國立臺灣大學醫學院藥理學研究所 博士論文
Graduate Institute of Pharmacology College of Medicine
National Taiwan University Doctoral Dissertation
以藥理學方法在大鼠中腦環導水管灰質腦切片研究 Nociceptin/Orphanin FQ 受體之功能異質性
A Study on the Functional Heterogeneity of
Nociceptin/Orphanin FQ Receptors in Rat Periaqueductal Gray Slices by Pharmacological Approach
廖彥昱 YAN-YU LIAO
指導教授:邱麗珠 博士 Advisor: LIH-CHU CHIOU, Ph.D.
中華民國 100 年 1 月
January, 2011
誌謝
唸博士班著實是一段漫長的路程,發生了不少五味雜陳的事,徬徨過、失落 過、激動過,背負了不少壓力,當然也收獲了許多、成長了許多,一路走來,最 感謝恩師邱麗珠教授的栽培,儘管我的資質駑鈍,老師都能不厭其煩的給予教導、
建議及鼓勵,使我得以順利畢業並一窺神經電生理領域的深奧,若沒有這麼棒的 指導教授,此刻的我可能還在苦苦掙扎,一段誌謝實遠遠不足表達內心的感謝。
謝謝口試委員:何英剛院士、簡伯武教授、陶寶綠教授、黃玲玲副教授及李立仁 助理教授對本論文內容的建議及指正,使本論文得以順利完成。
感謝實驗室的大夥:欣蓉、阡樺學姐、佳茹、惠萍、思穎、家旭、俐君、淑 方、冠伶、昱征、心慈、宏烈、旅揚、瑞峰、曉筠、騏翔、力威及晉嘉,在這幾 年快以實驗室為家的日子,因為有你/妳們的陪伴,不管是生活上的點滴、學術上 的討論或言不及義的閒扯,都讓研究生活變得絢麗多彩,其中特別感激欣蓉一直 以來的協助及不定時的搞笑放送,也感謝俐君及淑方在染色實驗上的幫忙,謝謝 好友明鋒陪我一起宅、謝謝仕鈞的關心與耀霆學長、范碧娟醫師的鼓勵,謝謝上 知的祝福,還有謝謝曾協助我及鼓勵我的人。
最後要感謝我最深愛的家人及女友,若沒有媽媽一直以來無怨無悔的付出支 撐著家裡,我不可能安心且無後顧之憂的完成學業,早該扛起一切的我現在會加 倍努力回報,謝謝您!也謝謝爸爸的容許,謝謝小弟在我很少回家的這段期間陪 在爸媽身邊,還要謝謝大哥的保佑,感謝親愛的嘉玲這些日子來無悔且勇敢的陪 伴,給予我最大的體諒、鼓勵與支持,幫我打點一切,總是在第一時間充當我的 精神支柱,聽我說著與妳工作領域完全不同的學術用語,有了你們的支持才能成 就這本論文,更是我前進與堅持之動力。
謹以此論文獻給我最摯愛的家人、女友及關心我的朋友們!!
彥昱 僅誌 民國100 年一月
摘要
在1994 年,一種新的鴉片受體被發現,因其結構相似於傳統鴉片受體,但卻
對傳統鴉片受體致效劑缺乏親和力,同時也未發現其內生性致效劑,所以一開始 被稱為類鴉片孤兒受體(opioid receptor like 1, ORL1)。它的內生性致效劑在一年後 被發現並命名為nociceptin 或 orphanin FQ,在 2002 年國際藥理學聯合會以其內生 性致效劑之名正式將此受體命名為 NOP 受體(nociceptin/orphanin FQ peptide receptor)。研究顯示此 N/OFQ-NOP 受體系統與許多神經及精神疾病有關,如疼痛、
焦慮、憂鬱、非自主運動、成癮、發作及痴呆等。
從 NOP 受體基因剪接變異體(splicing variants)被發現及一些受體結合和功能
性研究顯示有 NOP 受體異質性(heterogeneity)的存在。我們之前發現 Ro 64-6198
這個NOP 受體致效劑在中腦環導水管灰質腹側區(ventrolateral PAG)只能活化一部
份的NOP 受體,而在本研究中,我們發現另一個新的非胜肽 NOP 受體結合劑(+)-5a
Compound ((3aS,6aR)-1-(cis-4-Isopropylcyclohexyl)-5'-methyl-2'–phenylhexahydro spiro[piperidine-4,1'-pyrrolo[3,4-c]pyrrole])在相同的標本也只能活化一部份 NOP 受 體,這些可被(+)-5a Compound 活化的 NOP 受體與 Ro 64-6198 所活化的 NOP 受體 是屬於同一群,(+)-5a Compound (0.1-30 μM)能濃度相依的透過 NOP 受體活化 35%
所記錄神經細胞的G 蛋白偶合向內整流鉀離子通道(GIRK channels),其 IC50為605 nM,效價(potency)及所能引發反應的最大效力(efficacy)分別為 N/OFQ 的 1/12 倍及 47%,對(+)-5a Compound-insensitive 的細胞,給予 Ro 64-6198 同樣也無法引發反
應,但在這些細胞中N/OFQ 則可有效的活化 G 蛋白偶合向內整流鉀離子通道,另
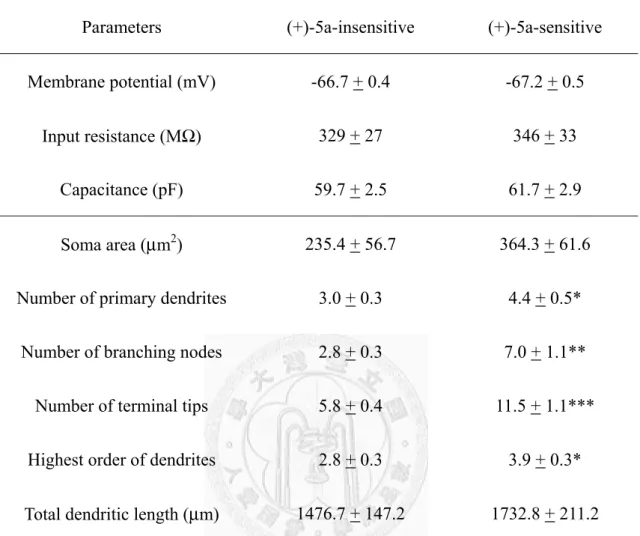
外在(+)-5a Compound-sensitive 的細胞,給予 Ro 64-6198 則反應程度並無差異,從 免疫螢光染色的實驗顯示(+)-5a Compound-sensitive 的細胞主要是 GABAergic,此 外 ,(+)-5a Compound-sensitive 的 細 胞 在 型 態 上 , 其 神 經 分 支 比 (+)-5a Compound-insensitive 的細胞來得複雜。
N/OFQ(1-11)是具藥理活性的 N/OFQ 代謝產物,從放射線標定的研究中,
125I-[Tyr10]N/OFQ(1-11)及 125I-[Tyr14]N/OFQ 這兩個 NOP 受體的放射線結合劑在鼠 腦 中 有 不 同 的 分 布 , 其 中 125I-[Tyr10]N/OFQ(1-11) 的 結 合 位 置 可 能 是
125I-[Tyr14]N/OFQ 所顯現出兩種不同親和力結合處中的高親和力結合處,基於這個
發現,我們於是做了在環導水管灰質區對 Ro 64-6198/(+)-5a Compound 有反應的
NOP 受體就是125I-[Tyr10]N/OFQ(1-11)這個結合位置的假設,因此,我們藉由測試 [Tyr10]N/OFQ(1-11)能否區別(+)-5a Compound-sensitive 及-insensitive 細胞來驗證這 個假設。在紀錄的環導水管灰質腹側區細胞中,[Tyr10]N/OFQ(1-11)在濃度 3-300 μM
時能引發向內整流性鉀離子電流,其EC50為9.0 μM,比 N/OFQ 弱了 173 倍,而
效力(efficacy)則與 N/OFQ 相似,[Tyr10]N/OFQ(1-11)顯露了與(+)-5a Compound 不 同的藥理特性,它不管在(+)-5a Compound-sensitive 或-insensitive 細胞中都有作用,
這些結果顯示 [Tyr10]N/OFQ(1-11)是一個效力比 N/OFQ 差的 NOP 受體全效致效
劑,而且在環導水管灰質腹側區細胞中,[Tyr10]N/OFQ(1-11)無法顯露出 NOP 受體
功能異質性。
為了闡明NOP 受體多樣性的可能,發展及鑑定新的 NOP 受體拮抗劑是很重
要的。因此在本研究中,我們也在中腦環導水管灰質腹側區細胞中測試Compound
24 (1-Benzyl-N-[3-[spiroisobenzofuran-1(3H),4’-piperidin-1-yl]propyl]pyrrolidine-2 -carboxamide)及 SB-612111 ((-)-cis-1- Methyl-7-[[4-(2,6- dichlorophenyl)piperidin-1 -yl]methyl]-6,7,8,9-tetrahydro-5H-benzocyclohepten-5-ol)這兩個新發展的 NOP 受體
拮抗劑是否具有能顯露出NOP 受體功能異質性的特性,然而在我們所紀錄到可被
N/OFQ 所活化的全部細胞,N/OFQ 的作用都會被 Compound 24 及 SB-612111 所拮
抗,因此這兩種NOP 受體拮抗劑並無法顯露 NOP 受體功能異質性。Compound 24
(0.3-10 μM)能濃度相依的抑制 N/OFQ 所誘導出的向內整流鉀離子電流,其 IC50為
2.6 μM,Compound 24 在環導水管灰質腹側區神經細胞的抑制效價比在小鼠輸精管
標本及表現人類NOP 受體的培養細胞所得到的效價還差,而且在相同的環導水管
灰質區標本,Compound 24 的作用比另一個胜肽類 NOP 受體拮抗劑 UFP-101 還要
慢,當Compound 24 濃度高達 10 μM 時對 NOP 受體並不具有內在致效劑的活性,
不過只要 Compound 24 的濃度高於 3 μM 時也會抑制 mu 鴉片受體致效劑
DAMGO(D-Ala2, N-Me-Phe4, Gly5-ol]-enkephalin)所誘導出的向內整流鉀離子電
流。而SB-612111 也能濃度相依的拮抗 N/OFQ 所引發的向內整流鉀離子電流,從
濃度反應曲線估出其IC50值為87.7 nM,而且 SB-612111 就算濃度高到 1 μM 並不
會產生內在致效劑活性,也不會影響 mu 鴉片受體致效劑 DAMGO 所引發的向內
整流鉀離子電流。
總結,在環導水管灰質腹側區(+)-5a Compound 與 Ro 64-6198 一樣都只活化
一部份的 NOP 受體,而這些對(+)-5a Compound 有反應的神經細胞大多屬於
GABAergic,而且對(+)-5a Compound 有反應及無反應的細胞,兩者在細胞型態上 並不相同,這些結果進一步支持在中腦環導水管灰質區的 NOP 受體有功能異質性 的存在。不過[Tyr10]N/OFQ(1-11)在藥理特性上與(+)-5a Compound 並不相同,而且 利用[Tyr10]N/OFQ(1-11)以及 Compound 24 與 SB-612111 這兩個新的 NOP 受體拮抗
劑也無法觀察到 NOP 受體功能異質性的現象。此外,在Compound 24 與 SB-612111
的定量性研究上,證明Compound 24 是一個效價普通且選擇性亦非理想的 NOP 受
體拮抗劑,另一方面,SB-612111 則是一個有效且具選擇性,同時又具非胜肽類優
勢的NOP 受體拮抗劑,它可說是能用來探究內生性 N/OFQ 生理角色最好的 NOP
受體拮抗劑。
關鍵詞:NOP 受體、環導水管灰質區、G 蛋白偶合向內整流鉀離子通道、鴉片受 體、異質性
Abstract
In 1994, a novel opioid receptor family, opioid receptor like 1 (ORL1) orphan receptors, was identified to be structurally similar to classical opioid receptors, but insensitive to traditional opioids. Its endogenous ligand was later identified to be a heptadecapeptide and termed nociceptin or orphanin FQ. This receptor was consensusly renamed after its endogenous ligand as nociceptin/orphanin FQ (N/OFQ) peptide (NOP) receptor in IUPHAR2002. The N/OFQ-NOP receptor system was implicated in several neurological and psychological disorders, such as pain, anxiety, depression, involuntary movement, addiction, seizure and dementia.
