國立臺灣大學醫學院暨工學院醫學工程學研究所 碩士論文
Graduate Institute of Biomedical Engineering College of Medicine and College of Engineering
National Taiwan University Master Thesis
利用電場調變整合素蛋白極化及引導細胞移動現象 Electric field modulation of integrin polarization and
directed cell migration
蔡承憲
Cheng-Hsien Tsai
指導教授:趙本秀 博士
Advisor: Pen-Hsiu Grace Chao, Ph.D.
中華民國 100 年 8 月
August, 2011
誌謝
兩年的學習與實驗終於到了一個段落,在這其中充斥著無數的感激,首先得 感謝趙本秀老師平時的教導,在不斷的討論、規劃與不厭其煩的指導中得到了論 文的方向,從無到有的做完這篇論文。感謝口試委員林峰輝教授與郭柏齡教授,
在百忙之中抽空參加我的口試,並且針對內容提出建議,使得這篇論文更加完備。
另外要更加感謝林峰輝教授提供了共軛焦顯微鏡實驗儀器,這大大的幫助我進行 論文中的螢光染色顯像的部分。
要特別感謝曾經在我實驗中投注心力的仕帆、晏彰、均源,嘉襄、祐甄、沿 毓、新民、守謙、向儀、維仁,在實驗的進行上給予協助也讓我順利而且歡樂的 完成研究所的學業。
最後,感謝永遠支持我的家人與親友們,感謝一路上陪伴自己走過的人,有 大家的支持,我才能順利的完成我的求學歷程
中文摘要
電刺激被臨床使用在疼痛的治療以及促進傷口癒合。在骨科的處理上,施予 電場可以促進骨頭的癒合以及促進韌帶的復原。在實驗中,我們選用物理治療上 時常使用的電刺激波形並檢視對於前十字韌帶纖維母細胞移動以及型態的影響,
我們選用的大部分刺激參數都可以增加纖維母細胞的移動,然而對於移動方向上 有著十分顯著的差異。此外,前十字韌帶纖維母細胞只有在直流電刺激下會造成 細胞有延伸排列的情形。實驗的結果使我們發現細胞移動速度與方向性上有不耦 合的情形,這可能是因為不同的機制在進行調控。我們發現整合素蛋白在電場誘 發細胞方向性改變的傳導機轉上扮演重要的控制角色,另外也發現整合素蛋白的 重新排列也會帶動 RhoA 排列在負極的現象,這表示電場藉由整合素蛋白的不平 均分佈調控了下游重要的傳導分子的分布狀態。本篇研究有助於了解物理治療臨 床電療在細胞層面的行為影響,以及建立與釐清電場誘導細胞方向性改變的傳導
機轉與整合素蛋白的關係。
關鍵字: 電場,移動,整合素蛋白,RhoA
ABSTRACT
Electrical stimulation is clinically used for the treatment of pain and to promote wound healing. In orthopaedic practices, applied electric fields (EFs) promote bone healing and improve lapine ligament repair in vivo. In the current study, several stimulation waveforms used in physical therapy were adapted to examine their effects on anterior cruciate ligament fibroblast (ACLF) migration and morphology. Most of the waveforms we tested resulted in enhanced fibroblast migration, while their effects on migration directionality were noticeably different. Furthermore, ACLFs elongation and alignment were only found in the DC groups. These findings suggest a
decoupling of migration speed and directionality, which may arise from disparate mechanisms. We found that integrin acts as a major player of EF-induced
directionality. We also found that integrin redistribution mediate the cathodal redistribution of RhoA. This introduces EF mediates one of the major signaling molecules, which is downstream from the integrin asymmetrically, with stronger redistribution on the cathode, is highly significant functionally. Results from this study may benefit our understanding the electro-therapy treatment on cell behavior and the relation between integrin and EF-induced directionality.
Key Words: Electric field, Migration, Integrin, RhoA
CONTENTS
國立臺灣大學碩士學位論文口試委員會審定書 ... i
誌謝 ... ii
中文摘要 ... iii
ABSTRACT ... iv
List of figures ... vi
Chapter 1 Introduction ... 1
Integrin: important role for mediating cell migration ... 2
EF-induced integrin redistribution ... 3
Members of the Rho Family of GTPases are regulated by Integrin-mediated Signals ... 4
Chapter 2 Materials and methods ... 7
Cell culture ... 7
Electric field ... 7
Pharmacological treatment ... 8
FRAP measurements ... 9
Migration analysis ... 10
Immunohistochemistry ... 10
Statistical analysis ... 11
Chapter 3 Results ... 12
EF-induced migration ... 12
EF- induced cell surface marker redistribution ... 13
Integrin distribution mediate EF-induced directionality ... 13
EF- induced RhoA redistribution ... 15
RhoA activity mediate EF-induced directionality ... 15
Chapter 4 Discussion ... 16
Reference ... 34
List of figures
Fig 1. The composition of a galvanotaxis chamber Fig 2. Electric waveforms
Fig 3. Electric waveform-induced fibroblast migration Fig 4. Morphology of fibroblast in applied EF
Fig 5. EF-induced integrin redistribution
Fig 6. Effect of cholesterol treatment on FRAP in membrane.
Fig 7. Pharmacological effect on EF-induced integrin redistribution and cell migration Fig 8. Short term EF- induced integrin redistribution
Fig 9. EF-induced RhoA redistribution
Fig 10. Pharmacological effect on EF-induced RhoA redistribution
Fig 11. The effect on EF-induced cell migration with inhibition of RhoA and PKA activation
Chapter 1 Introduction
Numerous approaches have been proposed to enhance the healing capability of the anterior cruciate ligament (ACL). Electrical stimulation is widely used for the treatment of pain and to promote wound healing [1]. In orthopaedic practices, applied electric fields (EFs) have been used clinically to promote bone healing [2]. EF has also been shown to improve lapine ligament repair in vivo [3]. We have previously
demonstrated that applied EF enhanced ligament fibroblast migration and collagen production, depending on the applied EF parameters[4]. Electric stimulation of various types has been found to be an effective treatment technique for soft and hard tissue healing. Besides the long known, low-frequency currents (galvanization, impulse galvanization, diadynamic currents, TENS, faradic current, HVPGS) are increasingly applied today, as they cause patients less discomfort. Low intensity direct current can accelerate the healing rate of both hard and soft tissue injuries [5]. Electrical
stimulation of osteogenesis involves the use of faradic waveform to promote the growth of new bone [6]. Faradic stimulation maximizes the restoration of elastic properties of the muscle by encouraging the correct parallel configurational
arrangement of the collagen fibres within the cellular matrix [7]. Diadynamic current is a low frequency and biphasic sinusoidal-like waveform. The phase duration (10~20
ms) is longer than faradic current. Clinical usage of diadynamic current (fluctuation current) is for pain [8], analgesia [9], electro-myo stimulation [10], throboembolism (circulating) [11], and suppurating wounds healing [12]. In particular, several recent reports that high-voltage, pulsed galvanic stimulation (HVPGS) significantly
increases proliferation, collagen synthesis and healing rate of skin wounds. Similarly, in animal models HVPGS have shown an accelerated rate of full-thickness burns healing [13, 14]. In the current study, several stimulation waveforms used in physical therapy for promoting tissue repair were adapted to examine their effects on ACL fibroblast migration. Here, we focused on clinical electro-therapy on cell migration behavior.
