台灣與鄰近島嶼瓊崖海棠族群遺傳變異 與親緣關係之研究
鄧書麟1,3) 傅昭憲1) 張坤城2) 楊倩如2) 黃瓊逸1)
摘 要
瓊崖海棠(Calophyllum inophyllum L.)為藤黃科(Clusiaceae)瓊崖海棠樹屬(Calophyllum)植物,廣 泛分布於東南亞、印度、琉球、澳洲與太平洋諸島;台灣係位於本種之邊際分布區域,產於恆春半島 與蘭嶼等地區。瓊崖海棠為海岸防風樹種之一,種子榨油可為化妝品及藥用,更可研發成生質能源,
經濟及國土保安價值高。本研究以取自台灣、蘭嶼、東沙群島與日本琉球群島等共4個地區,8個族 群,計79個瓊崖海棠樣株,應用簡單序列重覆分子指紋技術(inter-simple sequence repeat, ISSR),來探 討台灣與鄰近島嶼瓊崖海棠族群之遺傳結構及各族群間的親緣關係,共使用14個引子,獲得159個多 型性條帶。POPGENE分析結果總基因歧異度(H)為0.2021,族群間之遺傳分化係數(Gst)為0.6385,基 因流(Nm)為0.2832。由族群分子變方分析(analysis of molecular variance, AMOVA)結果,島嶼間之變 方成分為47.87% (p < 0.001),島嶼內族群間之變方成分為14.53% (p < 0.001),族群內個體間之變方成 分為37.60% (p < 0.001),顯示在不同島嶼與族群間已有分化趨勢。依歸群及主座標分析結果可區分為 4大群,台灣、蘭嶼、東沙群島與日本琉球群島的族群均各自集成群。透過Mantel test結果顯示,族群 間地理距離與遺傳距離並不顯著(r = 0.78,p = 0.99),各族群呈隨機散佈,遺傳漂變在族群分化上扮演 重要角色。綜言之,瓊崖海棠族群間基因流並不暢通,不同族群間已呈分化現象,且依歸群分析所得 各族群之群聚現象,亦佐證不同島嶼間已有顯著分化,推測應受島嶼隔離之影響,導致基因流受阻並 產生遺傳漂變所致。此外,瓊崖海棠族群總體的基因歧異度相較於其他濱海植物相對較低,因此建議 除應採取就地保育之措施來保存現有瓊崖海棠族群外,更需進行遷地保育來保護。
關鍵詞:瓊崖海棠、族群遺傳變異、遺傳歧異度、基因流。
鄧書麟、傅昭憲、張坤城、楊倩如、黃瓊逸。2017。台灣與鄰近島嶼瓊崖海棠族群遺傳變異與親緣關 係之研究。台灣林業科學32(2):145-57。
1)林業試驗所中埔研究中心,60081嘉義市文化路432巷65號 Taiwan Forestry Research Institute, No.
65, Lane 432, Wunhua Rd., Chiayi 60081, Taiwan.
2)嘉義大學林業暨自然資源研究所,60004嘉義市學府路300號 Department of Forest, National Chiayi University, 300 University Rd., Chiayi 60004, Taiwan.
3)通訊作者 Corresponding Author, e-mail:[email protected]
2016年4月送審 2016年7月通過 Received April 2016, Accepted July 2016.
Research paper
Population Genetic Variations of Calophyllum inophyllum in Taiwan and on Nearby Islands
Shu-Lin Deng,1.3) Chao-Hsien Fu,1) Kun-Cheng Chang,2) Chien-JuYang,2) Chiung-Yi Huang1)
【Summary】
Calophyllum inophyllum L. belongs to the genus Calophyllum (Clusiaceae) which is widely distributed in Southeast Asia, India, Okinawa (Japan), Australia, and the Pacific Islands. Taiwan is located at the edge of the species’ distribution area, and the species is mainly distributed on the Hengchun Peninsula and Lanyu. C. inophyllum is one of the coastal windbreak trees, and is seed oil can be extracted and used for cosmetics and medicinal purposes and also developed into bio- mass energy; so, it possesses high value for landscping and economics. In this study, genetic varia- tions and the population genetic structure of 79 individuals sampled from 8 populations in Taiwan, Lanyu, the Pratas Island, and Okinawa (Japan), were examined using inter-simple sequence repeat (ISSR) markers. For a total, 14 primers were used and 159 polymorphic sites were detected. A POPGENE analysis revealed that the total gene diversity (H) was 0.2021, the genetic differentia- tion index (Gst) was 0.6385, and the gene flow index (Nm) was 0.2832. An analysis of molecular variance (AMOVA) revealed that the variance component between population regions was 47.87%
(p < 0.001), that of populations within regions was 14.53% (p < 0.001), and the among individuals within populations was 37.60 (p < 0.001). The results indicated significant genetic differentiation among islands and among population within island. In addition, the Mantel test showed that there was no significant relation between the genetic distance and geographic distance (r = 0.78, p = 0.99), pointing to a random distribution among populations. In conclusion, the gene flow among popula- tions was relatively restricted, and populations on different islands showed significant genetic dif- ferentiation. Based on results of the UPGMA cluster analysis, the clustering of populations proved significant genetic differentiation among islands. It wasspeculated that geographic segmentation may be a significant factor in the genetic differentiation among the 4 regions. In addition, the ge- netic diversity of C. inophyllum is relatively low compared to those of other coastal plants. Based on the above findings, both in-situ and ex-situ conservation approaches are necessary for this species.
Key words: Calophyllum inophyllum L., population genetic variation, genetic diversity, gene flow.
Deng SL, Fu CH, Chang KC, Yang CJ, Huang CY. 2017. Population genetic variations of Calo- phyllum inophyllum in Taiwan and on nearby islands. Taiwan J For Sci 32(2):145-57.
