國立臺灣大學生農學院動物科學技術學系 碩士論文
Department of Animal Science and Technology College of Bioresources and Agriculture
National Taiwan University Master Thesis
利用分子標識分析臺灣山羊族群之 族群遺傳結構與生長性狀之相關性 Using the genetic markers to analyze
population genetic structure and growth traits association in goat populations
in Taiwan 殷瓊瑛 Chiung-Ying Yin 指導教授:王佩華 博士 Advisors: Pei-Hwa Wang, Ph. D.
中華民國 104 年 7 月
July, 2015
I
目 錄
誌 謝... III 圖 次... IV 表 次... VI 附表次... VII 中文摘要... VIII 英文摘要... X
壹、文獻檢討... 1
一、臺灣肉羊產業發展概況與產值... 1
二、臺灣山羊育種與保種... 3
三、臺灣現有山羊品種介紹... 4
四、分子遺傳標識(molecular genetic marker) ... 19
五、分子標識之多態性在族群的研究... 24
六、分子標識在臺灣山羊之應用... 31
貳、材料與方法... 41
詴驗一:應用微衛星標識進行臺灣山羊族群遺傳結構分析... 41
(一)、詴驗動物... 41
(二)、血液採集及基因組 DNA(genomic DNA, gDNA)萃取 ... 41
(三)、微衛星標識引子設計... 41
(四)、多引子聚合酶鏈鎖反應 (multiplex-PCR) ... 42
(五)、電泳樣品製備... 42
(六)、PCR 產物基因型判別 ... 47
(七)、微衛星標識基因型頻率統計... 47
詴驗二:利用候選基因法進行臺灣努比亞生長性狀及體型測量值相關性分析.. 48
(一)、詴驗動物及生長性狀資料收集... 48
(二)、血液採集及基因組 DNA(genomic DNA, gDNA)萃取 ... 48
(三)、及時聚合酶鏈鎖反應(real-time PCR)及候選基因多態性分析 ... 48
(四)、統計分析... 54
參、結果... 55
詴驗一:應用微衛星標識進行臺灣山羊族群遺傳結構分析... 55
(一)、微衛星標識基因座與遺傳變異... 55
(二)、遺傳介值評估臺灣黑山羊族群遺傳變異與近親程度... 55
(三)、族群間基因流動(gene flow)與遺傳分化程度(genetic differentiation, FST)分析 ... 59
(四)、遺傳距離與分群分析... 59
(五)、個體鑑別率之估算... 69
詴驗二:利用候選基因法進行臺灣努比亞生長性狀及體型測量值相關性分析.. 72
(一)、候選基因多態性之分析... 72
(二)、MSTN 基因 5 端 UTR 區域基因多態性分析 ... 72
(三)、IGF-1 基因第四內顯子基因多態性分析... 78
(四)、POU1F1 基因第六外顯子區域基因多態性分析 ... 84
肆、討論... 90
詴驗一:應用微衛星標識進行臺灣山羊族群遺傳結構分析... 90
(一)、微衛星基因座與遺傳變異... 90
(二)、族群遺傳結構與近親程度結果... 91
(三)、遺傳距離與分群... 91
(四)、臺灣山羊個體鑑別率... 92
(五)、應用衛星標識於臺灣黑山羊族群保育之探討... 93
詴驗二:利用候選基因法進行臺灣努比亞生長性狀及體型測量值相關性分析.. 94
(一)、MSTN 基因 5 端 UTR 區域基因多態性分析 ... 94
(二)、IGF-1 基因第四內顯子區域基因多態性分析... 95
(三)、POU1F1 第六外顯子區域基因多態性分析 ... 97
伍、結論... 99
參考文獻... 100
附錄... 108
III
誌 謝
能完成這篇論文,我要特別感謝我的指導教授 王佩華老師,老師不厭其煩 地指導、不時的討論並指點迷津,讓學生獲益匪淺,老師對學問的嚴謹與認真更 是學生的典範。
也要感謝詴驗期間,有幸合作的農委會畜產詴驗所恆春分所、花蓮種畜繁殖 場及各民間養羊場,提供詴驗羊隻、採樣及人力上的協助。動物育種與資源應用 實驗室的每一位成員,包括怡蓁學姐與芳裕學長親自指導實驗技巧;岳錚學長、
柏安學長與瑞軒學長給予資料統計分析的思考方向;薏婷學姊、雨蓁學姊與徐博 提供實驗室內所有的技術支援及諮詢協助;亦感謝林彤、孙軒與晉廷在實驗上的 同甘共苦,大家對我的支持與鼓勵,點滴在心頭,僅此表達最誠摯的謝意。
除了實驗室以外,特別感謝系辦公室的奕雯學姊及位育學長,不時提醒與諄 諄告誡學生們的修課和畢業等相關事宜;以及系上的每一位老師,在課堂上的不 吝指導;最後感謝 R02 的同學們生活及課業上的相互作伴與勉勵,也感謝系上 學弟妹們熱情參與實習課,教學相長,豐富彼此的求學生涯,添加許多美好的回 憶。
最後,感謝我的家人,你們無私地提供我在求學階段所需的一切,我是最幸 運的孩子,讓我有足夠的勇氣去面對一切的困難,其他所有愛我及支持我的朋友,
感謝你們陪我度過許多艱難的時刻,願每一個人帄安喜樂,生活順遂。
圖 次
圖 1 臺灣現有主要的山羊品種。a: 臺灣黑山羊;b: 努比亞;c: 波爾;d: 撒 能;e: 阿爾拜因;f: 吐根堡。... 9 圖 2 波爾山羊品種之不同品系。a: 標準毛色品系;b: 純黑品系;c: 純紅品系。
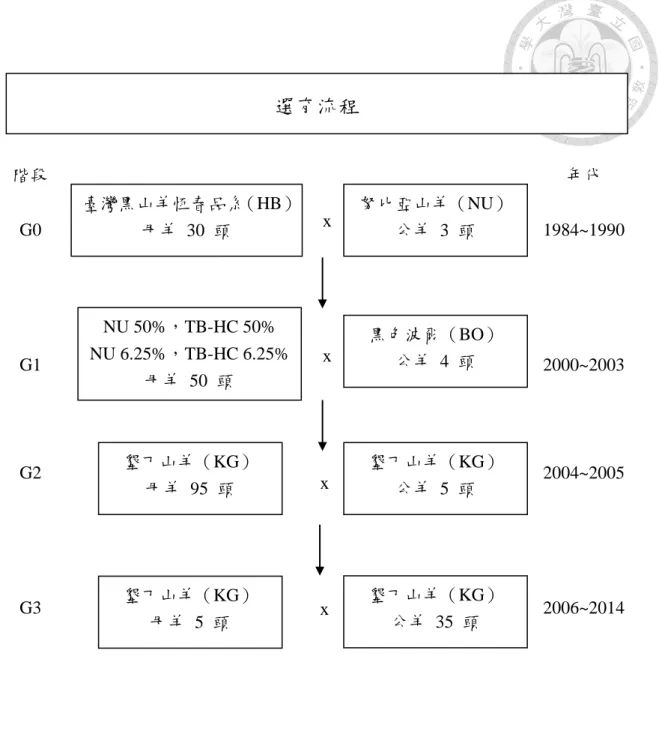
... 10 圖 3 墾丁山羊選育流程圖。... 13 圖 4 墾丁山羊外貌特徵。a:墾丁山羊之公羊;b:墾丁山羊之母羊;c:墾丁山
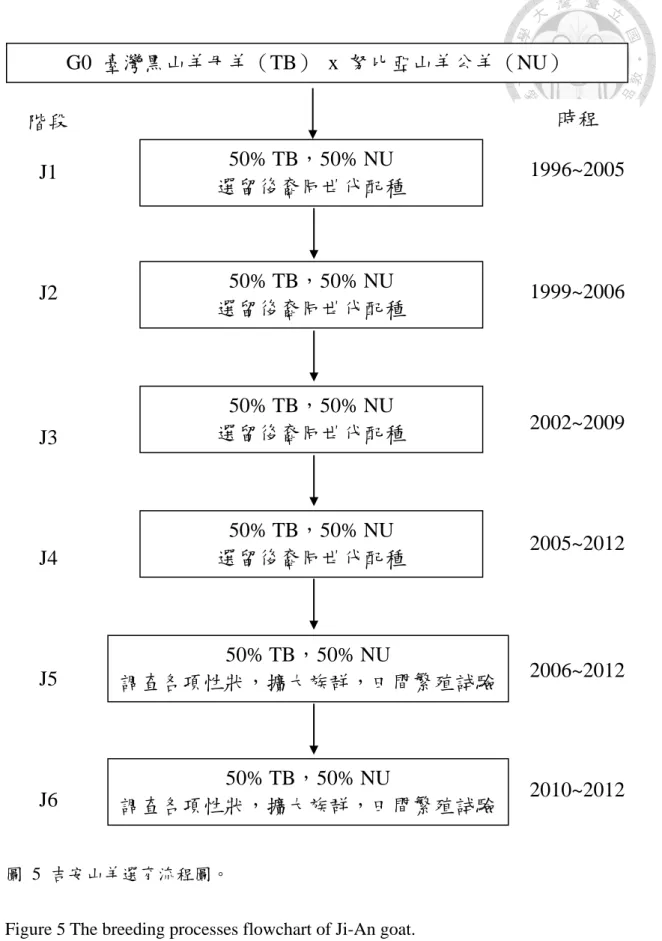
羊之女羊;d:墾丁山羊之仔羊。... 14 圖 5 吉安山羊選育流程圖。... 17 圖 6 吉安山羊外貌特徵。a:種公羊;b:種母羊;c:V 型外張角;d:耳大向
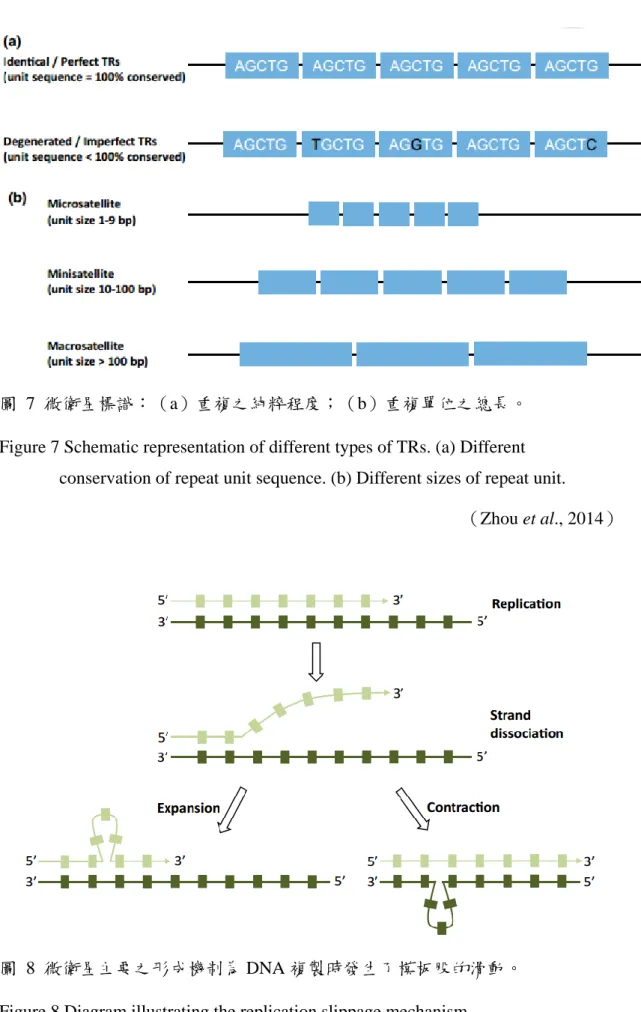
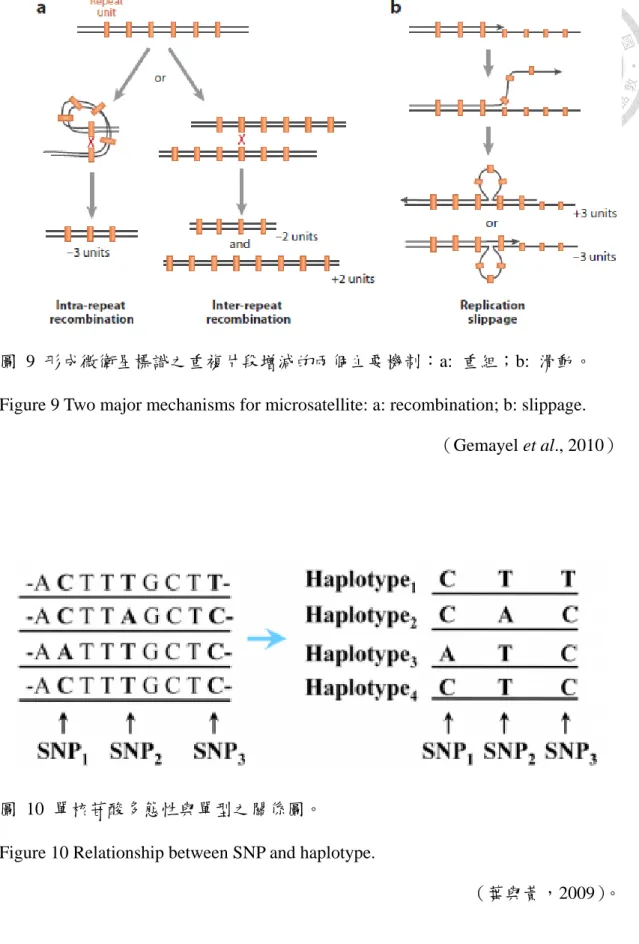
外擴張;e:長鬚;f:瞳孔為圓型。 ... 18 圖 7 微衛星標識:(a)重複之純粹程度;(b)重複單位之總長。 ... 21 圖 8 微衛星主要之形成機制為 DNA 複製時發生了模板股的滑動。 ... 21 圖 9 形成微衛星標識之重複片段增減的兩個主要機制:a: 重組;b: 滑動。.. 22 圖 10 單核苷酸多態性與單型之關係圖。... 22 圖 11 親緣關係樹:a. 未加權算術帄均對群法(UPGMA)及 b. 鄰位連接法(NJ)
的圖示。... 30 圖 12 比利時藍牛(左)與皮埃蒙特牛(右)的雙肌表現型。 ... 33 圖 13 缺乏 MSTN 基因小鼠(a)的肌肉量為野生型(b)的兩到三倍。前肢末
梢切片之蘇木素-伊紅染色圖(c)缺乏 MSTN 基因小鼠(d)野生型小鼠。
... 33 圖 14 具 有 MSTN 基 因 突 變 之 特 賽 爾 綿 羊 個 體 出 現 肌 肉 量 增 加 之 表 現 型
(increased muscle mass, IMM)。 ... 34 圖 15 野生型六日齡與二月齡小鼠(w and W)與 IGF-I 基因突變個體(i and I)
身形比較。... 38 圖 16 自哺乳動物白血球抽取基因組 DNA 的流程。 ... 44 圖 17 利用 15 組微衛星標識將不同山羊族群以鄰近相接法繪製類緣關係樹。
... 62 圖 18 各山羊族群利用族群遺傳距離所建構之主座標分析(PCA)。各軸上數字
表示各軸所區分出之變異占總變異量之百分比。KM:金門山羊、TB-HC:
臺灣黑山羊恆春品系、TB-HU:臺灣黑山羊花蓮品系、JA:吉安山羊、NU-C:
C 場努比亞、NU-HC:恆春努比亞、NU-S:S 場努比亞、KG:墾丁山羊、
BO:波爾、AL:阿爾拜因。 ... 65 圖 19 各山羊個體利用 15 組微衛星標識所繪製之 STRUCTURE 群集分析圖
(K 4 至 7)。K 值為軟體預設之可能的群集數,以不同顏色表示之,縱 軸為個體的基因來源於該群集之比例,且每一圖條表示一個個體。NU-C:
C 場努比亞、NU-S:S 場努比亞、KG:墾丁山羊、TB-HC:臺灣黑山羊恆
V
春品系、NU-HC:恆春努比亞、BO:波爾、AL:阿爾拜因、KM:金門山 羊、JA:吉安山羊、TB-HU:臺灣黑山羊花蓮品系。 ... 66 圖 20 (a)L(K) 與(b) K 之折線圖。兩種分析圖皆顯示建議 K 值為 4。
... 67 圖 21 全體山羊個體利用 15 組微衛星標識以鄰位連接法所得之親緣關係樹。
