Three pentacyclic triterpenes protect H9c2 cardiomyoblast cells against high glucose induced injury
Chien-yi Chan1, Mei-chin Mong2, Wen-hu Liu3, Chun-yin Huang1, Mei-chin Yin1,2,*
1Department of Nutrition, China Medical University, Taichung City, Taiwan
2Department of Health and Nutrition Biotechnology, Asia University, Taichung City, Taiwan
3Radiation Safety Office, Chung Shan Medical University, Taichung City, Taiwan
Running title: triterpenes protect cardiomyoblast cells
*To whom correspondence should be addressed: Dr. Mei-chin Yin, Professor, Department of Nutrition, China Medical University, 91, Hsueh-shih Rd., Taichung City, Taiwan
TEL: 886-4-22053366 ext. 7510, FAX: 886-4-22062891 Email: mcyin@ mail.cmu. edu.tw
1 2
3 4 5
7 8 9 10 11 12 13 14 15 16 17 18 19 20
Abstract
H9c2 cardiomyoblast cell line was used to examine the protection of three triterpenes, asiatic acid, boswellic acid and oleanolic acid, at 5 or 10 M against high glucose induced injury. High glucose stimulated reactive oxygen species (ROS), oxidized glutathione (GSSG), interleukin-6, tumor necrosis factor-alpha and monocyte chemoattractant protein- 1 production, as well as decreased glutathione peroxidase (GPX), glutathione reductase (GR) and catalase activities and protein expression. However, pre-treatments of three triterpenes reserved glutathione, maintained activity and expression of GPX, GR and catalase, as well as lowered ROS, GSSG and inflammatory cytokines generation. High glucose reduced Na+-K+-ATPase activity, raised nuclear factor kappa (NF-) B and caspase-3 activities, up-regulated protein expression of NF-B, mitogen-activated protein kinase, Bax and cleaved caspase-3, as well as down-regulated Bcl-2 expression. Pre- treatments of three triterpenes retained Na+-K+-ATPase activity, declined NF-B and caspase-3 activities, reserved Bcl-2 expression, as well as suppressed protein expression of NF-B, p-p38, Bax and cleaved caspase-3. These findings suggest that these triterpenes are potent cardiac-protective agents.
Keywords: H9c2 cell, asiatic acid, boswellic acid, ROS, NF-κB Abbreviations
AA, asiatic acid; BA, boswellic acid; DMEM, Dulbecco’s modified Eagle’s medium;
DMSO, dimethyl sulphoxide; GPX, glutathione peroxidase; GSH, glutathione; LDH, lactate dehydrogenase; IL, interleukin; MAPK, mitogen-activated protein kinase; MCP, monocyte chemoattractant protein; NF-B, nuclear factor kappa B; OA, oleanolic acid;
ROS, reactive oxygen species; TNF, tumor necrosis factor 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Introduction
Hyperglycemia induced cardiac injury is the major cause of mortality in people with diabetes [1]. Several studies indicated that hyperglycemia enhances cardiac production of free radicals such as reactive oxygen species (ROS) and inflammatory cytokines such as interleukin (IL)-6, tumor necrosis factor (TNF)-alpha and monocyte chemoattractant protein (MCP)-1; and these free radicals and cytokines impair cardiac contractile function and promote the development of myocardial infarction, cardiac hypertrophy and even heart failure [2,3]. Furthermore, it is reported that hyperglycemia activates both nuclear factor kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) pathways in myocardial cells, which in turn elicits transcription of genes encoded for oxidative and inflammatory factors in cardiac tissue, and facilitates progression of cardiac disorders [4,5]. In addition, mitochondrial apoptotic pathway is involved in high glucose caused myocardial cell death, in which the protein expression of apoptotic factors such as Bax and caspase-3 is raised, but the expression of anti-apoptotic factors such as Bcl-2 is reduced [6,7]. Thus, any agent(s) with anti-oxidative, anti-inflammatory and anti-apoptotic activities may potentially protect cardiomyocytes against high glucose induced injury.
Asiatic acid (AA), boswellic acid (BA) and oleanolic acid (OA) are pentacyclic triterpenes naturally occurring in many vegetables and fruits such as glossy privet fruit (Ligustrum lucidum Ait.), basil (Ocimum basilicum) and brown mustard (Brassica juncea) [8,9]. It is reported that these triterpenes could decline activity or expression of NF-κB and MAPK, and reduce apoptotic stress via lowering caspase-3 activity and Bax formation in neurons and brain [10-12]. Thus, these compounds might benefit cardiac cell survival through regulating these signaling pathways. The improvement of AA upon hyperglycemia in diabetic rats has been observed [13]; however, the cardiac protective effect of this agent under diabetic condition remains unknown. So far, less attention was 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
paid to the impact of BA upon diabetic complications. Our previous animal study found that OA via anti-oxidative and anti-inflammatory activities alleviated the progression of diabetic nephropathy in mice [14]. It is reasonable to speculate that this compound may also provide cardiac protection against the progression of hyperglycemia induced cardiac injury. In addition, Mapanga et al. [15] indicated that OA post-treatments, acting like a therapeutic agent, in high glucose treated H9c2 cells attenuated oxidative damage and improved cell viability. Thus, it is highly possible that pre-treatments of OA, acting like a preventive agent, in cardiac cells could exhibit protective effects to counteract subsequent high glucose evoked injury.