緒 言
瓊崖海棠(Calophyllum inophyllum L.)係 屬藤黃科(Clusiaceae)瓊崖海棠樹屬之常綠大
喬木,廣泛分布於東南亞、印度半島、澳洲與 太平洋諸島(Liu et al. 1994)。台灣則位於本種
之 邊 際 分 布 區 域 , 產 於 恆 春 半 島 與 蘭 嶼 等 地 區,為海岸防風林樹種之一,因樹形優美,樹 性強壯耐瘠,亦為良好之園林樹。其木材質地 細膩,具有特殊紋理,可用於傢具、建築及造 船;種子富含油脂,可作藥物、化妝品之原料 及生質燃料(Friday and Okano 2006, Ong et al.
2011)。此外,Patil et al. (1993)從瓊崖海棠中 分離的香豆素(coumarins)化合物inophyllum B 和inophyllum P,具顯著的抑制HIV (人類免疫 缺陷病毒)逆轉錄酶活性;Itoigawa et al. (2001) 從瓊崖海棠中分離到的pancibiflavonol等黃酮類 化合物也顯示出抗腫瘤活性,故本種實兼具經 濟栽培及國土保安之價值。
瓊 崖 海 棠 果 實 能 漂 浮 於 水 上 , 和 蓮 葉 桐 (Hernandia nymphiifolia (Presl) Kubitzki.)、
棋盤腳樹(Barringtonia asiatica Vidal.)及銀葉 樹(Heritiera littoralis Dryand.)等均屬於海漂 植物,其特性可抵抗強風及水淹環境。但近年 來由於台灣海岸林遭過度開發及經濟發展利用 等因素,不但適生環境遭破壞,原生族群數量 也大幅縮減,早已被列為墾丁國家公園內之稀 有植物(Hsu et al. 1985)。因此,為加強本物種 之保育與推廣,實應就此物種積極建立基礎資 料,以供後續研究之參考。
近 年 來 由 於DNA指紋分析技術的快速進 展,除可檢測遺傳物質的變異外,更可獲致大 量遺傳變異訊息,較之以往利用蛋白質序列及 化 學 分 析 為 基 礎 的 方 法 更 佳 , 更 彌 補 了 使 用 同 功 酶 分 析 時 其 基 因 座 數 目 偏 低 的 缺 憾 。 由 Zietkiewicz et al. (1994)所發展的簡單序列重覆 分子指紋技術(inter-simple sequence repeat;簡 稱ISSR),所得之產物比逢機擴增多型性DNA (random amplified polymorphic DNA, RAPD) 具更高的重覆性和穩定性,符合Mendelian遺 傳定律,因此可得到較高可信度的遺傳多型性 資料。加上ISSR在引子設計上比SSR (simple sequence repeats)簡單,無需知道DNA序列即 可運用引子進行擴增,且其基因座具中性演化 (neutral evolution)內涵,演化速率較快,可提 供更高的族群變異估值,故已被廣泛運用於植 物遺傳多樣性與親緣關係之研究上(Wei et al.
2008, Deng et al. 2014, Hsieh et al. 2014)。本研 究應用ISSR來分析台灣與鄰近島嶼瓊崖海棠族 群的遺傳變異,除探討其族群遺傳結構及族群 間親緣關係外,同時依據其族群遺傳結構特徵 提出有效的保護措施與經營策略,俾供後續研 究與保育推廣之參考。
材 料 與 方 法
一、植物材料
針對台灣、蘭嶼、東沙群島及日本琉球群 島等4個地區,計8個瓊崖海棠族群進行取樣,
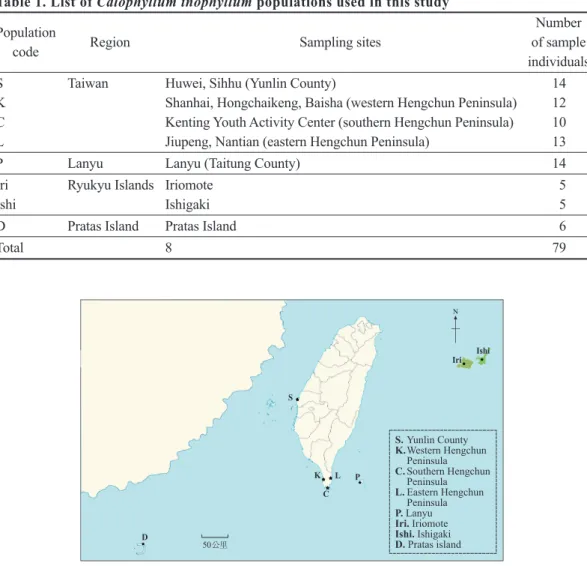
計79個有效樣株,採集已成熟之新生葉片,以 供相關分析。取樣時,採逢機取樣,每單株選 取無病蟲害之葉子5~10片,樣株與樣株間儘量 相隔10 m以上,並避免採集到同源單株及人工 栽植之林木。相關採集資訊如表1及圖1。
二、DNA之萃取與ISSR的擴增反應
以 乾 燥 的 成 熟 新 生 葉 片 為 材 料 , 使 用 Plant Genomic DNA Mini Kit (Viogene, Taiwan) 進 行 萃 取 , 分 離 出 的D N A 加 入 0 . 2 m l 的 T E Buffer (Amresco, USA)保存,並利用分光光 度計(Hitachi U-2001, Japan)測定DNA濃度。
Polymerase chain reaction (PCR)反應參考 Zietkiewicz et al. (1994)的方法進行,ISSR 引 子 取 自Operson Tech. (USA)的100個引子 (UBC 801 -- 900),經最佳化測試(optimization) 篩 選 共14個具多型性及再現性高的引子來進 行 分 析( 表 2 ) 。 反 應 溶 液 總 體 積 為 2 5 μ l , 配 方為2.5 mM dNTPs 1 μl (Protech, Taiwan),
1.0 mM MgCl2 0.025 μl (Protech),10X buffer 2.5 μl (Protech),DNA template 2 μl,primer (10 ng/uL) 2 μl,1 unit Taq polymerase 0.25 μl (Protech),ddH2O 17.225 μl,所有反應均 在溫度循環儀(thermal cycling machine, ABI 9700, city, ST, USA)進行,反應起始條件 (initial denaturing step)為94℃,6分鐘,再以 94℃,60秒;50.6~60.5℃,50秒,黏合溫度 (annealing temperature)依引子不同而定(表2);
72℃,2分15秒循環35次,最後以72℃,10
分 鐘 結 束 反 應 , 反 應 結 束 降 溫 至4℃保存反應 產物。
三、電泳、染色與照相
配製2% Agarose (SIA, city, ST, USA),
將PCR產物及分子量標記(100 bp Ladder DNA Marker II, YEA, city Taiwan)注入後,放入0.5x Tris base, boricacid and EDTA (TBE)緩衝液之 水平電泳槽(KODAK HR2025, city, ST, USA),
以電壓150 V進行電泳分離,約2~2.5小時。完
成電泳的膠片以溴化乙錠(ethidium bromide) (Sigma Aldrich Fluka, city, ST, USA)染色6分 鐘 , 退 染8分鐘後,置於260 nm波長的UV燈 上顯影觀察條帶,並以紫外燈照相系統(Vilber Lourmat CN-100, city Franch)拍照記錄。
四、族群遺傳參數分析
(一) 記 錄 強 度 較 強 且 清 晰 的 條 帶 , 有 條 帶 出 現 者 記 錄 為1 , 無 條 帶 者 為 0 。 利 用 NTSYS-pc ver. 2.01套裝軟體(Rohlf Fig. 1. Map showing locations of the 8 sampled Calophyllum inophyllum populations among the nearby regions Lanyu, Pratas Island, Okinawa, and Taiwan. Population codes are the same as in Table 1.
