The COMT L allele Modifies the Association between MAOB Polymorphism and PD in Taiwanese
R.M. Wu, MD, PhD; C.W. Cheng, PhD; K.H. Chen, PhD; S.L. Lu, MSc;
D.E. Shan, MD, PhD; Y.F. Ho, MD, PhD; and H.D. Chern, MD, PhD;
From the Department of Neurology, College of Medicine, National Taiwan University and National Taiwan University Hospital, Taipei, Taiwan (Dr. Wu), the Graduate Institute of Pharmaceutical Sciences, College of Medicine, National Taiwan University, Taipei, Taiwan
(Drs. Cheng, Chern, Ho, and Lu), the Institute of Epidemiology, College of Public Health, National Taiwan University, Taipei, Taiwan (Dr. Chen), and the Neurological Institute, Taipei
Veterans General Hospital & National Yang-Ming University School of Medicine, Taipei, Taiwan (Dr. Shan).
Address for correspondence and reprint requests: Dr. R.M. Wu, Department of Neurology, National Taiwan University Hospital, No 7, Chung-Shan South Road, Taipei, 100, Taiwan. Fax: +886-2-2341-8395, E-mail: [email protected]
Key wor ds: Parkinson's disease, monoamine oxidase B, catechol-O-methyltransferase, genetic
polymorphism.
Acknowledgement
This study was supported by research grant NSC 89-2314-B-002-143 from the National Science Council (Taipei, Taiwan, R.O.C.). The authors would like to thank Dr. T Barkas for revision of the English.
Ar ticle abstr act
Objective: Reports suggest that COMTL/L (Val158
/Met) and MAOB intron 13 genotype
polymorphism is associated with Parkinson’s disease (PD). To understand the
ethnicity-specific effects of genetic polymorphism, we performed a case-control study of the association
between PD susceptibility and polymorphism of MAOB and COMT, both separately and in
combination, in Taiwanese. Methods: 224 PD patients and 197 controls, matched for age,
gender, and birthplace, were recruited. MAOB and COMT polymorphism genotyping was
performed using PCR-based RFLP analyses. Chi-square, odds ratio, and Fisher’s exact tests
were used to compare differences in allelic frequencies and genotypes. Results: The MAOB G
genotype (G in men and G/G in women) was associated with a 2.07-fold increased relative risk
of PD. COMT polymorphism, considered alone, showed no correlation with PD risk; however,
a significant synergistic enhancement was found in PD patients harboring both the COMTL and
MAOB G genotypes. Conclusions: These results suggest that, in Taiwanese, PD risk is
associated with MAOB G intron 13 polymorphism and this association is augmented in the
presence of the COMTL genotype, indicating an interaction of these two dopamine
metabolizing enzymes in the pathogenesis of sporadic PD. However, the relatively low
frequencies of these combined genotypes in our study necessitates confirmation with a larger
Introduction
Since PD is characterized by the loss of at least 60% of midbrain dopaminergic neurons1
, it
has been suggested that genetic variants of the enzymes involved in the biosynthesis and
degradation of dopamine and related compounds influence susceptibility to development of this
disease. These enzymes include monoamine oxidase B (MAOB) and catechol-
O-methyltransferase (COMT). MAOB is potentially relevant to PD because of its role in
catabolizing dopamine, with the resultant production of reactive oxidative free radicals,2,3
and
in activating exogenous neurotoxins, such as 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
(MPTP), a neurotoxin that induces parkinsonism in intravenous users of synthetic heroin and in
nonhuman primates.4-6
Inhibition of MAOB activity by L-deprenyl prevents the development
of parkinsonism in nonhuman primates by blocking the conversion of MPTP to MPP+
,7
and
possibly reduces the rate of PD progression in humans.8
PD patients are reported to have higher
platelet MAOB activity than control individuals.9
Furthermore, inhibition of MAOB activity in
the brains of cigarette smokers has been suggested to have a protective effect against the
development of PD.10,11
Thus, there is an increasing body of evidence that MAOB may play a
crucial role in the pathogenesis of PD.
The gene encoding MAOB is located on the X chromosome. It contains a single-stranded
conformational polymorphism in intron 13, a transitional conversion of adenine (A) to guanine
(G) at a position 36 base pairs (bp) upstream from the 5’end of exon 14.12
studies have been carried out on a possible association between the G-A polymorphism and PD,
the findings have been inconsistent. Both the A12
and G13
alleles have been reported to be
associated with a risk of PD in Caucasians, while two studies failed to find any correlation in
Caucasian populations.14,15 In Asian populations, no correlation was found between PD
occurrence and MAOB G-A polymorphism in either Japanese16
or a small Taiwanese
population.17
COMT polymorphism has also been studied in terms of an association with PD risk.
COMT is a ubiquitous enzyme that catalyzes the O-methylation of biologically active or toxic
catechols, and plays a major role in the metabolism of drugs and neurotransmitters, such as
L-dopa, dopamine, noradrenaline, and adrenaline. A single gene, located on chromosome 22q11,
encodes both the acid-soluble and membrane-bound forms of this enzyme. A G to A transition
at codon 158 of the COMT gene, resulting in the substitution of methionine for valine, is linked
to low COMT enzyme activity and is designated the L (low activity) allele, in contrast to the H
(high activity) allele.18 In individuals with the L allele, the COMT protein is thermolabile.