In our present study, the protection of AA, BA and OA at various doses against high glucose induced oxidative, inflammatory and apoptotic stress in H9c2 cell line, a rat embryonic cardiomyoblast cell line, was examined. The effects of these compounds upon activity and protein expression of NF-B, MAPK, Bcl-2, Bax and caspase-3 were evaluated. OA was used not only for further demonstration regarding its cardiac-protective effects but also for comparison.
Materials and methods Chemicals
Asiatic acid (AA, 98%), boswellic acid (BA, 98.5%), oleanolic acid (OA, 99%), and other chemicals were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). Medium, plates, antibiotics, and chemicals used for cell culture were purchased from Difco Laboratory (Detroit, MI, USA). All chemicals used in these measurements were the highest purity commercially available.
Cell culture
H9c2 cells obtained from American Type Culture Collection (ATCC, Rockville, MD, 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
USA) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 1.5 g/l NaHCO3 and 5.5 mM glucose, supplemented with 10% fetal bovine serum, 100 U/ml penicillin, and 100 U/ml streptomycin under 95% air/ 5% CO2 at 37°C. The culture medium was changed every three days, and cells were subcultured once a week. The medium was changed to serum-deprived medium, and cells were washed with serum-free DMEM 24 hr before experiments and replanted in the 96 well plates. A phosphate buffer saline (PBS, pH 7.2) was used to adjust the cell number to 105/ml for various experiments and analyses.
Experimental design
AA, BA or OA was dissolved in dimethyl sulfoxide (DMSO), and diluted with the medium. The final concentration of DMSO in the medium was 0.5%. At this concentration, DMSO did not affect any measurements (data not shown). H9c2 cells (105 cells/ml) were treated with OA, AA and BA at 5 or 10 M for 48 hr at 37°C, which led to 97.5% incorporation of test agent into cells. After washing with serum-free DMEM, cells were further treated with medium containing glucose at 5.5 mM (G5.5) or 33 mM (G33) for 24 hr at 37°C. Cells treated with G5.5 alone (without triterpenes) were control groups.
Mannitol at 5.5 and 33 mM was also used to examine the influence of hyperosmolarity.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
MTT assay was performed to examine cell viability. Briefly, H9c2 cells were incubated with 0.25 mg MTT/ml for 3 hr at 37°C. The amount of MTT formazan product was determined by measuring absorbance at 570 nm (630 nm as a reference) using a microplate reader (Bio-Rad, Hercules, CA, USA). Cell viability was expressed as a percentage of control groups.
Lactate dehydrogenase (LDH) assay
The plasma membrane damage of H9c2 cells was evaluated by measuring LDH variation.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Fifty l of culture supernatants were collected from each well. LDH activity (U/l) was analyzed by a colorimetric LDH assay kit (Sigma Chemical Co., St. Louis, MO, USA).
Analyses for glutathione (GSH), oxidize glutathione (GSSG), ROS, and activity of glutathione peroxidase (GPX), glutathione reductase (GR) and catalase
Cells were washed twice with PBS (pH 7.2), then were scraped from the plates and followed by homogenizing in PBS containing 0.5 mM butylated hydroxytoluene to prevent sample further oxidation. The homogenate was centrifuged at 3000 xg for 20 min at 4°C, and the supernatant was used for these assays according to the manufacturer’s instructions.
GSH or GSSG concentration (ng/mg protein) was quantified by commercial colorimetric assay kits (OxisResearch, Portland, OR, USA). ROS level was determined using an oxidation sensitive dye, 2',7'-dichlorofluorescein diacetate. Cells were incubated with 50 μM dye for 30 min and washed with PBS. Fluorescence value was measured by using a fluorescence microplate reader at excitation and emission wavelengths of 485 and 530 nm, respectively. Relative fluorescence unit (RFU) was the difference in fluorescence values obtained at time 0 and 5 min. Results are expressed as RFU/mg protein. The activity (U/mg protein) of GPX, GR and catalase in H9c2 cells was assayed by using assay kits (EMD Biosciences, San Diego, CA, USA). Protein concentration was measured by Bio- Rad protein assay reagent (Bio-Rad Laboratories Inc. Hercules, CA, USA).
Cytokine measurement
After washed and homogenized, the released level of IL-6, TNF-alpha and MCP-1 in supernatant was measured by ELISA method using cytoscreen immunoassay kits (BioSource Int., Camarillo, CA, USA). The sensitivity of assay with the detection limit was 5 pg/mg protein for IL-6, and 10 pg/mg protein for TNF-alpha and MCP-1.
Na+-K+-ATPase activity assay
Na+-K+-ATPase activity was detected by measuring the amount of inorganic phosphate (Pi) 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
released from ATP according to the method of Torlinska and Grochowalska [16]. The reaction mixture contained 100 mM NaCl, 20 mM KCl, 2 mM ATP, 30 mM Tris-HCl buffer (pH 7.4) and the freshly isolated cellular mitochondria. The assay was initiated by adding ATP, and terminated by adding 15% trichloroacetic acid after 15 min incubation at 37°C. The released Pi was assayed by measuring the absorbance at 640 nm. A unit was defined as mol Pi/mg protein/hr. The values of treated cells were expressed as percentage of control groups.
Measurement of caspase-3 activity
Control or treated cells were lysed in 50 ml cold lysis buffer containing 10 mM HEPES (pH 7.9), 1.5 mM MgCl2, 10 mM KCl, 0.1 mM EDTA, 1 mM DTT, 1 mM PMSF and 0.6% Nonidet P-40, and followed by centrifugation at 10,000 xg for 10 min at 4°C. The supernatant from lysed cells was added to the reaction mixture containing dithiothreitol and caspase-3 substrate (N-acetyl-Asp-Glu-Val-Asp-p-nitroanilide), and further incubated for 1 hr at 37°C. Caspase-3 activity was determined by a commercial caspase-3 activity assay kit (Upstate, Lake Placid, NY, USA). Data were expressed as percentage of control groups.
