲鰻為無顯著相關 (p = 0.88, R² = 0.0144);沈氏黏盲鰻期感染強度皆為 1,結果顯示無相關;臺灣黏盲鰻 只有一尾樣本感染,故無法進行分析 (圖 23)。
盲鰻體長與盲鰻刺魚蚤豐富度之相關性測試結果。分析所有盲鰻體長和盲鰻 刺魚蚤感染強度,結果為顯著中度正相關 (p < 0.001, R² = 0.1685) (圖 24)。盲鰻刺 魚蚤之豐富度和個別四種盲鰻體長間皆無顯著相關,蒲氏黏盲鰻 (p = 0.05, R² = 0.0564)、楊氏黏盲鰻 (p = 0.53, R² = 0.0042)、沈氏黏盲鰻 (p = 0.16, R² = 0.0244)、
臺灣黏盲鰻 (p =0.51, R² = 0.0224)(圖 25)。
28
第 4 章 討論
4.1 盲鰻之寄生蟲相
本研究首次發現線蟲動物門海獸胃線蟲,以及節肢動物門盲鰻刺魚蚤寄生於 四種盲鰻。過去文獻研究紀載,寄生於盲鰻體表的四種Acanthocotylidae 科之單殖 吸 蟲 Myxinidocotyle eptatreti, M. californica, M. japonica and Lophocotyle novaezeelandica (Malmberg and Fernholm,1989; Vaughan and Christion, 2010),及寄 生於鰓囊的Archidactylinidae 科之橈足類 Archidactylina myxinicola (Izawa, 1996),
在本次研究中皆無發現。
四種單殖吸蟲皆於水族館養殖時發現,較高的養殖水溫加速蟲卵孵化週期,
而養殖空間狹隘、密閉,造成魚隻的緊迫、免疫力下降,可能使單殖吸蟲在寄主 間大量感染傳播 (Chen et al., 2010; Malmberg and Fernholm, 1989)。相對於野外環 境中,海水溫度穩定、族群密度低,因此在野外捕撈的盲鰻較難察覺單殖吸蟲的 寄生。另外,漁民捕獲盲鰻後,因離開海水許久,盲鰻大多已經死亡,其體外寄 生蟲可能在魚網中受到擠壓而脫落,因此在本研究中並無發現盲鰻感染單殖吸蟲。
橈足類寄生蟲 Archidactylina myxinicola 寄生於紫盲鰻及紫黏盲鰻的鰓囊 (Izawa, 1996),紫黏盲鰻分佈深度為 300-1000 m (Kuo et al., 1994),大於一般底拖網作業深 度,因此本研究並無採集到紫黏盲鰻;紫盲鰻分佈於日本,其分佈深度約 500-800 m (Jorgensen et al., 1998),推測此種橈足類以分佈深度較深的盲鰻為寄主,因此在 本研究中的四種盲鰻皆無發現。
海獸胃線蟲在臺灣海域已有許多寄生於硬骨魚及鯨豚的記錄,包含了花腹鯖、
白帶魚、真鰺、烏魚、大眼鯛等魚類,以及瓶鼻海豚、熱帶斑海豚、侏儒抹香鯨 等等 (吳,1996;周,2005;林,2006;查,2004;陳,2007;葉,2013)。其生 活史也已有許多研究,包括透過掠食者捕食獵物的方式傳遞,以橈足類與磷蝦等
29
甲殼類,以及多種硬骨魚類和頭足類為保蟲寄主,海洋哺乳類為其最終寄主 (Klimpel et al., 2004)。此外,當寄主食入含有海獸胃線蟲的保蟲寄主而遭受感染,
且海獸胃線蟲無法經該寄主完成其生活史,最終在寄主體內死亡,則稱意外寄主 (accidental host),如人類、海龜 (Santoro et al., 2010)。
歐洲的七鰓鰻是在寄生於鮭魚時,推測經由寄生的傷口感染了海獸胃線蟲,
也是第一次發現海獸胃線蟲寄生於無頷魚類 (Bao et al., 2013)。盲鰻則是經由腐食 行為食入海獸胃線蟲,實驗也在盲鰻消化道中發現魚鱗、蝦殼、未消化的頭足類。
將海獸胃線蟲置於生理食鹽水中並保存於4℃中,經過 21 天後,仍有 75%的線蟲 存活,顯示海獸胃線蟲在寄主死亡,或是離開寄主後仍可以存活一段時間 (Oh et al.