Table 1. List of Calophyllum inophyllum populations used in this study
Population Number
code Region Sampling sites of sample
individuals
S Taiwan Huwei, Sihhu (Yunlin County) 14
K Shanhai, Hongchaikeng, Baisha (western Hengchun Peninsula) 12 C Kenting Youth Activity Center (southern Hengchun Peninsula) 10 L Jiupeng, Nantian (eastern Hengchun Peninsula) 13
P Lanyu Lanyu (Taitung County) 14
Iri Ryukyu Islands Iriomote 5
Ishi Ishigaki 5
D Pratas Island Pratas Island 6
Total 8 79
1993),建立相似度矩陣(Ssm),另以 Dice (1945)公式計算兩兩樣本間之相似 度(SAB)。SAB = 2NAB / (2NAB + NA + NB)。
SAB = 兩兩樣本間之相似度。NA = 僅在樣 本A出現的條帶數。NB = 僅在樣本B出現 的條帶數。NAB = 在樣本A與在樣本B皆 出現的條帶數。
(二) 將各族群之條帶記錄以POPGENE vers.
1.31套裝軟體(Yeh et al. 1999)計算族群 間的遺傳變異資料,包含Nei’s基因歧異 度(Nei’s gene diversity; H) (Nei 1973),
Shannon’s多型性訊息指數(I) (Shannon and Weaver 1949)。另假設各族群近 親 交 配 指 數(Fis)為0,則可計算族群間 遺傳分化係數Gst (coefficient of gene differentiation),再利用Gst估算基因流 (gene flow)值(Nm),Nm = 0.25×(1 - Gst) / Gst (Slatkin and Barton 1989)。
(三) 依Excoffer et al. (1992)所發表兩兩族群 的距離公式求得距離矩陣D。D = N (1 - Ssm) = N (1 - M/N); M = 兩兩樣本相 同條帶數;N = 多型性條帶總數。將距
離矩陣以analysis of molecular variance (AMOVA) vers. l.55程式進行AMOVA分 析,求得地區間、地區內、族群間、族 群內個體間之變方成分值及各變方成分 所佔總變方成份的百分比,並用9999次 之隨機重排列檢定,測驗各變方成分之 顯著性。
(四) 依前述所建立之相似度矩陣,利用配對 不加權算數平均值(Unweighted Pair-Group Method with Arithmetic Averages, UPGMA) 的方法進行歸群分析(cluster analysis)並 建立個體歸群圖(dendogram) (Sokal and Michener 1958);同時運用主座標分析 (principal coordinate analysis, PCoA)來 繪 製 樣 本 分 布 圖 。 此 外 , 透 過M a n t e l test (Mantel 1967)來檢測遺傳距離矩陣 ( Φst)與相對地理矩陣間的相關性(r)。
結 果
一、瓊崖海棠多型性ISSR引子之篩選 共 測 試O p e r s o n Te c h . 1 0 0 個 隨 機 引 子 Table 2. Properties of 14 ISSR primers used in this study
Annealing
GC Number of Number of Percentage of Primer Sequence 5’→3’ temperature
(%) monomorphic polymorphic polymorphic
(℃) fragments fragments fragments
UBC807 5’–AGA GAG AGA GAG AGA GT–3’ 50.4 47.1 16 8 50.0 UBC810 5’–GAG AGA GAG AGA GAG AT–3’ 50.4 47.1 18 12 66.7 UBC818 5’–CAC ACA CAC ACA CAC AG–3’ 52.8 52.9 12 10 88.8 UBC823 5’–TCT CTC TCT CTC TCT CC–3’ 52.8 52.9 16 15 93.8 UBC825 5’–ACA CAC ACA CAC ACA CT–3’ 50.4 41.7 11 4 36.4 UBC827 5’–ACA CAC ACA CAC ACA CG–3’ 52.8 52.9 16 12 75.0 UBC842 5’–GAG AGA GAG AGA GAGA(CT)G–3’ 54.8 52.8 16 15 93.8 UBC844 5’–CTC TCT CTC TCT CTC T(AG)G–3’ 54.8 52.8 14 13 92.9 UBC848 