Integrin: important role for mediating cell migration
Cell adhesion underlies several important physiological processes, such as cell migration, spreading, polarity, anchoring, differentiation, morphogenesis and wound healing. Focal adhesions are provided by adhesion molecules and expressed at the cell surface of all nucleated cells, and they mediate extracellular binding to cell and tissue substrates and transmit mechanical docking to the intracellular actomyosin or
intermediate filament cytoskeleton.
During cell migration, many classes of membrane receptors fulfill adhesion and
cytoskeletal coupling functions and provide a range of adhesion strength, specificity and turn-over rates. High affinity adhesion to ECM ligands is predominantly provided by the integrin family [15].The integrin-mediated cell substrate interactions and
linkages to the actin cytoskeleton, which is mediating cell movement, and its form and turn-over dynamics and polarity determine cell speed and directionality.
EF-induced integrin redistribution
The electrocoupling mechanisms are confined to the cell surface. Because cell membrane receptors such as integrins have been demonstrated to laterally diffuse [16]
and redistributed by ES, the mechanism for altered receptor distribution patterns (e.g., clustering) could be offered as at least one of the electrocoupling pathways. In recent
years, laboratories use several ways to change the distribution of integrin to find out the linkage between integrin redistribution and cell migration. The DC electrical stimulus (0.1V/cm) facilitates the cellular focal adhesion formation of human fibroblast in three-dimensional collagen gel by clustering of integrins on the membrane [17]. Cell migration may be extended to postulate that application of ES triggers the initial event of integrin clustering [18, 19] that serves as the site for formation of a focal adhesion by recruiting other molecules such as actin-binding proteins and signaling molecules, and integrin ligation mediating its activation and/or
the clustering of adhesion components to initiate a nascent adhesion [20, 21].
Members of the Rho Family of GTPases are regulated by
Integrin-mediated Signals
Integrin engagement and subsequent clustering of these receptors in focal adhesions lead to the generation of intracellular macromolecular complexes[22]. Numerous enzymatic activity (GTPases) signaling proteins are also structural components of focal adhesions. The Rho family of GTPases contributes to these integrin-mediated signals; in particular, signals that control cytoskeletal organization involved in changes in cell motility. RhoA are part of the Ras superfamily of proteins that cycle between an active, GTP-bound state and an inactive, GDP-bound state.
Activated RhoA is capable of stimulating microfilament bundling in serum-starved cells that are already adherent [23]. Rho is also essential for the formation of focal complexes [24]
Studies devoted to how Ras superfamily protein induces integrin clustering into FAs, and much less are known about the clustering of integrins into focal complexes influences RhoA, Rac and Cdc42. Many studies have also demonstrated that
shear-induced RhoA activation is secondary to integrin activation [25] . With
increasing numbers of GEFs being identified [26]Rho can be activated by integrin ligation [27, 28], leading to the formation of stress fibers and FAs. Many studies have also indicated that RhoA is activatedupon integrin-mediated adhesion [29, 30].
Development in the field has been the recognition that integrin-mediated adhesion stimulates activation of RhoA. GTP-bound RhoA measurement reveals that integrin-mediated adhesion causes modest increase in RhoGTP levels [27].
Integrins not only control the activation of Rho family but also separately regulate the translocation of activated RhoA, Rac1 and Cdc42 to the plasma membrane [31]. Integrin signals regulate the location of membrane domains such as lipid rafts and thereby control domain-specific signaling events in
anchorage-dependent cells. RhoA is also thought to be concentrated in lipid rafts [32], and GTP-Rac1 binds more effectively to membranes from adherent than from suspended fibroblasts, indicating that integrins regulate RhoA and Rac1 membrane binding sites at the cell surface [31]
We anticipate that adhesion-mediated regulation of Rho family GTPases will play an important role in the complex process of cell migration behavior
In this experiment, we look further into the hypothesis that (1) different
waveform (direct current, diadynamic current, faradic current, sinusoidal current, and high voltage pulsed galvanic stimulation) caused different integrin distribution and
induced different fibroblast cell behavior (cell migration, orientation) (2) Electrical stimulation waveforms affects integrin redistribution which mediates EF-induced directionality. (3) integrin clustering and redistribution serves a mediator to activate and redistribute RhoA then induces cell migration directionality.
Chapter 2 Materials and methods
Cell culture
Anterior cruciate ligament (ACL) was harvested from porcine knee joint and cut into cubes. The explants were cultured in DMEM (Biowest) supplemented with 10% FBS (Sigma), penicillin and streptomycin, until the fibroblasts had grown out to confluence. For electric field studies, cells were released from high density monolayer cultures using a trypsin/EDTA solution and plated at 1.8×104cells / cm2 on to sterile glass slides. After attachment for 2 hrs in a 5 percent CO2 incubator at 37°C, the slide was gently rinsed with medium and then positioned in a custom galvanotaxis chamber, which was assembled under a sterile laminar flow hood.
Electric field
A customized electrical stimulation chamber was used as previously described [4]
(Figure 1). Constant direct current (DC) EF was applied at either 30 V or 0.19 V across the chamber using a Keithley SourceMeter. Other waveforms that were applied
using a custom stimulator (Dynaprog, MingQuo, Taiwan) as illustrated in Figure 2.
All waveforms and the constant 0.19 V group except for HVPGS were controlled to have the same amount of total current flow (1.9×10-5As). Faradic current (Far) represented a graduation series of triangular waves peaking at 15 V at 50 Hz.
High-voltage pulsed galvanic stimulation (HVPGS) consists of monophasic,
twin-spike pulses that have a fixed pulse duration of 100 μs and max intensity at 100 V with a frequency of 2.5 Hz. Sinusoidal waves (sin) have a peak intensity of 1.2 V at 50 Hz and the diadynamic waves (diadyn) are rectified monophasicc sinusoidal waves
Pharmacological treatment
Cholesterol treatment
Cells were preincubated for 30 min with 1 mM ChH (cholesteryl hemisuccinate) (Cat # C6512, Sigma) dissolved in dimethylsulfoxide and diluted in PBS then washed
[33].