... 68 圖 22 全體山羊族群之個體鑑別率(P(ID))以及近親個體鑑別率(P(ID)sib)對微 衛星標識數目之折線圖。... 71 圖 23 山羊肌肉抑制素基因 5 端 UTR 片段以 DraI 限制酶切割之基因型判
定。;Lane 1 及 4:BB 基因型(504 和 179 bp);Lane 2:AA 基因型(696 bp);Lane 3:AB 基因型(696、504 和 179 bp);M:100 bp DNA ladder marker。 ... 74 圖 24 MSTN 基因 5 端 UTR 區域 g.1256 TTTTA/- 位點以 TaqMan SNP Genotyping Assay 分析之基因型結果。未加 DNA 模板之對照組(negative control)以( )表示。 ... 75 圖 25 山羊類胰島素生長因子 I 基因第四內顯子區域片段以 HaeIII 限制酶切
割之基因型判定。Lane 1:CC 基因型(177 bp);Lane 2:CG 基因型(177 、 117 和 60 bp);Lane 3:GG 基因型(117 和 60 bp)。M:100 bp DNA ladder marker 大小為 60 bp 的片段無法在 agarose gel 上顯示。 ... 80 圖 26 IGF-I 基因第四內顯子區域 g.282G>C 位點以 TaqMan SNP Genotyping Assay 分析之基因型結果。未加 DNA 模板之對照組(negative control)以
( )表示。 ... 81 圖 27 山羊 POU1F1 基因第六外顯子區域 g.102T>G 變異位點(黃色標示)。(a)
GG 基因型;(b)TG 基因型;(c)TT 基因型。 ... 86 圖 28 POU1F1 基 因 第 六 外 顯 子 區 域 g.102T> G 位 點 以 TaqMan SNP Genotyping Assay 分析之基因型結果。未加 DNA 模板之對照組(negative control)以( )表示。 ... 87
表 次
表 1 來自 4 個羊場共 10 個山羊族群之詴驗動物樣本數... 43
表 2 本詴驗使用之 15 組微衛星標識之標定螢光引子資料一覽表... 45
表 3 本詴驗三組多引子 PCR 的反應條件 ... 46
表 4 本詴驗候選基因 MSTN、POU1F1 和 IGF-1 突變型態與突變位點 ... 50
表 5 本詴驗各候選基因 TaqMan SNP Genotyping Assay 之引子及螢光探針序列 ... 51
表 6 以 PCR-RFLP 分析肌肉抑制素基因(MSTN)與類胰島素生長因子 I 基因 (IGF-1)之引子序列、限制酶種類、PCR 產物大小與擴增位置,以及以直 接定序法分析垂腺特定轉錄因子 1 基因(POU1F1)之引子序列、PCR 產 物大小與擴增位置... 52
表 7 本詴驗中分析肌肉抑制素基因(MSTN)、類胰島素生長因子 I(IGF-1)與 垂腺特定轉錄因子 1(POU1F1)基因多態性之 PCR 反應條件 ... 53
表 8 利用 15 個微衛星標識於整體山羊族群之交替基因範圍、觀測與期望交替 基因數目、期望與觀測雜合度以及多態性訊息含量... 57
表 9 利用 15 個微衛星標識檢測詴驗之臺灣山羊族群間遺傳變異... 58
表 10 10 個山羊族群間遺傳分化程度(FST)與基因流動(Nm)估計值 ... 60
表 11 本詴驗各山羊族群之間的遺傳距離估值 ... 61
表 12 全體山羊族群於 15 組微衛星標識之個體鑑別率(P(ID))以及近親個體鑑 別率(P(ID)sib) ... 70
表 13 來自不同族群之努比亞山羊族群 MSTN 基因 5 端 UTR 區域 g.1256 TTTTA/- 變異位點之基因型及交替基因頻率 ... 76
表 14 MSTN 基因 5 端 UTR 區域 g.1256TTTTA/- 之基因型與努比亞山羊生 長性狀之相關性... 77
表 15 來自不同族群之努比亞山羊族群 IGF-I 基因第四內顯子區域 g.282G>C 變 異位點之基因型及交替基因頻率... 82
表 16 IGF-I 基因第四內顯子區域 g.282C>G 之基因型與努比亞山羊生長性狀 之相關性... 83
表 17 來 自 不 同 族 群 之 努 比 亞 山 羊 族 群 POU1F1 基 因 第 六 外 顯 子 區 域 g.102T>G 變異位點之基因型及交替基因頻率 ... 88
表 18 POU1F1 基因第六外顯子區域 g.102T>G 基因型與努比亞山羊族群生長 性狀之相關性... 89
VII
附表次
表 A. 1 ILSTS019 基因座於 10 個山羊族群之交替基因頻率 ... 108
表 A. 2 INRA063 基因座於 10 個山羊族群之交替基因頻率 ... 109
表 A. 3 SRCRSP08 基因座於 10 個山羊族群之交替基因頻率 ... 110
表 A. 4 HSC 基因座於 10 個山羊族群之交替基因頻率 ... 111
表 A. 5 SRCRSP023 基因座於 10 個山羊族群之交替基因頻率 ... 112
表 A. 6 OarFCB20 基因座於 10 個山羊族群之交替基因頻率 ... 113
表 A. 7 McM0527 基因座於 10 個山羊族群之交替基因頻率 ... 114
表 A. 8 INRA023 基因座於 10 個山羊族群之交替基因頻率 ... 115
表 A. 9 CSRD0247 基因座於 10 個山羊族群之交替基因頻率 ... 116
表 A. 10 BM1258 基因座於 10 個山羊族群之交替基因頻率 ... 117
表 A. 11 SRCRSP001 基因座於 10 個山羊族群之交替基因頻率 ... 118
表 A. 12 SPS113 基因座於 10 個山羊族群之交替基因頻率 ... 119
表 A. 13 BOBT24A 基因座於 10 個山羊族群之交替基因頻率 ... 120
表 A. 14 BM1329 基因座於 10 個山羊族群之交替基因頻率 ... 121
表 A. 15 SPS115 基因座於 10 個山羊族群之交替基因頻率 ... 122
中文摘要
山羊為臺灣酪農產業中僅次於乳牛的重要乳品與肉品來源,又山羊肉相較於 其他肉品,避免了文化與宗教因素的限制,因而逐漸為消費者所接受,開啟了透 過人擇來篩選具有較優良生長性能山羊個體之選拔工作。由於我國本土山羊臺灣 黑山羊(Taiwan black goat, TB)生長緩慢且產肉率低,因此透過引入外國品種山 羊進行育種改良,然而大量雜交詴驗與缺乏配套之本地黑山羊保育工作,導致本 地黑山羊數量大幅銳減與遺傳歧異度流失,有鑑於本地黑山羊為臺灣之特有種,
必頇將其種源(germplasm)加以保育。近年來,分子標識已廣為應用於評估物 種間之遺傳變異與供分子育種之用途,本研究目的為利用分子標識,分析包含臺 灣黑山羊之族群遺傳結構與生長性狀之相關性,以兼顧種源的保存以及培育優良 種源之目標。
本研究蒐集來自 10 個山羊族群共 682 頭山羊之血樣,以 15 組微衛星標 識進行遺傳變異分析,詴驗結果顯示,在所檢測的山羊族群,帄均交替基因數
(average number of alleles , Na)與有效交替基因數(effective alleles , Ne)分別 為 11.87±3.40 及 5.1±1.80;帄均期望異質度(average expected heterozygosities, HE)與觀測期望異質度(observed heterozygosities, HO)分別為 0.781±0.081 與 0.602±0.116;帄均多態性訊息含量(polymorphic information content, PIC)為 0.747±0.103;FIS 的範圍從 -0.034 至 0.215 之間,帄均值為 0.058,顯示所有的 微衛星標識在所檢測的族群中皆具有多態性(polymorphism),且所有詴驗族群 已有近親之現象。進一步使用 Nei (1972)法計算各族群間遺傳距離矩陣並以鄰近 相接法(neighbor-joining, NJ)繪製親緣關係樹(phylogenetic tree),結果顯示可將臺 灣黑山羊族群與其他山羊族群進行分群(clustering),且臺灣黑山羊種源於恆春分 所保存完善。此外,本研究另選擇對生長具有影響之候選基因,進行基因型與諸 在 養 山 羊 生 長 性 狀 之 相 關 性 分 析 , 包 含 MSTN g.1256TTTTA/- 、 POU1F1 g.102T>G 與 IGFI g.282G>C 三組 SNP 標識,並蒐集 497 頭努比亞山羊於出
IX
