中 國 醫 藥 大 學 基礎醫學研究所
碩士學位論文
Src 於 CpG 誘發巨噬細胞移行之相關探 討
The implication of Src in CpG-induced macrophage migration
指導教授:馬 明 琪
(Ming-Chei Maa,PH.D.)研究生:李 郁 芸
(Yu-Yun Li)中 華 民 國 九 十 七 年 七 月
目錄
中文摘要(Abstract in Chinese)... 2
英文摘要 (Abstract in English)... 3
第一章 序論(Introduction)... 4
第一節 免疫系統(immune system)及巨噬細胞(macrophage)... 4
第二節 Toll-like receptor 9(TLR9)及 CpG... 7
第三節 Src kinase...11
第四節 iNOS(inducible nitric oxide synthase) ... 14
第二章 實驗材料及方法(Materials and Methods)... 16
第一節 實驗材料 ... 16
第二節 實驗方法 ... 24
第三章 結果(Results)... 37
第四章 討論(Discussion)... 44
參考文獻(Reference) ... 50
圖表... 57
中文摘要(Abstract in Chinese)
Toll-like receptors 是近年來被發現在辨識微生物並活化免疫及發炎反 應上重要的感應器。未甲基化(unmethylated)胞嘧啶(cytosine)-鳥糞鹼
(guanine)雙核甘酸序列(CpG motifs),有強力免疫刺激的能力,在細 菌中遠大於脊椎動物。而未甲基化 CpG 序列能藉由 Toll-like receptor-9 (TLR9)去啟動先天性免疫並活化一連串訊息傳遞,引發許多生物體的反 應,包含巨噬細胞移行。在我們的研究中,證明了在CpG 刺激下,Raw264.7
巨噬細胞的移行和總體酪氨酸磷酸化明顯增加。但合併處理 PP2(Src
family kinases [SFKs] 的抑制劑)後,細胞的移行減少,透露出 SFKs 參 與在CpG 誘發的細胞移行中。進一步發現,Raw264.7 細胞在 CpG 刺激下,
只有 Src 表現增加,Lyn、Fgr 和 Hck 蛋白量沒有很大的差異,而取自老 鼠腹腔之巨噬細胞也有相同的現象。在Raw264.7 細胞用 RNA interference 的技術減少Src 表現量,會減弱 CpG 誘發的細胞移行,而回復 Src 表現,
則可恢復細胞移行能力。合併處理TLR9 抑制劑 chloroquine 和 quinacrine 後 ,CpG 誘 發 的 Src 表 現 和 細 胞 移 行 皆 減 弱 , 也 顯 示 此 過 程 為 TLR9-dependent。另外,我們也發現在缺少 iNOS 的巨噬細胞,其 CpG 引
發的Src 表現和細胞移行皆減少。由上述的結果,我們認為 iNOS 可以調
節Src 表現,進而影響 CpG 誘發的細胞移行能力。
關鍵字:CpG 雙核甘酸序列,Src,巨噬細胞移行能力
英文摘要 (Abstract in English)
Toll-like receptors (TLRs) have recently been identified as important sensors of infection that, upon microbial recognition, activate the immune and inflammatory responses. DNA containing cytosine-guanine dinucleotide (CpG) motifs have potent immunostimulatory activities. Unmethylated CpG dinucleotides in particular sequence contexts occur frequently in bacterial but not in vertebrate DNA. Unmethylated CpG motifs initiate innate immune responses via Toll-like receptor-9 (TLR9), which activates a spectrum of signaling molecules and triggers a variety of biological responses including migration in macrophages. In this study, we demonstrated that upon CpG stimulation, the motility and the content of total protein tyrosyl phosphorylation in Raw264.7 macrophages were greatly enhanced. The abrogation of CpG-induced migration by PP2 (an inhibitor for Src family kinases [SFKs]) suggested the involvement of SFKs in this process. Analysis of the expression of SFKs in Raw264.7 cells revealed that only the expression of Src (but not Lyn, Hck and Fgr) was CpG-inducible. And similar result was also detected in rat peritoneal macrophages. Remarkably, silencing of Src expression by RNA interference reduced CpG-evoked cell mobilization in Raw264.7 macrophages and restored Src expression could rescue this defect.