5’–CAC ACA CAC ACA CAC A(AG)G–3’ 54.8 52.8 15 14 93.3 UBC865 5’–ATG ATG ATG ATG ATG ATG–3’ 74.2 100 14 14 100 UBC886 5’–(AGC)(AGT)(AGC)CTC TCT CTC TCT CT–3’ 51.9 50.9 14 6 42.9 UBC887 5’–(AGT)(AGC)(AGT)TCT CTC TCT CTC TC–3’ 51.1 48.9 12 12 100 UBC889 5’–(AGT)(GCT)(AGT)ACA CAC ACA CAC AC–3’ 51.1 48.9 12 8 66.7 UBC890 5’–(AGC)(ACT)(AGC)GTG TGT GTG TGT GT–3’ 51.9 50.9 17 16 94.1
Total 203 159
Average 78.33
(UBC 801-900),並從中選取14個具多型性及 再現性高的引子來進行分析,共獲得159個多 型 性 條 帶(表2),個別引子多型性條帶比例介 於36.4~100%,平均78.33%,各引子中以UBC 890獲得的多型性條帶最多(16個),UBC 825 最 少(4個);多型性條帶比例最高是UBC 865 (100%),最低為UBC 825 (36.4%)。
二、 瓊崖海棠族群間遺傳歧異度、遺傳分化 係數、基因流之分析
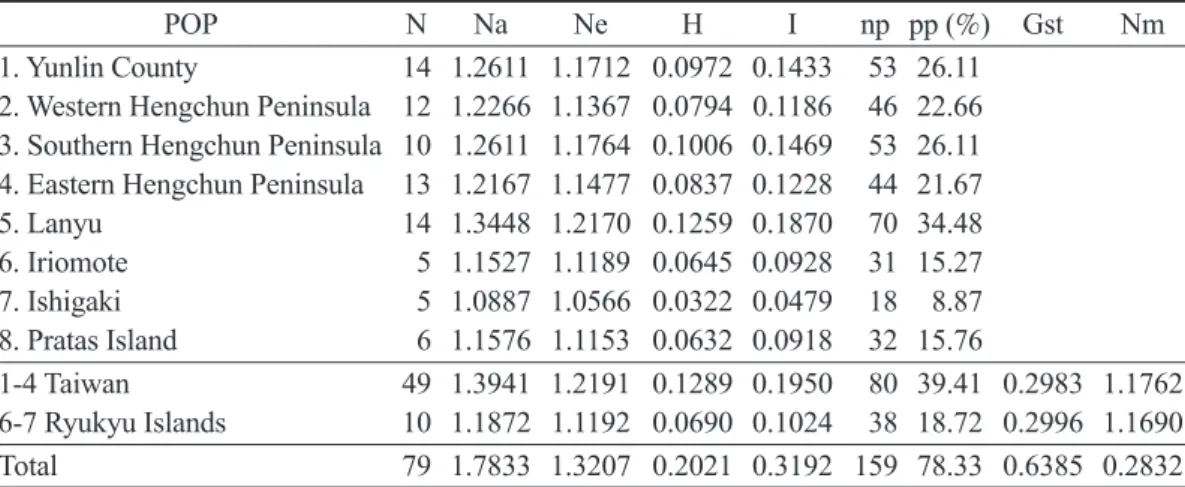
依據ISSR分析所得之條帶整理為0、1的數 值矩陣,並假設各族群自交指數(Fis)為0,以 POPGENE分析瓊崖海棠族群的遺傳變異與多 型性訊息指數(表3),結果顯示瓊崖海棠總體基 因歧異度(H)為0.2021;Shannon’s多型性訊息 指數(I)為0.3192;族群間遺傳分化係數(Gst)為 0.6385,基因流(Nm)為0.2832。各族群間之基 因歧異度值(H)從蘭嶼的族群(0.1259)至石垣島 的族群(0.0322),Shannon’s多型性指數(I)則介 於0.1870 (蘭嶼族群)至0.0479 (石垣島族群)。
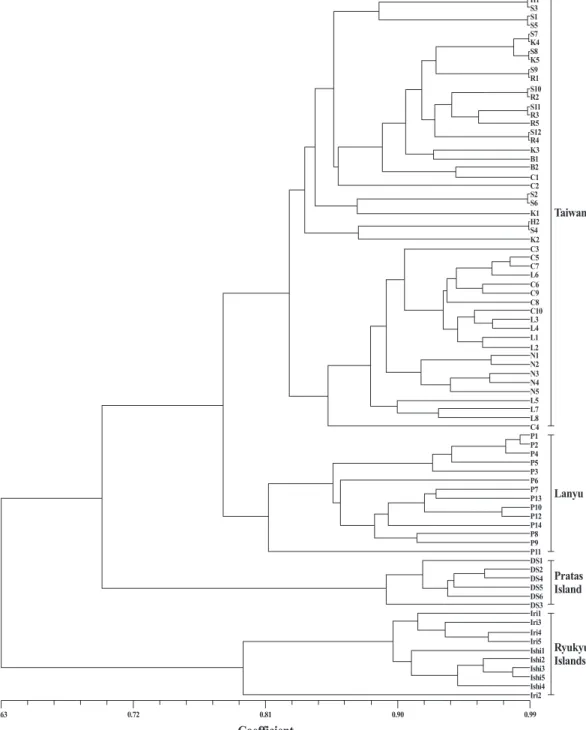
三、瓊崖海棠族群親緣關係與族群遺傳結構 依據ISSR分析所得之數值矩陣,以NTSYS- PC軟體進行個體間相似度矩陣計算,並運用
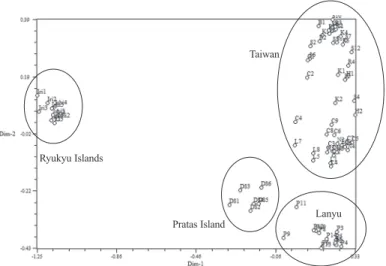
UPGMA進行歸群分析建構個體間之親緣關係 圖(圖2),經比對遺傳距離矩陣與樹狀圖關係矩 陣,得到協表相關係數(cophenetic correlation coeficient)為0.9503,其結果可於相似度0.79處 區 分 為4群,由台灣、蘭嶼、琉球群島及東沙 群島之族群各自集成群,而在台灣島內不同族 群間的個體則呈現不同程度的交錯現象,並形 成樹形圖上的小分枝。此結果再經主座標分析 (PCoA)後亦呈現出相似情況(圖3)。至於瓊崖海 棠族群遺傳距離(Φst)與地理距離相關性(r)之 關係,經Mantel test檢定結果並無顯著相關(r = 0.78,p = 0.99)。參考歸群結果將瓊崖海棠8個 族群依台灣、蘭嶼、琉球群島及東沙群島等4個 區域進行AMOVA分析,結果指出瓊崖海棠族 群於地理區域變方成分佔總變方成分的47.87%
(p < 0.001),地理區域內族群間之變方成分佔 14.53% (p < 0.001),族群內個體間則佔37.60%
(p < 0.001),顯示地理區域(島嶼)間的分化程度 較高(表4)。
討 論
一、族群的遺傳多樣性
遺 傳 多 樣 性 是 生 物 適 應 多 變 環 境 的 基
Table 3. POPGENE analysis of genes, genetic differentation, and gene flow of Calophyllum inophyllum
POP N Na Ne H I np pp (%) Gst Nm
1. Yunlin County 14 1.2611 1.1712 0.0972 0.1433 53 26.11 2. Western Hengchun Peninsula 12 1.2266 1.1367 0.0794 0.1186 46 22.66 3. Southern Hengchun Peninsula 10 1.2611 1.1764 0.1006 0.1469 53 26.11 4. Eastern Hengchun Peninsula 13 1.2167 1.1477 0.0837 0.1228 44 21.67
5. Lanyu 14 1.3448 1.2170 0.1259 0.1870 70 34.48
6. Iriomote 5 1.1527 1.1189 0.0645 0.0928 31 15.27 7. Ishigaki 5 1.0887 1.0566 0.0322 0.0479 18 8.87 8. Pratas Island 6 1.1576 1.1153 0.0632 0.0918 32 15.76
1-4 Taiwan 49 1.3941 1.2191 0.1289 0.1950 80 39.41 0.2983 1.1762 6-7 Ryukyu Islands 10 1.1872 1.1192 0.0690 0.1024 38 18.72 0.2996 1.1690
