Received 06 September 2005 | Accepted 4 September 2006 | Published 26 October 2006
Organogenesis results from a succession of complex pro-cesses including induction, patterning, specification, morpho-genesis, and differentiation [1]. Transcription factors includ-ing Pax6, Optx2, and Crx play important roles at different de-velopmental stages of oculogenesis and are ultimately impor-tant in cell fate decisions [2,3]. Mutations of these factors in-duce ocular developmental anomalies in humans, including aniridia, anophthalmia, and Leber congenital amaurosis [2].
Most important genes are evolutionally conservative, al-though functions of some genes may not be identical among species. Research in zebrafish (Danio rerio) is feasible for functional genomics and reverse genetics [3,4] because the fish develops rapidly, reaching maturity in three months. Fur-thermore, the fish can be induced to spawn by photoperiod, and has fecund, big, transparent, extra-uterine eggs that are suitable for studying embryonic development and gene modu-lation [1,3,5,6]. The development of the zebrafish brain,
ner-vous, and visual systems is similar to that of other vertebrates [6]. The remarkable evolutionary conservation among verte-brate eyes provides the basis for using zebrafish as a model system for detection and analysis of genetic defects related to human eye disorders [1,7], such as retinitis pigmentosa, cone dystrophy, and macular degeneration [3,8]. Zebrafish retina and lens show much the same morphology as other vertebrates including humans [7]. Because the zebrafish retina is simple and develops fast, in addition to its accessibility to genetic analysis, it has become an excellent model system for study-ing retinal neurogenesis in vertebrates [9]. The appearance of retinal ganglion cells between 27 and 28 h postfertilization (hpf) is the initial event in zebrafish retinal neurogenesis [3]. After 10 h, interneurons emerge and form the inner nuclear layer. At about 43 hpf, photoreceptor cells appear in the outer nuclear layer. Zebrafish retina becomes functional between 60 and 80 hpf [3,10,11]. The emmetropization of eye and lens is achieved at 72 hpf [10]. By then most embryos have hatched [12].
The reverse genetic approach is faster and discloses a more precise structure-function relationship than forward genetics
Egr1 gene knockdown affects embryonic ocular development in
zebrafish
Chao-Yu Hu,1 Chang-Hao Yang,2 Wei-Yu Chen,3 Chiu-Ju Huang,4 Hsing-Yen Huang,4 Muh-Shy Chen,2
Huai-Jen Tsai4
1Department of Ophthalmology, Far Eastern Memorial Hospital, 2Department of Ophthalmology, National Taiwan University
Hos-pital, 3Department of Pathology, Taipei Medical University, 4Institute of Molecular and Cellular Biology, National Taiwan
Univer-sity, Taipei, Taiwan
Purpose: To identify the changes in zebrafish embryonic ocular development after early growth response factor 1 (Egr1)
gene knockdown by Egr1-specific translation inhibitor, morpholino oligonucleotides (MO).
Methods: Two kinds of Egr1-MO were microinjected separately with various dosages into one to four celled zebrafish
embryos to find an optimal dose generating an acceptable mortality rate and high frequency of specific phenotype. Chordin-MO served as the positive control; a 5 mismatch Chordin-MO of Egr1-Chordin-MO1 and a nonspecific Chordin-MO served as negative controls. We graded the Egr1 morphants according to their gross abnormalities, and measured their ocular dimensions accordingly. Western blot analysis and synthetic Egr1 mRNA rescue experiments confirmed whether the deformities were caused by Egr1 gene knockdown. Histological examination and three kinds of immunohistochemical staining were applied to iden-tify glutamate receptor one expression in retinal ganglion cells and amacrine cells, to recognize acetylated α-tubulin
expression which indicated axonogenesis, and to label photoreceptor cells with zpr-1 antibody.
Results: After microinjection of 8 ng Egr1-MO1 or 2 ng Egr1-MO2, 81.8% and 97.3% of larvae at 72 h postfertilization
had specific defects, respectively. The gross phenotype included string-like heart, flat head, and deformed tail. The more severely deformed larvae had smaller eyes and pupils. Co-injection of 8 ng Egr1-MO1 and supplementary 12 pg synthetic Egr1 mRNA reduced the gross abnormality rate from 84.4% to 29.7%, and decreased the severity of deformities. Egr1 protein appeared in the wildtype and rescued morphants, but was lacking in the Egr1 morphants with specific deformities. Lenses of Egr1 morphants were smaller and had some residual nucleated lens fiber cells. Morphants’ retinal cells arranged disorderly and compactly with thin plexiform layers. Immunohistochemical studies showed that morphants had a mark-edly decreased number of mature retinal ganglion cells, amacrine cells, and photoreceptor cells. Retinal axonogenesis was prominently reduced in morphants.
Conclusions: The Egr1 gene plays an important role in zebrafish embryonic oculogenesis. Ocular structures including
lens and retina were primitive and lacked appropriate differentiation. Such arrested retinal and lenticular development in Egr1 morphants resulted in microphthalmos.
Correspondence to: Huai-Jen Tsai, PhD, Institute of Molecular and Cellular Biology, National Taiwan University, No.1, Sec. 4, Roosevelt Road, Taipei, Taiwan 106; Phone: 23641705; FAX: +886-2-23638483, email: [email protected]
[3]. Morpholino oligonucleotides (MO) are designed to bind a target mRNA, resulting in translation blockage by prevent-ing 40S subunit of ribosomes from bindprevent-ing, and ultimately suppress target gene expression. Alternatively, it can be de-signed to bind the junction of intron and exon of pre-mRNA, and causes the splicing machinery to effectively skip an entire exon [3,13,14]. Currently MO is the most widely used ap-proach for studying the loss-of-function of zebrafish genes [13,15,16].