Preparation of nuclear extract
Cells were washed in cold PBS, and re-suspended in above lysis buffer, and followed by centrifugation at 1200 xg for 10 min. Nuclear pellets were re-suspended in buffer containing 20 mM HEPES, 0.4 M NaCl, 1 mM EDTA, 1 mM DTT and 25% glycerol.
After incubation at 4°C for 30 min and centrifugation at 21,000 xg for 30 min, supernatants containing nuclear protein were collected.
NF-κB activity assay
NF-κB p50/65 DNA binding activity in the nuclear extract of H9c2 cells was determined by a commercial kit (Chemicon International Co., Temecula, CA, USA). The binding of 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
activated NF-κB was examined by adding a primary polyclonal anti-NF-κB p50/p65 antibody, a secondary antibody conjugated with horseradish peroxidase, and the 3,3′,5,5′- tetramethylbenzidine substrate. Absorbance at 450 nm was read. Values are expressed as relative optical density (OD)/mg protein.
Western blot analysis
H9c2 cells were homogenized in buffer containing 0.5% Triton X-100 and protease- inhibitor cocktail (1:1000, Sigma-Aldrich Chemical Co., St. Louis, MO, USA). This homogenate was further mixed with buffer (60 mM Tris-HCl, 2% SDS and 2% β- mercaptoethanol, pH 7.2), and boiled for 5 min. Sample at 40 μg protein was applied to 10% SDS-polyacrylamide gel electrophoresis, and transferred to a nitrocellulose membrane (Millipore, Bedford, MA, USA) for 1 hr. After blocking with a solution containing 5% nonfat milk for 1 hr to prevent non-specific binding of antibody, membrane was incubated with mouse anti-cleaved caspase-3 (1:1000), anti-Bcl-2 (1:2000), anti-Bax (1:1000), anti-GPX, anti-GR, anti-catalase (1:500), anti-NF-κB p65, anti-NF-κB p50 (1:1000) and anti-MAPK (1:2000) monoclonal antibody (Boehringer-Mannheim, Indianapolis, IN, USA) at 4ºC overnight, and followed by reacting with horseradish peroxidase-conjugated antibody for 3.5 hr at room temperature. The detected bands were quantified by an image analyzer (ATTO, Tokyo, Japan), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a loading control. The blot was quantified by densitometric analysis. Results were normalized to GAPDH, and given as arbitrary units (AU).
Statistical analysis
The effect of each treatment was analyzed from ten different preparations (n=10). Data were reported as means standard deviation (SD), and subjected to analysis of variance.
Differences among means were determined by the Least Significance Difference Test with 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