The inhibition of CpG-induced Src expression and migration in cells pretreated with chloroquine and quinacrine implicating these events were TLR9-dependent. Notably, this CpG-mediated mobilization and Src induction was suppressed in macrophages devoid of iNOS. With these findings, we concluded that iNOS-induced Src expression played a significant role in CpG-elicited macrophage migration.
第一章 序論(Introduction)
第一節 免疫系統(immune system)及巨噬細胞(macrophage)
免 疫 系 統 是 生 物 體 能 辨 識 出 自 體 (self ) 或 非 自 體 物 質
(non-self),進而消滅、排除非自體物質之整體生物反應 [1]。對人 體而言,這些非自體物質可以是微生物、食物、藥物、化學物質甚 至是花粉或動物的毛髮。我們的免疫系統藉由逐漸增加特異性的防 禦能力保護人體免於感染。一般而言,免疫反應可分為先天性免疫
(innate immunity)和後天性免疫(adaptive immunity)。先天性免疫 不具專一性,辨識致病原的能力是與生俱來的。動物體內阻隔感染 的有皮膚、黏膜等物理性屏障,唾液、胃液中酵素的化學屏障及能 與致病菌爭搶食物與生存空間位於泌尿生殖道與腸胃道內的共生 菌,而這是生物性屏障。但若是這一些屏障都無法有效的抵擋病原 菌入侵時,生物體即會啟動第二道防線,參與者主要為吞噬細胞,
這 是 先 天 性 免 疫 的 細 胞 性 屏 障 。 而 這 些 吞 噬 細 胞 包 含 單 核 球
(monocyte)、嗜中性球(neutrophil)和巨噬細胞(macrophage)等,
會藉由吞噬作用(phagocytosis)來消滅外來物,而在進行非特異性 吞噬的同時也會釋出細胞激素(cytokines)和化學激素(chemokines)
引發發炎反應。此外,抗原呈現細胞(antigen presenting cells)如樹
狀細胞(dendritic cells)和巨噬細胞(macrophages),將抗原呈現給 淋巴細胞(lymphocytes),活化 T 細胞分泌細胞激素,進而活化 B 細胞產生專一性的抗體,共同對抗病原菌,而這專一性的免疫也稱 後天性免疫。
單核球是由幹細胞分化而來,屬於顆粒性球,當受到刺激時會 分化成巨噬細胞。巨噬細胞在人體不同部分,有不同的命名,在肝 中 稱 Kupffer cells ; 在 肺 中 為 alveolar macrophages ; 在 腎 中 稱 mesangial cells;在腦中稱 microglial cells。巨噬細胞是一群特化的吞 噬細胞 [2],平時為休止狀態,但在外來物入侵時,會藉由 integrin 調節黏附(adhesive)能力而去和外來物結合,而這樣的交互作用可 以調節巨噬細胞的骨架重新排列、細胞移動、特殊細胞功能的活化、
基因的轉錄、細胞增生及存活等 [3]。而巨噬細胞移行(migration)
至外物入侵處,進而啟動其特殊功能對整體免疫而言至為重要。巨 噬細胞不但可將吞入的微生物由溶體(lysosome)分解,還能產生 H2O2、O2-及 NO 等物質,直接毒殺細菌並分泌細胞激素。另外,巨 噬細胞具有抗原呈現作用,可將抗原呈現給 T 淋巴球。活化的 T 細 胞能分泌各種細胞毒殺蛋白,去除大部分的病原;而 B 細胞則能產 生專一性的抗體對抗病原菌。
免疫系統藉由非特異性的先天免疫和具專一性的後天性免疫共
同作用,保護人體免於感染。巨噬細胞在此過程中扮演承先啟後的 角色,其重要性不可言喻。
第二節 Toll-like receptor 9(TLR9)及 CpG
1997年Medzhitov 等人發現人類有類似果蠅的Toll基因,可偵測 入侵的外來物,並引發免疫反應,故將其蛋白產物命名為Toll-like receptors,這個發現對於先天性免疫影響深遠 [4, 5]。迄今發現的 TLRs 已 有 16 種 之 多 [6] 。 由 於 宿 主 細 胞 上 的 pattern recognition receptors ( PRR ) 可 辨 識 pathogen-associated molecular patterns
(PAMPs),故Toll-like receptors家族即是一種PRRs,可專一性辨識 不同的PAMPs。有一些Toll-like receptors位於細胞表面,例如TLR4 位 於 細 胞 膜 上 , 可 辨 識 革 蘭 氏 陰 性 菌 內 毒 素lipopolysaccharides