Heterogeneity of NOP receptors has been proposed based on the findings of splicing variants and from binding and functional studies. We have previously reported that Ro 64-6198, a NOP receptor agonist, activated a subset, but not all, of N/OFQ-sensitive NOP receptors in midbrain ventrolateral periaqueductal gray (vlPAG). In this study, we found that a new non-peptide ligand of NOP receptors, (+)-5a Compound ((3aS,6aR)-1-(cis-4-Isopropylcyclohexyl)-5'-methyl-2'-phenylhexahydrospiro[piperidine -4,1'-pyrrolo[3,4-c]pyrrole]), also activated a subset of NOP receptors as the same subset affected by Ro 64-6198 in vlPAG neurons. (+)-5a Compound (0.1-30 μM) concentration-dependently activated G-protein-coupled inwardly rectifying potassium (GIRK) channels through the NOP receptors in about 35% of the recorded vlPAG neurons. (+)-5a Compound (EC50: 605 nM) was less potent (1/12) and efficacious (47%) than N/OFQ. In (+)-5a Compound-insensitive neurons, Ro 64-6198 was also ineffective, and vice versa, but N/OFQ was effective in activating GIRK channels through NOP receptors. In (+)-5a Compound-sensitive neurons, (+)-5a Compound precluded the effect of Ro 64-6198. Immunofluorecent and morphometric studies showed that most of the (+)-5a Compound-sensitive neurons were multipolar with intensive dendritic arborization and immunoreactive to glutamic acid decarboxylase-67.
N/OFQ(1-11) is a pharmacologically active metabolite of N/OFQ. The distribution of its radioligand, 125I-[Tyr10]N/OFQ(1-11), resembled the high affinity, but not all, binding sites of 125I-[Tyr14]N/OFQ in rodent brains. Based on this finding we hypothesize that the Ro 64-6198/(+)-5a Compound-sensitive NOP receptor in the PAG is the binding site of 125I-[Tyr10]N/OFQ(1-11). Here, we validated this hypothesis by examining if [Tyr10]N/OFQ(1-11) can differentiate (+)-5a Compound-sensitive and -insensitive vlPAG neurons. [Tyr10]N/OFQ(1-11), like N/OFQ, induced GIRK current through NOP receptors in the vlPAG neurons. It was 173 times less potent (EC50: 9.0 μM) but equi-efficacious, as compared with N/OFQ. [Tyr10]N/OFQ(1-11) displayed different pharmacological profiles as (+)-5a Compound. It was effective in both (+)-5a Compound-sensitive and -insensitive neurons. These results suggest that [Tyr10]N/OFQ(1-11) is an NOP full agonist and less potent than N/OFQ. The functional heterogeneity of NOP receptors, therefore, can not be revealed by [Tyr10]N/OFQ(1-11) in vlPAG neurons.
To clarify the possible diversity of NOP receptors, it is important to develop and characterize novel NOP receptor antagonists. In this study, we also examined if two newly developed NOP receptor antagonists, 1-Benzyl-N-[3-[spiroisobenzofuran -1(3H),4’-piperidin-1-yl]propyl]pyrrolidine-2-carboxamide (Compound 24) and (-)-cis-1-Methyl-7-[[4-(2,6-dichlorophenyl)piperidin-1-yl]methyl]-6,7,8,9-tetrahydro-5 H-benzocyclohepten-5-ol (SB-612111), can reveal the heterogeneity of NOP receptors in vlPAG slices. Both Compound 24 and SB-612111 antagonized the effect of N/OFQ in all the recorded neurons. Therefore, the heterogeneity of NOP receptors can not be revealed by these two antagonists. However, we further quantitatively characterized their interactions with N/OFQ in vlPAG slices. Compound 24, at 0.3-10 μM, attenuated N/OFQ-induced GIRK current concentration-dependently. The antagonistic potency of Compound 24 in vlPAG neurons (IC50: 2.6 + 0.6 μM) was, however, lower than that
obtained in mouse vas deferens preparations or expressed human NOP receptors. The action kinetic of Compound 24 was slower than UFP-101, a peptide antagonist, in the same preparations. Compound 24 had no intrinsic agonistic activity at NOP receptors at concentrations up to 10 μM. However, at concentrations higher than 3 μM, it also attenuated the GIRK current induced by DAMGO ([D-Ala2, N-Me-Phe4, Gly5-ol]-enkephalin), a mu-opioid receptor agonist. As for SB-612111, it also concentration-dependently antagonized N/OFQ-induced GIRK current in vlPAG neurons. The IC50 value of SB-612111 estimated from concentration-response curve is 87.7+1.2 nM. SB-612111 had no intrinsic agonistic activity and did not affect the GIRK current induced by DAMGO when tested at concentrations of up to 1 μM.
In conclusion, (+)-5a Compound activates a subset of NOP receptors, similar to the Ro 64-6198-sensitive subset, in vlPAG neurons which are mostly GABAergic.
Moreover, (+)-5a Compound-sensitive or -insensitive neurons are morphologically distinct. These results further support the presence of functional heterogeneity of NOP receptors in the PAG. However, the pharmacological profiles of [Tyr10]N/OFQ(1-11) are unlike that of (+)-5a Compound and the functional heterogeneity of NOP receptors can not be revealed by either [Tyr10]N/OFQ(1-11) or new NOP receptor antagonists, Compound 24 and SB-612111. Quantitative studies of Compound 24 and SB-612111 in the vlPAG showed that Compound 24 acts as a competitive NOP receptor antagonist but its potency and selectivity are moderate. On the other hand, SB-612111 is a pure, potent and selective antagonist of NOP receptors with the merits of non-peptide nature, high potency, and selectivity. SB-612111 is the best NOP receptor antagonist available for exploring the functional roles of endogenous N/OFQ.
Key words:Nociceptin/orphanin FQ Receptor; Periaqueductal Gray; Potassium channel;
(+)-5a Compound; [Tyr10]N/OFQ(1-11); Compound 24; SB-612111.