Antibody treatment
Cells were preincubated for 30 min with 2.8 μg/ml anti–porcine integrin α2β1 antibody (Cat # mab1998, 1 mg/ml) diluted in PBS and washed.
C3 transferase
Cells were preincubated for 2hrs with 2 μg/ml C3 transferase (Cat # CT04, cytoskeleton) diluted in serum free medium and washed.
PKA inhibitor
Cells were preincubated for 1hr with 10μmol/liter KT 5720 (Cat # k3761, sigma) dissolved in methanol and diluted in PBS then washed
FRAP measurements
FRAP studies were conducted on live cells labeled with DiI (2.5 μg/ml, sigma).
The cells were taken for FRAP experiments after 30 min cholesterol treatment.
Measurements were taken in PBS at room temperature. A Leica SP2 confocal microscope was used with a 60X objective for examining the stained cells. After a brief measurement at monitoring intensity, laser pulse (~3s) was used to bleach 50–75% of the fluorescence in the spot. In Fig. 6, the T1/2 values were extracted from the averaged FRAP curves, fitting to the analytical expression for a combination of diffusion coefficient D [34, 35] .
Migration analysis
Cell migration was analyzed by manually selecting centroid of each cell and tracking them with time. Cell migration distance and angle (θ) were measured and directional velocity, a representation of cell migration in the direction of the field, was calculated by multiplying migration speed with cos (θ).
Immunohistochemistry
Anti– integrin antibody (α2β1) was obtained from Chemicon (mab1998, 1 mg/ml) and RhoA antibody (ARH03, Cytoskeleton). Cells were fixed for 30 min with 4%
paraformaldehyde in PBS, and blocked with serum. Cells were incubated overnight in 40C with the respective monoclonal antibody diluted in PBS (1:200) supplemented with 1% serum. FITC-conjugated secondary antibodies (AP124F, 2 mg/ml, Chemicon) diluted in 1X PBS (1:200) supplemented with 1% BSA for 1hr was used for labeling the cell surface integrin. An Olympus microscope with a FluoView confocal system was used with a 20X objective for examining and photographing the stained cells. For quantifying the distribution of integrin, we divided the cell into four equal parts and
used image analyzing software (Image J) to measure the fluorescence intensity in the quarter then normalized to the overall cell average. After we measure the normalized intensity into the four regions, the asymetric index (AI) is calculated by subtracting the normalized intensity of the region facing cathode from quarter facing anode.
Statistical analysis
STATISTICA (Stat Soft) was used to perform ANOVA and post-hoc tests (LSD) with α=0.05. All bars represent 2-3 separate experiments.
Chapter 3 Results
EF-induced migration
To characterize the disparate waveform-induced cell migration behavior, we applied EFs used in physical therapy to ACLFs plated on glass.
Without an electric field, most fibroblasts migrate in random direction. After 1 hr exposure to DC EF, both 30 V and 0.19V EF caused most cells to elongate
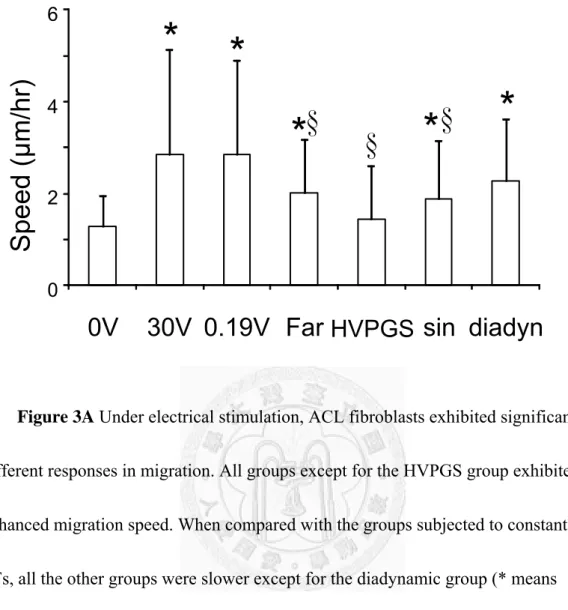
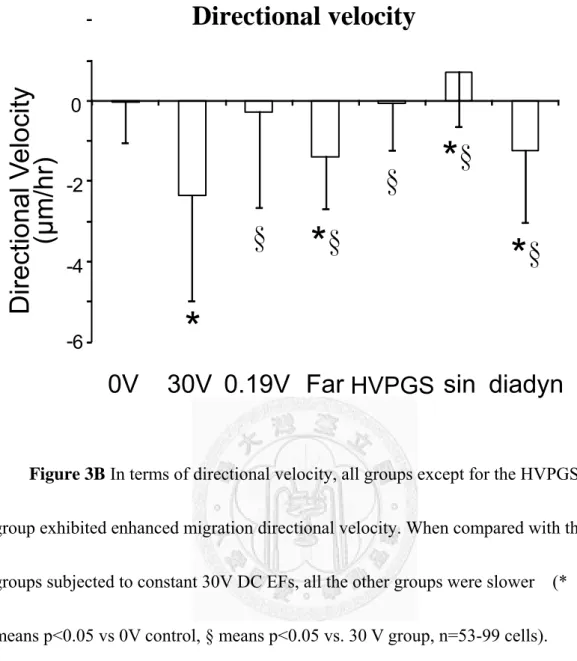
perpendicularly to the applied field and migrate toward the cathode. Other groups except for the HVPGS group exhibited enhanced migration speed but did not exhibit any significant elongation or orientation (Figure 4). When compared with the groups subjected to constant DC EFs, all groups were slower except for the diadynamic group (Figure 3A). In terms of directional velocity, which represents the speed toward the cathode, all groups except for the HVPGS group exhibited enhanced migration directionality. When compared with the groups subjected to constant 30V DC EFs, all other groups were slower. Interestingly, no directionality was found with the constant 0.19 V EF group even though it has similarly enhanced migration speed with the 30 V group (Figure 3B).