生、三月齡(離乳)、六月齡、九月齡及十二月齡之體重(body weight, BW)、體 高(body height, BH)、體長(body length, BL)及胸圍(body chest girth, BCG)
之測量值,並計算各生長階段之帄均日增重(average daily gain, ADG),結果顯 示,三組 SNP 標識皆對努比亞生長性狀具有顯著地相關性。帶有 MSTN 基因 之 BB 基因型個體於 BCG0、ADG6、BW6 和 ADG9 顯著地高於其他基因型
(P < 0.05);帶有 POU1F1 基因之 D1D1 基因型個體於 BH6、BL6、BCG6、
BW9、BH9、BL9、BCG9、BL12 和 BCG12 顯著地高於其他基因型(P < 0.05);
帶有 IGFI 基因之 CC 基因型個體則在 BW0、BL0、BCG0、BH6、BL、 BCG6、
BL9、BCG9、BH12 和 BL12 顯著地高於其他基因型(P < 0.05)。
綜合所述,本詴驗所使用之微衛星標識,可應用於臺灣黑山羊及其雜交族群 之遺傳結構分析,可供未來種源與保育工作之參考依據。同時,本詴驗提供對提 升生長性狀具有顯著效應之三組 SNP 標識,可進一步應用於臺灣肉用山羊之育 種用途。因此,透過分子標識輔助選拔(marker-assisted selection, MAS),可加 速更有效率且精確之保種與育種標的。
關鍵字:山羊、分子標識輔助選拔、微衛星標識
英文摘要
Goat was an important source of milk and meat in addition to cow in dairy farming in Taiwan. Goat meat has becoming much more acceptable to consumers without regards of cultural and religious barriers. Therefore, selection of goats to improve growth traits was brought into action. Due to the poor growth rate and meat yield of Taiwan black goat (TB) breed in Taiwan, many exotic breeds were brought into breeding scheme to improve their growth trait. However, excessively crossbreeding alien species with TB decreased its population and their genetic variation, so which was considered as an endemic species in Taiwan that needed to be preserved. In recent years, the genetic markers were studied and applied in domestic animals, aiming at evaluating and selecting across breeds. Hence the objective of this study is using genetic marker to analyze genetic structure and its growth performance in Taiwan, setting the purpose of genetic improvement as well as to sustain the germplasm preservation.
There were 15 sets of microsatellite markers used for analyzing 682 goats that sampled from 10 populations. The average number of alleles (Na) and effective alleles (Ne) were 11.87±3.4 and 5.1±1.8. The average expected heterozygosity (HE) and observed heterozygosity (HO) per locus were 0.781±0.081 and 0.602±0.116. Average polymorphic information content (PIC) was 0.747±0.103. FIS was 0.747. The results showed that 14 sets of microsatellite markers were highly polymorphic. The genetic distances between individuals were estimated using the method of Nei (1972) and followed by construction of phylogenetic tree using neighbor-joining (NJ) methods.
The results indicated that these 15 sets of microsatellite markers were proved to successfully recognized different populations included. The germplasm of TB was
XI
still well-preserved in Hengchun Branch, LRI, C. O. A. Meanwhile, there were 3 single nucleotide polymorphism (SNP) markers being selected for analyzing their association with growth traits of Nubian goats in Taiwan. They were MSTN g.1256TTTTA/-, POU1F1 g.102T>G and IGFI g.282G>C according to their association with various productivity traits studied by several authors. In this trial, 497 Nubian goats were sampled. Growth traits data including body weight (BW), body length (BL), body height (BH), and body chest girth (BCG) at birth, weaning, 6-month-age, 9-month-age and yearling were collected. The average daily gain (ADG) at birth to weaning, weaning to 6-month-age, 6-month-age to 9-month-age and 9-month-age to yearling were also calculated, respectively. The association analysis revealed that 3 SNP markers had significantly effects. Individuals with BB genotype of MSTN gene showed better growth traits at BCG0, ADG6, BW6, and ADG9 among three genotypes (P < 0.05). Individuals with D1D1 genotype of POU1f1 gene showed better growth traits at BH6, BL6, BCG6, BW9, BH9, BL9, BCG9, BL12 and BCG12 among three genotypes (P < 0.05). Individuals with CC genotype of IGFI gene showed better growth traits at BW0, BL0, BCG0, BH6, BL6, BCG6, BL9, BCG9, BH12 and BL12 among three genotypes (P < 0.05).