Table of Contents
Page 口試委員審定書 ... I 誌謝 ... II
中文摘要 ………... III
英文摘要 ……….. IV
Abbreviation ... 1
Introduction ………. 2
1. N/OFQ and NOP receptor ……….. 2
2. NOP receptor ligands ………. 3
2.1. NOP receptor agonist ………... 4
2.1.1. Peptide NOP receptor agonist ……… 4
2.1.2. Non-peptide NOP receptor agonist ……… 5
2.1.2.1. Ro 64-6198 ……….. 7
2.1.2.2. (+)-5a Compound ……… 8
2.2. NOP receptor antagonist ………. 9
2.2.1. Peptide NOP receptor antagonist ……….. 9
2.2.2. Non-peptide NOP receptor antagonist ……….. 10
2.2.2.1. Compound 24 ……….. 12
2.2.2.2. SB-612111 ……… 13
3. Physiological or pathological roles of endogenous N/OFQ ……….. 14
4. Pain ……… 15
4.1. Ascending pain pathway ………. 16
4.2. Descending inhibition pain pathway ……….. 16
4.3. Effects of N/OFQ on pain ……….. 17
4.3.1. Supraspinal pain regulation of N/OFQ ……… 17
4.3.2. Spinal pain regulation of N/OFQ ………. 19
5. Periaqueductal gray (PAG) ……… 20
5.1. Roles of N/OFQ in the PAG ……… 21
6. Heterogeneity of NOP receptors ……… 22
6.1. Splicing variants of NOP receptors ……… 22
6.2. Binding studies with 125I-[Tyr14]N/OFQ ……… 23
6.3. Autoradiographic studies with 125I-[Tyr10]N/OFQ(1-11) and 125I-[Tyr14]N/OFQ ……… 23
6.4. Functional heterogeneity of NOP receptor revealed from the results of Ro 64-6198 ……… ………. 24
Aim of study………. 26
Materials and methods ……….. 29
1. Brain slice preparations ……… 29
2. Electrophysiological recordings ……… 29
3. Quantitative analysis of NOP receptor ligands ………. 30
3.1. NOP receptor agonists ……….. 30
3.2. NOP receptor antagonists ……….. 32
4. Immunofluorescence staining ……… 33
5. Morphometric analysis ………. 34
6. Chemicals ……….. 35
7. Statistics ……… 36
Results ………. 37
1. Functional heterogeneity of NOP receptors revealed by (+)-5a Compound.. 37
1.1. (+)-5a Compound activated GIRK channels in vlPAG neurons ………. 37
1.2. (+)-5a Compound was less potent and less efficacious than N/OFQ ….. 37 1.3. (+)-5a Compound activated GIRK channels in about half of the
recorded neurons ………. 38
1.4. The effect of (+)-5a Compound was antagonized by NOP, but not
opioid or M2 muscarinic, receptor antagonists ……… 39 1.5. N/OFQ activated GIRK channels via NOP receptors in (+)-5a
Compound-insensitive vlPAG neurons ……… 40 1.6. (+)-5a Compound-insensitive neurons were also insensitive to Ro
64-6198, and vice versa ………... 41 1.7. (+)-5a Compound precluded the effect of Ro 64-6198 ……… 42 1.8. N/OFQ further enhanced GIRK current in (+)-5a Compound-sensitive
neurons ………. 42
1.9. Most of the (+)-5a Compound-sensitive neurons were GABAergic …… 43 2. Quantitative study of [Tyr10]NC(1-11) in vlPAG ………... 45 2.1. [Tyr10]N/OFQ(1-11) activated inwardly rectifying potassium channels... 45 2.2. [Tyr10]N/OFQ(1-11) was similar efficacious, but less potent than ,
N/OFQ ……….. 45
2.3. The effect of [Tyr10]N/OFQ(1-11) was antagonized by UFP-101 but not
naloxone ………... 46
2.4. [Tyr10]N/OFQ(1-11) further enhanced GIRK current in (+)-5a
Compound-sensitive neurons ……….. 47 2.5. [Tyr10]N/OFQ(1-11) induced GIRK currents in (+)-5a
Compound-insensitive neurons ……… 48 2.6. [Tyr10]N/OFQ(1-11) precludes the effect of N/OFQ ……… 49 3. Quantitative study of Compound 24 in vlPAG ……….. 49
3.1. N/OFQ activated GIRK channels in vlPAG neurons ………... 49
3.2. Compound 24 antagonized the effect of N/OFQ concentration -dependently ………... 50
3.3. Compound 24 reduced DAMGO-induced GIRK current at higher concentrations ……….. 51
3.4. Compound 24 had no effect on membrane current per se ………... 52
4. Quantitative study of SB-612111 in vlPAG ………... 53
4.1. SB-612111 antagonized the effect of N/OFQ in a concentration -dependently manner ……… 53
4.2. SB-612111 did not affect the membrane current per se ………... 53
4.3. SB-612111 did not affect DAMGO-induced GIRK current ……… 54
Discussion ………. 56
1. Functional heterogeneity of NOP receptors ………... 56
1.1. Functional heterogeneity of NOP receptors revealed by (+)-5a Compound and Ro 64-6198 ………. 56
1.1.1. (+)-5a Compound, like Ro 64-6198, is less potent and efficacious than N/OFQ in the vlPAG ………. 56
1.1.2. (+)-5a Compound activates GIRK channels through NOP, but not opioid-, σ-, H3-histamine or M2-muscarinic, receptors ……… 57
1.1.3. (+)-5a Compound activates a subset but not all of NOP receptors in vlPAG neurons ………... 58
1.1.4. (+)-5a Compound and Ro 64-6198 activate subset of NOP receptors in vlPAG neurons ……….. 59
1.1.5. A majority of the (+)-5a Compound-sensitive neurons are GABAergic ……… 59
1.2. Functional heterogeneity of NOP receptors can not be revealed by
[Tyr10]N/OFQ(1-11) ……… 60
1.2.1. [Tyr10]N/OFQ(1-11) activates GIRK mediated by NOP but not MOP receptors ……….. 60
1.2.2. [Tyr10]N/OFQ(1-11) is a full agonist of NOP receptor and less potent than N/OFQ ……… 61
1.2.3. [Tyr10]N/OFQ(1-11) affects both (+)-5a Compound-sensitive and -insensitive vlPAG neurons ………. 62
1.3. NOP receptor heterogeneity can not be differentiated by NOP receptor antagonist ……… 63
1.4. Heterogeneity of NOP receptors ……… 64
2. Pharmacological characterization of two NOP receptor antagonists, Compound 24 and SB-612111, in vlPAG neurons ……… 67
2.1. Compound 24 possessed moderate antagonist potency at native NOP receptors in PAG ………. 67
2.2. Compound 24 exhibiting moderate selectivity ……… 67
2.3. SB-612111 with potent antagonist activity at NOP receptor ……… 68
2.4. SB-612111 is devoid of intrinsic activity ………. 69
2.5. SB-612111 displaying highly selectivity for NOP receptors …………... 70
2.6. Development of NOP receptor antagonists ……….. 70
Conclusion ……… 72
References ……… 73
Bibliography ……… 93
Tables and Figures ………... 94
Abbreviation
ABC avidin-biotin-peroxidase complex
ACC anterior cingulated cortex
aCSF artificial cerebral spinal fluid BSA bovine serum albumin
DAB 3,3’-diaminobenzidine DAMGO [D-Ala2, N-Me-Phe4, Gly5-ol]-enkephalin
DMSO dimethyl sulfoxide
GAD67 glutamic acid decarboxylase-67 GABA γ-amino-butyric acid
GIRK G-protein coupled inwardly rectifying potassium channels GPCR G-protein coupled receptor
H2O2 hydrogen peroxide
I-140 membrane current at -140 mV
Ihold holding current
I-V current-voltage
LY lucifer yellow
NalBzOH naloxone benzoylhydrazone
N/OFQ nociceptin/orphanin FQ
NOP nociceptin/orphanin FQ peptide ORL1 opioid receptor-like 1
PAG periaqueductal gray
PBS phosphate buffered saline RVM rostral ventromedial medulla vlPAG ventrolateral periaqueductal gray
Introduction 1. N/OFQ and NOP receptor
An orphan G-protein coupled receptor (GPCR) was cloned in 1994 from a search to discover subtypes of opioid receptors and named initially as opioid receptor like 1 (ORL1), because, although having a structure highly homologous to classical opioid receptors, it displayed little affinity for opioids (Mollereau et al., 1994). One year later, this orphan receptor was de-orphanized through identifying its endogenous ligand, a heptadecapeptide named nociceptin (Meunier et al., 1995) or orphanin FQ (Reinscheid et al., 1995). The ORL1 receptor was classified as the fourth member of the opioid receptor family and renamed as nociceptin/orphanin FQ (N/OFQ) peptide (NOP) receptor. It is classified as a non-opioid branch of the opioid receptor family (NC-IUPHAR, 2004).
N/OFQ is derived from a precursor protein, preproN/OFQ (ppN/OFQ) which consists of 181, 187 and 176 amino acids, respectively, in the rat, mouse and human (Nothacker et al., 1996). The intracellular events triggered by the binding of N/OFQ with the NOP receptor, and mediated by the activation of G-protein, include inhibition of adenylyl cyclase, activation of phospholipase C and K+ channels and inhibition of Ca2+ channels.
The regional distribution of N/OFQ and the NOP receptor have been well
described. Regions with high levels of NOP receptor include the cerebral cortex, hypothalamus, hippocampus, amygdale, periaqueductal gray, substantia nigra, central gray, locus coeruleus and spinal cord (Anton et al., 1996; Berthele et al., 2003; Florin et al., 1997; Neal et al., 1999a). The localization of N/OFQ corresponds reasonably well with the NOP receptor. As with the receptor, N/OFQ immunoreactivity and mRNA levels detected using in situ hybridization are closely correlated. N/OFQ is found in ventral forebrain, hypothalamus, hippocampus, amygdala, ventral tegmentum, periaqueductal gray, substantia nigra, central gray, locus coeruleus, spinal cord (Boom et al., 1999; Neal et al., 1999b; Nothacker et al., 1996; Witta et al., 2004).
2. NOP receptor ligands
When the NOP receptor was cloned initially, several commonly used opioid drugs including etorphine and buprenorphine have been demonstrated to bind to NOP receptors (Wnendt et al., 1999). However, their binding affinities at the NOP receptors are relatively insignificant compared to their activity at classical opioid receptors.
Thereafter, a wide range of selective agonist and antagonist ligands for NOP receptors have been developed, which show little or no affinity to classical opioid receptors and so allow NOP receptor mediated responses to be studied in isolation (Chiou et al., 2007).