Early growth response factor 1 (Egr1) gene (also known as NGFI-A, zif268, Krox24, tis8, cef5, and ZENK) is an im-mediate early transcriptional regulator gene [17]. This factor encodes a highly conserved C2-H2 zinc finger DNA-binding protein, which is necessary for vertebrate development; this then leads to proliferation and differentiation [18-20]. How-ever, the functions of Egr1 on oculogenesis are still unclear. Postnatally induced myopia by a diffuser or a minus lens in chicks was associated with elongated eyeballs and decreased ZENK expression in retinal glucagon-containing amacrine cells [21,22]. By contrast, the Egr1 mRNA and protein, espe-cially in ganglion cells and bipolar cells, were strongly regu-lated by the transitions between light and darkness in form-deprived myopic mice, whereas image contrast exerted only a minor influence on Egr1 expression [23]. Since the mecha-nisms of eye growth control are different among species, stud-ies of gene expression patterns between specstud-ies during emmetropization and myopic progression are valuable [24]. By using NCBI protein-protein BLAST, the zebrafish Egr1 protein (NCBI accession number P26632) shares 55%, 56%, and 62% identities of amino acid sequence with human Egr1 protein (P18146), mouse Egr1 protein (NP_031939), and chicken ZENK protein (NP_989467), respectively. However, they share at least 95% identity in the three zinc finger C2-H2 domains that bind to the major groove of B-DNA [25]. In the zebrafish embryo, Egr1 mRNA is initially detected in the presomitic mesoderm between 11 and 20 hpf [19,26]. It is expressed in hypothalamus, mesencephalon, tegmentum, phar-ynx, otic vesicle, and heart between 24 and 48 hpf [19]. In retina, it is observed at 40 hpf and disappears at 48 hpf [19].
In this study, we microinjected MO targeting at Egr1-mRNA into zebrafish embryos and observed their changes in appearance and ocular histology after loss of Egr1 gene func-tion. Our findings should be helpful to further research in ocular development.
METHODS
Animals and embryos: Adult AB-strain zebrafish were bred with a controlled light cycle of 14 h of light and 10 h of dark-ness at 28.5 °C [26,27]. After the lights were turned on, we
removed the existing embryos, collected the newly laid eggs within 2 to 3 h, and then sorted them into petri dishes accord-ing to the developmental stages defined by Kimmel et al. [12]. The one to four celled embryos were used for microinjection [26]. All procedures conformed to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
Microinjection of morpholino oligonucleotides: Micro-injection is an effective delivery system for MO in zebrafish
[28]. In this study, two different sequences of Egr1-MO were produced by Gene Tools (Philomath, OR) referring to the ge-nomic DNA sequence reported by Drummond et al. (GenBank accession number U12895) [18]. The Egr1-MO1 is composed of 5'-GCA GCC ATC TCT CTG GAG TGT GCT C, which is the antisense of the last 17 nucleotides in the 5'-untranslated region (5'-UTR) and the leading 8 nucleotides of coding re-gion. The Egr1-MO2 is composed of 5'-GAG AGA ATT GTG CTG GAG AAA TCA G, which is complementary to the 5'-UTR sequence of Erg1 located at the upstream 24 to 48 nucle-otides from the start code (ATG). Since their effects were pre-sumed to be dose-dependent, various dosages of Egr1-MO, mixed with Phenol Red, were microinjected with Nanoject II (Drummond, Broomall, PA) into one to four celled zebrafish embryos to find an optimal dose generating a low mortality rate and a high frequency of specific phenotype [28]. We also microinjected 4.5 ng of chordin-MO per embryo as a positive control [28]. Theoretically, at least 75% of chordin morphants at 72 hpf should have severe defects including expanded blood island, u-shaped somites, reduced head size, and an abnormal tail fin with multiple folds [28]. A 5 base mismatch MO (5'-GCA CCC ATG TCT GTG GAC TGT CCT C) of Egr1-MO1 and a nonspecific MO (5'-CCT CTT ACC TCA GTT ACA ATT TAT A) served as negative controls [27,28]. These morphants and wildtypes were raised with a similar density of embryos in incubators at 28.5 °C.
Measurement of ocular dimensions: To decrease the pos-sible individual differences, we compared the ocular dimen-sions between the wildtype and Egr1 morphants collected from the same batch of embryos and cultured them in a similar den-sity under the same circumstances. After anesthesia with 0.02% volume of 2-phenoxyethanol (Sigma-Aldrich, St. Louis, MO) for 1 min, the 72-hpf larvae were observed with a stereo dis-secting microscope (MZFL III, Leica, Wetzlar, Germany) and photographed with a digital camera (FinePix S2pro, Fujifilm, Tokyo, Japan). Parameters were measured with image-pro-cessing software (PhotoImpact 6.0, Ulead, Taipei, Taiwan). The severity of the deformity was graded according to the changes in appearance. We measured the ocular and pupillary diameters of the left eye, which represented the gross devel-opment of retinal and lenticular structures, respectively [29]. The horizontal and vertical diameters were determined accord-ingly with the maximal length of line segments parallel to, or right-angled with the assumed line connecting its mouth and otic capsule. These measurements were analyzed with one-way analysis of variance, and the differences between every two groups were verified with Scheffe’s posthoc test (SPSS 11.0, SPSS, Chicago, IL). P values less than 0.05 were con-sidered significant.