Differences in COMT activity may determine individual variations in the therapeutic response
to levodopa19 and affect the individual’s susceptibility to PD. Studies suggest that COMTL is
less frequent in Asians than in Caucasians.20,21 Homozygosity for the COMT L allele has been
reported to be a genetic risk for PD in Japanese20, but recent studies in Caucasians and Chinese
The prevalence of PD varies world-wide. In general, this disease is less common in Asia
than in Western countries.24 Thus, understanding the ethnicity-specific effects of susceptibility
genes on PD risk in different races may provide valuable clues to potential causes of racial and
individual susceptibility to PD. Although several studies have investigated a possible
association of either MAOB or COMT polymorphism with PD susceptibility in Asian
populations,16,17, 20,23,25 the results have been inconsistent, possibly due to differences in ethnicity,
sample size, and the matched controls used. Moreover, all previous studies have focused on a
single gene effect of either MAOB or COMT. In the present study, we determined both the
individual and combined effects of MAOB and COMT genetic polymorphisms on PD risk in a
large population of Taiwanese PD patients (n=224) and in age-, sex-, and origin
(birthplace)-matched control subjects (n=197).
Mater ials and Methods Human subjects
After obtaining their informed consent, 224 patients with idiopathic PD (162 men and 62
women; average age: 67.2 ± 9.1 years) were recruited at the Movement Disorder Clinics of the
National Taiwan University Hospital, Taipei and the Taipei Veterans General Hospital. All
patients met the criteria for PD, which included the presence of two of the three cardinal signs
(tremor at rest, bradykinesia, and rigidity), improvement of symptoms with L-dopa therapy, and
or toxins, or of atypical parkinsonism. The control group consisted of 197 individuals (145 men
and 52 women; average age: 65.8 ± 9.2 years) recruited from the same two hospitals and from
community groups in the Taipei area, chosen on the basis that neither they nor their blood
relatives showed evidence of any neurological and psychiatric disorders. Controls and cases
were matched for age, sex, and birthplace (Table 1).
Labor ator y analysis (i) DNA extr action
Samples of buccal mucosa cells were collected from each PD patient or control subject by
rolling a buccal brush along the inner surface of the cheek. The brush was dried for 10-15
minutes at room temperature, then immediately stored at 4ºC until extraction of the genomic
DNA, performed using a QIAamp DNA Mini kit (QIAGEN Inc., Chatsworth, CA, U.S.A.). The
purified DNA was stored at -20ºC for genotype analysis.
(ii) Identification of MAOB intron 13 and COMT polymor phisms
PCR-based RFLP analyses were used to determine the MAOB and COMT genotypes of
the subjects.
In the MAOB study, a 232 bp DNA fragment of the MAOB gene containing the intron 13
polymorphism was amplified. The forward primer 5'-GGAACCTCTTATACCACAGG-3' and
reverse primer 5'-GACTGCCAGATTTCATCCTC-3' were used for partial MAOB DNA
fragment amplification.12
ng of genomic DNA as a template, 3 mM MgCl2, and 1 unit of Ampli Taq DNA polymerase
(Perkin Elmer Cetus, Norwalk, CT). The PCR procedures involved an initial denaturation at
94oC for 4 min, 34 cycles of 94oC for 40 sec, 54oC for 30 sec, and 72oC for 40 sec, and a final
extension at 72o
C for 10 min. To determine the MAOB polymorphism, 10 µL aliquots of the
PCR-amplified DNA product were digested with restriction enzyme Tsp45I and the digested
DNA pattern analyzed by electrophoresis on a 2% SeaKem LE agarose gel (FMC Crop.,
Rockland, ME). MAOB allele 1 (containing A and therefore the Tsp45I restriction site) was
detected as two bands of 146 and 86 bp, while allele 2 (containing G, no Tsp45I restriction site)
was detected as a single 232 bp band (Figure 1).
In the COMT study, the PCR mixture contained the same reagents used in MAOB
amplification, except that the forward primer was 5'-TCGTGGACGCCGTGATTCAGG-3' and
the reverse primer 5'-AGGTCTGACAACGGGTCAGGC-3'.25 The reaction mixture was
subjected to an initial denaturation at 94ºC for 4 min, followed by 34 cycles of 94ºC for 40 sec,
55ºC for 30 sec, and 72ºC for 30 sec, and a final extension at 72ºC for 10 min. Ten microliters
of the amplified double-stranded DNA was digested with restriction endonuclease NlaIII, then
the sample was subjected to electrophoresis. The COMTH allele was detected by the presence
of two bands of 136 and 81 bp and the COMTL allele by the presence of three bands of 96, 81,
Statistical analysis
In most cases, chi-square and odds ratio tests were used to compare either allelic
frequencies or the genotype frequencies of MAOB or COMT in PD patients and control
subjects. When more than 20% of the cell numbers that the expected number of cases was less
than 5.0 in a cell, or when the expected number of cases was less than 1.0 in a cell, Fisher’s
exact test was performed.26
Since the MAOB gene is located on the X chromosome, results for
the MAOB genotype were assessed separately in men and women. To comprehensively assess
the individual contributions of MAOB and COMT polymorphism to an association with PD
risk, we further examined the relationship of the individual susceptibility genotypes (the
MAOB G allele and the COMT L allele) with PD risk, stratified by age (≤ 60-years-old vs.
>60-years-old) or birthplace (Taiwan vs. mainland China).
Because of the possibility of a potential interaction of the two dopamine-metabolizing
genes in the development of PD, we then investigated PD risk associated with combinations of
these two susceptibility genotypes. All statistical tests were based on a two-tailed probability
and values of p<0.05 were considered significant.