2.1. NOP receptor agonist
2.1.1. Peptide NOP receptor agonist
Since N/OFQ was identified to be a NOP receptor agonist, several peptide-based NOP receptor agonists have been reported (Table 1). The shortest N/OFQ derivative possessing the same affinity for NOP receptor as N/OFQ is the truncated and amidated N/OFQ(1-13)-NH2 (Guerrini et al., 1997). C-terminal amidation protects from degradation by carboxypeptidases and is now a standard feature of most N/OFQ-based peptide ligands. Substituting the 14th and 15th residues with Arg and Lys, respectively, gave an agonist, [Arg14, Lys15]N/OFQ. It was reported to be the first N/OFQ derivative with higher affinity and biological potency in vitro than N/OFQ (Okada et al., 2000;
Rizzi et al., 2002b). Conformationally restricted peptide synthesis also generated an agonist, cyclo[Cys10, Cys14]N/OFQ, with comparable affinity to N/OFQ (Ambo et al., 2001). The cyclic peptide is the first conformationally restricted peptide with potent activity and may serve as a good template for studying the bioactive conformation of N/OFQ. [(pF)Phe4, Arg14, Lys15]N/OFQ-NH2 (UFP-102) appears to be a more promising one with higher potency and longer duration of action (Carra et al., 2005).
Substituting the 7th amino acid residue of UFP-102 with aminoisobutyric acid (Aib), a more potent agonist, UFP-112, of NOP receptors (Arduin et al., 2007) was developed. It behaves as a selective full agonist, producing long lasting effect in vivo. The
tetrapeptide OS-461 and its analogues, OS-462 and OS-500, developed by Nippon Shinyaku (Economidou et al., 2006b) were reported to have NOP agonistic activity and analgesic effects in mice. In 2007, a truncated cyclic analogue with cysteine residues at positions 7 and 10, c[Cys(7,10)]N/OFQ(1-13)-NH2, was developed to improve the metabolic stability (Kitayama et al., 2007). Although it acts as a high-potency full agonist of NOP recetpors, the expected metabolic enzymatic protection was not achieved.
2.1.2. Non-peptide NOP receptor agonist
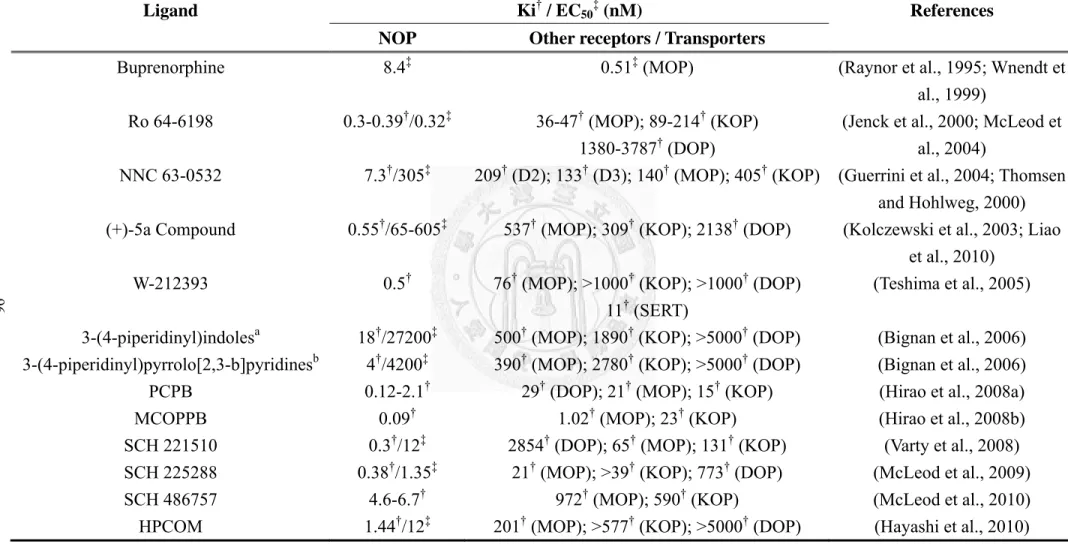
Buprenorphine and lofentanil are a few classic opioid drugs having agonistic activity at NOP receptors (Hawkinson et al., 2000). Ro 64-6198 is the first synthetic non-peptide agonist presented by F. Hoffmann-La Roche Ltd. in 2000 (Jenck et al., 2000). It was widely used to characterize the functional roles of the N/OFQ-NOP system. In 2003, F. Hoffmann-La Roche Ltd. developed another non-peptide agonist, (+)-5a Compound, with a structure backbone analogous to Ro 64-6198 (Kolczewski et al., 2003) (See the section 2.1.2.2). NNC 63-0532 is a spiroxatrine analogue with high affinity for NOP receptors but only 12-fold selectivity for NOP receptors as compared with MOP receptors (Thomsen and Hohlweg, 2000). In 2005, a potent non-peptide NOP receptor full agonist, W-212393, was development by Mitsubishi Pharma (Teshima et
al., 2005). This compound is fairly selective versus other opiate receptors, but also exhibits nanomolar affinity for the serotonin transporter. In 2006, Bignan et al. (2006) at Johnson & Johnson Company presented novel piperidin-indoles and pyrrolo-pyridines as NOP receptor agonists with low nanomolar affinity and more than 25-100 times selectivity versus other opioid receptors. In 2008, Hirao et al. (2008a) at Pfizer Company revealed two brain penetrating non-peptide agonists, PCPB and MCOPPB, which have subnanomolar affinity for NOP. In 2008, Varty et al. at Schering-Plough Company developed an NOP receptor agonist, SCH 221510 (Varty et al., 2008). It produced robust anxiolytic-like activity at doses that do not seem to produce nonspecific disruption of overt behaviors such as locomotor activity and did not tolerate after chronic dosing. In 2009, the same company developed another NOP receptor agonist, SCH 225288 (McLeod et al., 2009) with good oral absorption rate and effective in several cough models, including capsaicin-induced coughs in guinea pigs, and mechanically-evoked coughs in cats and infectious tracheobronchitis-induced coughs in canines. SCH 486757 (McLeod et al., 2010) is another orally active NOP receptor agonist from the same company. It has higher selectivity to NOP receptors than SCH 225288 and produces a level of antitussive efficacy equivalent to codeine in capsaicin-evoked cough model without the side effect liabilities associated with opioid antitussives. In addition, it does not show potential to induce abuse liabilities. This
compound has recently entered human clinical trials for cough treatment. In 2010, Pfizer Company also presented a new NOP receptor agonist, HPCOM (Hayashi et al., 2010). It showed potent affinity, high selectivity and long-lasting metabolic stability.
When administered by s.c. injection, it had high plasma concentration with high plasma-to-brain selectivity (s.c.), and exhibited good antiallodynic activity on injured paw in neuropathic pain model rats derived from peripheral-selective antiallodynic activity. These non-peptide NOP receptor agonists are shown in Table 2. Among these, Ro 64-6198 is the first developed NOP receptors and has been widely used to elucidate the function of NOP receptors and determine the potential of NOP receptor as a therapeutic target, especially in anxiety. Our laboratory also had contribution in this field using this compound (Chiou et al., 2004). We used this compound as a reference compound to further explore the effects of (+)-5a Compound. These two lignads are specially introduced in the followings.
2.1.2.1. Ro 64-6198
Ro 64-6198 (Fig. 1) was shown initially to have subnanomolar affinity and high selectivity for NOP receptors (Jenck et al., 2000). Functional studies showed that Ro 64-6198 acted as a potent NOP receptor agonist in CHOhNOP cells and peripheral and brain preparations (Chiou et al., 2004; Gehlert et al., 2006; Jenck et al., 2000; Rizzi et al., 2001; Wichmann et al., 2000). Ro 64-6198 has been used in several studies to reveal
the functional roles and nature of NOP receptor. In most studies, Ro 64-6198 mimicked the effects of N/OFQ, including memory impairment (Goeldner et al., 2008; Higgins et al., 2002; Kuzmin et al., 2009), high-dose N/OFQ-induced hypolocomotion (Kuzmin et al., 2004) and hyperphagia (Ciccocioppo et al., 2002), reduction of alcohol-induced conditioned place preference (Kuzmin et al., 2003), antitussion (McLeod et al., 2002) and orexigenic effect (Economidou et al., 2006a). However, Ro 64-6198 did not mimick all of the effects of N/OFQ in several functional studies (See the section 6.4). This raises the possibility that Ro 64-6198 and N/OFQ may activate a different functional subset of NOP receptors.
2.1.2.2. (+)-5a Compound
(+)-5a Compound ((3aS,6aR)-1-(cis-4-isopropylcyclohexyl)-5’-methyl-2’
-phenylhexahydrospiro[piperidine-4,1’-pyrrolo[3,4-c]pyrrole]) (Fig. 1) acts as a full agonist of NOP receptors and is 1.7- and 25-fold less potent than N/OFQ in expressed hNOP receptor-mediated GTPγS binding and cAMP reduction, respectively (Kolczewski et al., 2003). Importantly, (+)-5a Compound is more selective for NOP receptors over other opioid receptors than Ro 64-6198, with overall >1000-fold binding selectivity (Kolczewski et al., 2003).
2.2. NOP receptor antagonist
2.2.1. Peptide NOP receptor antagonist
Several peptide NOP receptor antagonists have been developed so far (Table 3).