(LPS);TLR2可辨識革蘭氏陽性菌的peptidoglycan(PGN)。另有一 些Toll-like receptors位於細胞內,如TLR9位於endosome中,可辨識細 菌 和 病 毒 DNA上的CpG motif [7]。一般TLRs的訊息傳遞經由 myeloid differentiation primary response protein 88(MyD88)、IL-1 receptor associated kinase (IRAK)、TNF receptor-associated factor 6
(TRAF6)、transforming growth factor-β -activated protein kinase 1
(TAK1)和IκB kinase(IKK)、IκB、NF-κB [8, 9]以啟動免疫反應。
未甲基化CpG雙核苷(unmethylated CpG dinucleotides)為核心 的特定核苷酸序列(CpG motifs)可被TLR9所辨識。CpG雙核苷酸
出現頻率在細菌裡遠比脊椎動物DNA多許多 [10, 11],而這是脊椎動 物DNA在長期進化過程中所形成區別自身和外源DNA的重要機制。
不同物種演化出不同的TLR9來精確的掌握在不同物種中特定核苷 酸序列(未甲基化CpG雙核苷和其相鄰的部份),而適時的啟動免 疫反應 [12]。鼠類和人類的TLR9在胺基酸序列約有24%的差異 [13]。與CpG相鄰的DNA在鼠類中皆有在5'端的兩個嘌呤(purines)
在3’兩個嘧啶(pyrimidines),例如GACGTT [14],和CpG相鄰的DNA 在5'端的兩個嘌呤為guanine(G)和adenine(A),3'兩個嘧啶皆 為thymine(T)。但人類和某些其他物種則以TCGTTC和/或TCGTA 呈現 [15],所以CpG相鄰的DNA依物種不同而不同。
合成的oligodeoxynucletides(ODNs)包含CpG motif可以模仿細 菌DNA刺激並引發寄主相似的免疫反應。依據cytokines的分泌,這 些ODN可分為D(CpG-A)、K(CpG-B)和CpG-C三大類 [16]。如 GGTGCATCGATGCAGGGGGG為D(CpG-A)類,在結構上只有單 一CpG motif和其相鄰的序列迴文(palindrome)所組成,在3'尾端 由許多G所組成。主要功能為使抗原呈現細胞(antigen presenting cells)成熟、調節interferon-α (IFN-α)且優先刺激plasmacytoid dendritic cells ( pDCs ) 分 泌 IFN-α 。 而 K ( CpG-B ) 類 其 序 列 如 TCCATGGACGTTCCTGAGCGTT 有多個CpG motif並由硫代磷酸
酯(phosphorothioate)組成背脊(backbone)能使pDCs成熟和製造 tumor-necrosis factor(TNF)並促使B細胞增生和製造IgM和IL-6。
C型的CpG ODN像TCGTCGTTCGAACGACGTTGAT和CpG-B 類相同有多個CpG motifs也是由硫代磷酸酯(phosphorothioate)組成 背脊,但在5'端有兩段TCG序列,實際的CpG motif則藏在序列迴文 中,其也會刺激B細胞製造IgM和IL-6並活化pDCs分泌IFN-α [7]。
TLR9表現於B細胞、巨噬細胞和樹狀細胞,在endosome酸性環 境下辨識CpG [17]。但其攝入DNA ODN的能力和劑量、時間及溫度 有關,而非只攝入含有CpG序列的DNA [18]。有些研究藉由標定螢 光(fluorescently labeled)的方式追蹤ODN;發現無論任何ODN皆會 被攝入細胞內,但只有CpG ODN能活化TLR9 [19]。TLR9的抑制劑 chloroquine 和 其 類 似 物 quinacrine 可 抑 制 CpG 和 TLR9 的 交 互 作 用
(interaction)而抑制免疫反應 [20, 21]。另外,TLR9 knockout 的老 鼠更證實了CpG和TLR9的密切關係 [13]。TLR9的訊息傳遞路徑包含 mitogen-activated protein kinases ( MAPKs ) , 像 是 p38, c-Jun NH2-terminal kinase (JNK),extracellular receptor kinase(ERK),
和NF-κB-inducing kinase(NIK)-IKK-IκB pathways [22-24]。最後會 活化transcription factors包含NF-κB、activating protein-1(AP-1)、
CCAAT/enhancer binding protein ( C/EBP ) 、 cAMP-responsive
element-binding protein(CREB)而影響細胞激素和化學激素的表現 [23, 25, 26]。
第三節 Src kinase