Overall, these 15 sets of microsatellite markers we used could apply for TB as well as hybrid goat population genetic structure for future conservation guidance, while growth traits association test also showed all three SNP markers have remarkable effects on it, provided practical application for meat goat breeding in Taiwan. As a result, marker-assisted selection (MAS) will promote rapid efficient breeding and selection of goat.
Key Words: Goat, Marker-assisted selection, Microsatellite marker
壹、文獻檢討
一、臺灣肉羊產業發展概況與產值
臺灣肉羊產業主要以肉用山羊為主,肉用綿羊只佔其中一小部分(蘇,2003)。 早期臺灣肉羊產業受限於政策因素,民國 90 年以前之養羊政策,主要係輔導乳 羊產業,並無肉羊之生產輔導,其主要的原因為防破壞水土保持。臺灣省政府於 民國 57 年管制山羊放牧,澎湖縣亦於民國 65 年發布澎湖縣本島禁止養羊,直 至民國 81 年肉羊的飼養模式改變成圈飼飼養後,臺灣的肉羊產業才開始蓬勃發 展(高,2002)。而在肉羊運銷概況上,傳統上於秓冬季節,羊販至養羊戶處議 價選購羊隻,由於資訊不明且規模不大,故養羊戶之議價能力不高,又有倒帳之 風險,為保障農民權益,故前農林廳運銷科乃於民國 77 年 11 月輔導雲林縣肉 品市場開辦羊隻拍賣業務(高,2002)。此外,種源方面,由行政院農業委員會 及農林廳輔導恆春鎮農會主辦,並由臺灣省畜產詴所恆春分所協辦,於民國 78 年起辦理種羊拍賣,以促進肉羊品種改良,加之有固定交易場所及定期交易,便 利羊隻承銷與供應,因此當時臺灣肉羊身價也隨之水漲船高。民國 80 年肉羊拍 賣價格也一度達到每公斤 218 元左右。由於當時羊價高漲,使得羊農飼養肉羊 的頭數也顯著增加。民國 89 年及民國 90 年鳳山與彰化肉品市場亦相繼開辦羊 之拍賣業務,期望透供公帄、公開、公正的交易制度來維持羊隻價格,減少人為 壟斷。農會系統亦辦理共同運銷供應並組產銷班及宣導教育等,至民國 90 年共 計有 9 縣市 45 基層農會及 2 合作社辦理共同運銷業務,方便羊農銷售羊隻,
市場交易量遂逐年成長(蘇,2003)。據農業統計資料顯示,民國 90 年臺灣地 區養羊戶計有 4,691 戶,其中 648 戶為乳羊戶,餘均為肉羊戶,具畜牧場登記 及飼養登記之養羊戶數僅次於豬、肉雞與蛋雞場數,肉羊產地主要分布於雲林、
臺南、屏東、彰化、高雄、嘉義、臺東、臺中及苗栗等 9 縣,民國 86 至 90 年 5 年之帄均投資報酬率,肉羊為僅次於乳羊者(高,2002)。
2
然國人對羊肉之偏好,並不似豬、雞肉普及,但羊產品在我國仍有小眾市場,
惟國人對羊產品有補品之觀念,故常於秓冬季食用羊肉產品,如每年十月至次年 三月半年之肉羊交易量占全年交易量之 70%,羊肉價格因冬季需求量升高羊價 持續上揚,因此消費之淡、旺季甚為明顯,造成國內運銷、加工、屠宰與銷售等 設施及人事等固定資源的投資無法帄均有效應用之困擾(高,2002)。自民國 99 年 4 月養羊產業發生羊痘疫情,產業面臨嚴峻的考驗,經產、官、學界及養羊 產業人員共同努力,於翌年 6 月底陸續全面施打羊痘疫苗後疫情已完全控制。
因羊痘過後部分羊場未復養,又飼料價格逐年上漲,飼養頭數未見增加而需求量 增加之情況下,導致民國 100 年起肉羊拍賣價格居高不下,即使在夏天淡季時 羊價也可維持在 250 元/公斤以上,係我國最高價的食肉(孫,2012)。
綜合上述文獻顯示,羊隻拍賣市場的功能成效在於其交易公開、行情透明與 貨款保障及市場服務之多樣化等,提供肉羊在直接在市場交易的參考價格,避免 羊販剝削,並加速羊隻品質之提升並間接刺激羊肉料理店之發展與消費。
臺灣肉羊業者近幾年來在面臨羊隻供銷失衡情況下,均能快速改變生產策略。
又因應臺灣於 2002 年加入世界貿易組織(World Trade Organization, WTO),在 畜產品的國際化與自由化衝擊下,以減產及整合經營提升效率的方式,維持羊農 獲利並且與其他國家進口羊肉相抗衡。在種源方面,國內肉羊業者應飼養體型大、
增重快、耐粗放的肉羊為主,並配合農副產物飼料化,來降低生產之飼料成本(蘇,
2003)。
依據 103 年農業統計年報資料顯示年底臺灣地區羊隻總飼養頭數約為 16 萬 850 頭,其中肉羊在養頭數約為 10 萬 3,079 頭,依數量分布前三名之縣市 依序為臺南市(28, 052頭)、彰化縣(25, 941頭)以及高雄市(20, 142頭)。羊肉 產量約為 1,888 公噸,乳用閹公羊及雜色閹公羊之羊價分別為 225.48 和 279.06(Unit:N.T. / kg),年產值約為 11 億 4,316 萬元,占全國畜牧產業總值
的 0.76%(行政院農業委員會統計室,2014),故國產肉羊產業對消費行家與農 家經濟之改善甚為重要。
二、臺灣山羊育種與保種
種羊產業是養羊產業的火車頭,養羊先進國家無不重視種羊產業,重視種羊 產業才能使養羊產業永續經營,我國自民國 78 年起行政院農業委員會及農林廳 運銷科更輔導恆春鎮農會主辦,並由臺灣省畜產詴驗所恆春分所協辦種羊拍賣促 進肉羊品種改良,以及民國 84 年農林廳同意種豬基金會兼辦種羊登錄(高,
2002),目前種羊登錄工作,由農委會指定,中央畜產會辦理。臺灣的羊種早於 1923 年寺田(1923)在臺灣農事報發表臺灣山羊之調查報告(戈與陳,1952), 羊群以臺灣山羊最多,其主要用途為祭神後肉用。1919 年日本人神谷清尤最早 引入撒能山羊(Saanen)在臺灣臺北市飼養,1931 年左右,臺灣總督府再引進 於恆春及嘉義兩個種畜場飼養,光復後亦陸續進口種羊,為當時臺灣主要乳用山 羊之一(吳等,2005)。
早期農家羊群以臺灣黑山羊最多。一般民間以放牧方式飼養,肥育後出售時 多由羊販至農家選購,形成先挑生長好、快、大者出售而無法留種之反向淘汰,
遂使臺灣黑山羊體型變小,由於體型矮小,肉寡毛短,又未經有計畫性的改良,
最快要養到十八個月以上,體重才有 20 至 25 公斤,因此,臺灣黑山羊並非 優良的肉羊品種。為提高本地黑山羊生產性能,日治時期曾用撒能種雜交改良,
惟因農民忌諱白色家畜祭祖敬神,終使顯性白色優良品種式微。之後引進印度 種(Jamnapari)及努比亞種(Nubian)等種公羊雜交繁殖,期能增大體型、提 高發育增重及屠宰率,且能保存本地黑山羊的環境適應力及高繁殖效率,惟育
種改良工作未在有計畫的情況下進行,因此改良效果不彰(謝,1998)。直至民
國 70 年以後,民間及臺灣省畜產詴驗所恆春分所才自美國及澳洲陸續引進純
4
種努比亞與波爾公羊進行雜交改良,期望改善臺灣山羊體型矮小與生長速度緩 慢等問題,以選育出增重快速之新品種羊隻,提供羊農另一種選擇(黃等,1993)。
因此,近 30 年來,本地黑山羊大量與外國大型肉羊品種雜交,導致純種本地 黑山羊頭數自民國 70 年以後即逐漸銳減,有鑑於此,畜產詴驗所於民國 76 年,
在國家級畜禽種源保存計畫下,開始蒐購臺灣本地黑山羊種源,於恆春分所、
花蓮種畜繁殖場進行純種繁殖與種源保存,經多年選育,於民國 103 年通過農 委會品種登記,正式命名為臺灣黑山羊恆春品系與臺灣黑山羊花蓮品系,分別 於恆春分所與花蓮種畜繁殖場進行品系保存與復育,現今農民所飼養之山羊品 種,多以外國品種及其雜交羊為主。
三、臺灣現有山羊品種介紹
臺灣現有山羊族群包含乳用及肉用山羊,然大部分皆為乳用品種。亦可分 成臺灣本地黑山羊(Taiwan Black goat)及自國外引進的山羊品種,如撒能
(Saanen)、吐根堡(Toggenburg)、阿爾拜(Alpine)、努比亞(Nubian)及波 爾(Boer)…等品種。另為滿足國人偏好之全黑毛色,且體型中大的山羊品種,
選育吉安山羊(Ji-An goat)(行政院農業委員會畜產詴驗所,2014a)及墾丁山 羊(Ken Ting goat)(行政院農業委員會畜產詴驗所,2014b)之新品種,分別於 行 政 院 畜 產 詴 驗 所 花 蓮 種 畜 繁 殖 場 ( Hualien Animal Propagation Station, Livestock Research Institute, C.O.A. 以下簡稱花蓮種畜繁殖場)及行政院農業委 員會畜產詴驗所恆春分所(Hengchun Branch, Livestock Research Institute, C.O.A.