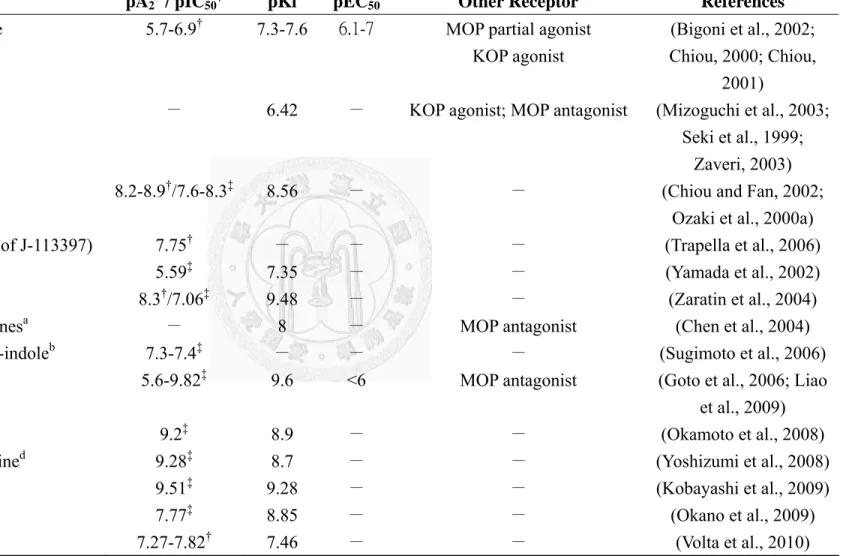
However, early developed peptide antagonists are flawed by their non-specificity, such as the conformational restricted peptide III-BTD or by their residual agonist activity, such as acetylated hexapeptides (Ac-RYYRIK-NH2 and Ac-RYYRWK-NH2) and the first proposed antagonist, [Phe1ψ(CH2-NH)Gly2]N/OFQ(1-13)-NH2 (Chiou et al., 2007).
ZP-120 (Ac-RYYRWKKKKKK-NH2) acted as a partial agonist at NOP receptor with higher potency and longer duration than N/OFQ. Its blood-brain barrier impermeable merit makes it useful in improving cardiovascular function without central effects (Kapusta et al., 2005). [Nphe1]N/OFQ(1-13)-NH2 is the first antagonist demonstrated to antagonize N/OFQ-effects but have no intrinsic activity at NOP receptors (Calo' et al., 2000b). Several physiological or pathological roles of N/OFQ have thereafter been revealed by using this pure and selective antagonist. [Nphe1,Arg14,Lys15]N/OFQ-NH2
(UFP-101), derived by a combination of two N/OFQ-modified peptides, [Arg14,Lys15]N/OFQ and [Nphe1]N/OFQ(1-13)-NH2, is a potent and competitive antagonist (Calo' et al., 2002). UFP-101 has been proven to be a more potent antagonist than [Nphe1]N/OFQ(1-13)-NH2 and have a longer duration in vivo. It has become a useful pharmacological tool available for the investigation of the central and peripheral
biological functions regulated by the N/OFQ-NOP receptor system and for defining the therapeutic potential of NOP receptor ligands. In 2006, Gunduz et al. synthesized a potent hexapeptide alcohol, Ac-RYYRIK-ol, by substituting the C-terianl amide of Ac-RYYRIK-NH2 with an alcoholic function (Gunduz et al., 2006). However, tt displayed a complex pharmacological profile which is likely due to the low efficacy agonistic activity. Recently, a novel NOP receptor antagonist, Ac-RYYRIR-ol, representing the Lys6 to Arg6 replacement of its parent compound, Ac-RYYRIK-ol, had been synthesized (Bojnik et al., 2010). It also displays a complex pharmacological profile but do not show expected higher binding affinity than Ac-RYYRIK-ol.
2.2.2. Non-peptide NOP receptor antagonist
Putative non-peptide NOP receptor antagonists are summarized in Table 4.
Naloxone benzoylhydrazone (NalBzOH) is a non-selective opioid receptor ligand, which was reported to competitively antagonize the effects of N/OFQ before any specific antagonist has been developed (Noda et al., 1998). A morphinan derivative, TRK-820, has been reported to interact with NOP receptor as an antagonist (Seki et al., 1999). This compound acts as a high affinity KOP agonist, whereas very high concentrations are required to reverse the actions of N/OFQ. In 1999, the first non-peptide antagonist selective for NOP receptor, J-113397, was developed by Banyu
Pharmaceutical in Japan. It is a very potent and selective antagonist of NOP receptors without intrinsic activity (Chiou and Fan, 2002; Ozaki et al., 2000b). However, it was later found to have psychomimetic central activity (Koizumi et al., 2004) which is likely attributed to its affinity at σ receptors (Chiou et al., 2007). Later, Calo’s group at University of Ferrara (Italy) synthesized a high yield achiral analogue of J-113397, termed Trap-101 (Trapella et al., 2006), which is equipotent to enatiomer J-113397. In 2000, Japan Tobacco Inc. presented a non-peptide quinoline antagonist, JTC-801 (Shinkai et al., 2000), which binds to the NOP receptor with high affinity in vitro, yet only about 10-fold selectivity over the MOP receptor. In 2004, a potent and selective non-peptide antagonist, SB-612111, was reported by GlaxoSmithKline Pharmaceuticals (See section 2.2.2.2). In 2006, Goto et al at Banyu Pharmaceutical developed several structural diverse analogs of spiropiperidines as NOP receptor antagonists, among these, Compound 24 is a potent one (See the next paragraph). The same company kept developed several NOP antagonists with different chemical cores, including the benzimidazole derivatives (compound 28 is the most potent one in [35S]GTPγS binding assay) (Okamoto et al., 2008), N-biarylmethyl spiropiperidines (compound 37 is a potent one) (Yoshizumi et al., 2008) and structureal diverse analogs of arylpyrazole (compound 31 is the most potent one) (Kobayashi et al., 2009). Recently, Volta et al in University of Ferrara (Italy) modified Trap-101 structure and introduced two methyl
groups in the hydroxymethyl function at the carbon 3 of the piperidine ring, thus obtaining a novel non-peptide antagonist, GF-4 (Volta et al., 2010). Such chemical modification did not alter the pharmacological activity of the compound (a pure and competitive NOP receptor antagonist) but slightly reduced its potency at recombinant NOP receptors and, more dramatically, in vitro selectivity over classical opioid receptors.
In this study, we have the gifts of Compound 24 and SB-612111 from international collaboratory laboratories. Their detailed informations are introduced in the followings.
2.2.2.1. Compound 24
Compound 24 (1-Benzyl-N-[3-spiro[isobenzofuran-1(3H),4’-piperidine]-1-yl]
propyl-D-proline amide) (Fig. 1) is a spiropiperidine analogue of NOP receptor antagonist developed by Goto et al. (2006). It was reported to have high affinity (Kd: 0.27 nM) at cloned human NOP receptors and be a potent antagonist (IC50: 0.15 nM) in inhibiting GTPγS binding at cloned hNOP receptors expressed in CHO cells. In the in
vivo study, Compound 24 significantly antagonized N/OFQ-induced reduction of
locomotor activity (Goto et al., 2006). It also acted as a compeptitive antagonist in the assays of GTPγS and electrically stiumulated mouse vas deferens contraction (Fischetti et al., 2008).
2.2.2.2. SB-612111
SB-612111, (-)-cis-1-Methyl-7-[[4-(2,6-dichlorophenyl)piperidin-1-yl]methyl]
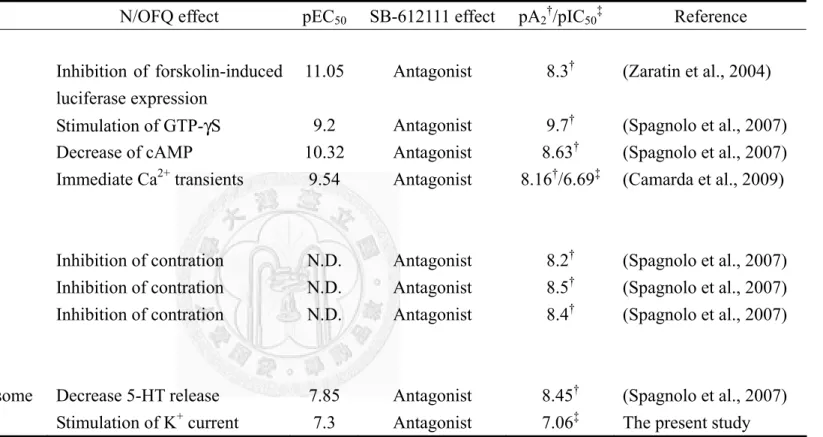
-6,7,8,9-tetrahydro-5H-benzocyclohepten-5-ol, (Fig. 1) is also an NOP receptor antagonist originally developed by GlaxoSmithKline Pharmaceuticals (Zaratin et al., 2004). It exhibits high affinity (Ki: 0.33-1.42 nM) at cloned human NOP receptors expressed in CHO cells and displayed 175-7594 times selectivity to NOP receptors among traditional opioid receptors (Khroyan et al., 2009; Spagnolo et al., 2007; Zaratin et al., 2004). Functional studies showed that SB-612111 acted as a potent antagonist (Kb: 5 nM) in reversing the inhibitory effect of N/OFQ on forskolin-induced luciferase expression (Zaratin et al., 2004). It also attenuated N/OFQ-induced inhibition of cAMP accumulation and GTPγS binding at CHO cell-expressed human NOP receptors, and inhibition of [3H]5-HT overflow in mouse cortical synaptosomes (Spagnolo et al., 2007).