EF- induced cell surface marker redistribution
To investigate the effects of PT waveforms on integrin redistribution, we measured the fluorescence intensity of the labeled-integrin. Faradic, diadynamic and 30V DC EF redistributed integrin toward cathode, and this observations on
EF-induced cell migration seem consistent withstudies have also shown that DC electric field can redistribute integrin toward the cathode [17] ,but exposure to HVPGS, 0.19V DC EF, integrin distribute randomly. Other than EF- induced integrin cathodal distribution, sine wave induce integrin move toward anode (Figure 5a). When
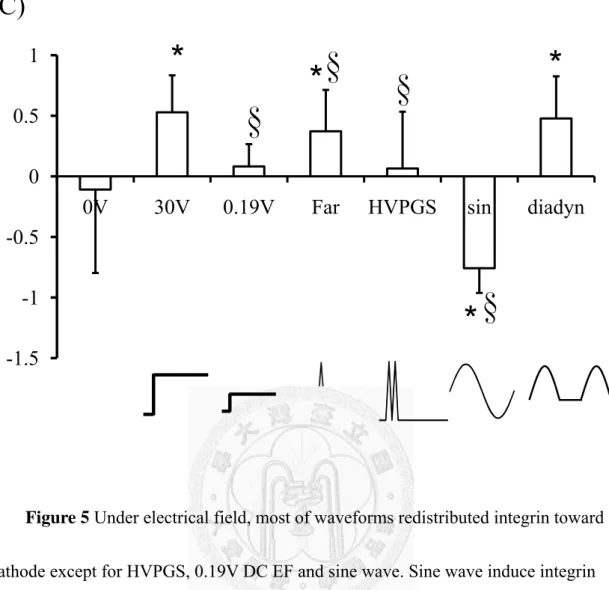
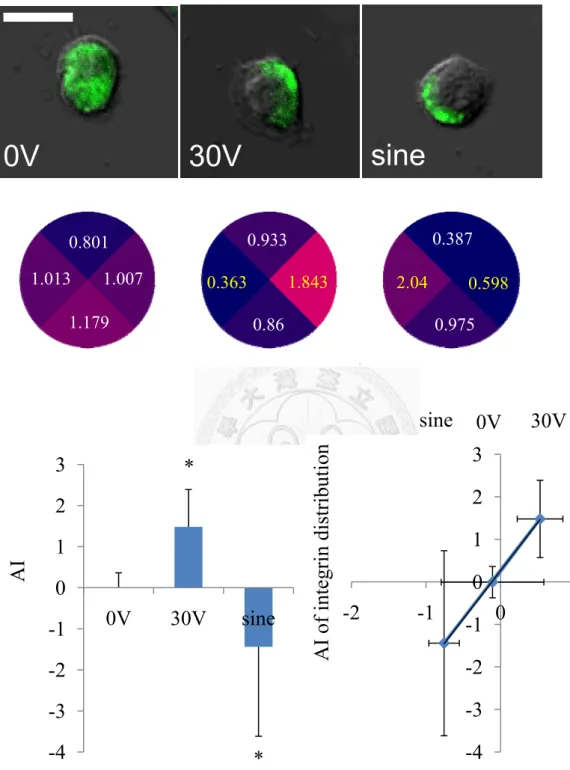
calculating the normalized fluorescence intensity in the quarter, Faradic, diadynamic and 30V DC EF parameters affect integrin staining stronger in the cathodal quarter than anode quarter but HVPGS causes integrin distribute randomly and sine wave affects integrin move toward opposite side (Figure 5b). After analyzing the asymetric index, we found out the asymmetric value between groups are related to EF-induced directionality (Figure 5c).
Integrin distribution mediate EF-induced directionality
To characterize the effects of the integrin distribution on cell migration behavior,
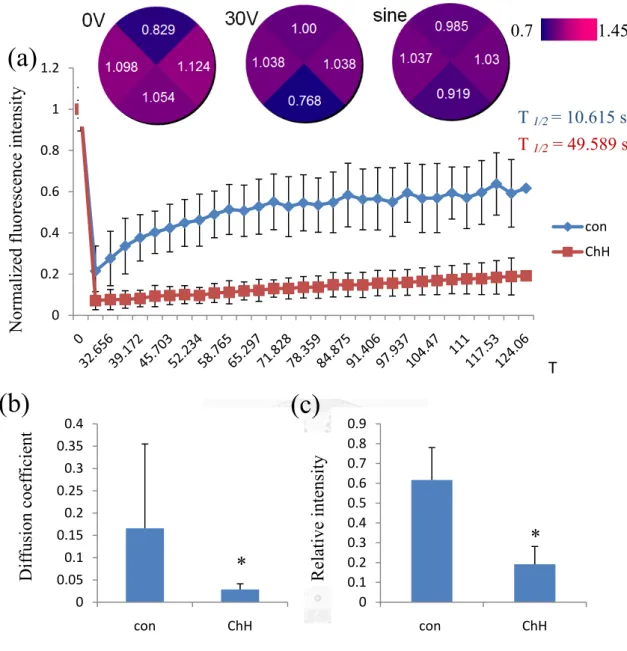
we investigated cell migration speed and directional velocity for changes induced by cholesterol hemisuccinate and integrin monoclonal antibody. To reduce membrane fluidity, we used cholesterol hemissuciate and analyzed membrane diffusion coefficient quantitatively and indicated that the rate of diffusion and fluorescence recovery intensity loss by cholesterol embedding (Figure. 6b, 6c). Our observations on membrane fluidity reduced by cholesterol (Figure. 6a) seem consistent with study
[33] found cholesterol induced random distribution of membrane receptor, manifested by decreasing membrane fluidity and diffusion rate. Moreover, we found that
randomly distributed integrin in electric field by cholesterol (Figure 7a) also
decreased the EF-induced migration directionality (Figure 7c) but did not affect speed (Figure 7b). However, the ChH treatment is the non-specific way to reduce the
integrin redistribution, we further blocked integrin function specifically by anti-integrin antibody, and the function loss of integrin also had same effect on directionality and speed (Figure 7b, 7c) by cholesterol even though it has asymmetric distribution (Figure 7a). Based on EF-induced cell migration would be an early and proximal event after EF onset, we observed the short-term EF exposure to integrin redistribution. We found out not only 1 hr DC EF exposure can affect integrin redistribution, in short-term EF expose; both 5 min and 10 min period also affected integrin redistribute toward cathode (Figure 8) then mediated the cell migration
directionality.
EF- induced RhoA redistribution
We also investigated RhoA distribution tried to find out the correlation with integrin distribution. For 1hr DC EF, the RhoA redistributed toward cathode. Other than EF- induced RhoA cathodal distribution, sine wave induced RhoA move toward anode (Figure 9). The trend of EF-induced RhoA redistribution coincides with
integrin (Figure 9). Although exposure to 30V DC EF, the RhoA redistributed toward cathode, but anti-integrin antibody treatment blocked RhoA redistribution
phenomenon and affected RhoA distribute randomly (Figure 10).
RhoA staining revealed a lack of RhoA redistribution with functional blocking of integrins, suggesting inhibited intracellular polarization, but inhibition of PKA
activity seems does not affect EF-induced RhoA redistribution.