Rescue experiment of Egr1 mRNA: The total RNA of 48-hpf zebrafish embryos was extracted with Trizol (ProTech, Taipei, Taiwan) and treated with RNase-free DNase (Promega, Madison, WI). The first strand of cDNA was obtained with the reaction of total RNA, oligo(dT)12-18 primers, dNTP, and M-MLV reverse transcriptase (Promega). The coding region of Egr1 DNA was acquired with polymerase chain reaction (PCR) using VioTaq DNA polymerase (Viogene, Taipei,
wan), the above product after reverse transcription and the primers of CCA GCA TGC ATC TCT CCT CTC CGA GCA CAC TCC AGA G and TTT CTG ACT CGA GGG AGA GCT TGG AGT TCT GGG GAA G. The expected 1.6-kb PCR prod-uct was extracted with a gel extraction kit (Geneaid, Taoyuan, Taiwan) and then ligated to the pGEM-T Easy vector (Promega). The resultant plasmid pGEM-egr1 was obtained from Escherichia coli DH5α after transformation. The Egr1
DNA was separated from pGEM-egr1 by SacII and was
tran-scribed to 5'-capped Egr1 mRNA with 3'-poly(A) tail with a MESSAGE mMACHINE™ Kit (Ambion, Austin, TX). Vari-ous amounts of the in vitro synthetic Egr1 mRNA, mixed with the selected dose of Egr1-MO, were microinjected into one to four celled embryos. The abnormality rates at 72 hpf were compared to confirm the effect of Egr1 mRNA rescue.
Western blot analysis: To establish the effect of Egr1 gene knockdown by Egr1-MO, we compared the amounts of 55-kD Egr1 protein (NCBI accession number NP_571323)
Figure 1. Morphological phenotypes of the wildtype and Egr1 morphants. Gross appearance of the wildtype (A) and Egr1 morphants (B: Grade 1, C: Grade 2, D: Grade 3, E: Grade 4, and F: Grade 5) at 72 h postfertilization. The grading criteria are defined in the Results section. The scale bar represents 100 µm in photo A, and is applicable to all photos.
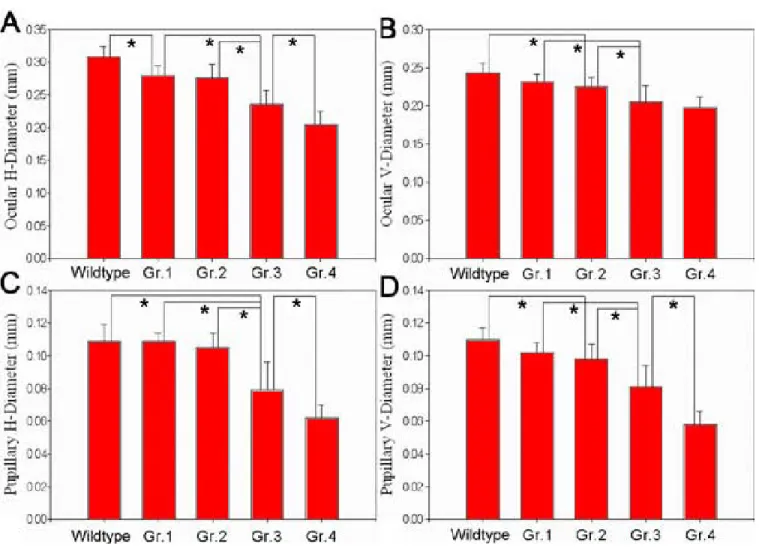
TABLE 1. COMPARISONOFOCULARDIMENSIONSAMONGWILDTYPEANDVARIOUSGRADESOF EGR1-MORPHOLINO-OLIGONUCLEOTIDE-INJECTED EMBRYOSAT 72 HPOST-FERTILIZATION
Wildtype Grade 1 Grade 2 Grade 3 Grade 4 P (n=20) (n=19) (n=13) (n=13) (n=20) value* ---- --- --- --- --- --- ---Eye H-diameter 0.31±0.02 0.28±0.01 0.28±0.02 0.24±0.02 0.21±0.02 <0.001 V-diameter 0.24±0.01 0.23±0.01 0.23±0.01 0.21±0.02 0.20±0.02 <0.001 H:V ratio 1.27±0.04 1.21±0.05 1.22±0.04 1.16±0.13 1.04±0.07 <0.001 Pupil H-diameter 0.11±0.01 0.11±0.01 0.11±0.01 0.08±0.02 0.06±0.01 <0.001 V-diameter 0.11±0.01 0.10±0.01 0.10±0.01 0.08±0.01 0.06±0.01 <0.001 H:V ratio 1.00±0.05 1.07±0.08 1.07±0.06 0.98±0.13 1.07±0.14 0.009
Values are means±SD in mm. Grading criteria are defined in the Results section. H represents horizontal, V represents vertical. Asterisk (*)
represents one-way analysis of variance.
between the Egr1-MO-injected embryos and the wildtype em-bryos at 24, 48, and 72 hpf. We also evaluated its expression in the mRNA rescued morphants. The loading dose of extracted total proteins was quantified separately by absorbance mea-surements at 595 nm (DU 640B, Beckman Coulter, Fullerton, CA). One piece of gel after SDS-PAGE served for coomassie blue stain to determine adequate loading. The other part of gel was blotted onto a polyvinylidene difluoride membrane, hy-bridized with the Egr1 antibody (sc-110, Santa Cruz Biotech-nology, Santa Cruz, CA) at 1:1000 dilution, washed exten-sively with Tris buffered saline, incubated with horseradish peroxidase-conjugated secondary antibody (number 12-348, Upstate Biological Inc., Lake Placid, NY) at 1:5000 dilution, and finally developed by using enhanced chemiluminescence (SuperSignal West Pico Chemiluminescent Substrate, Pierce, Rockford, IL).