1911 年 Peyton Rous 於 Rous sarcoma virus(RSV)發現第一個 oncogene,為 v-src [27]。而後科學家在正常細胞中發現其類似的 c-src 為 proto-oncogene。本論文提及之 Src 為 c-src 的蛋白產物。其
分子量為60 kDa,為 non-receptor protein-tyrosine kinase,是 Src family kinase(SFKs)的其中一員。可接受例如:細胞激素(cytokines)、
生長因子(growth factor)、黏附(adhesion)和抗原接受體(antigen receptor ) 的 刺 激 , 而 產 生 細 胞 生 長 ( cell growth )、 分 化
(differentiation )、 細 胞 形 狀 改 變 ( cell shape alteration )、 移 行
(migration)及生存(survival)等生理反應 [28]。Src 的蛋白質結 構從N-端到 C-端,依序為 SH4 domain、unique domain、SH3 domain、
SH2 domain、SH2-kinase linker、protein-tyrosine kinase domain(SH1 domain)和 C-terminal kinase domain。SH4 的 myristylation 可讓 Src 透過長鏈脂肪酸與細胞內膜結合;unique domain 為整個家族中的成 員各自不同之處;SH3 可與 proline rich 的胺基酸序列結合;SH2 則 是 與 phosphorylated tyrosine 有 很 強 的 鍵 結 ; kinase domain 與 regulatory domain 則是主要調控 Src 活性的功能區塊 [29]。
在調控 Src活性方面,有兩個最重要的磷酸化胺基酸,其分別是
(C-terminal Src kinase)及Chk(Csk的homolog)磷酸化後,pTyr527 結合至Src的SH2 domain,使Src存在於不活化狀態。當pTyr527與 SH2-binding pocket分開後,催化區就會顯露出來 [30],此時位於 activation loop的Tyr416就可以進行自體磷酸化,Src就能穩定存在於 活化態。因此藉由Tyr416與Tyr527兩個胺基酸的磷酸化可調控Src的 活性。此外若Src缺乏C端的tyrosine磷酸化則會使proteasome對Src造 成ubiquitination和degradation,因此在某種狀態下Csk對Src kinase是 正向調控的 [31, 32]。
Src family tyrosine kinases(SFKs)共有 11 個家族成員,分別為 Src、Yes、Fyn、Lyn、Fgr、Hck、Frk、Lck、Srm、 Blk 及 Brk。其 中Src、Fyn 和 Yes 存在於所有的細胞中 [33],Blk、Fgr、Hck、Lck 和Lyn 主要表現在造血細胞(hematopoietic cells)。其中 Lck 在 T 細 胞、Hck 在骨髓單核球細胞、Srm 存在角質細胞(keratinocytes)。
在過去文獻中認為Lyn、Hck、Fgr這三這Src family成員在巨噬細 胞中扮演不可或缺的角色。但在1997年Lowell的實驗室中 [34],使 用Lyn、Hck、Fgr這三種蛋白knockout的老鼠,取其腹腔和骨髓的巨 噬細胞,在LPS刺激下卻發現NO的釋放和IL-1、IL-6、TNF-α等類發 炎前驅物質的分泌,和對照組之巨噬細胞並無差別,所以他們認為 在這三種蛋白不存在下,應該有其他的tyrosin kinase取而代之來活化
巨噬細胞。本實驗室發現,巨噬細胞受LPS刺激下Src表現量增加並 且NO的釋出也增加,所以認為Src在巨噬細胞活化中扮演重要角色 [35]。
在許多人類的癌症像乳癌、大腸癌、胰臟癌、肺癌等,皆可以 發現Src蛋白表現量增加和其活性增加的現象 [36]。這些研究都突顯 出Src在人類癌症的發展(development)和進展(progression)有重 要 的 關 聯 性 。Src 可 以 和 一 些 tyrosine kinase receptors 、 cytokine receptors、steroid receptors、integrin receptors and G-protein coupled receptors作用,藉由這樣的調控讓Src參與了細胞生長、存活、黏附 和移行 [37-39]。在癌症的轉移(metastasis)中,Src藉由和focal adhesion kinase(FAK)的交互作用而參與在其中。一般而言,FAK 磷酸化後會使Src聚集,之後吸引Crk-associated substrate(Cas)。而 Paxillin(Pax)使Crk II聚集到Cas而影響DOCK180(180-kDa protein downstream of Crk)-ELMO(engulfment and cell motility)複合體,