Total 79 1.7833 1.3207 0.2021 0.3192 159 78.33 0.6385 0.2832
N, number of samples; Na, observed number of alleles; Ne, effective number of alleles; H, Nei’s (1973) gene diversity; I, Shannon’s information index; np, the number of polymorphic loci; pp, the percent- age of polymorphic loci; Gst, proportion of the total diversity among populations; Nm, gene flow.
礎 , 因 此 在 探 討 物 種 保 育 與 其 合 理 利 用 時 , 實 有 賴 於 能 了 解 其 遺 傳 多 樣 性 的 分 布 、 分 化 (differentiation)及其影響因子(Solbrig 1991)。
遺傳多樣性越高,適應環境的能力就越強,越 容易擴展其分佈範圍和開拓新的環境。一般而 言,廣泛分布物種趨向於擁有較高之遺傳多樣
Fig. 2. UPGMA dendrogram based on 159 polymorphic ISSR bands of 79 individuals of Calophyllum inophyllum.
性(Hamrick et al. 1991)。此外,物種在有生存 壓力的環境下也會導致遺傳多樣性的增高(Nevo 1998)。本樹種透過ISSR分析結果顯示,8個瓊 崖海棠族群所獲得的多型性條帶平均百分比為 78.33%,此數值相較於其他濱海植物運用ISSR 分析的結果,低於銀葉樹之83.6% (Deng et al.
2009),土沈香(Excoecaria agallocha Hayata.) 的87.21% (Zhang et al. 2005),但高於苦楝 (Melia azedarach Linn.)的66.67% (Ho and Ko 2010)及棋盤腳樹的66.67% (Deng et al. 2011)。
相較結果顯示瓊崖海棠各自族群間的遺傳多樣 性並不高,推測可能歸因於本研究範圍內之族 群係位於本物種天然分布上的邊緣區域,由於 環 境 逆 境 較 高 , 加 上 濱 海 地 區 常 年 遭 人 為 破 壞,遂導致該樹種於本研究調查範圍內之有效 族群數量過低所致。
此 外 , 瓊 崖 海 棠 之 基 因 歧 異 度( H ) 為 0 . 2 0 2 1 ; S h a n n o n ’ s 多 型 性 訊 息 指 數 ( I ) 為 0.3192,此數值相較於其他濱海植物運用ISSR 分析的結果,其基因歧異度值(H)顯然較低,如 銀葉樹為0.2888 (Deng et al. 2009)、土沈香為 0.2150 (Zhang et al. 2005)、苦楝為0.3930 (Ho and Ko 2010)、相思樹(Acacia confusa Merr.)為 0.4512 (Liu et al. 2007)及全緣冬青(Ilex integra Thunb.)為0.2220 (Leng et al. 2005)。另也低於 Deng et al. (2009)所整理的裸子植物平均值H = 0.2362,I = 0.3599及異交木本被子植物的平均 值H = 0.2764,I = 0.3883。綜言之,瓊崖海棠 的基因歧異度相對其他濱海植物而言並不高,
顯示各族群應歷經嚴重的瓶頸效應(bottleneck effect),推測同樣與本研究範圍內之族群係位 於本物種天然分布上的邊緣區域,加上濱海地 Fig. 3. Results of 2-dimensional principal coordinate analysis based on ISSR of 79
Calophyllum inophyllum L. individuals studied.
Table 4. Analysis of molecular variance for the 8 populations of Calophyllum inophyllum Source of variation df Ssd Msd Variance components variance% Total p-value
Nest analysis
Variance among regions 3 802.72 267.57 14.68 47.87 < 0.001 Variance among populations within regions 4 231.29 57.82 4.45 14.53 < 0.001 Variance within populations 71 818.40 11.53 11.52 37.60 < 0.001 df, degrees of freedom; Ssd, sum of squares; Msd, mean squares; p-value, probability.