Histological examination and immunohistochemical staining: The 72-hpf larvae derived from Egr1-MO-injected embryos and from the wildtype were fixed in 4% paraformal-dehyde, embedded in paraffin, cut into 5 µm sections, stained
with hematoxylin and eosin. They were examined with a light
microscope (BX51, Olympus, Tokyo, Japan) and photographed with a camera (PM-C35DX, Olympus). To identify the differ-entiation of retinal ganglion cells and amacrine cells, we per-formed immunohistochemical staining using 3 µg/ml of
glutamate receptor 1 antibody (AB-1504, Chemicon, Temecula, CA) at 4 °C overnight [26,30]. In addition, the
axonogenesis was identified by acetylated α-tubulin antibody
(T6793, Sigma-Aldrich) at 1:400 dilution, 4 °C overnight
[31,32]. After deparaffinization, rehydration, and antigen re-trieval with water bath in 121 °C citrate buffer for 5 min, the
sections were washed with phosphate buffered saline, and in-cubated with those primary antibodies. The sections then were incubated sequentially with the biotinylated horseradish per-oxidase, avidin, and diaminobenzidine following the manufacturer’s instructions (Ultratech HRP Kit, Immunotech, Marseilles, France). Finally, the sections were counterstained with hematoxylin and recorded with a camera mounted on a light microscope. On the other hand, to identify the differen-tiation of photoreceptor cells, we immersed the 72-hpf wildtype larvae and Egr1 morphants in OCT separately, stored them in -20 °C freezer, and cryosectioned them into 6 µm slices. The
Figure 2. Ocular dimensions of the wildtype and Egr1 morphants with different grades of deformity at 72 h postfertilization. The asterisks point out a significant difference with Scheffe’s posthoc test between the bars of differing Grades. A: Ocular horizontal diameter, B: ocular vertical diameter, C: pupillary horizontal diameter, and D: pupillary vertical diameter.
zpr-1 antibody labeled most cone cells at 1:100 dilution for 2 h at room temperature [33].
RESULTS
The effect of morpholino oligonucleotide injection on embry-onic development: About 82.6% of 72-hpf larvae (n=113) af-ter chordin-MO injection exhibited severe defects, whereas
only less than 1% of the nonspecific negative control (n=166) changed in appearance. In a pilot study, we microinjected vari-ous amount of Egr1-MO into the one to four celled embryos and found that higher dose of Egr1-MO produced more se-verely deformed embryos and more deaths. We chose 8 ng of Egr1-MO1 and 2 ng of Egr1-MO2 per embryo as the study concentration because they generated an acceptable mortality rate (<10%) and a high specific abnormality rate (>80%). The most common abnormalities that appeared in the Egr1-MO-injected embryos were enlarged pericardium, small eye, and
Figure 4. Western blot analysis of the Egr1 protein. A 55-kD Egr1 protein appears in wildtype (WT) embryos at 24, 48, and 72 h postfertilization (hpf), whereas the Grade-3 Egr1 morphants (Mo) do not exhibit this at the same periods. After Egr1 mRNA rescue, Egr1 protein appeared in the 48-hpf rescued morphants (ReMo). The leftmost lane represents the molecular weight marker (M). Figure 3. The effects of Egr1 mRNA rescue. Coinjection of 8 ng
Egr1-MO1 and 0 (n=142), 2 (n=194), 6 (n=179), 10 (n=180), 12 (n=139), or 14 (n=190) pg of synthetic Egr1 mRNA caused different survival rates and abnormality rates of embryos at 72 h postfertilization.
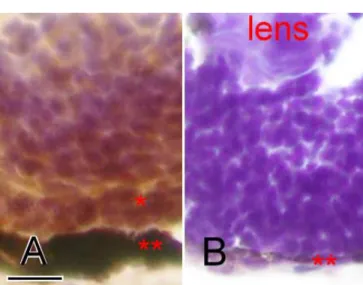
Figure 5. Histological exami-nation of the wildtype and Egr1 morphants. Hematoxylin and eosin stained transverse sec-tions of zebrafish eyes at 72 h postfertilization having A: wildtype, B: Grade 1, C: Grade 2, D: Grade 3, or E: Grade 4. Higher-grade morphants have smaller eyes and smaller lenses. Both the inner plexi-form layer between the gan-glion cell layer (gcl) and inner nuclear layer (inl) as well as the outer plexiform layer between the inl and outer nuclear layer (*) are thin. The outermost reti-nal pigment epithelium (**) layer is also thin and irregular. Scale bar represents 100 µm in
photo A, and is applicable to all photos.
deformed body axis. According to the severity of gross defor-mities at 72 hpf, Egr1 morphants were classified into five grades (Figure 1):
Grade 1: No difference between the morphant and wildtype; Grade 2: Enlarged pericardium only; Grade 3: Mildly deformed body axis; Grade 4: Markedly deformed body axis; Grade 5: Agenesis of tail or eyes.
About 81.8% of the 8 ng-Egr1-MO1-injected embryos (n=142) and 97.3% of the 2 ng-Egr1-MO2-injected embryos (n=400) exhibited Grade 3 or more severe deformity, whereas only 2.5% of embryos (n=200) injected with 8 ng of our five nucleotide mismatch control MO showed abnormal appear-ance. Besides pericardial edema found in Grade-2 morphants, Grade 3 morphants additionally had a string-like heart, bent tail and flat head. Similar deformities appeared but were more severe in Grade 4 morphants.