Results
MAOB G-A polymor phism in intron 13
Table 2 shows the allelic frequencies and genotype distribution for MAOB polymorphism in
PD patients and controls. There was no significant difference in total A and G MAOB allelic
frequencies between PD patients (81.6% and 18.4%, A:G ratio = 4.42) and controls (86.4% and
13.6%, A:G ratio = 6.33). However, MAOB genotype distribution showed a significant
difference between the two groups, the frequency of the G genotype (G in men and G/G in
women) being twice as high in PD patients compared to controls (OR = 2.07, 95% CI 1.12 to
3.81, p = 0.018 for χ2
test). This association showed a significant gender difference. In men,
the association between the MAOB G genotype and an increased PD risk was slightly stronger
(OR = 2.14, 95% CI 1.14 to 4.00, p=0.016 for χ2
test) than in all subjects combined, and a
significantly reduced risk of PD was found in subjects with the A allele (OR = 0.47, 95% CI
0.25 to 0.88), while, in women, there was no significant difference in the distribution of MAOB
genotypes between PD patients and controls. Compared with previous data for a Caucasian
population13, the frequency of the homozygous variant G in women was much lower in
Taiwanese (17.2% vs.1.6%).
Table 3 shows the results of analyses of the effects of age and birthplace on the association
of the susceptible MAOB G genotype with PD risk. A higher frequency of the MAOB G
genotype was seen in PD patients in both age groups (p values of 0.056 for subjects ≤ 60 years
and of 0.053 for subjects >60 years). Interestingly, the frequency of the G genotype was higher
Chinese group, the frequency was 25.2% in PD patients, 13.8% in controls, and 20.0% in all
subjects combined, while, in patients born in Taiwan, the corresponding frequencies were 9.6%,
4.8%, and 7.3%, a 2.6- to 2.9-fold difference. Statistically, an association between MAOB
genotype and PD risk was seen in subjects born in mainland China (χ2
= 3.86, p=0.049), but not
in those born in Taiwan (χ2
= 1.87, p = 0.171).
COMT polymor phism at codon 158
COMT genotype data were available for 222 patients (99.1%) and 191 controls (97.0 %).
Table 4 shows the allelic and genotypic frequencies for COMT polymorphism in PD patients
and controls. The allelic frequency of COMTL (Met) was 25.9% in PD patients, and 22.5% in
control subjects, this difference not being significantly different. There was also no significant
difference between the two groups in the frequencies of either the homozygous (H/H or L/L) or
heterozygous (H/L) genotypes. Subgroup analyses for gender, age, and birthplace also showed
no association between the susceptible COMT genotype (COMTL) and PD risk (Table 5).
Combined effect of the two susceptibility genes
Combined MAOB and COMT genotyping data were available for 218 PD patients (97.3%)
and 185 controls (94.0%). The distribution of the joint allele frequencies for these two loci is
shown in Table 6. Importantly, the frequency of the combined alleles MAOB A and COMT H
frequency of the combined alleles MAOB G and COMT L was 2.4 times higher in PD patients
than in controls (OR = 2.39, 95% CI 1.11-5.18, p = 0.023). Moreover, these associations
between PD and the combined MAOB and COMT alleles were stronger in men than in the total
population, but were not seen in women. In men, the combined alleles MAOB A and COMT H
were associated with a reduced relative risk of PD (OR 0.65, 95% CI 0.46-0.91, p = 0.013),
whereas the combined alleles MAOB G and COMT L markedly increased the relative risk of
PD by 7.24-fold (95% CI 1.65-31.80, p = 0.002). These findings suggest that the combination
of the MAOB A and COMT H alleles may be a protective factor for PD development, while the
combination of the uncommon MAOB G and COMT L alleles may increase the relative risk of
PD.
Table 7 shows the combined effect of these two susceptibility genotypes on the
development of PD. For MAOB polymorphism, the AA and A/G genotypes were combined
because the two genotypes were found to have a similar effect, relative to GG (Table 2). For
COMT polymorphism, because no association of the genotype with PD was found in this study
(Table 4), the heterozygous genotype (H/L) was combined with either the homozygous HH or
LL genotypes. Consistently, the presence of the MAOB G genotype (G for men and G/G for
women) and of at least one copy of the COMT L allele significantly increased the relative risk
of PD by a factor of 5.0-6.0, suggesting a strong synergistic effect of the low activity COMT L
frequency of the combined genotypes of MAOB A, A/A, and A/G with COMT H/H and H/L
was lower in PD patients than in controls (p = 0.02), implying a protective effect of the
combined wild and heterozygous genotypes of these two dopamine metabolizing enzymes on
the occurrence of PD (OR = 0.55).
Discussion
The frequency of the MAOB G-A polymorphism in intron 13 differs in the ethnic groups
that have been studied. In Caucasians, the frequency of intron 13 allele 1 (A) of the MAOB
gene is reported to be 45-51% in PD patients and 49-58% in controls.12-15 Consistent with other
studies in Asians,16,17 our present results showed a predominant distribution of the A allele (A/G
ratio = 4.43) in control subjects, and no association between PD and MAOB polymorphism in
terms of the total A and G allele frequencies. However, when we assessed the gender
distribution of the MAOB genotypes, we found an increased frequency of the MAOB G variant
both in men and in the whole group, a finding not previously reported in studies on Asians.