Furthermore, SB-612111 antagonized the inhibitory effect of N/OFQ on electrically stimulated contractions in mouse vas deferens and guinea pig ileum preparations (Spagnolo et al., 2007). In GTPγS binding, cAMP accumulation and rat vas deferens contraction assays, SB-612111 did not display intrinsic activity at NOP receptors (Spagnolo et al., 2007). It also effectively antagonized immediate calcium transients evoked by N/OFQ in CHOhNOP cells stably expressing the chimeric Gαqi5 protein (Camarda et al., 2009b). SB-612111 also antagonized the effects of N/OFQ in vivo,
including the hyperalgesia and the reversal of morphine tolerance induced by N/OFQ in the mouse hot plate test (Zaratin et al., 2004), the antinociceptive and the pronociceptive actions of N/OFQ given by i.t. and i.c.v. injections, respectively, in the tail-flick test (Rizzi et al., 2007), and the orexigenic effect of N/OFQ (Rizzi et al., 2007). SB-612111 also potentiated the antinociception induced by buprenorphine and other mixed NOP/MOP receptor agonists, but not by selective MOP receptor agonists, in mouse tail-flick test (Khroyan et al., 2009). Several in vivo studies reported that SB-612111 alone did not affect the nociceptive response in the mouse hot-plate test when given by
i.c.v. injection (Zaratin et al., 2004) or in the tail-flick test when given by i.p. (Rizzi et
al., 2007) or s.c. (Khroyan et al., 2009) administration. However, SB-612111 (i.v.) effectively reversed the thermal hyperalgesia in a rat carrageenan inflammatory model (Zaratin et al., 2004). It (i.p.) also induced a depressant-like effect in the mouse forced swimming test in a manner reversed by N/OFQ, and this effect was inactive in NOP receptor knockout mice (Rizzi et al., 2007). The latter two studies might suggest that endogenous N/OFQ has pronociceptive and antidepressant functions.
3. Physiological or pathological roles of endogenous N/OFQ
Studies on the role of the endogenous N/OFQ in physiological or pathological functions began from the time of the discovery of the NOP receptor and first relied on
administration of N/OFQ. Several approaches have been conducted in revealing the physiological or pathological roles of endogenous N/OFQ. Before the selective NOP receptor antagonists or transgenic mice were developed, the antisense oligonucleotide against NOP receptor had been applied (Rossi et al., 1997). Knockout mice have been developed without the gene encoding NOP receptors (Nishi et al., 1997) or ppN/OFQ (Kest et al., 2001; Koster et al., 1999). Many physiological or pathological roles of N/OFQ have been reported from animal studies, including pain regulation, anxiety, depression, addiction, feeding, learning and memory, motor control and central cardiovascular regulation (Chiou et al., 2007; Lambert, 2008). N/OFQ also regulates neurotransmitter release in the autonomic nervous system and plays a role peripherally in the pathogenesis of asthma, urine retention or cardiovascular function (Chiou et al., 2007; Lambert, 2008).
4. Pain
Pain generally starts with a physical event, including a cut, burn, tear or bump.
The sensation of pain usually depends on the activation of a set of neurons that includes primary afferent nociceptors, interneurons in the spinal cord, cells of the ascending tracts, thalamic neurons and neurons of the cerebral cortex. Hence, the pain system involves a set of ascending pathways that convey nociceptive information from
peripheral nociceptors to higher levels of the central nervous system, as well as descending pathway that modulate that information (Fig. 2).
4.1. Ascending pain pathway
Nociceptive information is transmitted mainly through the ascending pain pathway.
Primary afferent nociceptors convey noxious information to the secondary sensory neurons within the dorsal horn of the spinal cord which sends projections upwardly. A subset of these projection neurons transmits information to the somatosensory cortex via the thalamus, providing information about the location and intensity of the painful stimulus. Other projection neurons engage the cingulated and insular cortices via connections in the brainstem and amygdale, cortributing to the affective component of the pain experience.
4.2. Descending inhibition pain pathway
The descending inhibition pain pathway is a well-characterized anatomical network that enables us to regulate nociceptive processing. The brain uses this pathway to send chemical substances and nerve impulses back down to the sensory neurons in the spinal cord to act against the pain message sent up by the pain receptors. The midbrain PAG, of pain-inhibiting circuitry is a crucial part, is best known and
contributes to opiate analgesia. In addition to PAG, a variety of brain regions are involved in this descending modulation, including the frontal lobe, anterior cingulated cortex (ACC), amygdale, hypothalamus and rostral ventromedial medulla (RVM).
Figure 2 illustrates the key anatomical features of the descending inhibition pain pathway.
4.3. Effects of N/OFQ on pain
The sequence similarities between NOP and classical opioid receptors drive the studies that investigate the roles of endogenous N/OFQ focusing on pain regulation.
N/OFQ shares similar cellular actions with opioids, such as inhibition of cAMP formation, activation of K+ channels and inhibition of Ca2+ channels (Darland et al., 1998). However, different from conventional opioids that usually produce analgesia, N/OFQ produces controversial effects in pain regulation, such as supraspinal hyperalgesic but spinal analgesic effects (Mogil and Pasternak, 2001).
4.3.1 Supraspinal pain regulation of N/OFQ
Supraspinal hyperalgesia is the first reported action of N/OFQ in vivo using the hot-plate (Meunier et al., 1995) and tail-flick (Reinscheid et al., 1995) tests in the mouse.
However, the hyperalgesic effect of i.c.v. N/OFQ in mice was found to be resulted from
an inhibition of stress-induced analgesia (Mogil et al., 1996). N/OFQ administered supraspinally reverses the effects of exogenous opioids (Pan 2000).
Intracerebroventricular injection of N/OFQ was stressful, resulting in the release of central endogenous opioid peptide with their effects subsequently reversed by the delivered dose of N/OFQ. Indeed, when compared with non-injected animals rather than with i.c.v. vehicle-injected ones, i.c.v. N/OFQ-injected animals showed normal sensitivity to noxious stimulation, while i.c.v. vehicle-injected animals displayed a decreased nociceptive sensitivity (Mogil et al., 1996). In contrast, N/OFQ was ineffective in reversing analgesia induced by morphine given i.t. (Tian et al., 1997), indicating that the peptide acted as a supraspinal, but not a spinal anti-opioid peptide. In addition, all the studies, except the study of Yamamoto et al. (2001), show that i.c.v.
blockade of NOP receptors produces antinociceptive responses by various approaches, including i.c.v. injection of J-113397 in rat carrageenan test (Okuda et al., 2000), or [Nphe1]N/OFQ(1-13)-NH2 (Calo' et al., 2000b), UFP-101 (Calo' et al., 2002) or NOP antisense oligonucleotide (Rossi et al., 1997) in rat tail-flick test. Morphine-induced supraspinal analgesia is also enhanced by N/OFQ blockade (Candeletti and Ferri, 2000;
Rizzi et al., 2000). This may suggest the N/OFQ produces a basal nociceptive tone at the supraspinal level. The nociceptive tone of endogenous N/OFQ might be - at least in part - an effect secondary to anti-stress effect of N/OFQ. The tolerance induced by low
doses of morphine was reversed by NOP receptor antagonists and was absent in NOP receptor or N/OFQ knockout mice. However, the tolerance developed by escalating dose of morphine was not prevented in N/OFQ knockout mice (Chung et al., 2006; Kest et al., 2001). Therefore, the supraspinal nociceptive tone of endogenous N/OFQ may contribute to the tolerance development by low dose of morphine.
4.3.2 Spinal pain regulation of N/OFQ
At the spinal level, although N/OFQ injected intrathecally was initially reported to have no effect on thermal nociception (Reinscheid et al., 1995), a subsequent study reported a trend toward enhanced morphine analgesia by i.t. N/OFQ (Grisel et al., 1996) followed by additional support for spinal N/OFQ analgesia (King et al., 1997). In contrast to the supraspinal anti-opioid/hyperalgesic action, spinal administration produces a classical (opioid-like) antinociceptive response by inhibiting transmitter release at primary nocieptive afferent terminals (Inoue et al., 2003; Ko et al., 2006).
Indeed N/OFQ has been shown to inhibit the release of transmitters involved in this pathway including glutamate (Faber et al., 1996). Some studies using different NOP antagonist injections intrathecally suggest that endogenous N/OFQ in the spinal cord palys a protective role in the inflammatory but not acute pain status (Rizzi et al., 2006;
Yamamoto et al., 2001). However, negative results have also been reported by i.t.
injection with J-113397 in an inflammatory pain model (Fu et al., 2006) or the Nphe (Corradini et al., 2001) or JTC-801 (Muratani et al., 2002) in neuropathic pain models.
5. Periaqueductal gray (PAG)
The periaqueductal gray (PAG) is the gray matter located around the cerebral aqueduct within the tectum of the midbrain. It is a major component of a descending pain inhibitory system. The PAG projects to the RVM, which in turn sends its output to dorsal horn laminae important in nociceptive function. The PAG-RVM system is recognized as the central site of action of analgesic agents including opioids, cyclooxygenase inhibitors and cannabinoids (Hohmann et al., 2005; Leith et al., 2007;
Yaksh et al., 1976). The major intrinsic circuit within the PAG is a tonically active GABAergic net work and inhibition of this network is an important mechanism for activation of outputs of the PAG. The PAG is subdivided from statistical cluster analysis and developmental studies into four subdivisions, dorsal, dorsolateral, ventrolateral and medial (Beitz and Shepard, 1985). The medial subdivision contains the smallest neurons and exhibits the lowest cell density. The highest cell density occurred in the dorsal subdivision. The lateral division of the PAG contained the largest neurons with cells in the dorsolateral portion being slightly larger than those located ventrolaterally. The study of the projections from the PAG to the nucleus raphe magnus (NRM) indicated
that the ventrolateral subdivision provides about 70% of the total PAG output to NRM, suggesting that the ventrolateral PAG is critically involved in descending inhibition pain pathway (Beitz, 1985).