Egr1-morphants with higher grade states had smaller eyes and pupils (Table 1, Figure 2). These changes were especially prominent in Grade 2 and above. The horizontal ocular diam-eter was the most sensitive indicator of Egr1 depression be-cause it reduced significantly even when there was no detect-able change in appearance of the Grade-1 morphants. It also decreased more significantly than did the vertical ocular di-ameter. By contrast, the horizontal pupillary diameter changed in proportion to the vertical pupillary diameter.
Expression of Egr1 gene in wildtype, morphants, and Egr1-mRNA-rescued morphants: Coinjecting various amounts of synthetic Egr1 mRNA and 8 ng Egr1-MO1 resulted in
dif-ferent rescue effects (Figure 3). When 12 pg of Egr1 mRNA was supplied, the abnormality rate decreased prominently from 84.4% to 29.7% without altering the survival rate. Among the larvae with abnormalities in appearance, the average grade of deformity in the rescued morphants also decreased markedly from 3.5 to 2.0.
Western blot analysis demonstrated that the wildtype had the presumed 55-kD Egr1 protein at 24, 48, and 72 hpf, whereas the Grade-3 Egr1 morphants did not (Figure 4). When supple-mentary 12 pg Egr1 mRNA was coinjected with 8 ng Egr1-MO1 into one to four celled embryos, the Egr1 protein ap-peared in the rescued morphants (Figure 4). None of our blot-ting showed a band like the 75 kDa nuclear Egr1 protein found in mouse retina [34].
Histological observation and immunohistochemical stain-ing: In histological examination, lenses of the 72-hpf Egr1 morphants were smaller than those of the wildtype and had some residual nucleated lens fiber cells; however, their nas were more markedly affected (Figure 5). Morphants’ reti-nal cells arranged disorderly and compactly with little extra-cellular matrix, but there were no apparent vacuoles or py-knotic cells in their retinas. Their inner and outer plexiform layers were thin. The retinal pigmented epithelium (RPE) layer was also thin and irregular. In the morphants of higher-grade deformity, these changes became more prominent. The Grade 4 morphants were even devoid of retinal lamination.
Immunohistochemical study showed that morphants’ retina had markedly less glutamate receptor 1 expression,
Figure 6. Immunohistochemi-cal staining of the wildtype and Egr1 morphants. Horizontal sections of zebrafish eyes at 72 h postfertilization with immu-nohistochemical stain for glutamate receptor 1 (A, B) and acetylated α-tubulin (C,
D). Compared with the
wildtype (A, C), retinal cells of the Grade-3 Egr1 morphant (B,
D) arranged more compactly
and disorderly. Significantly smaller areas of staining for both glutamate receptor 1 and acetylated α-tubulin appear in
the morphant’s retina. Scale bar represents 100 µm in panel
A, and is applicable to all
pan-els.
namely decreased cellular differentiation of ganglion cells and amacrine cells (Figure 6A,B). The reduced expression of acety-lated α-tubulin in retina represented decreased axonogenesis
(Figure 6C,D). The remaining retinal axonongenesis of morphants was sometimes found in the presumed ganglion cell layer or inner nuclear layer. Zpr-1 antibody can label most cone cells in the wildtype, but it failed in the Grade 3 morphants (Figure 7). The decrement of all protein markers in the present study was parallel to the increasing grading and became obvi-ous in the Grade 3 morphants and higher.
DISCUSSION
MO has been applied successfully in zebrafish genetic stud-ies [3,27]. The increasing number of defined gene sequences and the success of MO technology in zebrafish open the door for modeling nearly any inherited developmental defect [8,16,28,35,36]. Previous researchers have successfully ap-plied this technique to prove the essential role of receptor pro-tein-tyrosine phosphatase α in zebrafish retinal development
[32]. In the present study, Egr1-MO effectively inhibited Egr1 gene expression for at least three days and generated pheno-types including string-like heart, small eyes, flat head, and deformed tail. The vertical eye diameter of wildtype in this study is comparable with a former histological study that mea-sured 240 µm at 74 hpf [10]. Small ocular and pupillary
di-mensions of Egr1 morphants suggest arrested retinal and len-ticular development, respectively [29]. Such reduced retinal and lenticular mass generated the picture of microphthalmos. The disarranged retinal cells were primitive proven by the depressed axonogenesis, and decreased cellular differentia-tion in ganglion cells, amacrine cells, and photoreceptor cells. Most of the Egr1 morphants could not survive longer than 96 h and did not hatch out. Such a fatal “inborn error” prohibited further evaluation [21-23]. It is reasonable that if a vital gene is inhibited immediately after MO injection, embryos cannot survive long or recover from their severe developmental re-tardation. Although few low-grade hypomorphic larvae sur-vived for more than seven days, their changes in expression were too obscure to be differentiated. In addition, because MO spreads evenly in the embryo [13,28], we cannot confine its effect to a specific tissue only. In this study, the defective phe-notypes we observed are a result of Egr1 gene knockdown in the whole body. Nevertheless, we cannot rule out the possible influence from other tissues. Such problems should also exist in knockout animals, although researchers did not find that Egr1 knockout mice had obvious developmental or behavioral defects except female infertility [37,38]. To conquer the prob-lems of limited temporal and unlimited spatial expression, we planned to apply a tetracycline-controlled transcriptional regu-lation system [39,40]; however, the lack of a cell-type-spe-cific promoter of the presumed target cells like the glucagon-containing amacrine cell prohibited its use [22]. The other method of specific gene knockdown, double-strand RNA in-terference (RNAi), has been also considered [3,41]. But in zebrafish, RNAi is limited by frequent nonspecific effects [3,36,42,43]. Other shortcomings includes problems of deliv-ery, off-target effects, poor tissue distribution, varying response
between genes, and difficulty in tackling neurological targets [44].