Moreover, an inverse relationship between PD and A polymorphism was seen in men, with a
reduced relative risk of 0.47(95% CI 0.25,0.88). The sample size in the current study was
substantially greater than those in previous studies involving Japanese16 and Taiwanese,17 thus
providing a greater power to detect a significant difference in the distribution of genotype
frequencies between PD and control subjects. The prevalence rate of PD is 2- to 4-fold higher
development, a lower frequency of this genotype would be expected in Asians than in
Caucasians. Moreover, the MAOB gene is on the X-chromosome, suggesting that any effect of
MAOB on PD risk should be higher in men than in women. Importantly, our findings are
consistent with both of these propositions. Although the G-A polymorphism occurs in an intron
and, therefore, does not directly alter the amino acid sequence of the enzyme, linkage
disequilibrium of this polymorphism with other genes that confer PD susceptibility might
explain this association.13,27 PD patients are reported to have higher MAOB activity than
controls.9 The positive association between the MAOB G polymorphism and PD seen in this
study justifies further investigations to determine whether this polymorphism affects brain
MAOB activity, since high MAOB activity may increase oxidative stress and PD risk.3
The Taiwanese population is ethnically diverse. The majority of the population is Han
Chinese and the rest aboriginal. The Taiwan aborigines consist of at least 11-13 linguistically
distinct populations and those groups settled in the plains or valleys are known collectively as
Peen-Poof.28 Several centuries ago, the Han Chinese began to migrate from mainland China to
Taiwan. In the early migration periods, they settled in the south of Taiwan. Most were single
men who married Peen-Poof women, resulting in an ethnic admixture.29 This migration steadily
continued for several centuries. However, a recent large migration of Han Chinese to Taiwan
occurred approximately 50 years ago after the Second World War; this group of Chinese
and previous Han immigrants. Interestingly, we found a significant difference in the frequency
distribution of MAOB polymorphism and its association with PD in the two subgroups of
populations born in mainland China or Taiwan. This finding is compatible with the ethnic
diversity of Taiwanese resulting from the history of migration. Thus, any genetic association
study of PD in Taiwanese should be conducted using patient and control populations in which
ethnicity is closely matched.
The level of COMT enzyme activity is genetically polymorphic. Some ethnic differences
have been recognized and may explain variations in the individual response to L-dopa
therapy.19 The low COMT activity allele (COMTL) is common in Caucasians, with an allelic
frequency of 40-50%21,22, but less common in Asians in whom the frequency is
20-30%.20,21,23,25,30 In agreement with the results of another population study in Taiwan,30 we found
a low COMTL allele frequency in our subjects; in controls, the frequency of the L allele was
22.5% and the frequency of the L/L homozygote 6.3%. As in a previous study in Hong-Kong
Chinese,23, no correlation was found in the present study between COMT 158Met/Val
polymorphism and PD.
In this study, the most noteworthy finding was that individuals harboring both the MAOB
G genotype (G for men and G/G for women) and the COMTL
allele had a much higher risk of
PD than those harboring only the MAOB G genotype. This synergistic effect of the low activity