5.1. Roles of N/OFQ in the PAG
Several brain regions that are involved in pain regulation and enriched with NOP receptors can be the sites of action for the analgesic effect of i.c.v. NOP receptor antagonists. The midbrain PAG is one of the very dense distribution areas in the brain of NOP receptors either by immunolabeling (Anton et al., 1996), in situ hybridization (Neal et al., 1999a) or autoradiography in rats (Neal et al., 1999a) or in mice (Florin et al., 1997). This area is enriched with ppN/OFQ mRNA in rats (Neal et al., 1999b;
Nothacker et al., 1996) or in mice (Boom et al., 1999) or N/OFQ peptides in rats (Neal et al., 1999b) or in human (Witta et al., 2004). The midbrain PAG is the site initiating descending pain inhibitory pathway. The ventrolateral region of the PAG has been proven to be a crucial site for morphine-induced supraspinal analgesia (Yaksh et al., 1976). It is also the site of action that N/OFQ reverses morphine-induced analgesia (Morgan et al., 1997). J-113397 (i.p.) has been reported to have antinociceptive effect in rat carrageenan test, nerve injured rats and mouse formalin test but not in mouse hot-plate test (Okuda et al., 2000). It is interesting to note that Yamamoto et al. (2001)
found that i.c.v. injection of J-113397 increased the phase II, but not phase I, nociceptive responses in rat formalin test. This result is in contrast to the antinociceptive responses observed by other studies when NOP receptor antagonists were administered
i.c.v. (Calo' et al., 2000b; Calo' et al., 2002; Okuda et al., 2000; Rizzi et al., 2006).
6. Heterogeneity of NOP receptors
The NOP receptor has been proposed to be heterogeneous from several aspects, including the presence of 1) splicing variants of NOP receptors, 2) two binding sites of
125I-[Tyr14]N/OFQ in the brain, 3) distict distriubiton in the bindings sites of
125I-[Tyr10]N/OFQ(1-11) and 125I-[Tyr14]N/OFQ, and 4) functional heterogeneity of
NOP receptors.
6.1. Splicing variants of NOP receptors
The gene encoding murine NOP receptors, which is believed to be a single copy gene, termed Oprl1 and localized on chromosome 2, was found to have alternative splicing sites (Nishi et al., 1997). Several splicing variant mRNAs have been found in the brain, although none of the encoded protein has yet been identified (Mogil and Pasternak, 2001).
6.2. Binding studies with 125I-[Tyr14]N/OFQ
Binding studies with 125I-[Tyr14]N/OFQ as a radioligand showed curvilinear scatchard plots in the mouse brain which are best fit by nonlinear regression analysis to two sites (Mathis et al., 1997). The high affinity site (KD: 3.8 pM) is far less abundant in brain tissue (Bmax: 31.6 fmol/mg protein) than the lower affinity site (KD: 896 pM; Bmax: 233 fmol/mg protein). The curvilinear scatchard plot in brain membranes contrasts sharply with linear scatchard plots seen in CHO cells transfected with NOP receptors (KD: 36 pM) (Table 5). These results suggest there are high and low affinity binding sites of N/OFQ in rodent brains.
6.3. Autoradiographic studies with 125I-[Tyr10]N/OFQ(1-11) and 125I-[Tyr14]N/OFQ
Autoradiographic studies, using 125I-[Tyr10]N/OFQ(1-11) and 125I-[Tyr14]N/OFQ as radioligands, showed that their distributions in the brain of either mice (Mathis et al., 1999) or rats (Letchworth et al., 2000) are different. Binding parameters of
125I-[Tyr10]N/OFQ(1-11) revealed an affinity (KD) of 235 pM, which is over 100-fold
lower than its Ki in inhibition of against 125I-[Tyr14]N/OFQ binding in mouse brain. The binding site of 125I-[Tyr10]N/OFQ(1-11) was, therefore, proposed to be the high affinity site for 125I-[Tyr14]N/OFQ, based from their similar maximal binding densities (Bmax) (Table 5). The binding density of 125I-[Tyr10]N/OFQ(1-11) is one sixth of that of
125I-[Tyr14]N/OFQ (1.3+0.2 and 7.8+0.3 fmol/mg, respectively) in the rat midbrain PAG
(Letchworth et al., 2000), suggesting that [Tyr10]N/OFQ(1-11) might affect a portion of N/OFQ-sensitive NOP receptors. Recently, Dr. Pasternak’s group showed that, in both wild type and NOP-knockout mice, N/OFQ displayed similar Ki values in competing the binding of 125I-[Tyr10]N/OFQ(1-11) at the high affinity site (Majumdar et al., 2009).
This suggests that the high affinity binding site of N/OFQ still exists in NOP-knockout mice.
6.4. Functional heterogeneity of NOP receptors revealed from the results of Ro
64-6198
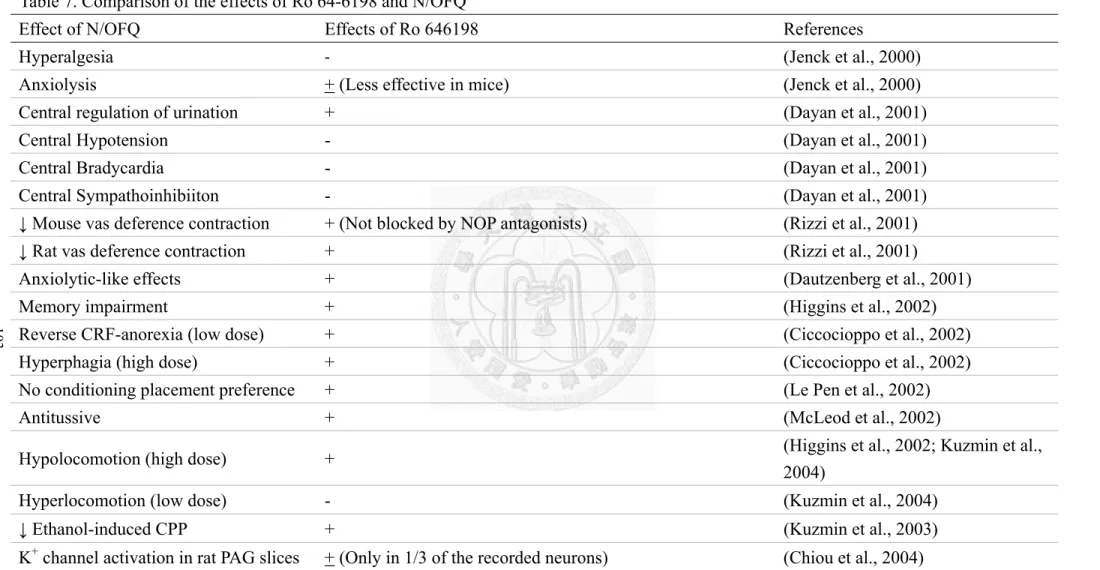
As reported previously, we have proven that Ro 64-6198 activates only a subset of the N/OFQ-sensitive NOP receptors in rat PAG (Chiou et al., 2004). Several studies also indicate that Ro 64-6198 mimicked some, but not all, of the effects of N/OFQ (Table 7).
N/OFQ (i.c.v.) produced anxiolytic effect in both rats and mice but Ro 64-6198 (i.p.) was less effective in producing anxiolytic effect in mice at the doses unaffecting motor activity (Jenck et al., 2000). N/OFQ affects locomotor activity in a biphasic manner, increasing locomotion at lower doses but decreasing it at higher doses. Ro 64-6198 failed to fully reproduce the effect of N/OFQ, instead, it inhibited locomotor activity at all doses (Kuzmin et al., 2004). Ro 64-6198 mimicked the effects of N/OFQ to inhibit
contraction of mouse vas deferens, but the effects of Ro 64-6198 were not blocked by neither NOP nor MOP receptor antagonists (Rizzi et al., 2001). Ro 64-6198 regulated urination, but not hypotension or bradycardia, centrally as did N/OFQ (Dayan et al., 2001). In a neuropathic pain model, Ro 64-6198 was less effective in reducing allodynic responses than N/OFQ if given by i.t. administration, but was equopotent as N/OFQ if given by intraplantar injection.(Obara et al., 2005). In addition, Ro 64-6198 attenuated the expression of morphine sensitization at higher doses via a mechanism not blocked by a NOP receptor antagonist (Kotlinska et al., 2005). Although Ro 64-6198 increased food intake as N/OFQ, it did not decrease alcohol consumption as N/OFQ (Economidou et al., 2006a). However, it reduced alcohol self-administrtion and prevented relapse-like alcohol drinking (Kuzmin et al., 2007). Taken together, these effects of Ro 64-6198 suggest that there are functional heterogeneity of NOP receptors.
Aims of Study Hypotheses:
1. NOP receptors are functionally heterogeneous in vlPAG neurons. This was
proven by our previous study using Ro 64-6198 and can also be revealed by
(+)-5a Compound, [Tyr10]N/OFQ(1-11) or new NOP receptor antagonists.
2. [Tyr10]N/OFQ(1-11) is an agonist for the high, but not low, affinity binding site
of N/OFQ in rodent brain. The Ro 64-6198-sensitive NOP receptors might be
the NOP receptors exhibiting high binding affinity for N/OFQ.
Aim 1: To investigate if (+)-5a Compound can also affect a portion of NOP
receptors in vlPAG neurons.
(+)-5a Compound has a structure backbone analogous to Ro 64-6198 and Ro 64-6198 acts differently from N/OFQ at NOP receptors in rat PAG slices. If our hypothesis 1 is true, (+)-5a Compound should, like Ro 64-6198, also affect a portion of NOP receptors in vlPAG neurons.
Aim 2: To investigate if (+)-5a Compound affects the same subset of NOP
receptors that Ro 64-6198 effects.
If both (+)-5a Compound and Ro 64-6198 affect a portion of NOP receptor in
vlPAG neurons, if they affect the same subset of NOP recpeotrs will be examined.
Aim 3: To characterize the neurochemical and morphological properties of
the neurons sensitive to Ro 64-6198 and/or (+)-5a Compound.
If (+)-5a Compound affects the same subset of NOP receptors that Ro 64-6198 effects. The neurochemical and morphological properties of the neurons sensitive and insensitive to Ro 64-6198 and (+)-5a Compound will be examined.