The Egr1 gene is multipotent and important for cell pro-liferation, differentiation, embryonic ocular development, and neuronal plasticity [18-20,45]. As an inducible transcription factor, its expression can be used as a functional assay for neuronal inputs and receptor systems [46]. Basal retinal Egr1 expression is maintained by synaptic plasticity occurring in response to physiological stimuli, and it is higher in the highly visual animals like monkeys [46,47]. In adult monkey retinas, the Egr1 protein is located mainly in cells of the inner nuclear layer and also in the ganglion cells. Amacrine cells are likely to mediate neural plasticity [47]. However, Zhong et al. [48] showed that at least 90% of Egr1 protein was found in the ON-bipolar cells and the remaining in the GABAergic ama-crine cells of infant macaque retinas. These cells were stimu-lated to produce more Egr1 when retinal images were enriched in focus without form-deprivation [48]. Similarly, Egr1 pro-tein in young mice appears in retinal ganglion cells, amacrine cells, and bipolar cells [23]. However, mouse retinal Egr1 mRNA and protein, especially in ganglion cells and bipolar cells, are strongly regulated by the transitions between light and darkness, whereas differences in retinal illumination ei-ther play no role or only a minor one [23]. Rat retinal Egr1 mRNA also increases transiently after the rat is exposed to light from the dark [49], but enhanced environments dramati-cally increase the Egr1 expression in retinal ganglion cell layer and inner nuclear layer of young rats [50]. The number of ZENK-expressing bipolar cells and amacrine cells in chick retinas increase after light onset and declined later [22]. The light intensity but not focus-defocus modulates ZENK syn-thesis in chick retinal ON-bipolar cells [22]. Thus, there exist
Figure 7. Cryosection and immunostaining of the wildtype and Egr1 morphants. Cryosections of zebrafish eyes at 72 h postfertilization with zpr-1 immunostaining for photoreceptor cells. The wildtype (A) has markedly more labeled retinal cells in the outer nuclear layer (*) than the Grade-3 Egr1 morphant (B). The retinal pigmented epithe-lial layer (**) is also much thinner in the morphant. The scale bar represents 10 µm in photo A, and is applicable to photo B.
minor but notable differences in Egr1 response among spe-cies, and this evidence suggests a circadian influence on Egr1 expression as well [23]. In rodents, zif268 mRNA is increased in suprachiasmatic nuclei (SCN), the circadian pacemaker, by retinal illumination at night [51]. The SCN clock is reset by daily exposure to light by transmission of photic information from retina via the retinohypothalamic tract, a monosynaptic glutamatergic projection [52]. The photosensitive retinal gan-glion cells innervating SCN express melanopsin and might be the primary photoreceptor for this system [53]. However, there are pronounced species differences in such zif268 response as well as melanopsin expression [51,54]. In zebrafish, the melanopsin mRNA is confined to the eye, merely in a subset of retinal horizontal cells (type B) [54]. In contrast, chicken melanopsin is expressed in brain, pineal gland, and all retinal layers [55]. ZENK gene in chick retina is downregulated in postnatal myopic chicks with elongated eyeballs [21,22], but the causal relationship between ZENK gene expression and ocular growth has not been well established. Except ZENK studies, no other research documents the association between Egr1 expression and ocular growth.
Egr1 expression is also upregulated in response to diverse stimuli including basic fibroblast growth factor (bFGF), focal ischemia, hypoxia, physical forces, injury, etc. [56]. It is acti-vated through multiple protein-protein interactions with co-activators such as CREB-binding protein (CBP) and p300 [56,57]. The expression of numerous target genes like plate-let-derived growth factor (PDGF) A-chain, PDGF B-chain, transforming growth factor-β1, and tumor necrosis factor is
regulated by Egr1 protein [46]. Similarly, microphthalmia-associated transcription factor (Mitf) is also responsive to bFGF, activated by CBP/p300, and associated with cellular differentiation as well as cell division [58]. The mutation of Mitf in mice results in retinal degeneration and microphthalmia [59]. The Mitf/Pax6 interactions are important for RPE devel-opment [58]. Our study proves the effect of Egr1 expression on embryonic oculogenesis. Egr1 and Mitf may share some common downstream mechanisms in oculogenesis, although the latter mainly affects RPE and is critical for RPE-neural retina communication [59]. Leung et al. proposed that cyclin-dependent kinase 5 (Cdk5)/p35 activity was essential for eye development and retinal lamination in zebrafish and was likely regulated by Egr1 [60].
In conclusion, early knockdown of zebrafish embryonic Egr1 gene arrested retinal and lenticular development and then caused microphthalmos. This is quite different from the find-ings in postnatal chicks and implies Egr1 may have diverse functions at various developmental stages in varied species. MO is a powerful tool to study gene function precisely and quickly. However, since the outcomes result from gene knock-down in all tissues, further research to identify the role of Egr1 in the isolated tissue is favored.
ACKNOWLEDGEMENTS
The whole study was supported by the National Science Coun-cil Grant 92-2314-B-418-006, Taiwan. We thank the Zebrafish
International Resource Center (NIH-NCRR Grant number RR12546) for providing zpr-1 antibody.