these two enzyme polymorphisms may be one of the underlying mechanisms of PD etiology. In
humans, both COMT and MAOB metabolize dopamine and other catecholamines. Low
COMT enzyme activity results in dopamine being metabolized mainly by MAO, which may
increase oxidative stress on midbrain dopamine neurons as a result of free radical formation3,31
and thus trigger or enhance the development of PD in susceptible individuals. Accordingly, the
MAOB G genotype may be a major susceptible risk factor and COMTL
may facilitate the
development of PD in Taiwanese. However, because of the relatively low frequencies of the
combined MAOB G and COMT L alleles in this study, further confirmation with a larger
sample size is necessary.
A full understanding of the pathogenesis of PD requires extensive investigation to evaluate
whether a cluster of related genes is involved in the nigrostriatal degeneration of dopamine
neurons. To date, most studies have focused on single genetic risk susceptibility to PD.
However, such isolated single gene studies may overlook the complexity of dopamine
metabolism, which involves sequential biochemical processes, and the effect of multiple genes
on PD development. As far as we are aware, our study is the first to address the issue of
dopamine catabolism in relation to PD risk using a two-gene model. This approach should
allow more precise evaluation of PD risk associated with MAOB and COMT polymorphisms
and could also be used to assess associations between PD and other candidate genes, such as
Tables
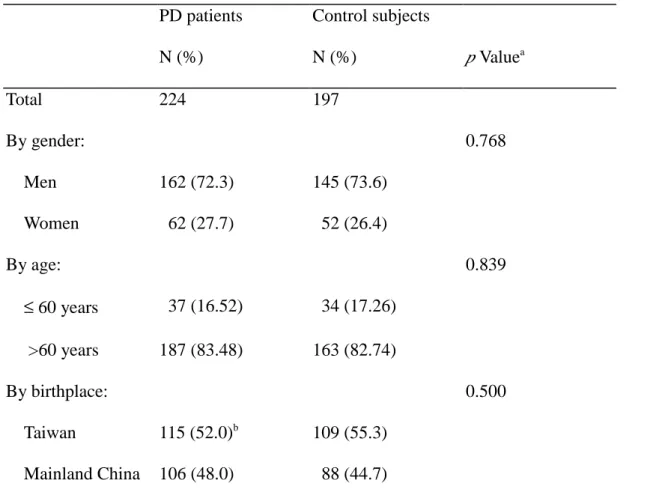
Table 1 Demographic data for PD patients and control subjects PD patients N (%) Control subjects N (%) p Valuea Total 224 197 By gender: 0.768 Men 162 (72.3) 145 (73.6) Women 62 (27.7) 52 (26.4) By age: 0.839 ≤ 60 years 37 (16.52) 34 (17.26) >60 years 187 (83.48) 163 (82.74) By birthplace: 0.500 Taiwan 115 (52.0)b 109 (55.3) Mainland China 106 (48.0) 88 (44.7) aχ2 test. b
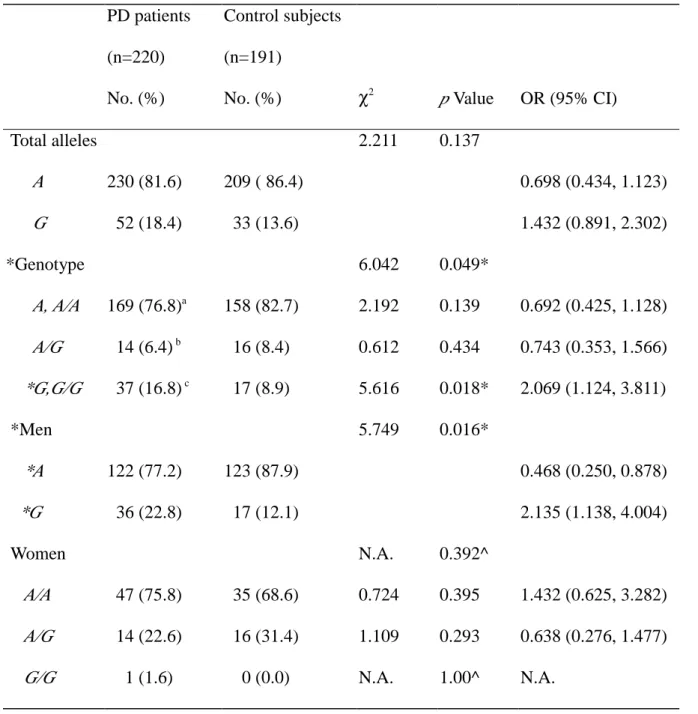
Table 2 Distribution of MAOB allele frequency and genotype polymorphism, and estimated OR in relation to PD risk PD patients (n=220) No. (%) Control subjects (n=191)
No. (%) χ2 p Value OR (95% CI)
Total alleles 2.211 0.137 A 230 (81.6) 209 ( 86.4) 0.698 (0.434, 1.123) G 52 (18.4) 33 (13.6) 1.432 (0.891, 2.302) *Genotype 6.042 0.049* A, A/A 169 (76.8)a 158 (82.7) 2.192 0.139 0.692 (0.425, 1.128) A/G 14 (6.4) b 16 (8.4) 0.612 0.434 0.743 (0.353, 1.566) *G,G/G 37 (16.8) c 17 (8.9) 5.616 0.018* 2.069 (1.124, 3.811) *Men 5.749 0.016* *A 122 (77.2) 123 (87.9) 0.468 (0.250, 0.878) *G 36 (22.8) 17 (12.1) 2.135 (1.138, 4.004) Women N.A. 0.392^ A/A 47 (75.8) 35 (68.6) 0.724 0.395 1.432 (0.625, 3.282) A/G 14 (22.6) 16 (31.4) 1.109 0.293 0.638 (0.276, 1.477) G/G 1 (1.6) 0 (0.0) N.A. 1.00^ N.A.
* : Significant difference between the two groups.
^ : Fisher’s exact test.
N.A. : not applicable.
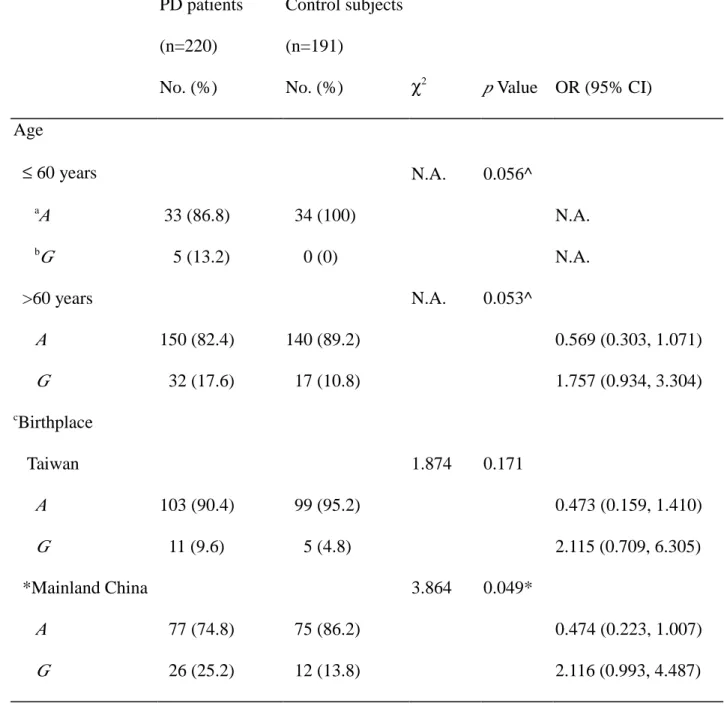
Table 3 Odds ratio for MAOB genotype polymorphism in relation to PD risk stratified by age or birthplace PD patients (n=220) No. (%) Control subjects (n=191)
No. (%) χ2 p Value OR (95% CI)
Age ≤ 60 years N.A. 0.056^ a A 33 (86.8) 34 (100) N.A. b G 5 (13.2) 0 (0) N.A. >60 years N.A. 0.053^ A 150 (82.4) 140 (89.2) 0.569 (0.303, 1.071) G 32 (17.6) 17 (10.8) 1.757 (0.934, 3.304) c Birthplace Taiwan 1.874 0.171 A 103 (90.4) 99 (95.2) 0.473 (0.159, 1.410) G 11 (9.6) 5 (4.8) 2.115 (0.709, 6.305) *Mainland China 3.864 0.049* A 77 (74.8) 75 (86.2) 0.474 (0.223, 1.007) G 26 (25.2) 12 (13.8) 2.116 (0.993, 4.487)
* : Significant difference between groups. ^ : Fisher’s exact test. N.A. : not applicable.