Aim 4: To investigate if [Tyr10]N/OFQ(1-11), an agonist for the high affinity
binding site of N/OFQ in rodent brain, also affect a portion of NOP receptors in
vlPAG neurons.
Binding studies showed that 125I[Tyr14]N/OFQ (low-affinity site) and
125I[Tyr10]N/OFQ(1-11) (high-affinity site) displayed distinct distribution sites in rodent
brains. The binding densities of high-affinity site is one sixth of that of low-affinity sites in the rat PAG, suggesting that [Tyr10]N/OFQ(1-11), which had higher selectivity for the high-affinity site than N/OFQ(1-11), might affect a portion of N/OFQ-sensitive NOP receptors. [Tyr10]N/OFQ(1-11) had higher selectivity for the specific binding site of
125I-[Tyr10]N/OFQ(1-11) than its parent peptide, N/OFQ(1-11) (Table 6). Therefore, we
used [Tyr10]N/OFQ(1-11) as a tool to investigate if it can reveal the heterogeneity of
NOP receptors in vlPAG neurons.
Aim 5: To investigate if [Tyr10]N/OFQ(1-11) affects the same subset of NOP
receptors that (+)-5a Compound does.
If the aim 4 is true, then the pharmacological files of [Tyr10]N/OFQ(1-11) will be compared with that of (+)-5a Compound to validate if they affect the same subset of NOP receptors.
Aim 6: To characterize the pharmacological properties of two novel NOP receptor
antagonists, Compound 24 and SB-612111, and examine if they can differentiate
the NOP receptors which are all sensitive to N/OFQ in vlPAG neurons.
We will further examine if the heterogeneity of NOP receptors can be revealed by NOP receptor antagonists, Compound 24 and SB-612111. In addition, their pharmacological properties in vlPAG neurons will be quantitatively characterized.
Materials and methods:
All experiments were conducted with Wistar rats of 9-18 days old and conformed to the guidelines of the Institutional Animal Care and Use Committee of the College of Medicine, National Taiwan University. All efforts were made to minimize the number of animals used.
1. Brain slice preparations
The midbrain blocks containing the PAG were rapidly dissected from postnatal Wistar rats. Coronal slices (300 or 400 μm) were then sectioned with a vibrotome (microslicer DTK-100, Dosaka) and equilibrated at room temperature in the artificial cerebral spinal fluid (aCSF). The aCSF consisted of (in mM) 117 NaCl, 4.5 KCl, 2.5 CaCl2, 1.2 MgCl2, 1.2 NaH2PO4, 25 NaHCO3 and 11.4 dextrose (pH=7.4), and was oxygenated with 95% O2/5% CO2. After equilibration for at least one hour, one slices was mounted on a submerged recording chamber and continuously perfused with aCSF at a rate of 2-3 ml/min.
2. Electrophysiological recordings
Blind patch-clamp whole cell recording was performed with 4-8 MΩ glass microelectrodes filled with the internal solution consisting of (in mM): 125 K+
gluconate, 5 KCl, 0.5 CaCl2, 5 BAPTA, 10 HEPES, 5 MgATP and 0.33 GTPtris (pH=7.3). To elucidate if (+)-5a Compound, like N/OFQ, also induced the NOP receptor-mediated GIRK currents, a hyperpolarization voltage ramp protocol was applied. After whole cell configuration was formed, the potential of the recorded neuron was held at -70 mV, stepped to -60 mV for 100 ms, ramped from -60 mV to -140 mV for 400 ms, and then stepped back to -70 mV (Fig. 3, inset). The membrane currents elicited by voltage ramps were acquired through an Axopatch 200B amplifier (Molecular Devices/Axon Instruments, Union City, CA) with a pClamp 7 software (Molecular Devices/Axon Instruments, Union City, CA) and simultaneously recorded with a chart recorder (Gould RS3200) at a low frequency response of 10 Hz to monitor the time course of drug effects. The access resistance (10-15 MΩ) was monitored during the recording period. Only those neurons with unchanged access resistance before and after drug treatments were accepted to ensure that the clamp efficiency was not deteriorated during the recording period.
3. Quantitative analysis of NOP receptor ligands
3.1. NOP receptor agonists
The effect of the tested NOP receptor agonist was quantified by the percent increment of the membrane current at -140 mV (I-140), taking its own I-140 before
treatment as 100%. An increment of I-140 greater than 5% and the induced-current having a reversal potential at around -90 mV (the equilibrium potential of K+ ions) was considered to be effective (defined as agonist-sensitive). For establishing the concentration-response curves of (+)-5a Compound or [Tyr10]N/OFQ(1-11), the percent increment of I-140 in each neuron was normalized to the maximal effect (Emax) produced by 1 μM N/OFQ, which was 39.4%+4% increment (n=26) (Chiou et al., 2002a). The EC50 values of (+)-5a Compound and [Tyr10]N/OFQ(1-11) were determined by the refection point of its concentration-response curve produced by logistic fitting:
E=Emax/[1+(D/EC50)n], where E represents the percentage of increment, Emax the maximal increment, D the concentration of agonist and n the Hill coefficient. To quantitatively evaluate the antagonistic effect of various receptor antagonists, their interactions with (+)-5a Compound or [Tyr10]N/OFQ(1-11) were examined in the same neuron. Given that not all neurons were sensitive to (+)-5a Compound or [Tyr10]N/OFQ(1-11) (see Results), it was not practical to pre-apply the intended antagonist. Therefore, the tested receptor antagonistwas applied to (+)-5a Compound or [Tyr10]N/OFQ(1-11)-sensitive neurons after the response of (+)-5a Compound or [Tyr10]N/OFQ(1-11) had reached a steady state, which usually took 20-25 min. The response of the tested antagonist was continuously monitored thereafter. In the study verifying whether NOP receptor-mediated GIRK channels were functional in those
(+)-5a Compound or [Tyr10]N/OFQ(1-11)-unresponsive neurons,N/OFQ was examined after (+)-5a Compound or [Tyr10]N/OFQ(1-11) had been applied for atleast 20 min. In this set of experiments, Ro 64-6198 was tested in some (+)-5a Compound-unresponsive neurons before N/OFQ was added in order to verify if the neurons unresponsive to (+)-5a Compound were also insensitive to Ro 64-6198. [Tyr10]N/OFQ(1-11) was tested in the (+)-5a Compound-treated neurons in order to verify the correlation with (+)-5a Compound.
3.2. NOP receptor antagonists
To quantitatively estimate the antagonistic effect of Compound 24 or SB-612111 against N/OFQ- or DAMGO-induced GIRK current, Compound 24 or SB-612111 was applied after the response to N/OFQ or DAMGO had reached a steady state. The GIRK current induced by N/OFQ or DAMGO was quantified as the percent increment of the membrane current recorded at -140 mV (I-140) in each neuron, taking its own I-140 before treatment as 100%. For establishing the concentration-response curve of Compound 24 or SB-612111 against N/OFQ- or DAMGO-induced GIRK current, the inhibitory effect of Compound 24 or SB-612111 in each neuron was calculated, taking the effect of N/OFQ or DAMGO in the same neuron as 100 %. The IC50 values of Compound 24 and SB-612111 were determined by the refection point of its concentration-response curve
in its inhibition of 0.1 μM N/OFQ-induced GIRK current produced by logistic fitting:
I=Imax/[1+(D/IC50)n], where I represents the percentage of inhibition, Imax the maximal inhibition, D the concentration of antagonist and n the Hill coefficient.
4. Immunofluorescence staining
For immunofluorescence studies, 0.2% Lucifer yellow (LY) was added in the internal solution. After recording, the slice containing the recorded neuron which had been filled with Lucifer yellow was fixed, re-sectioned and subjected to an immunofluorescent staining of glutamic acid decarboxylase-67 (GAD67), a synthesizing enzyme of GABA (Erlander et al., 1991). Briefly, after recording, the slices were fixed with 4% paraformaldehyde at 4 °C for one day, and then dehydrated in 30%
sucrose. Dehydrated slices were embedded and re-sectioned into 50 μm sections with a cryostat microtome (Leica CM3050S, Leica Microsystems, Nussloch, Germany). Slice sections were rinsed and washed with phosphate buffered saline (PBS) 3 times, followed by 0.3% Triton X-100 containing PBS (PBST) plus 0.5% bovine serum albumin (BSA) and then blocked in PBST containing 1% BSA and 10% normal goat serum (NGS) for 1 h. Then, slice sections were incubated with the mouse monoclonal antibody against GAD67 (diluted 1:1000) (Chemicon, Temecula, CA) in PBST containing 1% BSA overnight at 4 °C. Slice sections were then washed with PBST 3
![Table 5. Saturation analysis of 125 I-[Tyr 14 ]N/OFQ and 125 I-[Tyr 10 ]N/OFQ(1-11) binding in transfected CHO cells and mouse brain](https://thumb-ap.123doks.com/thumbv2/9libinfo/9607213.632992/115.892.125.762.183.349/table-saturation-analysis-binding-transfected-cells-mouse-brain.webp)
![Table 6. Binding affinity of N/OFQ derived peptides at 125 I-[Tyr 10 ]N/OFQ(1-11) and 125 I-[Tyr 14 ]N/OFQ binding sites in mouse brain and expressed NOP receptors in CHO cells](https://thumb-ap.123doks.com/thumbv2/9libinfo/9607213.632992/116.1263.110.1174.183.567/table-binding-affinity-derived-peptides-binding-expressed-receptors.webp)
![Table 9. Quantitative in vitro studies of N/OFQ(1-11) and [Tyr 10 ]N/OFQ(1-11) N/OFQ(1-11) Preparation N/OFQ effect Ki †/EC 50 ‡](https://thumb-ap.123doks.com/thumbv2/9libinfo/9607213.632992/120.1263.384.1162.184.651/table-quantitative-vitro-studies-ofq-tyr-preparation-effect.webp)