significance defined at P<0.05.
Results
As shown in Figure 1a and 1b, 5.5 and 33 mM mannitol did not affect MTT or LDH assays when compared with 5.5 mM glucose (P>0.05). Thus, the influence of hyperosmolarity from 33 mM glucose was ruled out. High glucose (33 mM) significantly reduced 55% cell viability (1c) and raised 220 U/l LDH activity (1d) (P<0.05); however, triterpene pre- treatments enhanced 21-42% cell survival and decreased 67-181 U/l LDH activity (P<0.05), in which 10 M AA treatment resulted in the lowest LDH activity (P<0.05).
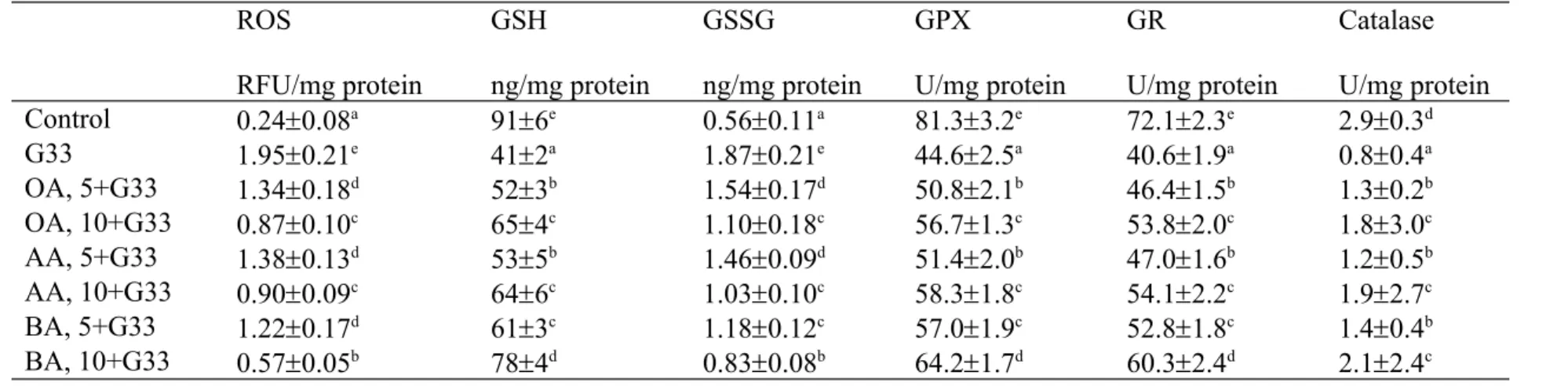
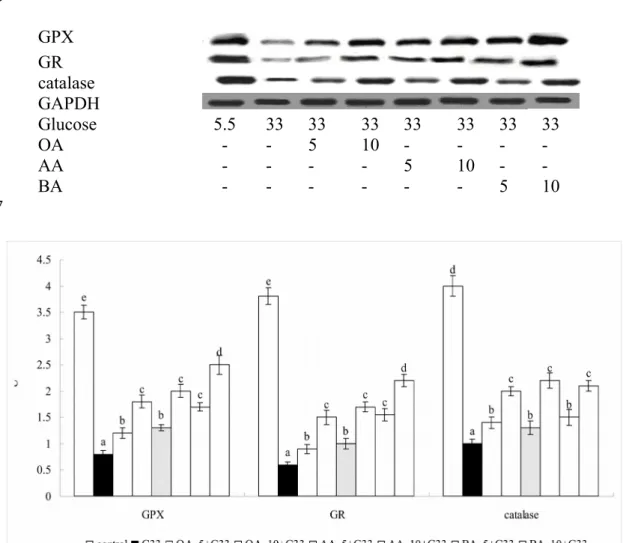
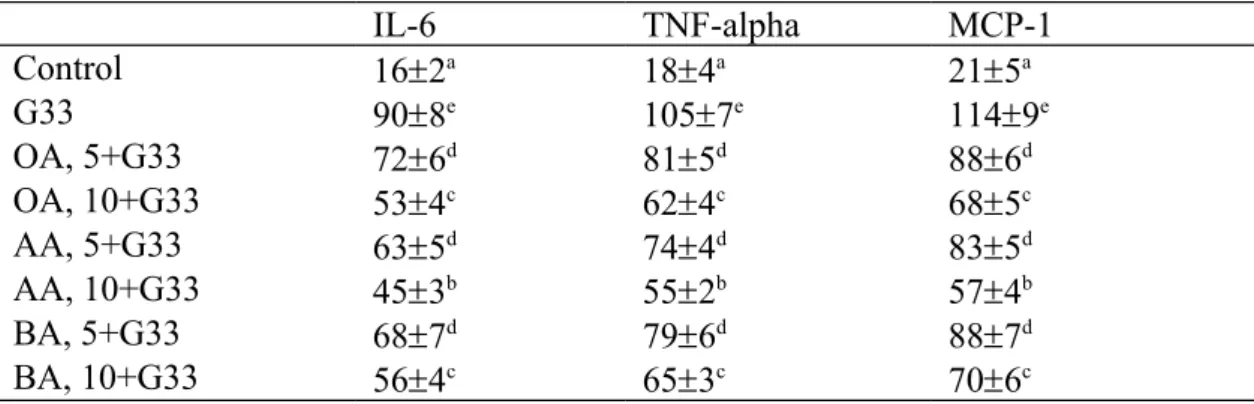
High glucose lowered GSH level, stimulated ROS and GSSG production; also decreased activity and expression of GPX, GR and catalase in H9c2 cells (Table 1 and Figure 2, P<0.05). However, pre-treatments of triterpenes significantly preserved GSH level at 11-37 ng/mg protein, maintained GPX activity at 6.2-19.6 U/mg, GR activity at 5.9-19.9 U/mg protein, catalase activity at 0.4-1.3 U/mg protein, as well as reduced ROS and GSSG levels at 0.57-1.38 RFU/mg protein and 0.33-1.04 ng/mg protein, respectively (P<0.05), in which 10 M BA treatment had the highest GSH content and GPX activity, and the lowest ROS level when compared with other triterpenes treatments (P<0.05). As shown in Table 2, high glucose increased the release of IL-6, TNF-alpha and MCP-1 (P<0.05); and triterpene pre-treatments decreased these cytokines level at 18-54, 24-51 and 26-57 pg/mg protein, respectively (P<0.05), in which cells pre-treated with 10 M AA had the lowest levels of IL-6, TNF-alpha and MCP-1 when compared with other triterpenes treatments (P<0.05).
As shown in Figure 3, high glucose reduced 63% Na+-K+-ATPase activity (3a) and raised 2.7 folds caspase-3 activity (3b) (P<0.05). The pre-treatments of test triterpenes increased 13-43% retain in Na+-K+-ATPase activity and lowered caspase-3 activity to 1.5- 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
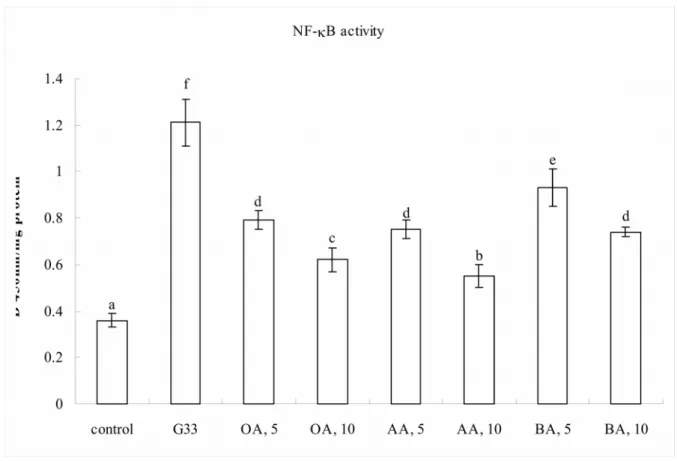
2.1 folds of controls (P<0.05). High glucose increased 3.3 folds NF-B activity (Figure 4a), and enhanced 4.4, 3.1 and 5.7 folds protein expression of NF-B p50, NF-B p65 and p-p38 (4b, P<0.05). Triterpenes pre-treatments declined NF-B activity to 1.5-2.5 folds of controls; and suppressed NF-B p50 expression to 1.7-3.8 folds of controls (P<0.05).