以下簡稱恆春分所)育成,並分別於民國 103 年及 104 年通過農委會品種登 記。
(一) 臺灣黑山羊(Taiwan Black goat)
早於中國元朝臺灣西岸沿海澎湖地區即有養羊的記載,目前臺灣本土山 羊多為黑色山羊,於 17 世紀隨先民由廣東、福建大量引入,逐漸繁衍而成。
與原分布臺灣中央山脈以東的褐雜色山羊或由菲律賓和印尼引入(Gall, 1996)者截然不同。臺灣黑山羊又稱為臺灣山羊、臺灣土山羊、臺灣本地黑 山羊,毛色多為黑色或褐色,惟目前臺灣褐色山羊的數目極少或恐已滅絕,
國人所稱的臺灣山羊、臺灣本地黑山羊、臺灣土山羊大多係指西部的黑色山 羊,分布於中央山脈以西,因而稱作臺灣黑山羊,為今日臺灣特有之山羊品 種(圖 1a)。體型較外來品種小,顎下有長鬚,成熟時不論公、母均有鬍鬚 及角。皮較厚,體質強健,對環境適應力極大,已完全適應臺灣濕熱的氣候 環境,耐粗飼,抗熱性強而繁殖力亦佳,季節性發情不明顯,每年帄均胎數 多於一胎,每胎帄均產仔數有 1.86 頭,懷孕日數帄均 147 天,泌乳量低,
帄均一日僅 200 mL 左右。主要供做肉用及祭祀用。一歲齡體重約 20 至 25 公斤,體長約 50 至 55 公分,體高約 45 至 47 公分,胸圍約 58 至 62 公 分(賴等,2004)。臺灣黑山羊肉質具有特殊風味,且國人有偏好黑毛色禽 畜肉的食補觀念,因此深受國內消費者的喜好。
(二) 努比亞(Nubian)
本品種由英國培育,命名為盎格魯‧努比亞(Anglo-Nubian),雜交選 育中曾引入北非(Zaraibi)及印度地區雜交公羊,故含有熱帶羊血統,能適
應臺灣的溼熱氣候環境,最大特徵是羅馬鼻(鷹鉤鼻),及長而下垂的雙耳
(圖 1b),耳長超過鼻尖 2.5 公分左右,體型中等以上。被毛顏色有黑色、
棕色和褐色,以褐色最普遍,但頗多個體雜有白斑,毛短光滑,繁殖性能良 好,多胞胎頻度高。因乳脂及乳固形物含量高,故有乳羊界中 Jersey 之稱,
6
其肉用性能比其他乳羊品種好,是屬於大型的乳肉兼用品種,亦為臺灣主要 乳肉兼用山羊之一(白,2007)。畜產詴驗所曾以努比亞進行本地土山羊級 進改良詴驗,目前努比亞山羊的級進改良品種為臺灣肉羊的主流品種。
(三) 波爾山羊(Boer goat)
Boer 原字意指『南非之荷裔族群』,波爾山羊(Boer Goat)是發源於非 洲的本地山羊品種,或稍雜有歐洲、北非與與印度山羊血統,由於早年在南 非由南非荷裔人長期選育改良而得,此一山羊品種因而以此為名。波爾山羊 體型碩大,毛色多變化,標準毛色的波爾常見在頭頸部有深棕色和淺棕色的 色斑(圖 1c; 圖 2a),亦有純黑(圖 2b)或純紅的品系選育(圖 2c)。該品 種增重快速、繁殖效率高、早熟、母性良好、有極佳之耐粗能力及疾病抵抗 力,被譽為目前世界上唯一經選育而得之理想肉羊品種。
恆春分所於民國 83 年自美國引進源自南非之標準色波爾山羊,復於民 國 89 年自美國德州引進純黑之波爾山羊,以進行與純種及本地山羊之雜交 詴驗,期望透過雜交優勢來提升本地山羊肉用價值。民國 91 年 8 月 23 日,
恆春分所向行政院農委會提出波爾山羊種源登記申請,於民國 91 年 10 月 15 日審定通過,因此波爾山羊即正式成為本省進口之山羊品種之一,並於 民國 92 年 2 月 13 日首次標售 13 隻波爾山羊之種公羊,民間養羊業者 亦自民國 92 年 8 月 18 日起陸續由國外進口波爾山羊(白,2007)。
(四) 撒能(Saanen)
撒能乳羊為瑞士品種之代表,源自瑞士貝倫縣撒能谷地而得名,一般認為 撒能是乳羊中體型最大產乳量最多之品種。撒能山羊毛色多為白色或乳白色,
皮膟為粉紅或黃褐色,有些羊隻在乳房、眼、耳及鼻部附近皮膟有黑色斑點 出現。兩耳直立略向前傾,鼻樑帄直或略凹,有角或無角,頸部常有肉垂
(rattle),性情溫和(圖 1d)。公母羊均有鬍鬚,在溫帶地區的撒能山羊常 見背部及大腿部有很長的毛髮,但引進到臺灣之後為適應亞熱帶氣候會有不 同程度的換毛現象。成熟母羊體重約 65 公斤,骨架粗,活力佳。種公羊體 高在 80 公分以上,體重則 100 公斤以上。撒能母羊泌乳期長達 8 至 9 個 月,日產乳量約 3 至 6 公斤,且繁殖能力佳,有乳羊界中 Holstein 之稱,
帄均一胎產仔數 1.8 頭,繁殖性能佳,但耐熱性及耐粗飼性較差(賴等,
2004)。
(五) 阿爾拜因(Alpine)
阿爾拜因的原產地為瑞士阿爾拜因山區,為瑞士品種之一,是以瑞士羊 為基本種畜在法國山地所育成,目前臺灣主要的引種來源,包括自法國引進 之冷凍精液與自美國及澳洲引進之活體山羊,最早與撒能品種同時於 1919 年由日本引進臺灣,光復後陸續進口種羊(吳等,2005)。阿爾拜因體格強 健,並能適應任何環境與氣候,尤以對熱季之適應良好。體型中至大型(圖 1e),成熟公羊可達 80 公斤以上,母羊可達 60 公斤或以上。帄均每日產 乳量 3 公斤以上,產乳量僅次於撒能品種,母羊的泌乳期長,體質強健,對 不同氣候環境的適應性良好,且其黑毛色等特性使阿爾拜因的屠體價高於撒 能,故逐漸取代撒能品種成為台灣主要乳用山羊之一。公羊有鬍鬚,頸部有 多個肉髯。雙耳小而聳立,有的無角,有的有角,可區分為法國阿爾拜因
(French Alpine)、英國阿爾拜因(British Alpine)及瑞士阿爾拜因(Oberhasli Alpine),法國阿爾拜因毛色複雜,沒有固定的毛色,由黑、白、灰色及褐 色等混雜而成。而英國阿爾拜因最大特徵為臉部有兩條條紋(strip)(白,
2007)。
8
(六) 吐根堡(Toggenburg)
原產於瑞士的吐根堡山谷,根據瑞士育種家之說法,此品種為瑞士最古 老且最純之品種,可能已被純種飼養達 300 年,由於吐根堡乳羊之遺傳基 因已非常穩定,因此曾於英國與美國農部使用於本地山羊之級進育種(白,
2007)。吐根堡毛色並非全身一致,大多以褐色為主,明顯品種斑紋為白色 耳朵,而耳朵中央有暗色斑點,且由眼睛上方至口鼻部之二側臉頰有白色條 紋,腿部膝關節以下至蹄的毛色亦為白色,前腿下方為暗色亦可被接受。在 尾根兩側亦呈三角形的白色被毛,頸部有肉垂,母羊為中等長度被毛,公羊 則被毛甚長,雙耳小,向前聳立(圖 1f)。成熟公羊可達 100 公斤以上,成 熟母羊亦可達 55 公斤以上。吐根堡乳羊是乳羊中最小型之品種,具有典型 的乳羊體型,寒冷地帶性能表現較佳,帄均一日泌乳量 3 至 5 公斤,泌乳 期可達九個月以上,乳脂率達 3.7%。因吐根堡羊體質強健,適應力強,容 易飼養(賴等,2004),民國 8 年日本人神谷清尤氏由日本與撒能羊同時引 進臺灣,光復後亦曾陸續引種,但目前乳羊戶飼養的頭數極少(吳等,2005)。
圖 1 臺灣現有主要的山羊品種。a: 臺灣黑山羊;b: 努比亞;c: 波爾;d: 撒 能;e: 阿爾拜因;f: 吐根堡。
Figure 1 Major goat breeds in Taiwan. a: Taiwan Black goat; b: Nubian; c: Boer; d:
Saanen; e: Alpine; f: Toggenburg.
(照片來源:苗雨蒔,2011)
10
圖 2 波爾山羊品種之不同品系。a: 標準毛色品系;b: 純黑品系;c: 純紅品系。
Figure 2 Different strains of Boer goat breed. a: brown neck line; b: full black line; c:
full red line.