REFERENCES
1. Thisse C, Zon LI. Organogenesis—heart and blood formation from the zebrafish point of view. Science 2002; 295:457-62. 2. Wawersik S, Maas RL. Vertebrate eye development as modeled in
Drosophila. Hum Mol Genet 2000; 9:917-25.
3. Malicki JJ, Pujic Z, Thisse C, Thisse B, Wei X. Forward and re-verse genetic approaches to the analysis of eye development in zebrafish. Vision Res 2002; 42:527-33.
4. Cheng KC, Levenson R, Robishaw JD. Functional genomic dis-section of multimeric protein families in zebrafish. Dev Dyn 2003; 228:555-67.
5. Fadool JM. Understanding retinal cell fate determination through genetic manipulations. Prog Brain Res 2001; 131:541-54. 6. Bilotta J, Saszik S. The zebrafish as a model visual system. Int J
Dev Neurosci 2001; 19:621-9.
7. Glass AS, Dahm R. The zebrafish as a model organism for eye development. Ophthalmic Res 2004; 36:4-24.
8. Penberthy WT, Shafizadeh E, Lin S. The zebrafish as a model for human disease. Front Biosci 2002; 7:d1439-53.
9. Pujic Z, Malicki J. Retinal pattern and the genetic basis of its for-mation in zebrafish. Semin Cell Dev Biol 2004; 15:105-14. 10. Schmitt EA, Dowling JE. Early retinal development in the
zebrafish, Danio rerio: light and electron microscopic analyses. J Comp Neurol 1999; 404:515-36.
11. Easter SS Jr, Nicola GN. The development of vision in the zebrafish (Danio rerio). Dev Biol 1996; 180:646-63.
12. Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of embryonic development of the zebrafish. Dev Dyn 1995; 203:253-310.
13. Heasman J. Morpholino oligos: making sense of antisense? Dev Biol 2002; 243:209-14.
14. Ekker SC, Larson JD. Morphant technology in model develop-mental systems. Genesis 2001; 30:89-93.
15. Udvadia AJ, Linney E. Windows into development: historic, cur-rent, and future perspectives on transgenic zebrafish. Dev Biol 2003; 256:1-17.
16. Corey DR, Abrams JM. Morpholino antisense oligonucleotides: tools for investigating vertebrate development. Genome Biol 2001; 2:REVIEWS1015.
17. Long KD, Salbaum JM. Evolutionary conservation of the imme-diate-early gene ZENK. Mol Biol Evol 1998; 15:284-92. 18. Drummond IA, Rohwer-Nutter P, Sukhatme VP. The zebrafish
egr1 gene encodes a highly conserved, zinc-finger transcrip-tional regulator. DNA Cell Biol 1994; 13:1047-55.
19. Close R, Toro S, Martial JA, Muller M. Expression of the zinc finger Egr1 gene during zebrafish embryonic development. Mech Dev 2002; 118:269-72.
20. Carman JA, Monroe JG. The EGR1 protein contains a discrete transcriptional regulatory domain whose deletion results in a truncated protein that blocks EGR1-induced transcription. DNA Cell Biol 1995; 14:581-9.
21. Bitzer M, Schaeffel F. Defocus-induced changes in ZENK ex-pression in the chicken retina. Invest Ophthalmol Vis Sci 2002; 43:246-52.
22. Fischer AJ, McGuire JJ, Schaeffel F, Stell WK. Light- and focus-dependent expression of the transcription factor ZENK in the chick retina. Nat Neurosci 1999; 2:706-12.
23. Brand C, Burkhardt E, Schaeffel F, Choi JW, Feldkaemper MP. Regulation of Egr-1, VIP, and Shh mRNA and Egr-1 protein in 1257
the mouse retina by light and image quality. Mol Vis 2005; 11:309-20.
24. Lawrence MS, Azar DT. Myopia and models and mechanisms of refractive error control. Ophthalmol Clin North Am 2002; 15:127-33.
25. Pavletich NP, Pabo CO. Zinc finger-DNA recognition: crystal structure of a Zif268-DNA complex at 2.1 A. Science 1991; 252:809-17.
26. Li Z, Joseph NM, Easter SS Jr. The morphogenesis of the zebrafish eye, including a fate map of the optic vesicle. Dev Dyn 2000; 218:175-88.
27. Chen YH, Tsai HJ. Treatment with Myf5-morpholino results in somite patterning and brain formation defects in zebrafish. Dif-ferentiation 2002; 70:447-56.
28. Nasevicius A, Ekker SC. Effective targeted gene ‘knockdown’ in zebrafish. Nat Genet 2000; 26:216-20.
29. Vihtelic TS, Hyde DR. Zebrafish mutagenesis yields eye mor-phological mutants with retinal and lens defects. Vision Res 2002; 42:535-40.
30. Yazulla S, Studholme KM. Neurochemical anatomy of the zebrafish retina as determined by immunocytochemistry. J Neurocytol 2001; 30:551-92.
31. Devine CA, Key B. Identifying axon guidance defects in the embryonic zebrafish brain. Methods Cell Sci 2003; 25:33-7. 32. van der Sar AM, Zivkovie1/2 D, den Hertog J. Eye defects in
receptor protein-tyrosine phosphatase alpha knock-down zebrafish. Dev Dyn 2002; 223:292-7.
33. Larison KD, Bremiller R. Early onset of phenotype and cell pat-terning in the embryonic zebrafish retina. Development 1990; 109:567-76.
34. Simon P, Schott K, Williams RW, Schaeffel F. Posttranscriptional regulation of the immediate-early gene EGR1 by light in the mouse retina. Eur J Neurosci 2004; 20:3371-7.