其後刺激Rac-GTP使肌動蛋白聚合(actin polymerization)。另外 Paxillin刺激p21-activated kinase(PAK)促進細胞migration。另外,
透過phosphatidylinositol 3(PtdInsP3)-kinase(PI3K)的路徑也可使 細胞增加黏附和移行 [40]。
第四節 iNOS(inducible nitric oxide synthase)
Nitric oxide synthase(NOS),為NO的合成酵素,分子量約130 kDa,主要的功能為氧化L-arginine變成L-citrulline並釋放出NO。NOS 在體內可分為三大類,分別是eNOS(endothelial NOS;NOS I),iNOS
(inducible NOS;NOS II)以及nNOS(neuronal NOS;NOS III)。
這三種NOS彼此間有著50~60%的相似性,但nNOS與eNOS是屬於持 續性表現的蛋白,存在於神經以及內皮細胞中,且會被鈣離子所活 化。相較於上述兩者,iNOS則是個誘發型的蛋白,表現於內皮細胞、
肝細胞、平滑肌和巨噬細胞,其產生NO的過程中是不需要鈣離子的 [41]。iNOS的結構包含氧化(oxygenase)及還原(reductase)的功 能區塊(domain),iNOS必須在二聚體的存在下才具有活性。當iNOS 單體與heme結合後會誘發結構型上的改變,使得iNOS可透過本身的 oxygenase domain去形成二聚體,此時的iNOS尚未具有活性,當 arginine或H4B連接到heme與oxygenase domain所形成的結合位置 時,iNOS結構上就會變緊實進而產生活性。可與iNOS promoter結合 的轉錄因子有NF-κB、AP1(Jun/Fos)、C/EBT、CREB以及STAT家 族。而NF-κB及JAK-STAT的路徑可以促進巨噬細胞中inos的基因轉 錄 [42, 43]。
在許多癌症中也可發現 iNOS 的過度表現,其產物 NO(nitric oxide)在過去的研究當中發現可以藉由影響細胞增生、死亡、移行 和血管增生去調控癌症的生成(tumorigenesis)[44]。iNOS-deficient mice 比一般正常 mice 不易引發腫瘤形成,說明了來自 iNOS 的 NO 在癌症上扮演相當重要的角色 [45]。另外,iNOS 高度表現於巨噬 細胞,尤其當受到外在刺激包含內毒素或細胞激素等,會誘使巨噬 細胞產生NO 去毒殺入侵的病原菌,因此 iNOS 的表現量增加象徵著 巨噬細胞的活化以及發炎反應的產生 [46]。
在過去的研究中發現巨噬細胞可以被細菌外膜內毒素包含革蘭 氏陰性菌的LPS和革蘭氏陽性菌的PGN所活化。但無論是細菌是革蘭 氏陰性菌或是陽性菌,只要它的DNA包含有CpG motif就能啟動免疫 反應,而巨噬細胞要移行至外來物入侵處才可將其進行吞噬作用,
進而藉由抗原呈現作用啟動更高層級免疫。因此,移行作用對巨噬 細胞而言為重要且不可缺的功能。由於Src已知可參與在癌細胞的移 行,並引發癌症轉移,所以本論文探討在CpG誘發巨噬細胞移行當 中,Src 扮演何種角色。另外,本實驗室發現在LPS刺激下,iNOS 的存在與否會調控Src而影響細胞移行。在此我們也想知道在CpG誘 發的Src表現及細胞移行是否也受iNOS的調控。
第二章 實驗材料及方法(Materials and Methods)
第一節 實驗材料
一、細胞:
本實驗所用到的細胞 Raw264.7,其為老鼠巨噬細胞株(murine macrophage-like cell line),此株細胞來自於 ATCC(American Type Culture Collection),並且經過v-Abl(abelson)transform。而製作 src siRNA(siRNA-1 和 siRNA-2)細胞和做為對照組非特異性 siRNA
(Ctrl)的細胞則是利用 Raw264.7 巨噬細胞,使用 transfected plasmid DNA pLKO.1-msrc(puro),其目標序列(targeted sequence)為 5’-CGAGCCGCCAATATCCTAGTA-3’或 negative control 的 pSilencer plasmid(其包含的序列在老鼠的基因上不會表現,為 Ambion Ins 所 提供),經由 Lipofectamine invitrogen 的方式,再由 puromycin 或 hygromycin 篩選得到 Src siRNA 細胞。而將 Src 再放回 Src-attenuated 細胞是用 Raw264.7 細胞表現 src siRNA 再 co-transfected pBabe
(hygro)DNA 和 DNA encoding avian c-Src [47]以 hygromycin 篩選 後得到。
活體老鼠之巨噬細胞(rat peritoneal macrophages),簡稱為PEM 細胞,取自於公的 SD rat 或 C57BL/6 老鼠的巨噬細胞,利用 TG
(thioglycolate)注射至老鼠腹腔,使其巨噬細胞聚集,再以 PBS 反 覆沖出細胞後收集。