區 常 年 遭 人 為 破 壞 嚴 重 , 致 族 群 數 量 銳 減 , 致 使 族 群 歧 異 度 呈 現 偏 低 的 情 況 。 此 結 果 與 Aranaud-Haond et al. (2006)運用SSR來比較海 茄苳位於分布邊緣區域和核心區域族群的遺傳 多樣性之結果相似,同樣以位居分布邊際區域 的族群之遺傳多樣性較低,其原因推測可能歸 因於分布邊緣的族群,由於在分布邊緣上其有 效族群大小組成過低且缺乏有效傳粉昆蟲,加 上太高的環境逆境所致。
二、族群的遺傳分化
族 群 遺 傳 結 構 常 受 到 許 多 因 素 的 影 響 , 如 繁 殖 系 統 、 分 布 範 圍 、 基 因 流 、 以 及 花 粉 與種子的傳播機制等(Schaal et al. 1998, Ge et al. 2003)。族群的遺傳分化則能反應出物種的 長期進化歷史、基因突變、遺傳漂變(genetic drift)、交配系統及基因流等不同過程之相互 作用(Schaal et al. 1998)。瓊崖海棠總體之遺 傳分化係數(Gst)為0.6385,此數值相較於其他 濱 海 植 物 運 用ISSR分析結果,高於土沈香族 群的Gst值0.2210 (Zhang et al. 2005)、苦楝的 Gst值0.4153 (Ho and Ko 2010)及海南草海桐 (Scaevola hainanensis Hance.)族群的Gst值 0.1720 (Ho et al. 2005)。此外,亦較Deng et al.
(2009)整理所得濱海植物的Gst平均值0.2606為 高,顯見族群間已存有一定程度之遺傳變異。
族群的遺傳變異主要係受到三種自然力之 左右,分別為基因流、天擇(natural selection) 與遺傳漂變,其中基因流(Nm)是重要的演化媒 介 , 它 決 定 一 個 族 群 的 地 方 性(endemic)遺傳 分化程度(Huang 1991)。遺傳變異是進化的動 力,故進化過程勢必伴隨著大小不一的遺傳變 異,是種化的重要關鍵(Grant 1981)。瓊崖海棠 為異交植物,其果實可藉海水漂流散佈繁殖,
並藉由適應新環境而長成植株,拓展其族群。
據Wright (1969)的觀點,若Nm > 1,基因流 就足以克服遺傳漂變的作用,防止族群分化的 發生,如Nm < 1則反之;若依據Ellstrand and Elam (1993)的意見,Nm只要大於0.5則就被認 為足以克服隨機遺傳漂變所造成的分化。由於 植物族群間的基因流傳是借助於花粉、種子、
孢子或營養體的遷移來進行,尤以花粉和種子 的擴散與傳播為主(Harmrick et al. 1991),當異 交率越高的植物其基因流傳強度將越大,如為 自交植物則其基因交流為零。本研究瓊崖海棠 族群之基因流指數為0.2832 (Nm < 0.5),無論 依據Wright (1969)或Ellstrand and Elam (1993) 較嚴格的意見,均說明本樹種各族群間的基因 流並不通暢。若依此數值與其他濱海植物運用 ISSR分析結果相比,不僅低於土沈香的Nm值 0.8840 (Zhang et al. 2005)、苦楝的Nm值0.7040 (Ho and Ko 2010)及海南草海桐的Nm值2.4000 (Ho et al. 2005),同時與Deng et al. (2009)整 理所得濱海植物的平均值(Nm = 1.4459)與異交 木本植物的平均值(Nm = 1.2905)相較結果亦較 低。雖然瓊崖海棠果徑約2~3 cm,外有硬脆質 之果皮,不但能阻隔海水滲入,並能漂浮於海 水上進行傳佈(Liu et al. 1994);但限於瓊崖海 棠難以忍受低溫的逆境,當最冷月份平均溫度<
12℃時即無法生長(Friday and Okano 2006),故 其傳播距離與效率遂受限制,近距離族群間的 傳播可能性較大。Duke (1995)亦提出雖然很多 紅樹植物均具可藉海流傳播的種子或繁殖體,
但其長距離傳播的能力值得懷疑,尤其是廣泛 分布的紅樹植物。因此,類此海漂植物如要跨 越島嶼間的遠距隔離仍有困難,並非所有種子 均可順利著陸且適應環境而長成植株。
另本樹種的授粉媒介主要係透過昆蟲來完 成,如蜜蜂及甲蟲類等(Wahyuni et al. 2012),
由 於 透 過 弱 飛 行 能 力 的 動 物 授 粉 和 靠 種 子 遷 移的植物其族群間的遺傳分化會較顯著(Avise 1994),主要限於島嶼間飛行能力,島嶼間之 傳粉過程受阻礙,導致族群間的基因流受阻,
加上海洋的阻礙與人為的破壞,因此在不同島 嶼 間 呈 現 出 分 化 之 現 象 。 此 結 果 與Kwon and Morden (2002)認為島嶼間地理隔離會限制花粉 及種子傳播的有限範圍相吻合,換言之,瓊崖 海棠實際傳播距離和效率均受限制。
另由個體間親緣關係圖(圖2)顯示,於相似 度0.79處劃分時台灣、蘭嶼、琉球群島及東沙 群 島 等 地 區 的 族 群 各 自 聚 為 一 群 , 以 上4群的 親緣關係以台灣與蘭嶼的族群較接近先聚為一
群,再與東沙群島歸為一群;而以琉球群島的 遺傳距離最遠。但就台灣島內不同族群間其個 體的群聚關係而言,並無單一族群的所有個體 可共同歸於同一支系下,不同族群間的個體係 以不等程度的交錯情況形成小分支;另依主座 標分析(PCoA)結果(圖3),同樣也呈現出一致 結果,顯示台灣島內不同族群間無明顯差異;
惟在不同島嶼間的族群有較明顯的差異。據此 再將瓊崖海棠族群依島嶼區分為台灣、蘭嶼、
琉球群島及東沙群島等4個地區來進行AMOVA 分析,結果各變方成分均達顯著水準,亦佐證 瓊崖海棠在島嶼間及各族群間已有分化趨勢,
其中族群地理區域間變方成分(佔47.87%)較族 群內個體間(37.60%)為大,顯示族群在地理島 嶼間的分化程度更加顯著。此應與小族群較易 受 到 遺 傳 漂 變 導 致 基 因 的 喪 失 及 基 因 流 的 受 限,致使族群間的分化程度增加有關(Ellstrand and Elam 1993),此外瓊崖海棠分布範圍包括 東南亞、印度、琉球、澳洲與太平洋諸島等地 區 , 相 對 於 本 研 究 範 圍 實 係 屬 本 種 之 邊 際 分 布 區 域 , 故 亦 受 分 布 極 限 、 邊 緣 效 應 之 影 響 (Aranaud-Haond et al. 2006)。
另 以Mantel test檢定族群間的遺傳距離 (Φst)與地理距離(r),結果兩者之間並無顯著相 關(r = 0.78, p = 0.99),據Fisher et al. (2000)認 為當遺傳距離與地理距離之間缺乏顯著相關性 時,意味著遺傳漂變在族群間的分化上扮演不 可忽視之力量,當一個族群與其他族群相互隔 離時,遺傳漂變成為影響族群遺傳結構和族群 間遺傳變異的重要因素。由於海岸林的開發破 壞,生育地縮減致使現存族群太少,如隔離時 間長,可能會因近交衰退,加上島嶼地理位置 封閉基因無法交流而遺傳漂變造成某些基因的 喪失而降低族群的遺傳變異。至於琉球群島族 群與其他族群差異較大之原因,推測係受到島 嶼隔離與海流方向之影響,由於黑潮(Kuroshio) 的海流方向主要沿菲律賓東岸向北流,經呂宋 海峽由台灣東岸外海經過(Wessells and Hopson 1988),基於海流之不可逆遂限制琉球群島族 群與其他族群的交流。此外,加上黑潮於日本 的東或東北方與由極地南流的親潮(Oyashio)相