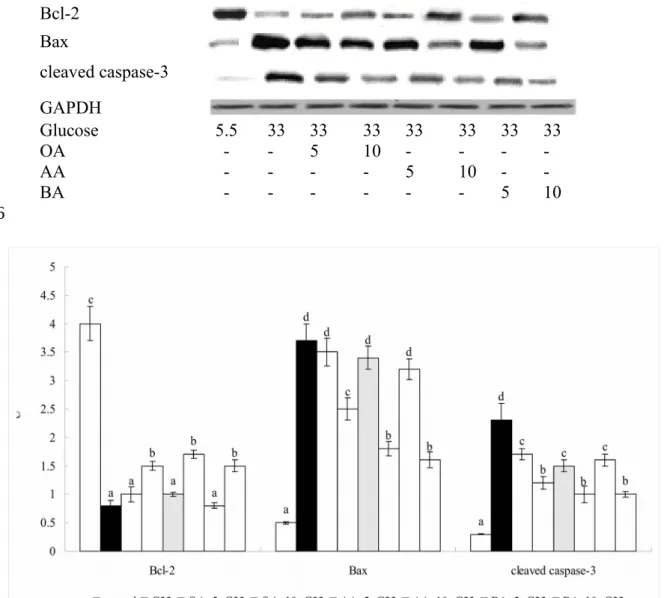
These triterpenes only at 10 M down-regulated NF-B p65 and p-p38 expression, in which 10 M AA had the lowest p-p38 expression (P<0.05). High glucose suppressed 78% Bcl-2 expression, and increased 7.4 and 7.6 folds protein production of Bax and cleaved caspase-3 (Figure 5, P<0.05). Test triterpenes dose-dependently down-regulated cleaved caspase-3 expression to 3.3-5.8 folds of controls (P<0.05), but only at 10 M reserved Bcl-2 expression and suppressed Bax expression (P<0.05).
Discussion
Mapanga et al. [15] reported that post-addition of oleanolic acid at 20 and 50 M in high glucose treated H9c2 cells could decrease oxidative stress and increase cell viability. Our present study revealed that pre-treatments from oleanolic acid and other two triterpenes, asiatic acid and boswellic acid, at 5 or 10 M effectively protected H9c2 cardiomyoblast cells to resist subsequent high glucose evoked oxidative, inflammatory and apoptotic stress, which consequently benefited membrane integration and survival of H9c2 cells.
Furthermore, we found that these triterpenes maintained Na+-K+-ATPase activity, and suppressed NF-B and p38 activation in this cardiomyoblast cell line. These findings support that oleanolic acid, asiatic acid and boswellic acid at lower doses may act as preventive agents to protect cardiac cells against high glucose induced injury.
Oxidative stress derived from hyperglycemia is a crucial factor responsible for the progression of diabetes related cardiac disorders [17,18]. Particularly, it is documented 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
that ROS overproduction facilitated deterioration in insulin-resistant or obesity related heart diseases [19,20]. Our present study found that three triterpenes pre-treatments at two doses efficiently decreased ROS production in H9c2 cells, which contributed to reduce oxidative stimulation in these cells. Furthermore, these triterpenes effectively preserved GSH content, lowered GSSG formation, as well as retained GPX and GR activities and protein expression in H9c2 cells. These data revealed that these agents could promote glutathione redox cycle to exert anti-oxidative action. In addition, we found that test triterpenes substantially reserved catalase activity and protein expression. Apparently, these agents could enhance anti-oxidative defense of H9c2 cells through mediating multiple factors, and counteracted subsequent high glucose evoked oxidative stress. On the other hand, ROS has been considered as a crucial stimulator for cardiac mitochondrial apoptotic pathway in diabetic progression [21,22]. Since triterpenes treatments already lowered ROS formation, the observed greater mitochondrial stability and less apoptotic stress in triterpenes-treated H9c2 cells could be explained. The development of diabetic cardiomyopathy is usually accompanied with a local rise in cytokines such as IL-6 and TNF-alpha in cardiac cells [23,24]. Our present study found that pre-treatments of these triterpenes diminished subsequent high glucose elicited inflammatory stress in H9c2 cells via lowering IL-6, TNF-alpha and MCP-1 formation. These results agreed that these agents could provide anti-inflammatory protection for cardiac cells. Younce et al. [25]
indicated that MCP-1 was mainly responsible for hyperglycemia-induced cardiomyocyte death because its overproduction exacerbated ROS formation and endoplasmic reticulum oxidative stress, which finally caused autophagy. In our present study, triterpenes-treated H9c2 cells had lower MCP-1 and ROS levels. Thus, the observed protective effects from these compounds could be partially ascribed to their anti-MCP-1 activity. Since oxidative and inflammatory injury were mitigated, it was reasonable to observe the greater cell 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
viability and less LDH activity in these triterpenes-treated H9c2 cells.