a
A corresponds to genotypes A/A and A/G b
Table 4 COMT allele and genotype frequencies in PD patients and control subjects PD patients (n=222) No. (%) Control subjects (n=191)
No. (%) χ2 p Value OR (95% CI)
Total alleles 1.280 0.258 H(Val) 329 (74.1) 296 (77.5) 0.831 (0.603, 1.145) L(Met) 115 (25.9) 86 (22.5) 1.203 (0.873, 1.658) Genotype 1.194 0.550 H/H 125 (56.3) 117 (61.2) 1.037 0.309 0.815 (0.550, 1.208) H/L 79 (35.6) 62 (32.5) 0.446 0.504 1.149 (0.764, 1.730) L/L 18 (8.1) 12 (6.3) 0.508 0.476 1.316 (0.617, 2.807)
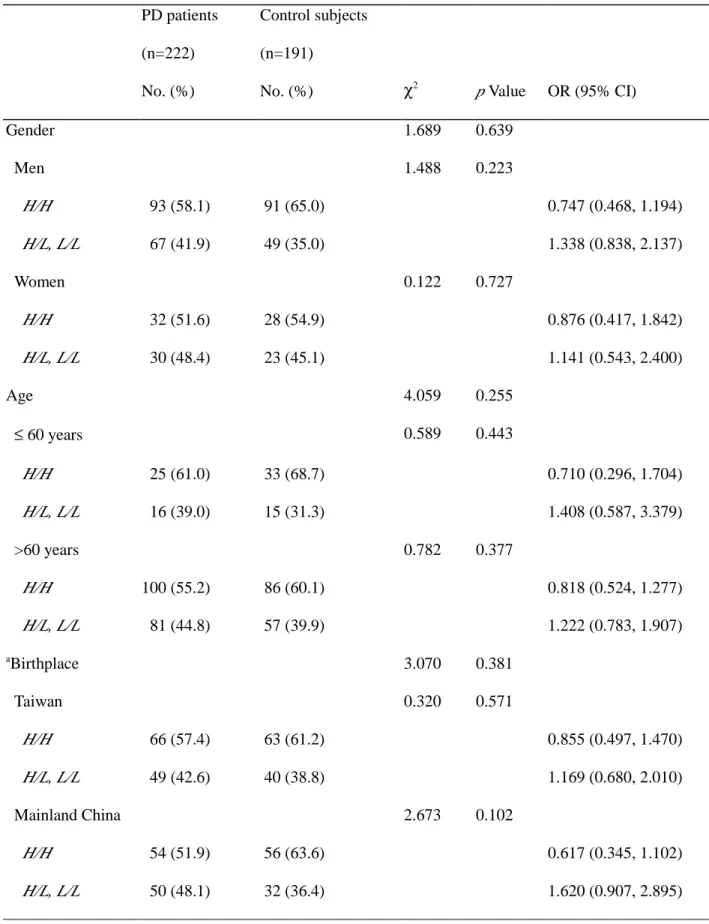
Table 5 Odds ratio for COMT genotype polymorphism in relation to PD risk stratified by age or birthplace PD patients (n=222) No. (%) Control subjects (n=191)
No. (%) χ2 p Value OR (95% CI)
Gender 1.689 0.639 Men 1.488 0.223 H/H 93 (58.1) 91 (65.0) 0.747 (0.468, 1.194) H/L, L/L 67 (41.9) 49 (35.0) 1.338 (0.838, 2.137) Women 0.122 0.727 H/H 32 (51.6) 28 (54.9) 0.876 (0.417, 1.842) H/L, L/L 30 (48.4) 23 (45.1) 1.141 (0.543, 2.400) Age 4.059 0.255 ≤ 60 years 0.589 0.443 H/H 25 (61.0) 33 (68.7) 0.710 (0.296, 1.704) H/L, L/L 16 (39.0) 15 (31.3) 1.408 (0.587, 3.379) >60 years 0.782 0.377 H/H 100 (55.2) 86 (60.1) 0.818 (0.524, 1.277) H/L, L/L 81 (44.8) 57 (39.9) 1.222 (0.783, 1.907) aBirthplace 3.070 0.381 Taiwan 0.320 0.571 H/H 66 (57.4) 63 (61.2) 0.855 (0.497, 1.470) H/L, L/L 49 (42.6) 40 (38.8) 1.169 (0.680, 2.010) Mainland China 2.673 0.102 H/H 54 (51.9) 56 (63.6) 0.617 (0.345, 1.102) H/L, L/L 50 (48.1) 32 (36.4) 1.620 (0.907, 2.895)
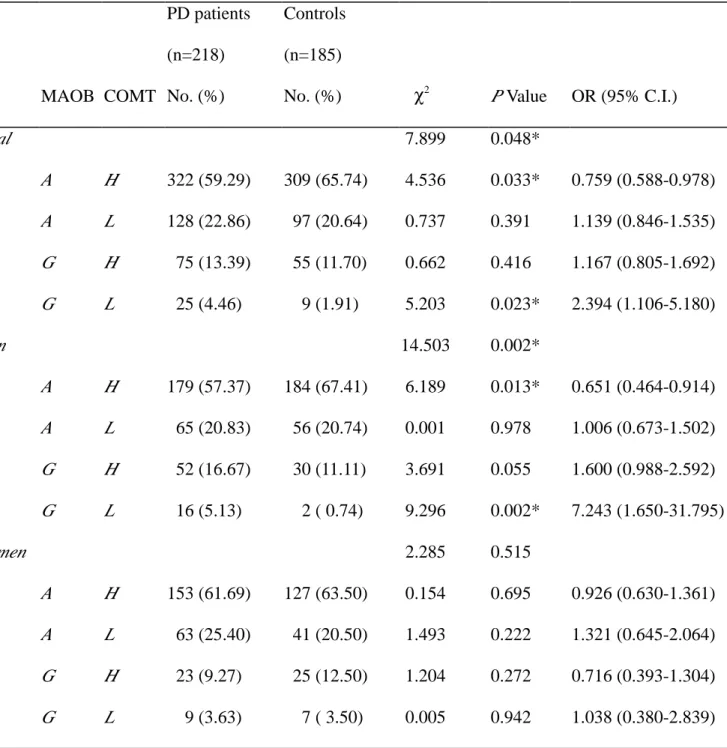
Table 6 Distribution of the combined allele frequencies of MAOB and COMT, and estimated OR in relation to PD risk MAOB COMT PD patients (n=218) No. (%) Controls (n=185) No. (%) χ2 P Value OR (95% C.I.) Total 7.899 0.048* A H 322 (59.29) 309 (65.74) 4.536 0.033* 0.759 (0.588-0.978) A L 128 (22.86) 97 (20.64) 0.737 0.391 1.139 (0.846-1.535) G H 75 (13.39) 55 (11.70) 0.662 0.416 1.167 (0.805-1.692) G L 25 (4.46) 9 (1.91) 5.203 0.023* 2.394 (1.106-5.180) Men 14.503 0.002* A H 179 (57.37) 184 (67.41) 6.189 0.013* 0.651 (0.464-0.914) A L 65 (20.83) 56 (20.74) 0.001 0.978 1.006 (0.673-1.502) G H 52 (16.67) 30 (11.11) 3.691 0.055 1.600 (0.988-2.592) G L 16 (5.13) 2 ( 0.74) 9.296 0.002* 7.243 (1.650-31.795) Women 2.285 0.515 A H 153 (61.69) 127 (63.50) 0.154 0.695 0.926 (0.630-1.361) A L 63 (25.40) 41 (20.50) 1.493 0.222 1.321 (0.645-2.064) G H 23 (9.27) 25 (12.50) 1.204 0.272 0.716 (0.393-1.304) G L 9 (3.63) 7 ( 3.50) 0.005 0.942 1.038 (0.380-2.839)