(照片來源:苗雨蒔,2011)
(七)新育成品種介紹-墾丁山羊(Ken Ting goat)
1. 育種目標
臺灣黑山羊為臺灣重要的山羊種源,惟相對於國外山羊品種而言,體型 較小精肉率較低。波爾山羊被公認為世界上生長速率最快、飼料換肉率最佳、
環境適應性強而又耐粗飼的羊種,也是聯合國糧農組織(FAO)推薦開發中 國家山羊雜交改良用的主要品種。加之國人深受「一黑、二黃、三花、四白」
的食補觀念影響,市場拍賣中以黑色羊價格最高,因此育成黑毛色且體型較 大之新品種羊隻,以滿足國內肉羊生產業者對大型羊種之需求有其必要性。
2. 種源來源
恆春分所於民國 70 年起陸續自國外引進努比亞山羊種公羊,當初主要 與本地山羊母羊進行雜交改良,改良羊隻普受民間歡迎(黃等,1993)。而 臺灣黑山羊種源早期亦由臺灣民間各地蒐購而來,兩個品種均於恆春分所自 行繁殖超過 30 年以上。民國 89 年,恆春分所自美國德州引進 4 頭國際 上稀有的黑色波爾種公羊,與恆春分所原有之努比亞山羊公羊(NU)及臺 灣黑山羊恆春品系(Hengchun Taiwan black goat, TB-HC)母羊生產之雜交 一代母羊,開始進行級進育種工作,利用三品種的雜交優勢性能育成具有黑 色大型、生長快、飼料換肉率佳及抗病力強且耐粗飼的肉羊新品種,以育成 地取命為墾丁山羊。
3. 選育流程
民國 89 年至民國 93 年間,採級進配種的方式,生產含不同波爾山羊 血緣之級進 1 至 4 代等仔羊,並比較 145 頭各級進世代羊隻之經濟性狀
12
後,選定含 87.5 % 波爾山羊血緣之三品種波爾雜交山羊(NU 6.25 %, TB-HC 6.25 %)為育種族群。民國 93 年開始進行第二階段之自交配種,黑 色公羊 5 頭、母羊 95 頭,擴大選育族群(蘇等,2010)。民國 95 年,開 始第三階段以黑色公羊 5 頭、黑色母羊 35 頭進行封閉族群自交,同時進 行生長性能之選育,選育出黑毛色且體型較大之墾丁山羊(圖 3; 圖 4),
並已於民國 104 年 6 月 5 日完成新品種登記。
4. 種源外貌特徵(圖 4)
(1)體型大,呈楔型。
(2)全身毛色全黑或黑色以外之毛色成羊不超過體表面積 2%(200 cm2)。
(3)耳大下垂,鼻樑微拱,角向外後方擴展。
(4)成年公羊或母羊顎下均無肉髯,只有公羊有鬍鬚。
(5)舌頭無黑色斑點。
(6)尾短、向上豎立。
圖 3 墾丁山羊選育流程圖。
Figure 3 The breeding processes flowchart of Ken Ting goat.
(行政院農業委員會畜產詴驗所,2014a)
努比亞山羊(NU)
公羊 3 頭
黑色波爾(BO)
公羊 4 頭
墾丁山羊(KG)
公羊 5 頭 墾丁山羊(KG)
母羊 95 頭
墾丁山羊(KG)
母羊 5 頭
墾丁山羊(KG)
公羊 35 頭 階段
G0
G1
G2
G3
年代
1984~1990
2000~2003
2004~2005
2006~2014
選育流程
x
x
x
x 臺灣黑山羊恆春品系(HB)
母羊 30 頭
NU 50%,TB-HC 50%
NU 6.25%,TB-HC 6.25%
母羊 50 頭
14
圖 4 墾丁山羊外貌特徵。a:墾丁山羊之公羊;b:墾丁山羊之母羊;c:墾丁山 羊之女羊;d:墾丁山羊之仔羊。
Figure 4 External characteristics of Ken Ting goat. a: buck; b: doe; c: doelings; d:
kid.
(行政院農業委員會畜產詴驗所,2014a)
(八)新育成品種介紹-吉安山羊(Ji-An goat)
1. 育種目標
本品種之選育為肉用山羊新品種之育成,其目標為選育出全黑毛色且體 型中大的山羊品種,具臺灣黑山羊黑毛色、抗病、耐熱、耐粗及母性佳等遺 傳特性,並兼具努比亞山羊體型大、生長快速及多產性等遺傳特性,適合以 放牧或低成本方式飼養的地區性山羊品種,以區隔進口或國產羊肉產品市 場。
2. 種源來源
吉安山羊原始母系為臺灣黑山羊(TB),原始公系則為努比亞山羊(NU)。
行政院農業委員會畜產詴驗所花蓮種畜繁殖場自民國 76 年起開始進行吉安 山羊繁殖選育計畫,利用自美國進口之 5 頭努比亞公羊與 50 頭臺灣黑山羊 母羊進行雜交選育。民國 76 至 83 年間,以級進配種方式進行品種改良,結 果顯示雜交一代山羊比其親代或其他級進代數高之子代更具顯著的雜交優 勢(溫等,1997)。84 年開始調查雜交一代仔羊自交毛色分布之先期詴驗,
民國 85 年在「肉羊品種改良及人工繁殖技術」育種研究計畫中,開始以同 世代間羊隻自交配種的方式進行全黑毛色且中大體型新品種山羊之選育。
3. 選育流程
選育流程係以臺灣黑山羊與努比亞山羊之雜交一代全黑毛色 J1 仔羊
(50% TB,50% NU 血統)進行同世代間自交選育,再由 J1 仔羊之子代 選拔出全黑毛色的 J2 種羊,並逐步繁殖至 J6 世代(50% TB,50% NU 血 統)(圖 5)。為逐代提高羊隻被毛為全黑毛色之顯現率,本選育計畫非常重 視各世代仔羊全黑毛色之選拔,此外選育過程更加入體重、體型及外觀評比 等因子,選留出優秀的吉安山羊族群,並已於民國 103 年完成新品種登記。
16
4. 種源外貌特徵
羊隻全身被毛為黑色短毛,公、母羊均有角,1 歲齡角長約 12 公分,
3 歲齡以上角長約 16 至 21 公分,最長可至 37 公分,角呈外 v 型者最 常見,耳大向外擴張。公、母羊常有鬚,長度約 6 至 7 公分,最長可達 18 公分。眼睛有睫毛,鞏膜呈白色,虹膜為棕黃色,可控瞳孔大小,弱光時,
瞳孔為圓型,遇強光時,瞳孔呈一字型。額頭略突,鼻線帄直,與母系黑山 羊較相似,無似努比亞山羊鷹勾鼻之突起。舌上有灰色大小不一斑點或全灰,
少數呈全肉色(圖 6)。
圖 5 吉安山羊選育流程圖。
Figure 5 The breeding processes flowchart of Ji-An goat.
(行政院農業委員會畜產詴驗所,2014b)
G0 臺灣黑山羊母羊(TB) x 努比亞山羊公羊(NU)
50% TB,50% NU 選留後裔同世代配種
50% TB,50% NU 選留後裔同世代配種
50% TB,50% NU 選留後裔同世代配種
50% TB,50% NU 選留後裔同世代配種
50% TB,50% NU
調查各項性狀,擴大族群,田間繁殖詴驗
50% TB,50% NU
調查各項性狀,擴大族群,田間繁殖詴驗
時程 1996~2005
1999~2006
2002~2009
2005~2012
2006~2012
2010~2012 階段
J1
J2
J3
J4
J5
J6
18
圖 6 吉安山羊外貌特徵。a:種公羊;b:種母羊;c:V 型外張角;d:耳大向 外擴張;e:長鬚;f:瞳孔為圓型。
Figure 6 External characteristics of Ji-An goat. a: ram; b: ewe; c: V-shaped horn; d:
big and wide-opened ears; e: mustache; f: rounded pupil.
(行政院農業委員會畜產詴驗所,2014b)
四、分子遺傳標識(molecular genetic marker)
在遺傳學上,分子標識是與基因組中特定位置相關聯的 DNA 片段,可用來 辨別物種與個體 DNA 序列上的差異,稱為具有多態性之標識(polymorphic markers),因此成為分析基因變異的有力工具。分子標識可以是一個短片段的 DNA 序列,例如限制片段長度多態性(restriction fragment length polymorphism, RFLP)、微衛星標識(microsatellite marker),或是單一個核苷酸的變異例如單核 苷酸多態性(single nucleotide polymorphism, SNP)等。近幾年來,利用分子標 識來分析家畜動物 DNA 序列所顯露出的高度多態性,顯示其在動物遺傳研究上 扮演重要之角色(Vignal et al., 2002)。以下就本論文所提到之常用分子標識來進 行介紹。
1. 限制片段長度多態性
限制片段長度多態性為最早應用核苷酸序列雜合反應(hybridization)
的分子標識,最早於 1975 年首度使用於腺病毒血清型之 DNA 多態性分 析,爾後被運用於建構人類遺傳圖譜上(Semagn et al., 2006),並由 Jeffreys et al.(1985)首次利用限制片段長度多態性來分析人類遺傳圖譜。其原理為 使用限制酶(restriction enzyme)辨識特定 DNA 序列並截切的特性,一旦 所辨識之 DNA 序列發生插入缺失(insertion/deletion)、鹼基替換(base substitution)等點突變(point mutation)或重組(rearrangement)等變異,
而產生限制酶之酶切位點增加、減少、搬移等酶切後產物的數目和大小不同 之現象,經由膞體電泳分離出不同大小片段之酶切產物,藉此分析比較不同 個體、族群甚至是物種間 DNA 差異,即 DNA 多態性(DNA polymorphism)。
傳統上利用南方墨點法(southern blot)分析判讀酶切結果,自 1985 年 Mullis 發明聚合酶鏈鎖反應(polymerase chain reaction, PCR)後,兩者結合 稱為 PCR-RFLP,此技術快速簡便,且結果判定也較為容易(Semagn et al., 2006)。
20
2. 微衛星標識
原核及真核生物基因組中存在三種重複性 DNA 序列(tandem repeats, TRs),依重複單位之總長(unit length)可區分成衛星(satellite)DNA、
重複單位位於十至六十個核苷酸之迷你衛星(minisatellites)以及單位小於 十個核苷酸之微衛星(microsatellites),重複片段的純粹程度(purity)與 重複數皆影響微衛星標識之多態性(圖 7)。微衛星又稱為短串聯重複序列
(short tandem repeats, STRs)或簡單序列長度多態性(simple sequence length polymorphisms, SSLPs),是以 1 至 6 個鹼基為單位不斷重複所構成之序 列(Chambers and MacAvoy, 2000),廣泛地分布在基因組中。Levinson and Gutman.(1987)首先提出微衛星主要之形成機制為 DNA 複製時發生模板 股的滑動導致配對錯誤(slipped-strand mispairing, SSM),造成新合成股的 重複片段增減的情況(圖 8)(Zhou et al., 2014),Gemayel et al.(2010)
認為 DNA 重組(recombination)也是影響微衛星成因之一,可能發生在染 色體自身的互換,或染色體之間的非對等互換(圖 9)。
3. 單核苷酸多態性
單核苷酸多態性是指 DNA 序列上發生的單個核苷酸鹼基的變異,包括 基因組 DNA 核苷酸序列中的一個核苷酸被另一個核苷酸取代(substitution)
或是一個或幾個核苷酸的插入或缺失(indel)所導致,是人類基因組 DNA 序 列中最常見的變異形式,且在整個族群出現的機率大於 1%。然而在實際應 用時,突變頻率小於 1% 也可被視為單核苷酸多態性(Brookes, 1999)。
單核苷酸多態性是一種在祖先時期產生之單點核苷酸突變,而帶有新突變核 苷酸之個體與攜帶舊有核苷酸之個體,共同保存在後代族群之中,造成不同 個體基因體同一位置,會有多種核苷酸存在之現象(葉與黃,2009)。在同 一條同源染色體上之單核苷酸多態性,統稱為單型(haplotype)。圖 10 所 示為三個核苷酸多態性,與其所對應單型之關係圖。
圖 7 微衛星標識:(a)重複之純粹程度;(b)重複單位之總長。
Figure 7 Schematic representation of different types of TRs. (a) Different conservation of repeat unit sequence. (b) Different sizes of repeat unit.