Enhanced activation of NF-B and MAPK pathways promoted the progression of diabetic cardiomyopathy via up-regulating several sets of genes involved in the reactions of oxidation, inflammation, endothelial dysfunctions and apoptosis [26-28]. Westermann et al. [29] reported that the inhibition of p38 MAPK prevented cardiac inflammation and attenuated left ventricular dysfunction in diabetic cardiomyopathy. We found the pre- treatments from test triterpenes markedly abated subsequent high glucose stimulated NF-
B activation and p38 phosphorylation in H9c2 cells, which contributed to suppress oxidative and inflammatory reactions, and decrease ROS and cytokines production. These findings implied that these compounds might prevent high glucose induced cardiotoxicity through declining NF-B and p38 activation. In addition, we notified that asiatic acid exhibited greater effects than other two agents in lowering NF-B activity, NF-B protein expression and inflammatory cytokines production; but boswellic acid was more effective in decreasing ROS generation and p38 phosphorylation. It seems that the protective mechanisms of these three triterpenes upon H9c2 cells were not identical. It is highly possible that other pathways are involved in the action modes of these agents for cardiac cells against high glucose induced injury. Further studies using NF-B and p38 inhibitors, and examining other pathways should be processed in order to clearly elucidate the roles of these pathways upon the mechanism of these triterpenes.
The impairment of Na+-K+-ATPase activity in mitochondria and sarcoplasmic reticulum of cardiac cells affected sodium, potassium and calcium homeostasis, and retarded contraction and relaxation of myocardia [30,31]. Cai et al. [32] indicated that hyperglycemia related myocardial apoptosis was partially mediated by mitochondrial caspase-3 activation pathway. In our present study, test triterpenes effectively maintained Na+-K+-ATPase activity and repressed caspase-3 activity. These results revealed that these 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
triterpenes were able to stabilize mitochondrial membrane of H9c2 cells against subsequent high glucose caused disturbance. Bcl-2 is an anti-apoptotic molecule, and Bax and caspase-3 are pro-apoptotic molecules. We found test triterpenes at 10 M retained Bcl-2 expression, and down-regulated Bax and cleaved caspase-3 production, which consequently diminished apoptotic injury and benefited cell survival. These findings implied that these compounds could penetrate into H9c2 cells and mitigate apoptotic stress under high glucose condition by regulating both anti-apoptotic and pro-apoptotic molecules. The study of Yin et al. [9] reported that dietary intake of several triterpenes including oleanolic acid, asiatic acid and boswellic acid resulted in their deposit in heart of normal mice. Thus, the application of these triterpenes through dietary intake to increase cardiac protection against diabetic cardiomyopathy seems feasible. Further studies including other cardiac cell lines such as HL-1 could be helpful to verify the cardiac protection from these triterpenes. Also, further in vivo study is encouraged to ensure the effects and safety of these agents upon the progression of diabetes associated cardiac disorders.
In conclusion, pre-treatments from oleanolic acid, asiatic acid, and boswellic acid protected H9c2 cells to resist subsequent high glucose induced injury. These agents exhibited anti-oxidative, anti-inflammatory and anti-apoptotic effects via lowering ROS production, enhancing glutathione redox cycle, suppressing NF-B and p38 activation, and down-regulating the protein expression of Bax and cleaved caspase-3. These findings suggest that these triterpenes are potent cardiac protective agents.
Conflict of interest None.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
References
[1] Aneja A, Tang WH, Bansilal S, Garcia MJ, Farkouh ME. Diabetic cardiomyopathy : insights into pathogenesis, diagnostic challenges, and therapeutic options. Am J Med 2008;121:748-757.
[2] Rajamani U, Essop MF. Hyperglycemia-mediated activation of the hexosamine biosynthetic pathway results in myocardial apoptosis. Am J Physiol Cell Physiol 2010;299:C139-C147.
[3] Drimal J, Knezl V, Navarova J, Nedelcevova J, Paulovicova E, Sotnikova VR, et al.
Role of inflammatory cytokines and chemoattractants in rat model of streptozotocin- induced diabetic heart failure. Endocr Regul 2008;42:129-135.
[4] Lorenzo O, Picatoste B, Ares-Carrasco S, Ramírez E, Egido J, Tuñón J. Potential role of nuclear factor κB in diabetic cardiomyopathy. Mediators Inflamm 2011;2011;652097.
[5] Thandavarayan RA, Giridharan VV, Sari FR, Arumugam S, Veeraveedu PT, Pandian GN, et al. Depletion of 14-3-3 protein exacerbates cardiac oxidative stress, inflammation and remodeling process via modulation of MAPK/NF-ĸB signaling pathways after streptozotocin-induced diabetes mellitus. Cell Physiol Biochem 2011;28:911-922.
[6] Pollack M, Phaneuf S, Dirks A, Leeuwenburgh C. The role of apoptosis in the normal aging brain, skeletal muscle, and heart. Ann NY Acad Sci 2002;959:93-107.
[7] Zhou G, Li X, Hein DW, Xiang X, Marshall JP, Prabhu SD, et al. Metallothionein suppresses angiotensin II-induced nicotinamide adenine dinucleotide phosphate oxidase activation, nitrosative stress, apoptosis, and pathological remodeling in the diabetic heart. J Am Coll Cardiol 2008;52:655-666.
[8] Yin MC, Chan KC. Nonenzymatic antioxidative and antiglycative effects of oleanolic 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
acid and ursolic acid. J Agric Food Chem 2007;55:7177-7181.
[9] Yin MC, Lin MC, Mong MC, Lin CY. Bioavailability, distribution, and antioxidative effects of selected triterpenes in mice. J Agric Food Chem 2012;60:7697-7701.
[10] Cuaz-Pérolin C, Billiet L, Baugé E, Copin C, Scott-Algara D, Genze F, et al.
Antiinflammatory and antiatherogenic effects of the NF-kappaB inhibitor acetyl-11- keto-beta-boswellic acid in LPS-challenged ApoE-/- mice. Arterioscler Thromb Vasc Biol 2008;28:272-277.