二、動物:
使用的動物有SD rat,約 250 g 五到七週大之公鼠,訂購自財團 法 人 國 家 實 驗 動 物 中 心 。 另 外 C57BL/6 inducible nitric oxide synthesized knockout(C57BL/6-NOStm1Lau)mice,約 20 g,自行繁殖 於無特定病原(specific pathogen free)環境中,iNOS knockout mice 有正常發育,外觀及標準之顯形(phenotype)。
三、試劑:
1. CpG ODN 1826和control CpG ODN 1826
CpG ODN 1826 為 合 成 之 oligonucleotide , 含 為 甲 基 化 CpG dinucleotides序列,5’- tcc atg acg ttc ctg acg tt -3’共20 mer,為murine TLR9 ligand , 在 分 類 上 為 K ( CpG-B )。 以 200 μg 溶 於 1 ml endotoxin-free water 配 製 為 stock concentration 200 μg/ml , final concentration 0.3 μg/ml。此藥品購自於Invivogen,保存於-20℃。另 外,ODN 1826 control又稱nCpG,作為negative control,其序列為5’- tcc atg agc ttc ctg agc tt -3’,相同的為20 mer,因為將cytosine和
guanine位置對調使其不具活性。
2. PP2
取1 mg 的PP2 溶於5 ml 之DMSO中,配製為stock concentration 20 μg/ml,final concentration 10 μM。藥品購自於Merck,保存於-20
℃。
3. Chloroquine diphosphate salt
取 10 mg 之 chloroquine 溶於 pH 值 4-6 之溶液中,配製為 stock concentration 5 mM,final concentration 10 μM。此藥品對光敏感須 避光,購自於Sigma,保存於室溫下。
4. Quinacrine dihydrochloride
取 5 mg 之 quinacrine 溶於滅菌後之二次水中,配製為 stock concentration 0.5 mM,final concentration 1 μM。此藥品對光敏感須 避光,購自於Sigma,保存於室溫下。
5. 1400W
取 10 mg 之 1400W 溶於 PBS 中,配製為 stock concentration 10 mM,final concentration 100 μM。此藥品購自於 Sigma,保存於-20
℃。
6. ODQ
取 5 mg 之 ODQ 溶於 DMSO 中,配製為 stock concentration 50
mM,final concentration 100 μM。此藥品購自於 Sigma,保存於-20
℃。
四、抗體
1. 購自 Sigma:
Anti-actin(anti-actin mouse monoclonal antibody),保存於-20℃,
抗體比例1:5000~10000。
2. 購自 Upstate:
Anti-iNOS(anti-iNOS rabbit polyclonal antibody),保存於-20℃,
抗體比例1:1000~2000。
3. 購自 Abcam:
Anti-COX-2(anti-COX-2 rabbit polyclonal antibody),保存於-20
℃,抗體比例1:5000。
4. 購自 Santa Cruz:
(1)Anti-Lyn(anti-Lyn rabbit polyclonal antibody),保存於 4℃,抗 體比例 1:1000。
(2)Anti-Hck(anti-Hck goat polyclonal antibody),保存於 4℃,抗
體比例 1:500。
(3)Anti-Fgr(anti-Fgr rabbit polyclonal antibody),保存於 4℃,抗 體比例 1:200。
(4)pTyr(HRP-conjugated mouse pTyr monoclonal antibody),保存 於 4℃,抗體比例 1:1000。
5. 購自 Jackson:
(1)HRP-rabbit anti-goat antibody,保存於-20℃,抗體比例 1:2000
~3000。
(2)HRP- goat anti-mouse antibody,保存於-20℃,抗體比例 1:5000。
(3)HRP-goat anti-rabit antibody,保存於-20℃,抗體比例 1:5000。
6. Anti-Src(anti-Src mouse monoclonal antibody),特別感謝由台灣台 南成功大學呂增宏教授實驗室提供,保存於 4℃,抗體比例 1:200。
以上抗體稀釋後皆保存於-20℃。