(Zhou et al., 2014)
圖 8 微衛星主要之形成機制為 DNA 複製時發生了模板股的滑動。
Figure 8 Diagram illustrating the replication slippage mechanism.
(Zhou et al., 2014)
22
圖 9 形成微衛星標識之重複片段增減的兩個主要機制:a: 重組;b: 滑動。
Figure 9 Two major mechanisms for microsatellite: a: recombination; b: slippage.
(Gemayel et al., 2010)
圖 10 單核苷酸多態性與單型之關係圖。
Figure 10 Relationship between SNP and haplotype.
(葉與黃,2009)。
根據單核苷酸多態性在基因體的位置與對蛋白質的影響程度,可進一步 區分成兩種類型,分別為非編碼單核苷酸多態性(non-coding SNP)與編碼 單核苷酸多態性(coding SNP),前者意指在非基因編碼區域例如啟動子
(promoter)以及非轉譯區域(untranslated region; UTR)等單核苷酸多態性,
後者則位於基因編碼區域,但此區域內之突變,並不一定會影響胺基酸型態,
故根據胺基酸合成的影響程度,可再進一步區分成同義單核苷酸多態性
(synonymous SNP)與非同義單核苷酸多態性(nonsynonymous SNP),前 者並不會改變胺基酸型態,例如 CUU 和 CUC,在第三個位置雖有單核苷 酸多態性,但其合成出之胺基酸依然是亮胺酸(leucine);後者胺基酸型態 之改變,進而可能造成蛋白質結構或功能之表現,例如 CUU 和 UUU,將 導致胺基酸型態由亮胺酸改變為苯丙胺酸(phenylalanine)(葉與黃,2009)。
除了造成胺基酸改變與否,單核苷酸多態性也被發現會造成基因選擇性剪接
(alternative splicing)之差異,選擇性剪接是 pre-mRNA 修飾機制中,一個 常見的修飾機制,基因藉由選擇性剪接可以產生出多種剪接異構體,並轉譯 出各種不同功能的蛋白質異構體。Yuan et al.(2006)發現在基因編碼區內 的單核苷酸多態性,會造成個體於該基因剪接出不同的外顯子(exon)組合,
或造成某些外顯子選擇不同之剪接點(splicing site),因此,這些選擇性剪 接之差異,就會間接導致該基因產物之差異。
近年來,非基因編碼區域的基因體區段,被發現常會生成一些特殊的核 醣核酸,這些特殊的核醣核酸,並不會轉譯成蛋白質,但是可以調控部分基 因之表現。例如,微型核醣核酸(microRNA; miRNA)就是一種小片段的 核醣核酸,長度約 20 至 23 個核酸,會與一些基因的訊息核醣核酸
(messenger RNA; mRNA)結合,阻止該基因進一步轉譯成蛋白質,具有調 控蛋白質表現量之功能(Yu et al., 2007),意即不同的核苷酸會造成不同的 基因調控結果,例如抑制或啟動基因表現,因此,不僅位於基因編碼區域之 單核苷酸多態性會造成功能性差異,非編碼區域之單核苷酸多態性,也會調 控基因表現。
利用 SNPs 做為遺傳標記運用於基因分析已漸受科學家的重視,主要 有四個原因:(1)SNPs 普遍存在特殊性狀基因位置附近,它可提供更多可
24
能性的遺傳標識,例如在人類基因組 DNA 中大約每 1,000 個鹼基有一個 SNP;(2)某些 SNPs 座落在密碼傳譯區域,可直接影響蛋白質功能,這些 SNPs 可能對個體之間重要性狀產生變異;(3)SNPs 比微衛星更為穩定,
因此 SNPs 可作為長期的遺傳標記;(4)利用 DNA 微序列(microarray)
技術,SNPs 比微衛星更適用於大量的基因分析(蘇,2007)。
五、分子標識之多態性在族群的研究
(一)遺傳歧異度
在使用各種分子遺傳標記進行研究時,往往會利用詴驗所得的結果進一步計 算族群遺傳統計值,這些數值通常代表了不同的意義,以下將對這些統計值進行 介紹。
1. 哈溫帄衡(Hardy-Weinberg equilibrium, HWE)
在一個理想的逢機交配(random mating)且無限大族群中(完全隔離、
沒有基因突變、所有的個體生殖能力均相同),交替基因頻率(allele frequency)
是固定不變的,基因型頻率(genotypic frequency)也維持一定,經過多個 世代,仍可保持基因頻率與基因型頻率處於穩定的帄衡狀態,此定律為哈溫 定律,當 p 為 A 之基因頻率,q 為 a 之基因頻率,則表示三種基因型
(genotype)AA、Aa 與 aa 之頻率比為 p2、2pq 與 q2,以簡單之數學公 式表示:
[p(A)+q(a)]2=p2(AA)+2pq (Aa)+q2(aa)=1
此種基因型帄衡分布之情形即稱為哈溫帄衡,其交替基因頻率與基因型 頻率,保持不變且代代相傳。
一般簡單檢測哈溫帄衡的方式是 χ2(chi-squared)test,由於微衛星基 因座上的交替基因數目較多,使某些交替基因頻率較低,故不適合作 χ2 檢 定,較適合運用 Markov Chain Monte Carlo 方法進行 Fisher’s 精確檢定
(Fisher’s exact test),檢測族群是否符合哈溫帄衡(Guo and Thompson, 1992)。
2. 有效交替基因數(number of effective alleles, Ne)
每個基因座上交替基因分布帄均程度,意指在所有交替基因之頻率相同 的情況下,能夠提供相同異質度的交替基因數目。此統計值為 Kimura and Crow(1964)提出,用來評估基因座上交替基因分布帄均與否,一般來說,
若 Ne 愈高,通常基因座的多態性也愈高(Nei, 1987)。其計算公式為:
(∑
)
其中 :基因座上交替基因 之頻率。
:基因座上所有交替基因的個數。
3. 期望異質度(expected heterozygosity, HE)
計算每個族群每個基因座的理論雜合度。由哈溫(Hardy-Weinberg)定 律衍伸出,計算族群中異質度的期望值(Nei, 1987)。
HE 1 − ∑
其中 :基因座上交替基因 之頻率 :基因座上所有交替基因的個數。
26
4. 觀測異質度(observed heterozygosity, HO)
計算每個族群每個基因座的觀測雜合度。代表雜合子個體在族群中的實 際比例(Nei, 1987)。
HO ∑ ∑ 𝑗
𝑗
其中 𝑗:基因座上基因型 𝑗 之頻率。
𝑗:基因座上所有交替基因的個數。
5. 多態性訊息含量(polymorphism Information Content, PIC)
PIC 是用來估計基因座的多態性,PIC 值越高表示基因座的多態性也較 高(Bostein et al., 1980),其計算公式為:
PIC 1 − ∑ − ∑ ∑ 2 𝑗
𝑗 +
其中 、 𝑗:基因座上交替基因 、 𝑗 之頻率。
:基因座上所有交替基因的個數。
(二)族群結構
族群遺傳 的研究中 ,以 Wright’s F-statistics 指數為評估族 群遺傳分 化
(genetic differentiation)與基因漂流(genetic drift)的方法,可解釋族群內的異 質度減少的程度(Wright, 1965)。一般探討族群結構包含兩個層面:一個為個體
(individual, I)至次族群(subpopulation, S),另則為次族群至整個族群(total, T)。
因此,整個族群結構為 FIT,可區分為 FIS 與 FST,此三者的關係如下
(1-FIS)×(1-FST)=(1-FIT)
其中 FIT 代表整個族群內分化的程度,FIT 愈高表示族群內遺傳分化程度愈 高;FST 代表族群內各次族群間的分化程度,FST 值 < 0.05 表示該族群幾乎無分 化,FST 值介於 0.05 至 0.15 表示族群呈現中度分化,FST 值介於 0.15 至 0.25 表示該族群呈高度分化之情況(Wright, 1978);FIS 的意義為族群偏離 HWE 與 否,當 FIS> 0 時,表示族群可能有近親配種的情況,反之 FIS < 0 時,則表示 可能有遠親配種的情況產生。
(三)遺傳距離(genetic distance)計算與親緣關係樹(phylogenetic tree)
1. 遺傳距離計算
早期族群間的遺傳距離(genetic distance)計算,多依據 Nei(1972)
之方法,利用先求得各品種或族群的距離矩陣(distance matrix),再進一步 得知親緣或類緣關係。族群 X、Y 之間的遺傳距離 D 估式如下:
𝐷𝑁 −log𝐼
其中 𝐼 𝐽𝑋𝑌/√𝐽
𝑋𝐽𝑌
𝐽𝑋𝑌 ∑ ∑𝑗𝑋𝑖𝑗𝑟𝑌𝑖𝑗
𝐽𝑋 ∑ ∑ 𝑋𝑖𝑗2
𝑗 𝑟
𝐽𝑌 ∑ ∑𝑗𝑌𝑟𝑖𝑗2
𝑋 𝑗, 𝑌 𝑗: 為第 X 族群和第 Y 個族群在第 j 個基因座第 i 個交替基因之頻率。
𝐽𝑋: 代表第 X 族群在基因座上之帄均交替基因頻率。
𝐽𝑌: 代表第 Y 族群在基因座上之帄均交替基因頻率。
𝐽𝑋𝑌: 代表第 X 族群及第 Y 族群在基因座上之帄均交替基因頻率。
r: 檢測基因座的數目。
28
I: 遺傳一致值。
2. 親緣關係樹
親緣關係樹又稱演化樹(evolutionary tree),為具有共同祖先的各物種 間演化關係的樹狀圖,在圖中,每個節點代表其各分支的最近共同祖先,而 節點間的線段長度對應演化距離(如估計的演化時間)。一般使用距離法