35. Pichler FB, Laurenson S, Williams LC, Dodd A, Copp BR, Love DR. Chemical discovery and global gene expression analysis in zebrafish. Nat Biotechnol 2003; 21:879-83.
36. Ekker SC. Morphants: a new systematic vertebrate functional genomics approach. Yeast 2000; 17:302-306.
37. Mataga N, Fujishima S, Condie BG, Hensch TK. Experience-dependent plasticity of mouse visual cortex in the absence of the neuronal activity-dependent marker egr1/zif268. J Neurosci 2001; 21:9724-32.
38. Lee SL, Sadovsky Y, Swirnoff AH, Polish JA, Goda P, Gavrilina G, Milbrandt J. Luteinizing hormone deficiency and female in-fertility in mice lacking the transcription factor NGFI-A (Egr-1). Science 1996; 273:1219-21.
39. Zhu Z, Zheng T, Lee CG, Homer RJ, Elias JA. Tetracycline-con-trolled transcriptional regulation systems: advances and appli-cation in transgenic animal modeling. Semin Cell Dev Biol 2002; 13:121-8.
40. Huang CJ, Jou TS, Ho YL, Lee WH, Jeng YT, Hsieh FJ, Tsai HJ. Conditional expression of a myocardium-specific transgene in zebrafish transgenic lines. Dev Dyn 2005; 233:1294-303. 41. Hjorth JT, Connor RM, Key B. Role of hlx1 in zebrafish brain
morphogenesis. Int J Dev Biol 2002; 46:583-96.
42. Oates AC, Bruce AE, Ho RK. Too much interference: injection of double-stranded RNA has nonspecific effects in the zebrafish embryo. Dev Biol 2000; 224:20-8.
43. Wargelius A, Ellingsen S, Fjose A. Double-stranded RNA in-duces specific developmental defects in zebrafish embryos. Biochem Biophys Res Commun 1999; 263:156-61.
44. Jain KK. RNAi and siRNA in target validation. Drug Discov Today 2004; 9:307-9.
45. Knapska E, Kaczmarek L. A gene for neuronal plasticity in the mammalian brain: Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK? Prog Neurobiol 2004; 74:183-211.
46. Beckmann AM, Wilce PA. Egr transcription factors in the ner-vous system. Neurochem Int 1997; 31:477-510; discussion 517-6.
47. Pinaud R, De Weerd P, Currie RW, Fiorani Jr M, Hess FF, Tremere LA. Ngfi-a immunoreactivity in the primate retina: implications for genetic regulation of plasticity. Int J Neurosci 2003; 113:1275-85.
48. Zhong X, Ge J, Smith EL 3rd, Stell WK. Image defocus modu-lates activity of bipolar and amacrine cells in macaque retina. Invest Ophthalmol Vis Sci 2004; 45:2065-74.
49. Gudehithlu KP, Neff NH, Hadjiconstantinou M. c-fos and NGFI-A mRNNGFI-A of rat retina: evidence for light-induced augmentation and a role for cholinergic and glutamate receptors. Brain Res 1993; 631:77-82.
50. Pinaud R, Tremere LA, Penner MR, Hess FF, Barnes S, Robertson HA, Currie RW. Plasticity-driven gene expression in the rat retina. Brain Res Mol Brain Res 2002; 98:93-101.
51. Dong YN, Goguen D, Robertson HA, Rusak B. Anatomical and temporal differences in the regulation of ZIF268 (NGFI-A) pro-tein in the hamster and mouse suprachiasmatic nucleus. Neuro-science 2002; 111:567-74.
52. Coogan AN, Piggins HD. Circadian and photic regulation of phos-phorylation of ERK1/2 and Elk-1 in the suprachiasmatic nuclei of the Syrian hamster. J Neurosci 2003; 23:3085-93.
53. Hattar S, Liao HW, Takao M, Berson DM, Yau KW. Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity. Science 2002; 295:1065-70. 54. Bellingham J, Whitmore D, Philp AR, Wells DJ, Foster RG.
Zebrafish melanopsin: isolation, tissue localisation and phylo-genetic position. Brain Res Mol Brain Res 2002; 107:128-36. 55. Chaurasia SS, Rollag MD, Jiang G, Hayes WP, Haque R, Natesan
A, Zatz M, Tosini G, Liu C, Korf HW, Iuvone PM, Provencio I. Molecular cloning, localization and circadian expression of chicken melanopsin (Opn4): differential regulation of expres-sion in pineal and retinal cell types. J Neurochem 2005; 92:158-70.
56. Silverman ES, Collins T. Pathways of Egr-1-mediated gene tran-scription in vascular biology. Am J Pathol 1999; 154:665-70. 57. Khachigian LM, Collins T. Early growth response factor 1: a
pleiotropic mediator of inducible gene expression. J Mol Med 1998; 76:613-6.
58. Planque N, Leconte L, Coquelle FM, Martin P, Saule S. Specific Pax-6/microphthalmia transcription factor interactions involve their DNA-binding domains and inhibit transcriptional proper-ties of both proteins. J Biol Chem 2001; 276:29330-7. 59. Moore KJ. Insight into the microphthalmia gene. Trends Genet
1995; 11:442-8.
60. Leung YF, Link BA, Dowling JE. Cdk5/p35 activity is essential for eye growth and retinal lamination in zebrafish. ARVO An-nual Meeting; 2005 May 1-5; Fort Lauderdale (FL).
1258
The print version of this article was created on 26 Oct 2006. This reflects all typographical corrections and errata to the article through that date. Details of any changes may be found in the online version of the article. α