會,因受寒冷洋流的限制,琉球群島的族群遂 成為本物種天然分布的最北限亦有關聯(Friday and Okano 2006, Hanaoka et al. 2014);至於 東沙島的族群與台灣族群較接近之原因,主要 是黑潮流經呂宋海峽時部份海流轉向北南海,
並於台灣南端流出再重新注入黑潮,但仍有部 份 海 流 會 繼 續 流 入 南 海 或 經 台 灣 海 峽 流 入 東 海,因此黑潮終年可經呂宋海峽流入南海,再 藉由大陸的沿岸流與台灣西岸的族群進行交流 (Wessells and Hopson 1988)。
結 論
就物種保育而言,瞭解族群各項特徵為最 基礎且必要之工作,而瞭解族群的遺傳結構則 有利於了解基因庫大小、遺傳變異及分化程度 等,對後續保育策略的擬訂,可提供一個有效 的參考依據。本研究分析瓊崖海棠族群遺傳訊 息顯示,本樹種的基因歧異度(H)相較於其他濱 海植物而言並不高,此應與瓊崖海棠現存各族 群之生育地較小且遭嚴重破壞,導致遺傳歧異 度偏低所致。從族群遺傳變異與遺傳結構分析 (表3與4)亦顯示其族群間已有遺傳分化產生,
族群間的基因流並不通暢,不同島嶼間呈現明 顯分化現象。族群散布之情形,依Mantel test的 結果,其族群為隨機之散佈,故遺傳漂變在族 群分化上扮演重要角色,不同島嶼間所呈現分 化現象,推測仍係受到島嶼隔離、分布極限及 海流方向等因素之影響。瓊崖海棠雖為廣泛分 布的物種,但由於人為的破壞、生育地惡化甚 至喪失等因素,造成在海岸地區形成片斷化分 布,漸成狹隘分布的單一小種群;由於種群內 個體數量的減少,遂導致族群瓶頸效應顯著,
進而造成基因流傳的下降和遺傳漂變的產生。
因此,有關瓊崖海棠族群的保育策略,考量島 嶼與族群間已存有明顯遺傳分化,因此單獨保 護其中任一個族群對保護整個物種的遺傳變異 顯有不足,除採取就地(in situ)保育之措施來保 存所有的族群外,並應實施遷地(ex situ)保育 措施來加以保存,其種源的蒐集應涵蓋現有之 族群。
致 謝
本研究承行政院農業委員會林業試驗所科 技計畫(104農科-13.1.2-森-G2)經費補助,特此 致謝。
引 用 文 獻
Aranaud-Haond S, Teixeira S, Massa SI, Billot C, Saenger P, Coupland G, et al. 2006.
Genetic structure at range edge: low diversity and high inbreeding in Southeast Asian man- grove (Avicennia marina) populations. Mol Ecol 15(12):3515-25.
Avise JC. 1994. Molecular markers, natural history, and evolution. New York: Chapman Hall. 511 p.
Deng SL, Lu FY, Hsui YR, Lin YD, Wu HC.
2009. Population genetic variation of Heritiera littoralis among Taiwan, Iriomote-Island and Hong Kong. Q J Chin For 42(4):503-18. [in Chinese with English summary].
Deng SL, Hsui YR, Lu FY, Chang KC, Chang YH, Wu HC. 2011. Population genetic variation of Barringtonia asiatica between Hengchun Peninsula and Lanyu. J Agric Assoc Taiwan 12(4):364-81. [in Chinese with English summary].
Deng SL, Yu HM, Hsui YR, Hwong JL.
2014. Introduction of genetic variation of Jat- ropha curcus germplasm resources to Taiwan.
J Agric Assoc Taiwan 15(1):76-93. [in Chinese with English summary].
Dice LR. 1945. Measures of the amount of ecologic association between species. Ecology 26:297-302.
Excoffer L, Smouse PE, Quattro LM. 1992.
Analysis of molecular variance inferred from metric distances among DNA haplotypes: ap- plication to human mitochondrial DNA restric- tion data. Genetics 131:479-91.
Ellstrand NC, Elam DR. 1993. Population
genetics consequence of small population size:
implications for plant conservation. Annu Rev Ecol Syst 24:217-42.
Fisher M, Husi R, Prati D, Peintinger M, Kleunen M, Schmid B. 2000. RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). Am J Bot 87(8):1128-37.
Friday JB, Okano D. 2006. Species profiles for Pacific island agroforestry: Calophyllum inophyllum (Kamani). Hawaii, USA: Perma- nent Agriculture Resources.
Ge XJ, Yan Y, Zhao NX, Chen HS, Qi WQ.
2003. Genetic variation in the endangered In- ner Mongolia endemic shrub Tetraena mon- golica Maxim. (Zygophyllaceae). Biological Conservation 111:427-34.
Grant V. 1981. Plant speciation, 2nd edition.
New York: Columbia Univ. Press. p 149-80.