RhoA activity mediate EF-induced directionality
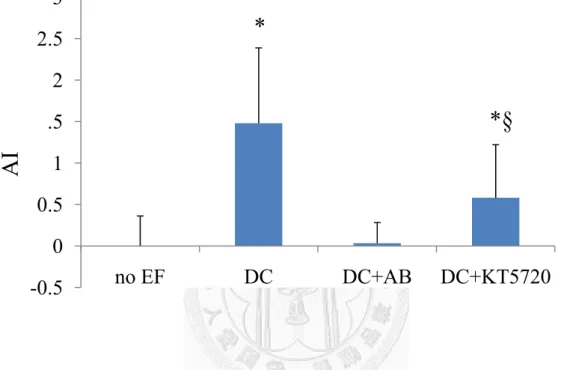
To characterize the effects of the RhoA activity on cell migration behavior, we inhibited RhoA activation by C3 transferanse, and found out similar effect on cell migration in applied EF with integrin blocking and PKA inhibition (Figure 11).
Chapter 4 Discussion
Electric stimulation in physical therapy has been used for many years in repairing non-healing soft and hard tissue. DC mode ES can redistribute cell membrane receptors such as integrin via electrophoresis and electroosmosis mechanism[16] even in the 3-D environment [17]. The EF-induced integrin
redistribution serves as the site for formation of a focal adhesion by recruiting other molecules such as actin-binding proteins and signaling molecules[19] and influences the cell migration directionality.
Based on these proposed mechanisms, we first hypothesize that different
waveform (direct current, diadynamic current, faradic current, sinusoidal current, and high voltage pulsed galvanic stimulation) caused different integrin distribution and induced different fibroblast cell behavior (cell migration, orientation). Therefore, we investigated the effects of physical therapy electrical stimulation on ligament cell migration and morphology. Most of the waveforms we tested resulted in enhanced fibroblast migration speed, while their effects on cell migration directionality were noticeably different. As we have previously shown, bovine ACL fibroblast and chondrocytes both exhibit different EF threshold response in motility and
directionality[4, 5]. Furthermore, ACL fibroblast exhibit frequency dependent
migration behaviors [4]. Using different waveform and field strengths, we discovered a decoupling of cell shape and directionality, which may suggest disparate mechanisms in the two responses [36]. Faradic ad diadynamic stimulation promoted the most significant response in both speed and directionality. Faradic stimulation has been shown to promote collagen organization while diadynamic stimulation promotes skin wound healing [7, 12]. Interestingly, while HVPGS has been demonstrated to increase proliferation, collagen synthesis and skin wound healing, no significant effect of increased cell migration is shown here [37].
In terms of cell migration behavior under different field strengths DC EF, interestingly, no directionality was found with the constant 0.19 V EF group even though it has similarly enhanced migration speed with the 30 V group. In here, we hypothesize disparate mechanism control migrate speed and directionality. Zhao et al.
[38] demonstrated using pharmacological inhibitors to find that directionality and migration rate are regulated by separable pathways. To determine the effects of the electric waveforms on cell migration behavior and the major factors control migration mechanism, we investigated EF-induced integrin redistribution.
When analyzing the asymetric index, we found out the asymmetric value between groups are related to EF-induced directionality. We further look into the
cause and effect between integrin redistribution and EF-induced directionality to clarify the signaling mechanism.
We investigated the time dependent redistribution effectiveness firstly.
Asymmetric distribution of integrin receptors labeled with specific antibodies was evident as early as 5 min after EF onset. Integrin redistribution occurs within minutes, which is the early and proximal event like EF-induced cell directionality. Furthermore, we decreased cell membrane fluidity by cholesterol hemisuccinate to reduce
EF-induced integrin redistribution. Membrane lipid fluidity depends mainly on the cholesterol content as well as on the degree of saturation of phospholipid acyl chains and phospholipid polar head group composition [39, 40] . Therefore we modify the physical state of membrane lipid matrix (i.e., the membrane lipid fluidity or microviscosity) by alteration of the cholesterol content. The fluorescence recovery after photobleaching (FRAP) experiment shows ChH-treated cell membrane diffusion coefficient is lower than the normal cell membrane. Moreover the ChH-treated
membrane eliminated polarization of integrin in the EF, and decreased EF-induced directionality, but did not affect the migration speed. Since the ChH treatment is the non-specific way to reduce the integrin redistribution, we further blocked integrin function specifically by anti-integrin antibody. The Ab-treated cell also decreased EF-induced directionality but did not affect speed; its effect on migration behavior is
same as ChH-treated cell. When using confocal microscopy to measure EF-induced integrin distribution, we found that integrin also redistributed toward cathode. Even though applied EF polarized the antibody-bound integrin, the functional blocking treatment abolished the directional response. Taken together, our results indicate that activation and polarization of α2β1 integrin mediates EF-induced directionality.
EF not only directs cathodal redistribution of integrin but up-regulates the integrin receptor [30]. Numerous steps have been taken toward understanding the Rho GTPases during cell polarization and directional migration, and Rho GTPases have emerged as key regulators in cell migration directionality [43].This introduces EF activates one of the major signaling molecules, which is downstream factor of mediating migration from the integrin asymmetrically.
Our current data show that this occurs in parallel with a cathodal redistribution of RhoA. We report on the involvement of α2β1 integrin mediated RhoA polarization in EF-induced migration. α2β1 integrin specifically binds to the main ligament matrix component, type I collagen, and α2 integrin has been shown to modulate ligament fibroblast migration. By functionally blocking α2β1 integrin with its antibody, we inhibited directionality without changing motility. This response coincides with the lack of RhoA polarization. Inhibition of RhoA also abolishes EF-induced
directionality.
The mechanism underlying the key role between RhoA and integrin
redistribution is not clearly understood. One likely possibility role on redistributing and activating RhoA is PKA. RhoA is a PKA substrate [41] and PKA activity at the leading edge exhibited a close temporal and spatial correlation with the formation of
activation of RhoA and cell protrusions [42]. We report on the inhibition of PKA activity by inhibitor also diminished EF-induced directionality, but EF-induced RhoA polarization still existed. Even though applied EF polarized the RhoA, the PKA inhibitor treatment abolished the directional response.
In summary, we have shown that EF stimulates the formation of adhesion molecules redistribution during cell migration, and cell migration directionality is dependent on integrin and RhoA polarization. Moreover RhoA redistribution is dependent on integrin-mediated signals. These findings have contributed to the understanding of some of the modes of action of EF in cell migration directionality.
Finally, our studies additionally identified waveform-induced disparate migration speed and directionality, and signaling molecules polarization under EF that will be mediators for EF-induced directionality. Results from this study may benefit our understanding the electro-therapy treatment on cell behavior and provide further treatment options.