[11] Yun KJ, Kim JY, Kim JB, Lee KW, Jeong SY, Park HJ, et al. Inhibition of LPS- induced NO and PGE2 production by asiatic acid via NF-kappa B inactivation in RAW 264.7 macrophages: possible involvement of the IKK and MAPK pathways. Int Immunopharmacol 2008;8:431-441.
[12] Tsai SJ, Yin MC. Anti-oxidative, anti-glycative and anti-apoptotic effects of oleanolic acid in brain of mice treated by d-galactose. Eur J Pharmacol 2012;689:81-88.
[13] Liu J, He T, Lu Q, Shang J, Sun H, Zhang L. Asiatic acid preserves beta cell mass and mitigates hyperglycemia in streptozocin-induced diabetic rats. Diabetes Metab Res Rev 2010;26:448-454.
[14] Wang ZH, Hsu CC, Huang CN, Yin MC. Anti-glycative effects of oleanolic acid and ursolic acid in kidney of diabetic mice. Eur J Pharmacol 2010;628:255-260.
[15] Mapanga RF, Rajamani U, Dlamini N, Zungu-Edmondson M, Kelly-Laubscher R, Shafiullah M, et al. Oleanolic acid: a novel cardioprotective agent that blunts hyperglycemia-induced contractile dysfunction. PLoS One 2012;7:e47322.
[16] Torlinska T, Grochowalska A. Age-related changes of Na+, K+-ATPase, Ca2+-ATPase and Mg2+-ATPase activities in rat brain synaptosomes. J Physiol Pharmacol 2004;55:457-465.
[17] Grieve DJ, Shah AM. Oxidative stress in heart failure. More than just damage. Eur 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Heart J 2003;24:2161–2163.
[18] Turan B. Role of antioxidants in redox regulation of diabetic cardiovascular complications. Curr Pharm Biotechnol 2010;11:819-836.
[19] Mellor KM, Ritchie RH, Delbridge LM. Reactive oxygen species and insulin-resistant cardiomyopathy . Clin Exp Pharmacol Physiol 2010;37:222-228.
[20] Huang CY, Lee SD. Possible pathophysiology of heart failure in obesity: Cardiac apoptosis. BioMedicine 2012; 2:36-40.
[21] Maalouf RM, Eid AA, Gorin YC, Block K, Escobar GP, Bailey S, Abboud HE. Nox4- derived reactive oxygen species mediate cardiomyocyte injury in early type 1 diabetes. Am J Physiol Cell Physiol 2012;302:C597-C604.
[22] Giacco F, Brownlee M. Oxidative stress and diabetic complications. Circ Res 2010;107:1058-1070.
[23] Chao PC, Hsu CC, Yin MC. Anti-inflammatory and anti-coagulatory activities of caffeic acid and ellagic acid in cardiac tissue of diabetic mice. Nutr Metab (Lond) 2009;6:33.
[24] Palomer X, Salvadó L, Barroso E, Vázquez-Carrera M. An overview of the crosstalk between inflammatory processes and metabolic dysregulation during diabetic cardiomyopathy . Int J Cardiol 2013;168:3160-3172.
[25] Younce CW, Wang K, Kolattukudy PE. Hyperglycaemia-induced cardiomyocyte death is mediated via MCP-1 production and induction of a novel zinc-finger protein MCPIP. Cardiovasc Res 2010;87:665-674.
[26] Min W, Bin ZW, Quan ZB, Hui ZJ, Sheng FG. The signal transduction pathway of PKC/NF-kappa B/c-fos may be involved in the influence of high glucose on the cardiomyocytes of neonatal rats. Cardiovasc Diabetol 2009;8:8.
[27] Yar AS, Menevse S, Alp E. The effects of resveratrol on cyclooxygenase-1 and -2, 1
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
nuclear factor kappa beta, matrix metalloproteinase-9, and sirtuin 1 mRNA expression in hearts of streptozotocin-induced diabetic rats. Genet Mol Res 2011;10:2962-2975.
[28] Chen S, Khan ZA, Karmazyn M, Chakrabarti S. Role of endothelin-1, sodium hydrogen exchanger-1 and mitogen activated protein kinase ( MAPK ) activation in glucose-induced cardiomyocyte hypertrophy. Diabetes Metab Res Rev 2007;23:356- 367.
[29] Westermann D, Rutschow S, Van Linthout S, Linderer A, Bücker-Gärtner C, Sobirey M, et al. Inhibition of p38 mitogen-activated protein kinase attenuates left ventricular dysfunction by mediating pro-inflammatory cardiac cytokine levels in a mouse model of diabetes mellitus. Diabetologia 2006;49:2507-2513.
[30] Jia JH, Chen KP, Chen SX, Liu KZ, Fan TL, Chen YC. Breviscapine, a traditional Chinese medicine, alleviates myocardial ischaemia reperfusion injury in diabetic rats.
Acta Cardiol 2008;63:757-762.
[31] Dhalla NS, Rangi S, Zieroth S, Xu YJ. Alterations in sarcoplasmic reticulum and mitochondrial functions in diabetic cardiomyopathy . Exp Clin Cardiol 2012;17:115- 120.