2014),因此盲鰻可經由攝食這些帶有線蟲的動物屍體而感染,同時在海獸胃線蟲 生活史中增加一條盲鰻經腐食其他保蟲寄主而感染的途徑 (圖 26),有別於以往利 用寄主掠食方式傳播。雖然目前無直接證據證實海獸胃線蟲是否能經由盲鰻完成 其生活史,但盲鰻為多種硬骨魚及鯨豚類的獵物 (Jorgensen et al., 1998; Zintzen et al., 2011),推測盲鰻可作為海獸胃線蟲之保蟲寄主,同時研究中的四種盲鰻也為海 獸胃線蟲的寄主新記錄。
刺魚蚤屬 (Genus Acanthochondria) 目前確定有 52 個物種,主要寄生於深海或 底棲硬骨魚類的口腔、鰓腔、鼻腔及泄殖腔,如牛尾魚科、鮟鱇科、鰈科等 (Poly and Mah, 2001; Ho et al., 2011),這也是第一次發現刺魚蚤屬寄生於無頷魚類的盲鰻。
盲鰻呼吸器官為鰓囊,不同於硬骨魚的鰓,也沒有鰓蓋及鰓腔;水經由鼻孔流入 咽部,再經過鰓囊進行氣體交換,最後從鰓孔流出。盲鰻刺魚蚤皆寄生於咽部,
流過的水流可供盲鰻刺魚蚤進行物質的交換,且推測盲鰻刺魚蚤的卵在孵化成無 節幼體後,可經由鰓孔排出到海水中,接著發育成有游泳能力的橈足幼體,再寄 生於其他盲鰻寄主。而盲鰻刺魚蚤對寄主專ㄧ性同樣無法定論,軟刺魚蚤科之寄 生蟲,其自由生活時期的橈足幼蟲被認為可以在不同科的底棲魚類寄主之間進行
30
轉移 (Paterson and Poulin, 1999)。本研究只針對臺灣東北海域盲鰻,四種盲鰻皆為 黏盲鰻屬 (Genus Eptatretus),盲鰻刺魚蚤是否會寄生在其他盲鰻屬的盲鰻還需要 更多研究。
本研究共捕獲四種盲鰻皆感染盲鰻刺魚蚤,為盲鰻之寄主新紀錄和臺灣海域 之地理新記錄。並於2014 年正式發表為新種 (Cheng et al., 2014),文章記錄蒲氏 黏盲鰻、沈氏黏盲鰻、楊氏黏盲鰻,三種盲鰻感染盲鰻刺魚蚤。後續研究也發現 臺灣黏盲鰻檢出盲鰻刺魚蚤感染,但臺灣黏盲鰻樣本數少,且只有一尾受到感染,
因此需要更多樣本來確定盲鰻刺魚蚤是否以臺灣黏盲鰻為寄主。
4.2 海獸胃線蟲物種
線蟲 DNA 樣本經過 PCR-RFLP 分析後共得到 Anisakis 屬線蟲六種,為 A.
pegreffii、Anisakis sp.、A. simplex s.s.、A. typica、A. brevispiculata、A. pegreffii 與 A. simplex s.s.的重組基因型 (recombinant genotype),及 Hysterothylacium 屬ㄧ種 H.
amoyense。盲鳗為上述寄生蟲之寄主新記錄,其中 Anisakis sp.及 H. amoyense 為臺 灣海域之地理新記錄。
A. pegreffii 是臺灣多種硬骨魚中最主要的寄生性線蟲物種,在四種盲鰻體內也 有寄生,且為蒲氏、楊氏、臺灣黏盲鰻主要的寄生物種。A. pegreffii 主要分佈於地 中海、東北大西洋、南大西洋及西北太平洋,其中包含臺灣、日本海及南韓海域,
以海豚科 (Delphinidae) 為最終寄主 (Kuhn et al., 2011),臺灣周遭海域也有瓶鼻海 豚 (Tursiops truncatus)、熱帶斑海豚 (Stenella attenuata) 等多種海豚分佈。
Anisakis sp.為本研究中數量第二多的物種,主要寄生於沈氏黏盲鰻,少部分寄 生於楊氏黏盲鰻。過去在南非海域的黑胸燧鯛 (Hoplostethus cadenati)(Kijewska et al., 2009) 及葡萄牙海域的黑斑小鯛 (Pagellus bogaraveo)(Hermida et al., 2012) 也
31
有該種線蟲寄生的紀錄;在最終寄主上,菲律賓群島擱淺的侏儒抹香鯨體內也發 現此種線蟲寄生 (Quiazon et al., 2013)。黑胸燧鯛與黑斑小鯛同樣為底棲性魚類,
分佈深度分別為200-700 m 與 150-300 m (Froese and Pauly, 2014),與沈氏黏盲鰻的 的分佈深度200-800 m 相似 (Jorgensen et al., 1998),而這兩種魚類皆以甲殼類為主 食,而侏儒抹香鯨則以深海頭足類為食。另外,Anisakis sp.與 A. paggiae 及 A.
ziphidarum 親緣關係最接近 (Mattiucci and Nascetti, 2006; Quiazon et al., 2013)。A.