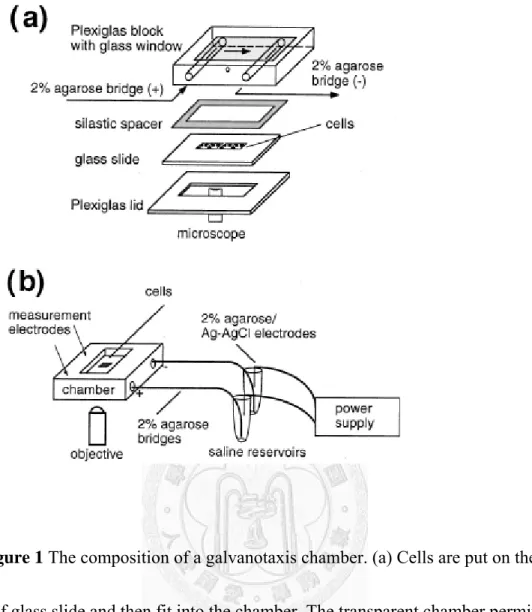
Figure 1 The composition of a galvanotaxis chamber. (a) Cells are put on the center of glass slide and then fit into the chamber. The transparent chamber permits direct observation for cell behaviors with a microscope. (b) The chamber is connected to saline reservoirs with 2% agarose salt bridge. We establish stable electric field across the chamber with external power supply. (Adaped from Chao PH, Roy R, Mauck RL, Liu W, Valhmu WB, & Hung CT (2000) J Biomech Eng 122, 261-267.)
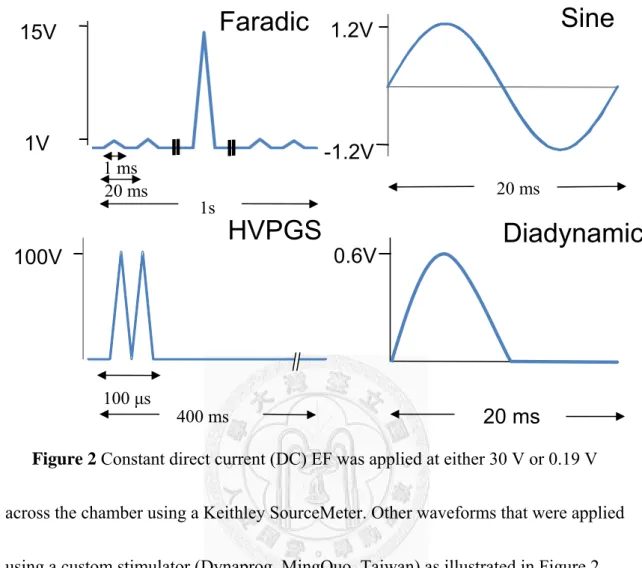
Figure 2 Constant direct current (DC) EF was applied at either 30 V or 0.19 V across the chamber using a Keithley SourceMeter. Other waveforms that were applied using a custom stimulator (Dynaprog, MingQuo, Taiwan) as illustrated in Figure 2.
All waveforms and the constant 0.19 V group except for HVPGS were controlled to have the same amount of total current flow (1.9×10-5As). Faradic current (Far) represented a graduation series of triangular waves peaking at 15 V at 50 Hz.
High-voltage pulsed galvanic stimulation (HVPGS) consists of monophasic,
twin-spike pulses that have a fixed pulse duration of 100 μs and max intensity at 100 V with a frequency of 2.5 Hz. Sinusoidal waves (sin) have a peak intensity of 1.2 V at 50 Hz and the diadynamic waves (diadyn) are rectified monophasicc sinusoidal waves
Faradic
HVPGS
Sine
Diadynamic
100 μs
400 ms
20 ms
20 ms 100V
1.2V
0.6V -1.2V 15V
1V
1s 20 ms
1 ms
Speed
Figure 3A Under electrical stimulation, ACL fibroblasts exhibited significantly different responses in migration. All groups except for the HVPGS group exhibited enhanced migration speed. When compared with the groups subjected to constant DC EFs, all the other groups were slower except for the diadynamic group (* means p<0.05 vs 0V control, § means p<0.05 vs. 30 V group, n=53-99 cells).
0 2 4 6
0V 30V 0.19V Far HVPGS sin diadyn
Speed ( μ m/hr)
* *
* * *
§ §
§
Directional velocity
Figure 3B In terms of directional velocity, all groups except for the HVPGS group exhibited enhanced migration directional velocity. When compared with the groups subjected to constant 30V DC EFs, all the other groups were slower (*
means p<0.05 vs 0V control, § means p<0.05 vs. 30 V group, n=53-99 cells).
Interestingly, no directionality was found with the constant 0.19 V EF group even though it has similarly enhanced migration speed with the 30 V group.
-6 -4 -2 0
Di rect iona l V el oc ity (μ m/hr )
*
§
§
* §
* §
* §
0V 30V 0.19V Far HVPGS sin diadyn
Figure 4 Morphological examination revealed differential cell shape responses.
In the constant 30 V group, ACL fibroblasts exhibited significant elongation and aligned perpendicular to the constant applied EF groups. All other groups did not exhibit any significant elongation or orientation (*p<0.05 vs. the 0V control, §p<0.05 vs. the 30 V group, n= 41-156 cells)
(a)
0 1 2 3
0V 30V 0.19V Far HVPGS sindiadyn
A spect Ratio
* *
§
0V 30V 0.19V
Faradic HVPGS sine
diadynamic (b)
§ §
§
20 μm
26
+ -
0.788 (a)
(b) 0.7 1.45
(C)
Figure 5 Under electrical field, most of waveforms redistributed integrin toward cathode except for HVPGS, 0.19V DC EF and sine wave. Sine wave induce integrin move toward anode. (a) immunostaining for integrin α2β1 (b) normalized integrin distribution (c) asymmetric Index (AI: I cathode- I anode). *p<0.05 vs. the 0V control,
§p<0.05 vs. the 30 V group, n= 8-30 cells
-1.5
-1 -0.5 0 0.5 1
0V 30V 0.19V Far HVPGS sin diadyn
*
* *
* §
§
§
§
Figure 6 effect of cholesterol treatment on FRAP in membrane. Typical FRAP curves (a) of DiI labeled membrane, with (red) or without cholesterol hemisuccinate (blue). Average T 1/2 values are shown (n= 15–20). Mean ± s.e.m. (n= 15–20) of the diffusion coefficient (D= A/ 4T1/2) are shown on the lower-left panels (*P< 0.05).
Cholesterol induced a decrease in the diffusion rates and fluorescence recovery intensity, accompanied by lower diffusion coefficient (b) and relative intensity (c).