Table 7 Estimated odds ratio for PD development associated with the coexistence of both MAOB and
COMT susceptibility genotypes
MAOB COMT PD patients (n=218) No. (%) Controls (n=185) No.(%) χ2 P Value OR (95% CI) Combined genotypes 9.845 0.080 A, A/A, A/G H/H 99 (45.41) 100 (54.05) 2.990 0.084 0.707(0.477-1.048) A, A/A, A/G H/L 67 (30.73) 58 (31.35) 0.018 0.894 0.972 (0.636-1.484 A, A/A, A/G L/L 17 (7.80) 11 (5.95) 0.531 0.466 1.338 (0.610-2.933) G, G/G H/H, 21 (9.63) 14 (7.57) 0.538 0.463 1.302 (0.642-2.639) G, G/G H/L 12 (5.50) 2 (1.08) 5.840 0.016* 5.330 (1.177-24.131) G, G/G L/L 2 (0.92) 0 (0.00) N.A. 0.502^ N.A. Combined genotypes 6.724 0.081 A, A/A, A/G H/H, H/L 166 (76.15) 158 (85.41) 5.443 0.020* 0.546 (0.326-0.912) A, A/A, A/G L/L 17 (7.80) 11 (5.95) 0.531 0.466 1.338 (0.610-2.933) G, G/G H/H, H/L 33 (15.14) 16 (8.65) 3.945 0.047* 1.884 (1.001-3.546) G, G/G L/L 2 (0.92) 0 (0.00) N.A 0.502^ N.A Combined genotypes 9.235 0.026* A, A/A, A/G H/H 99 (45.41) 100 (54.05) 2.990 0.084 0.707 (0.477-1.048) A, A/A, A/G H/L,L/L 84 (38.53) 69 (37.30) 0.065 0.799 1.054 (0.704-1.578 G, G/G H/H, 21 (9.63) 14 (7.57) 0.538 0.463 1.302 (0.642-2.639) G, G/G H/L,L/L 14 (6.42) 2 (1.08) 7.088 0.006* 6.279 (1.408-28.001)
*: Significant difference between groups
Figur e Legend
Figure 1. PCR-based RFLP analysis of genetic polymorphisms of MAOB (A) and COMT (B). M : molecular weight marker. Lanes 1 to 3 show MAOB intron 13 polymorphisms : lane 1, the A allele; lane 2, the A/G heterozygote; and lane 3, the G allele. Lanes 4 to 6 show COMT
polymorphisms : lane 4; the wild-type allele (high enzyme activity); lane 5, the heterozygote; and lane 6, the homozygous variant allele (low enzyme activity). Lane 7 shows the PCR-amplified COMT product.
References
1. Riedl AG, Watts PM, Jenner P, Marsden CD. P450 enzymes and Parkinson’s disease: the
story so far. Mov Disord 1998;13:212-220.
2. Riderer P, Konradi C, Hebestreit G, Youdim MBH. Neurochemical perspective to function
of monoamine oxidase. Acta Neurol Scand 1989;126:41-45.
3. Fahn S, Cohen G. The oxidative stress hypothesis in Parkinson’s disease : evidence
supporting it. Ann Neurol 1992;32:804-812.