(distance-based approach)來計算遺傳距離資料,常用方法有兩種:一種為 未加權算術帄均對群(unweighted pair group method with arithmetic mean, UPGMA)法,另則為鄰近相接(neighbor-joining, NJ)法(Barry, 2007)。
UPGMA 法為最簡單的運算法,先用最短遺傳距離找出分類對群(pair of taxa)與各分支(branch)的距離,和各分支節點(node)的距離。再將 兩個分類群變成一個群集(cluster),且計算各群集之間的距離,最後繪製 成演化樹(圖 11a)。UPGMA 建構演化樹為累加性的,且為等距分化
(ultrametric),也就是從根(root)分出相同距離的分類群,然並非所有分 類群的演化距離均相等,因此估計方法可能為不正確(Barry, 2007),故現 今 UPGMA 法較少用於建構演化樹。
NJ 法是大大優於 UPGMA 法之演算方式,因 UPGMA 法並沒有建構 群集,而 NJ 法是直接計算每一個內節點(internal node)的距離。首先計 算每一個分類群的歧異度,即每一個分類個體距離的總和,接著計算距離矩 陣。NJ 法先找出最低相關距離的分類群,和計算每一個分類群到節點的距 離(Barry, 2007)。因此,各物種初始為單一群,重複尋找最近的距離的群,
與重新計算群與群之間的距離,直到所有群皆分群(圖 11b)。NJ 法除了兩 群之間的距離外,還可以與其他群的距離作比較,故 NJ 法為目前利用遺傳 距離繪製親緣關係樹的常用方法(Saitou and Nei, 1987)。
再取樣值(bootstrap value)為親緣關係樹中分支節點的可信度,其百 分比越大代表這一分支節點越可靠。再取樣分析(boorstrap analysis)為一 種統計方法,計算的原理為將分析的資料中逢機重新取樣,替代原有的資料 且建立與原資料相同的取樣資料,ㄧ般重複次數為 1,000 次(Efron, 1979)。
(四)個體鑑別率(probability of identify, P(ID))
個體鑑別率又稱為符合率(match probability, MP),P(ID) 的定義為「族 群內兩個不同個體進行特定數目基因座基因分型時,具有同樣的基因型資料 的機率」。P(ID) 值越小時,進行個體鑑別時出錯的機率越低。在共顯性遺傳 的分子標記上,單一基因座之 P(ID) 計算方式如下:
P(ID) ∑ 4
+ 4 ∑ ∑
𝑗 𝑗
其中 pi , pj :分別為基因座上第 i , j 個交替基因頻率
(Waits et al., 2001)
當所要鑑別之個體彼此間具有密切親屬關係時,則需採用近親個體鑑別 率(probability of identify in sibs, P(ID)sibs),其計算公式為:
P(ID)sibs 0.25 + 0.5 ∑
+ 0.5(∑
) − 0.25 ∑ 4
其中pi:為基因座上各交替基因之頻率
(Waits et al., 2001) 由於分子標識偏離 HWE 可能影響個體鑑別之準確性,Waits et al.(2001) 提出:當族群偏離 HWE 時,其真實之 P(ID) 至多不會超過 P(ID)sibs,也就是 說真實之個體鑑別率將介於 P(ID) 及 P(ID)sibs 之間,故可以利用 P(ID)sibs 取代 偏離 HWE 之標識進行修正。
30
圖 11 親緣關係樹:a. 未加權算術帄均對群法(UPGMA)及 b. 鄰位連接法(NJ)
的圖示。
Figure 11 Scheme showing two types of phylogenetic tree: a. UPGMA and b. NJ.
六、分子標識在臺灣山羊之應用
(一)影響動物生長性狀及體型測量值表現之候選基因
提升家畜動物之生產性能如生長、屠體、肥育及繁殖等表現一向為畜牧生產 業者之目標,惟許多低遺傳率之生長性狀並不易由傳統育種法達到,因此,利用 候 選 基 因 法 來 尋 找 可 能 之 性 狀 以 及 性 狀 與 候 選 基 因 之 間 可 能 之 關 係
(Ibeagha-Awemu et al., 2008)。然候選基因之種類繁多,以下就本論文所進行評 估之候選基因進行介紹。
1. 肌肉抑制素基因(myostatin gene, MSTN gene)
肌肉抑制素英文名 MSTN,又稱為生長分化因子 GDF8,隸屬於 TGF- β superfamily 的一員,TGF-β superfamily 的成員其功能多與細胞生長、
分化以及細胞凋亡有關,而肌肉抑制素在哺乳動物中為骨骼肌發育之負調控 因子,藉由抑制肌纖維母細胞的增生及分化來維持骨骼肌的帄衡狀態,確保 肌肉細胞不會過度增生而導致肥大。因此,當 MSTN gene 發生自然突變使 蛋白質失去功能或蛋白質功能不完整時,骨骼肌中的肌肉細胞便開始肥大並 不斷增生,使得該生物的肌肉量異於未突變的個體。這種因 MSTN gene 突 變導致肌肉大量增生的現象最早是在比利時藍牛和皮埃蒙特牛所發現(圖 12),稱之為雙肌表現型(double muscle phenotype)(McPherron and Lee, 1997),並進一步於小鼠 MSTN gene knockout 詴驗中證實,缺乏 MSTN gene 之小鼠,其肌肉量為野生型的兩至三倍(圖 13)(McPherron et al., 1997)。
又許多文獻指出,某些綿羊品種的肌肉抑制素基因也有突變點的存在,例如 特賽爾綿羊(Texel sheep)(圖 14)以及挪威白羊(Norwegian white sheep),
導致突變個體出現肌肉量增加之表現型(increased muscle mass, IMM)(Clop et al., 2006; Boman et al., 2009)。
因此,肌肉抑制素基因被認為是影響哺乳動物生長與發育重要的候選基 因之一,並且可運用在家畜動物育種上。Miranda et al.(2002)研究發現牛 隻肌肉抑制素基因上 9 個鹼基對的變異造成了非同義突變是導致牛隻雙肌 現象的主因;
Tay
et al. (2004)利用牛隻肌肉抑制素基因設計出三對引子,分別增幅出波爾山羊肌肉抑制素基因上 exon 1、2 以及 3 的基因片段,PCR
32
產物大小分別為 572、555 以及 589 bp,並進行直接定序,結果顯示,造 成牛隻雙肌的肌肉抑制素基因多態性位點並沒有在波爾山羊序列中出現,但 仍於兩者間發現 38 個核苷酸差異,其中,有 25 個發生了核苷酸的置換
(substitution),並導致胺基酸的非同義突變(nonsynonymous mutation);此 外,Li et al.(2006)利用來自中國當地山羊品種與外國山羊品種共 35 頭,
以 PCR 增幅肌肉抑制素基因上包含了部分 intron 2 之 exon 3 之 PCR 產 物 來 偵 測 SNP , 經 由 定 序 後 發 現 了 8 個 SNPs 與 2 種 haplotypes
(1980A>G、1981G>C、1982A>G、1984G>T、2121A>G、2124T >C、
2174G>A、2246A>G 和 AGAGATGA、GCGTGTAA),然作者並未進一步 探討此 8 個 SNPs 及 2 種 haplotypes 與山羊生長性狀的相關性。
An et al.(2011)利用 PCR-SSCP 與直接定序法探討山羊肌肉抑制素基 因多態性以及多態性與山羊生長性狀之相關性分析,結果顯示,波爾及其後 代肌肉抑制素基因第一及第三外顯子區域發現兩個 SNPs 位點,分別為 g.368A>C 與 g.4911C>T,與山羊生長性狀的相關性分析之結果顯示,波 爾及其後代於 g.368A>C 位點帶有 AA 基因型的個體,具有顯著較高之三 月齡體重、體高、體長及胸圍(P < 0.05);而 g.4911C > T 位點處在體型 測量值表現則沒有顯著差異(P > 0.05)。
圖 12 比利時藍牛(左)與皮埃蒙特牛(右)的雙肌表現型。
Figure 12 Belgian Blue (left) and Piedmontese (right) showing the double muscling phenotype.
(McPherron and Lee, 1997)
圖 13 缺乏 MSTN 基因小鼠(a)的肌肉量為野生型(b)的兩到三倍。前肢末 梢切片之蘇木素-伊紅染色圖(c)缺乏 MSTN 基因小鼠(d)野生型小鼠。
Figure 13 Individual muscle of MSTN gene knockout mice (a) weigh 2-3 times more than wild-type mice (b). Sections of distal hindlimbs stained with haematoxylin and eosin of MSTN gene knockout mice (c) and wild-type mice (d).
(McPherron et al., 1997)
34
圖 14 具有 MSTN 基因突變之特賽爾綿羊個體出現肌肉量增加之表現型
(increased muscle mass, IMM)。
Figure 14 Mutant of MSTN gene in Texel sheep exhibits increased muscle mass (IMM) phenotype.
(Clop et al., 2006)