0 0.2 0.4 0.6 0.8 1 1.2
con ChH
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4
con ChH
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
con ChH
Diffusion coefficient Relative intensity
Normalized fluorescence intensity
T
*
*
(a)
(b) (c)
T 1/2 = 10.615 s T 1/2 = 49.589 s
0.7 1.45
Figure 7 In 30V and sine group, both control and integrin antibody treatment can induce integrin redistribution, but cholesterol treatment cause integrin distribute randomly (a) (★p<0.05 vs. 0V, ※p<0.05 vs. the 30V control, §p<0.05 vs. the sine control group, n= 8-30 cells). Both ChH and antibody treatment, the migration speed is significant faster on the 30V DC EF and sine wave group (b), but they slower EF-induced directionality (c). (★p<0.05 vs. 0V, ※p<0.05 vs. the 30V control,
§p<0.05 vs. the sine control group, n= 71-194 cells) -1.5
-1 -0.5 0 0.5 1 1.5 2
0V 30V sine
★
0 1 2 3 4 5 6 7
0V 30V sine
-6 -5 -4 -3 -2 -1 0 1 2 3
Speed (μm/hr)AI
control ChH Antibody
★
★
Directional Velocity (μm/hr)
※
§
※ §
★ ★
※
★ §
★
★ ★
★ ★ ★
0V 30V sine(a)
(b) (c)
Figure 8 In short-term EF expose, both 5 min and 10 min period affect integrin redistribute toward cathode (* p< 0.05 to 0V control, § p< 0.05 to 10 min period 30V DC EF, n= 15-30 cells).
-1 -0.8 -0.6 -0.4 -0.2 0 0.2 0.4 0.6 0.8 1
0V 30V (5min) 30V (10min) 30V (1hr)
0.803
1.022 1.177 0.999
5 min
0.787 0.871
1.503 0.839
10 min
AI
*
* *
§
Figure 9 For 1hr DC EF, the RhoA redistributed toward cathode. Other than EF- induced RhoA cathodal distribution, sine wave induce RhoA move toward anode (*means p<0.05 vs. 0V, n= 10-20 cells). According to AI of RhoA and integrin distribution, the EF-induced RhoA redistribution coincides with integrin
-4 -3 -2 -1 0 1 2 3
0V 30V sine
-4 -3 -2 -1 0 1 2 3
-2 -1 0 1
AI of integrin distribution
0V 30V sine
1.013 1.179
1.007 0.801
0.363 0.933
1.843 0.86
2.04 0.387
0.975 0.598
AI
*
*
AI of RhoA distribution
sine 0V 30V
Figure 10 For 1hr exposure to 30V DC EF, the RhoA redistributed toward cathode, furthermore, inhibition of RhoA activation had a similar effect on RhoA redistribution in applied EF, but anti-integrin antibody treatment caused RhoA distribute randomly. (*means p<0.01 vs. all other groups, § means p<0.01 vs. DC group, n=10-20 cells)
-0.5 0 0.5 1 1.5 2 2.5 3
no EF DC DC+AB DC+KT5720
*
*§
AI
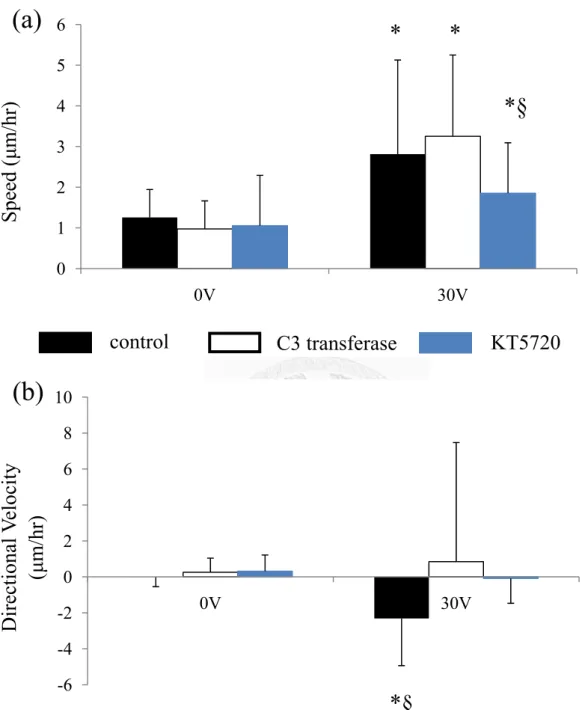
Figure 11 Inhibition of RhoA and PKA activation by C3 transferase and KT 5720 had similar effect on cell migration. The migration speed is significant faster on the 30V DC EF (a) but they slower EF-induced directionality (b). (*p<0.05 vs. 0V,
§p<0.05 vs. other treatment groups, n= 35-118 cells) 0
1 2 3 4 5 6
0V 30V
-6 -4 -2 0 2 4 6 8 10
0V 30V
Speed (μm/hr)
control C3 transferase KT5720
Directional Velocity (μm/hr)
* *
*§
(a)
(b)
*§
Reference
1. Kloth, L.C., Electrical stimulation for wound healing: a review of evidence
from in vitro studies, animal experiments, and clinical trials. Int J Low Extrem
Wounds, 2005. 4(1): p. 23-44.
2. Aaron, R.K., D.M. Ciombor, and B.J. Simon, Treatment of nonunions with
electric and electromagnetic fields. Clin Orthop Relat Res, 2004(419): p. 21-9.
3. Akai, M., et al., Electrical stimulation of ligament healing. An experimental
study of the patellar ligament of rabbits. Clin Orthop Relat Res, 1988(235): p.
296-301.
4. Chao, P.H., et al., Effects of applied DC electric field on ligament fibroblast
migration and wound healing. Connect Tissue Res, 2007. 48(4): p. 188-97.
5. Carley, P.J. and S.F. Wainapel, Electrotherapy for acceleration of wound
healing: low intensity direct current. Arch Phys Med Rehabil, 1985. 66(7): p.
443-6.
6. Campbell, C.E., D.V. Higginbotham, and T.J. Baranowski, Jr., A constant
cathodic potential device for faradic stimulation of osteogenesis. Med Eng
Phys, 1995. 17(5): p. 337-46.
7. Ciullo, J.V. and B. Zarins, Biomechanics of the musculotendinous unit:
relation to athletic performance and injury. Clin Sports Med, 1983. 2(1): p.
71-86.
8. Philipson, T., et al., [The effect of diadynamic current on chronic soft-tissue
pain in the neck and shoulder girdle]. Ugeskr Laeger, 1983. 145(7): p. 479-81.