paggiae 最終寄主為侏儒抹香鯨 (Kogia sp.) , A. ziphidarum 最終寄主為喙鯨 (Mesoplodon sp.),兩種鯨類皆於深海覓食,推測此種海獸胃線蟲可能以深海甲殼 類、魚類及頭足類為保蟲寄主,而盲鰻經由腐食行為吃入這些帶有線蟲的保蟲寄 主而感染。
A. brevispiculata 寄生於蒲氏黏盲鰻,於劍旗魚有寄生記錄,最終寄主為侏儒 抹香鯨 (Mattiucci et al., 2007; Quiazon et al., 2013),其他寄主的紀錄也相當稀少,
此兩種動物皆以魷魚等頭足類為主食,因此魷魚可能為 A. brevispiculata 生活史中 重要的寄主,臺灣周遭海域皆盛產鎖管、花枝等頭足類,雖然沒有寄生於臺灣頭 足類之紀錄,但在臺灣東北部捕獲之花腹鯖有寄生紀錄 (Chen and Shih, 2015),此 次發現盲鰻受 A. brevispiculata 寄生,可能因食入帶有此線蟲的鯖魚或是頭足類的 屍體而受到感染。
A. simplex s.s. 目前已知分佈限於北半球北緯 20 度以北海域,包含北大西洋、
東北太平洋及西北太平洋的日本東部海域,較少於熱帶及亞熱帶海域發現,寄生 於多種海魚、頭足類及海洋哺乳類 (Mattiucci et al., 2007; Kuhn et al., 2011)。A.
simplex s.s. 於日本東部海域的日本鯖 (Scomber japonicus) 體內極為常見 (Suzuki and Murata, 2011),臺灣東北部則是以花腹鯖且以此 A. pegreffii 寄生為主 (周,2005),
此次研究於沈氏黏盲鰻中發現 A. simplex s.s.,雖然檢獲數量僅有一隻,但仍然能 顯示盲鳗之海獸胃線蟲項可能因其腐食性及底棲性而與大洋性魚類有所差異。
32
A. simplex s.s.與 A. pegreffii 同樣以海豚科為最終寄主,A. simplex s.s.主要分佈 於溫帶及寒帶海域,A. pegreffii 則是分佈於熱帶、亞熱帶海域 (Kuhn et al., 2013),
然而在這兩種分佈重疊處,常常可以發現此二物種的重組基因型,如地中海與大 西洋交接海域,以及日本與臺灣中間海域皆為此二物種分佈重疊的地區,在過去 也都有文獻證實此重組基因型的存在 (Abollo et al., 2003; Mattiucci et al., 2007)。臺 灣過去也同樣於真鰺、花腹鯖體內發現 (林,2006;周,2005),此次研究也同樣 發現重組基因型存在於蒲氏黏盲鰻。
A. typica 主要分佈於熱帶及亞熱帶海域,與其最終寄主分佈相關,如瓶鼻海豚 及熱帶斑海豚,分佈於熱帶至溫帶海域,也包含了臺灣週遭海域,如臺灣西南海 域的花腹鯖,以及東北海域真鰺都有寄生紀錄 (Kuhn et al., 2011;Chen and Shih, 2015; 周,2005;林,2006),這次於蒲氏黏盲鰻也同樣有 A. typica 寄生。
以卡方檢定分析 A. pegreffii 在四種盲鰻感染線蟲物種百分比中發現,沈氏黏 盲鰻體內 A. pegreffii 百分比顯著低於其他三種,而蒲氏、楊氏、臺灣三種黏盲鰻 間無顯著差異。不同物種的海獸胃線蟲可能因寄主的攝食行為、棲地而有不同的 深度分佈;如地中海底棲性的歐洲狗鱈,其海獸胃線蟲物種數多於同一地區的竹 莢魚,顯示底棲性的歐洲狗鱈有較豐富的食性;A. physeteris 主要寄生於魷魚,劍 旗魚 (Xiphias gladius) 感染高比率的 A. physeteris,推測劍旗魚以魷魚為主要食物 之一 (Mattiucci et al., 2007)。盲鰻以動物屍體為食,包含各種甲殼類、頭足類、魚 類、海洋哺乳類,對於物種的選擇性較低,相較於大洋性魚類,更能接收來自不 同寄主體內的海獸胃線蟲,也更能顯示出海獸胃線蟲在海洋中的分佈。蒲氏黏盲 鰻受5 種線蟲感染,其較廣的棲地分布 (日本、台灣、東南亞) 及較高的盛行率與 感染強度,使得蒲氏黏盲鰻有機會食入較多的線蟲物種。
蒲氏、楊氏、臺灣三種黏盲鰻皆分佈於深度較淺的海底,分別為10-270 m、
20-261 m、20-427 m (Mok and Chen, 2001; Jorgensen et al., 1998),在分佈深度上有