Hamrick JL, Godt MJW, Murawski DA, Loveless MD. 1991. Correlations between species traits and allozyme diversity: implica- tions for conservation biology. In: Falk DA, Holsinger KE, editors. Genetics and conserva- tion of rare plants. New York: Oxford Univ.
Press. p 75-86.
Hanaoka S, Chien CT, Chen SY, Watanabe A, Setsuko S, Kato K. 2014. Genetic struc- tures of Calophyllum inophyllum L., a tree em- ploying sea-drift seed dispersal in the northern extreme of its distribution. Ann For Sci 71(5):
575-84.
Ho KY, Deng SL, Chang YH, Tsai CS, Kao MF, Hsiao JY. 2005. Genetic variation of endangered Scaevola hainanensis (Goodenia- ceae) in the Jiangiun Stream mouth. Taiwan J For Sci 20(3):193-202.
Ho KY, Ko NY. 2010. Genetic diversity among Melia azedarach L. populations at different climatic regions in Taiwan. Crop Environ Bio- inform 7:195-211. [in Chinese with English summary].
Hsieh KH, Lin SJ, Wu YP. 2014. Genetic di- versity of Zingiberaceae crop. J Taiwan Agric Res 63(3):235-48. [in Chinese with English summary].
Hsu KS, Lin TT, Lu SY, Chiou WL. 1985.
Investigation and report of the rare plants of the Kenting National Park. Taipei, Taiwan:
Taiwan Forestry Research Institute. 101 p. [in Chinese].
Huang S. 1991. Gene flow and population differentiation. Chin Biosci 34(2):19-32. [in Chinese with English summary].
Itoigawa M, Ito M, Tan HT, Kuchide M, Tokuda H, Nishino H, Furukawa H. 2001.
Cancer chemopreventive agents, 4-phenylcou- marins from Calophyllum inophyllum. Cancer Lett 169(1):15-19.
Kwon JA, Morden CW. 2002. Population ge- netic structure of two rare tree species (Colu- brina oppositifolia and Alphitonia ponderosa, Rhamnaceae) from Hawaiian dry and mesic forests using random amplified polymorphic DNA markers. Mol Ecol 11(6):991-1001.
Leng X, Wang Z, An S, Feng J, Liu Y, Wang G. 2005. ISSR analysis of genetic diversity of Ilex integra, an insular endemic plant. Chinese Biodiversity 13(6):546-54. [in Chinese with English summary].
Liu YC, Lu FY, Ou CH. 1994. Trees of Tai- wan. City, Taiwan; College of Agriculture, Na- tional Chung-Hsing University. Monographic Publication no. 7. 925 p. [in Chinese].
Liu YY, Liao YK, Hu TK. 2007. Study of genetic variation among the populations of Acacia confusa at west regions in Taiwan. J Agric For (NCYU) 4(2):32-46. [in Chinese with English summary].
Mantel NA. 1967. The detection of disease clustering and a generalized regression ap- proach. Cancer Res 27(2):209-20.
Nei M. 1973. Analysis of gene diversity in subdivided populations. Proc Natl Acad Sci
USA 70:3321-3.
Nevo E. 1998. Molecular evolution and ecolog- ical stress at global, regional and local scales:
the Israeli perspective. J Exp Zool 282(1-2):
95-119.
Ong HC, Mahlia TIM, Masjuki HH, Nor- hasyima RS. 2011. Comparison of palmoil, Jatropha curcas and Calophyllum inophyllum for biodiesel: a review. Renewable and Sus- tainable Energy Reviews 15(8):3501-15.
Patil AD, Freyer AJ, Eggleston DS, Halti- wanger RC, Bean MF, Taylor PB, et al.
1993. Inophyllums novel inhibitor of HIV- reverse transcriptase isolated from Malaysian tree, Calophyllum inophyllum Linn. J Med Chem 36(26):4131-8.
Rohlf FJ. 1993. NTSYS-pc Numerical Taxon- omy and Multivariate Analysis System, Vers.
1.55. New York: Exeter Software. 7 p.
Shannon CE, Weaver W. 1949. The math- ematical theory of communication. Urbana:
Univ. of Illinois Press. 54 p.
Schaal BA, Haywdrth DA, Olsen KM, Rauscher JT, Smith WA. 1998. Phylogeo- graphic studies in plants: problems and pros- pects. Mol Ecol 7:465-74.
Slatkin M, Barton NH. 1989. A comparison of three indirect methods for estimating aver- age levels of gene flow. Evolution 43:1349-68.
Solbrig OT. 1991. From genes to ecosystems:
a research agenda for biodiversity. Cambridge IUBS. 124 p.
Sokal RR, Michener CD. 1958. A statistical method for evaluation systematic relationships.
Univ. Kans Sci Bull 38:1409-38.
Wahyuni R, Handoko C, Agustarini R.
2012. Preliminary study on the flowering and fruiting behaviors of Nyamplung (Calophyllum inophyllum Linn.). J For Res 9(1):1-10.
Wessells NK, Hopson JL. 1988. Biology.
New York: Random House. 1251 p.
Wright S. 1969. Evolution and the genetics
of populations, Vol. 2. The theory of gene fre- quencies. Chicago: Univ. of Chicago Press.
Wei X, Cao HL, Jiang YS, YE WH, Ge XJ, Li F. 2008. Population genetic structure of Ca- mellia nitidissima (Theaceae) and conservation implications. Bot Stud 49:147-53.
Yeh FC, Yang RC, Boyle T. 1999. POPGENE Vers. 1.31: Microsoft Windows-based Free- ware for Population Genetic Analysis. Quick user’s guide. City Canada; Univ. of Alberta.
Zhang ZH, Tang T, Zhou RC, Wang YG, Jian SG, Zhong CR, Shi SH. 2005. Effects of divergent habitat on genetic structure of popu- lation of Excoecaria agallocha, a mangrove associate. Acta Gene Sin32(12):1286-92. [in Chinese with English summary].
Zietkiewicz E, Rafalski A, Labuda D. 1994.
Genome fingerprinting by simple sequence repeat (SSR) anchored polymerase chain reac- tion amplification. Genomics 20(2):176-83.