4. Langston JW, Ballard P, Tetrud JW, Irwin I. Chronic parkinsonism in humans due to a
product of meperidine-analog synthesis. Science 1983;219:979-980.
5. Burns RS., Chiueh CC, Markey SP, Ebert MH, Jacobowitz DM, Kopin IJ. A primate model
of parkinsonism: selective destruction of dopaminergic neurons in the pars compacta of the
substantia nigra by N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Proc Natl Acad Sci
USA 1983;80:4546-4550.
6. Ballard PA, Tetrud JW, Langston JW. Permanent human parkinsonism due to
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP): seven cases. Neurology 1985;35:949-956.
7. Chiba K, Trevor A, Castagnoli N Jr. Metabolism of the neurotoxic tertiary amine, MPTP, by
brain monoamine oxidase. Biochem Biophys Res Commun 1984;120:574-578.
8. Parkinson Study Group. Effect of deprenyl on the progression of disability in early
9. Steventon GB, Sturman SG, Heafield MTE, Waring RH, Napier J, Williams AC. Platelet
monoamine oxidase-B activity in Parkinson’s disease. J Neural Transm [P-D Sect]
1989;1:255-261.
10. Checkoway H, Frankin GM, Costa-Mallen P, et al. A genetic polymorphism of MAO-B
modifies the association of cigarette smoking and Parkinson's disease. Neurology 1998;50:
1458-1461.
11. Fowler JS, Volkow ND, Wang G.-J, et al. Inhibition of monoamine oxidase B in the brains
of smokers. Nature 1996;379:733-736.
12. Kurth JH, Kurth MC, Poduslo SE, Schwankhaus JD. Association of a monoamine oxidase
B allele with Parkinson’s disease. Ann Neurol 1993;33:368-372.
13. Costa P, Checkoway H, Levy D, et al. Association of a polymorphism in intron 13 of the
monoamine oxidase B gene with Parkinson’s disease. Am J Med Genet 1997;74:154-156.
14. Ho SL, Kapadi AL, Ramsden DB, Williams AC. An allelic association study of monoamine
oxidase B in Parkinson’s disease. Ann Neurol 1995;37:403-405.
15. Mellick GD, Buchanan DD, McCann SJ, et al. Variations in the monoamine oxidase B
(MAOB) gene are associated with Parkinson's disease. Mov Disord 1999;14:219-224.
16. Morimoto Y, Murayama N, Kuwano A, Kondo I, Yamashita Y, Mizuno Y. Association of a
polymorphism of the monoamine oxidase B gene with Parkinson’s disease in a Japanese
17. Hwang WJ, Lai ML, Tasi TT, Lai MD. Genetic polymorphism of monoamine oxidase B and
susceptibility of Parkinson’s disease. Chin Med J (Taipei) 1997;60:137-141.
18. Lotta T, Vidgren J, Tilgmann C, et al. Kinetics of human soluble and membrane-bound
catechol-O-methyltransferase: a revised mechanism and description of the thermolabile
variant of the enzyme. Biochemistry 1995;34:4202-4210.
19. Ricera-Calimlim L, Reilly DK. Difference in erythrocyte catechol-O-methyltransferase
activity between Orientals and Caucasians: difference in levodopa tolerance. Clin
Pharmacol Ther 1984;35:804-809.
20. Kunigi H, Nanko S, Ueki A, et al. High and low activity alleles of catechol-
O-methyltransferase gene: ethnic difference and possible association with Parkinson's disease.
Neurosci Lett 1997;221:202-204.
21. Mcleod HL, Syvanen AC, Githang’a J, et al. Ethnic differences in catechol-
O-methyltransferase pharmacogenetics: frequency of the codon 108/158 low activity allele is
lower in Kenyan than Caucasian or South-west Asian individuals. Pharmacogenetics
1998;8:195-199.
22. Hoda F, Nicholl D, Bennett P, et al. No association between Parkinson's disease and
low-activity alleles of catechol-O-methyltransferase. Biochem Biophy Res Commun
1996;228:780-784.
(COMT) gene in Parkinson's disease. Mov Disord 1997;12:426-427.
24. Zhang ZX, Roman GC. Worldwide occurrence of Parkinson‘s disease: an updated review.
Neuroepidemiology 1993;12:195-203.`
25. Yoritaka A, Hattori N, Yoshini H, Mizuno Y. Catechol-O-methyltransferase genotype and
susceptibility to Parkinson's disease in Japan. J Neurol Transm 1997;104:1313-1317.
26. Cochran WG. Some methods of strengthening the common χ2
tests. Biometrics
1954;10:417-451.
27. Hotamisligil GS, Girmen AS, Fink JS, et al. Hereditary variations in monoamine oxidase as
a risk factor for Parkinson’s disease. Mov Disord 1994;9:305-310.
28. Ferrell R, ed. Taiwan aboriginal groups: problems in cultural and linguistic classification.
Taipei: Institute of Ethnology, Academia Sinica, 1969.
29. Chen KH, Cann H, Chen TC, Van West B, Cavalli-Sforza L. Genetic markers of an
aboriginal Taiwanese population. Am J Phys Anthropol 1985;66:327-337.
30. Chen CH, Lee YR, Wei FC, Koong FJ, Hwu HG, Hsiao KJ. Association study of NlaIII and
MspI genetic polymorphisms of catechol-O-methyltransferase gene and susceptibility to
schizophrenia. Biol Psychiatry 1997;41: 985-987.
31. Jenner P. Oxidative mechanisms in nigral cell death in Parkinson’s disease. Mov Disord