[32] Cai L, Li W, Wang G, Guo L, Jiang Y, Kang YJ. Hyperglycemia-induced apoptosis in mouse myocardium: mitochondrial cytochrome C-mediated caspase-3 activation pathway. Diabetes 2002;51:1938-1948.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Figure 1. Effect of glucose or mannitol at 5.5 or 33 mM alone upon cell viability determined by MTT assay (1a) and plasma membrane damage determined by LDH assay (1b). Effects of OA, AA or BA at 5 or 10 M, and post-treated by 33 mM glucose (G33) upon MTT assay (1c) and LDH assay (1d). Cells contained no triterpenes and treated with 5.5 mM glucose were used as control. Data are meanSD (n=10). a-eMeans among bars without a common letter differ, P<0.05.
1 2 3 4 5 6
1a.
1b.
1
2 3
4
1c.
1d.
1
2 3
4
Table 1. Effect of OA, AA or BA upon level of ROS, GSH, GSSG, and activity of GPX, GR and catalase. H9c2 cells were pre-treated with OA, AA or BA at 5 or 10 M, and followed by adding 33 mM glucose (G33). Cells contained no triterpenes and treated with 5.5 mM glucose were used as control. Data are meanSD (n=10).
ROS
RFU/mg protein
GSH
ng/mg protein
GSSG
ng/mg protein
GPX
U/mg protein
GR
U/mg protein
Catalase U/mg protein
Control 0.240.08a 916e 0.560.11a 81.33.2e 72.12.3e 2.90.3d
G33 1.950.21e 412a 1.870.21e 44.62.5a 40.61.9a 0.80.4a
OA, 5+G33 1.340.18d 523b 1.540.17d 50.82.1b 46.41.5b 1.30.2b
OA, 10+G33 0.870.10c 654c 1.100.18c 56.71.3c 53.82.0c 1.83.0c
AA, 5+G33 1.380.13d 535b 1.460.09d 51.42.0b 47.01.6b 1.20.5b
AA, 10+G33 0.900.09c 646c 1.030.10c 58.31.8c 54.12.2c 1.92.7c
BA, 5+G33 1.220.17d 613c 1.180.12c 57.01.9c 52.81.8c 1.40.4b
BA, 10+G33 0.570.05b 784d 0.830.08b 64.21.7d 60.32.4d 2.12.4c
a-eMeans in a column without a common letter differ, P<0.05.
1 2 3
4
Figure 2. Effect of OA, AA or BA upon protein expression of GPX, GR and catalase. H9c2 cells were pre-treated with OA, AA or BA at 5 or 10 M and followed by adding 33 mM glucose (G33). Cells contained no triterpenes and treated with 5.5 mM glucose (G5.5) were used as control. Data are meanSD (n=10). a-eMeans among bars without a common letter differ, P<0.05.
GPX GR catalase GAPDH
Glucose 5.5 33 33 33 33 33 33 33
OA - - 5 10 - - - -
AA - - - - 5 10 - -
BA - - - 5 10
1 2 3 4 5 6
7
8
Table 2. Effect of OA, AA or BA upon level (pg/mg protein) of IL-6, TNF-alpha and MCP-1. H9c2 cells were pre-treated with OA, AA or BA at 5 or 10 M, and followed by adding 33 mM glucose (G33). Cells contained no triterpenes and treated with 5.5 mM glucose were used as control. Data are meanSD (n=10).
IL-6 TNF-alpha MCP-1
Control 162a 184a 215a
G33 908e 1057e 1149e
OA, 5+G33 726d 815d 886d
OA, 10+G33 534c 624c 685c
AA, 5+G33 635d 744d 835d
AA, 10+G33 453b 552b 574b
BA, 5+G33 687d 796d 887d
BA, 10+G33 564c 653c 706c
a-eMeans in a column without a common letter differ, P<0.05.
1 2 3 4 5
6
Figure 3. Effect of OA, AA or BA upon activity of Na+-K+-ATPase (a) and caspase-3 (b).
H9c2 cells were pre-treated with OA, AA or BA at 5 or 10 M, and followed by adding 33 mM glucose (G33). Cells contained no triterpenes and treated with 5.5 mM glucose were used as control. Data are meanSD (n=10). a-eMeans among bars without a common letter differ, P<0.05.
1 2 3 4 5 6
3a.
3b.
1
2 3 4
5
Figure 4. Effect of OA, AA or BA upon NF-B activity (a), expression of NF-B and MAPK (b). H9c2 cells were pre-treated with OA, AA or BA at 5 or 10 M, and followed by adding 33 mM glucose (G33). Cells contained no triterpenes and treated with 5.5 mM glucose (G5.5) were used as control. Data are meanSD (n=10). a-fMeans among bars without a common letter differ, P<0.05.
4a.
1 2 3 4 5 6 7
8
4b.
NF-B p50 NF-B p65 p-38 p-p38 JNK p-JNK ERK1/2 p-ERK1/2 GAPDH
Glucose 5.5 33 33 33 33 33 33 33
OA - - 5 10 - - - -
AA - - - - 5 10 - -
BA - - - 5 10
1
1
Figure 5. Effect of OA, AA or BA upon expression of Bcl-2, Bax and cleaved caspase-3. H9c2 cells were pre-treated with OA, AA or BA at 5 or 10 M and followed by adding 33 mM glucose (G33). Cells contained no triterpenes and treated with 5.5 mM glucose (G5.5) were used as control. Data are meanSD (n=10). a-dMeans among bars without a common letter differ, P<0.05.
Bcl-2 Bax
cleaved caspase-3 GAPDH
Glucose 5.5 33 33 33 33 33 33 33
OA - - 5 10 - - - -
AA - - - - 5 10 - -
BA - - - 5 10
1 2 3 4 5
6
7 8