9. Lisinski, P., W. Zapalski, and W. Stryla, [Physical agents for pain
management in patients with gonarthrosis]. Ortop Traumatol Rehabil, 2005.
7(3): p. 317-21.
10. Romanenko, S.G., O.P. Tokarev, and S. Vasilenko Iu, [Electrostimulation of
laryngeal muscles with fluctuating currents in the treatment of patients with
unilateral laryngeal paralysis]. Vestn Otorinolaringol, 2001(3): p. 52-4.
11. Rusiaev, V.F., Z.N. Salienko, and V.F. Pavlenko, [Effect of a diadynamic
current on the development of thromboembolism]. Patol Fiziol Eksp Ter,
1983(1): p. 10-4.
12. Dobrova, A.M., et al., [Use of diadynamic currents to treat suppurating
wounds]. Sov Med, 1979(9): p. 55-8.
13. Brown, M., et al., High-voltage galvanic stimulation on wound healing in
guinea pigs: longer-term effects. Arch Phys Med Rehabil, 1995. 76(12): p.
1134-7.
14. Gogia, P.P., R.R. Marquez, and G.M. Minerbo, Effects of high voltage
galvanic stimulation on wound healing. Ostomy Wound Manage, 1992. 38(1):
p. 29-35.
15. Hynes, R.O., Integrins: versatility, modulation, and signaling in cell adhesion.
Cell, 1992. 69(1): p. 11-25.
16. Cho, M.R., et al., Integrin-dependent human macrophage migration induced
by oscillatory electrical stimulation. Ann Biomed Eng, 2000. 28(3): p. 234-43.
17. Sun, S. and M. Cho, Human fibroblast migration in three-dimensional
collagen gel in response to noninvasive electrical stimulus. II. Identification of
electrocoupling molecular mechanisms. Tissue Eng, 2004. 10(9-10): p.
1558-65.
18. Cho, M.R., et al., Induced redistribution of cell surface receptors by
alternating current electric fields. FASEB J, 1994. 8(10): p. 771-6.
19. Giancotti, F.G. and E. Ruoslahti, Integrin signaling. Science, 1999. 285(5430):
p. 1028-32.
20. Calderwood, D.A., S.J. Shattil, and M.H. Ginsberg, Integrins and actin
filaments: reciprocal regulation of cell adhesion and signaling. J Biol Chem,
2000. 275(30): p. 22607-10.
21. Carman, C.V. and T.A. Springer, Integrin avidity regulation: are changes in
affinity and conformation underemphasized? Curr Opin Cell Biol, 2003. 15(5):
p. 547-56.
22. Yamada, K.M. and S. Miyamoto, Integrin transmembrane signaling and
cytoskeletal control. Curr Opin Cell Biol, 1995. 7(5): p. 681-9.
23. Ridley, A.J. and A. Hall, The small GTP-binding protein rho regulates the
assembly of focal adhesions and actin stress fibers in response to growth
factors. Cell, 1992. 70(3): p. 389-99.
24. Hotchin, N.A. and A. Hall, The assembly of integrin adhesion complexes
requires both extracellular matrix and intracellular rho/rac GTPases. J Cell
Biol, 1995. 131(6 Pt 2): p. 1857-65.
25. Yang, B., et al., p190 RhoGTPase-activating protein links the beta1
integrin/caveolin-1 mechanosignaling complex to RhoA and actin remodeling.
Arterioscler Thromb Vasc Biol. 31(2): p. 376-83.
26. Whitehead, I.P., et al., Dbl family proteins. Biochim Biophys Acta, 1997.
1332(1): p. F1-23.
27. Ren, X.D., W.B. Kiosses, and M.A. Schwartz, Regulation of the small
GTP-binding protein Rho by cell adhesion and the cytoskeleton. EMBO J,
1999. 18(3): p. 578-85.
28. Barry, S.T., et al., Requirement for Rho in integrin signalling. Cell Adhes Commun, 1997. 4(6): p. 387-98.
29. Chong, L.D., et al., The small GTP-binding protein Rho regulates a
phosphatidylinositol 4-phosphate 5-kinase in mammalian cells. Cell, 1994.
79(3): p. 507-13.
30. Clark, E.A., et al., Integrin-mediated signals regulated by members of the rho
family of GTPases. J Cell Biol, 1998. 142(2): p. 573-86.
31. Pozo, D., et al., Identification of G-protein coupled receptor subunits in
normal human dental pulp. J Endod, 2000. 26(1): p. 16-9.
32. Kumanogoh, H., et al., Biochemical and morphological analysis on the
localization of Rac1 in neurons. Neurosci Res, 2001. 39(2): p. 189-96.
33. Carmena, M.J., et al., Cholesterol modulation of membrane fluidity and VIP
receptor/effector system in rat prostatic epithelial cells. Regul Pept, 1991.
33(3): p. 287-97.
34. Axelrod, D., et al., Mobility measurement by analysis of fluorescence
photobleaching recovery kinetics. Biophys J, 1976. 16(9): p. 1055-69.
35. Sprague, B.L., et al., Analysis of binding reactions by fluorescence recovery
after photobleaching. Biophys J, 2004. 86(6): p. 3473-95.
36. McLaughlin, S. and M.M. Poo, The role of electro-osmosis in the
electric-field-induced movement of charged macromolecules on the surfaces of
cells. Biophys J, 1981. 34(1): p. 85-93.
37. Bourguignon, G.J. and L.Y. Bourguignon, Electric stimulation of protein and
DNA synthesis in human fibroblasts. FASEB J, 1987. 1(5): p. 398-402.
38. Zhao, M., et al., Membrane lipids, EGF receptors, and intracellular signals
colocalize and are polarized in epithelial cells moving directionally in a
physiological electric field. FASEB J, 2002. 16(8): p. 857-9.
39. van Ginkel, G., H. van Langen, and Y.K. Levine, The membrane fluidity
concept revisited by polarized fluorescence spectroscopy on different model
membranes containing unsaturated lipids and sterols. Biochimie, 1989. 71(1):
p. 23-32.
40. Yeagle, P.L., Lipid regulation of cell membrane structure and function.
FASEB J, 1989. 3(7): p. 1833-42.
41. Lang, P., et al., Protein kinase A phosphorylation of RhoA mediates the
morphological and functional effects of cyclic AMP in cytotoxic lymphocytes.
EMBO J, 1996. 15(3): p. 510-9.
42. Machacek, M., et al., Coordination of Rho GTPase activities during cell
protrusion. Nature, 2009. 461(7260): p. 99-103.
43. Fukata, M., M. Nakagawa, and K. Kaibuchi, Roles of Rho-family GTPases in
cell polarisation and directional migration. Curr Opin Cell Biol, 2003. 15(5):
p. 590-7.