五、實驗儀器與耗材:
儀器 廠牌
Centrifuge Eppendrof 5424
Centrifuge Beckman Allegra
Hemacytometer BOECO Hoefer semiPhor Pharmacia Biotech
TE70PH meter JENCO 6071 Power supply EPS 1001 pH meter JENCO 6071
Shaker TKS RS01
Vortex GENIE SI-2 G560 Universal Microplate
Spectrophotometer
BIO-TEK
Automated Strip Washer BIO-TEK
Stirrer/Hot plate CORNING PC640 Vertical slab gel unit Hoefer SE400 Vortex GENIE SI-2 G560 Water bath TKS ZX-400
Boyden chamber Neuro probe
耗材 廠牌
Protein Assay Kit Bio-Rad Polycarbonate filter(8 μm) Neuro probe PVDF transfer membrane NEN
X-15RDigital analysis system Kodak EDAS 120
六、實驗藥品:
1. 購自 Acors(New Jersey, USA):
Deoxycholate、Sodium orthovanadate。
2. 購自 Fisher Scientific(New Jersey, USA):
Methanol、Tween 20
3. 購自 Fisons Scientific Equipment(Bishop Meadow Road, England): Sodium chloride。
4. 購自 Amresco:Bis-acrylamide。
5. 購自 Gibco BRL(New York,NY,USA):
Ammonium persulfate,EDTA,Fetal bovine serum,Glycine,L- Glutamine,Penicillin-streptomycin,Sodium Bicarbonate,TEMED,
trypan blue。
6. 購自 Nunc:
100 X 20 mm culture dish,60 X 15 mm culture dish,15 ml cornical tube,50 ml cornical tube。
7. 購自 Merck:Thioglycolate。
8. 購自 Sigma:
β- mercaptoethanol,Aprotinin,Bromophenol blue,EGTA,IGEPAL CA-630,PMSF,Acrylamide,1400W, ODQ, DMSO。
9. 購自美國 USB:
Coomassie brilliant blue R-250,Tris-base,SDS,Glycine,Tween 20。
第二節 實驗方法
一、細胞培養(cell culture):
1. 試劑:
(1)培養液(RPMI-1640)
培養細胞的培養液為 RPMI-1640,將一單位的 RPMI powder 和2.0 g 的 NaHCO3及5.957 g 的 HEPES 溶於滅菌後的二次水 中,將pH 值調至 7.4。在無菌操作台中,以 0.22 μm 的濾膜過 濾後分裝,保存於4℃。
(2)培養液添加物
小牛的血清白蛋白(fetal bovine serum,FBS),購自 Hyclone 或 Sigma,保存於-20℃,培養液中目標濃度為 10%。
(3)Penicillin-Streptomycin(10X;10000 unit/ml-10 mg/ml),購自 Sigma,保存於-20℃,培養液中目標濃度為 1%。
(4)L-Glutamine(10X;200 mM),購自 Sigma,保存於-20℃,培 養液中目標濃度為 1%。
(5)1X PBS(phosphate balanced solution)
以8 g NaCl、1.2 g NaH2PO4.H2O 加二次水配製,將其 pH 調 成7.4 後補二次水到 1L,滅菌後保存於 4℃。
(6)冷凍培養液
以 2 ml 的 DMSO 加上 8 ml 的 RPMI (只添加 P-S 與 L-Gln, 無含血清)配製成 20% DMSO,若無使用完可存放於-20℃保 存。冷凍培養液,以2.5 ml 的 20% DMSO 與 fetal bovine serum 2.5 ml 混合,最後的 DMSO 濃度為 10%。
2. 步驟:
(1)細胞分盤(subculture)
細胞培養於RPMI medium(10% FBS、1% P-S、1% L-Gln)中,
放置於細胞培養箱(37℃,5% CO2),待細胞成長約八至九分 滿時,進行分盤培養。分盤時將舊的培養液吸掉,以回溫後之 1X PBS 約 3~5 ml 沖洗細胞,共二至三次,將死去的細胞去 除。再加入8~10 ml 回溫後的培養液,將細胞全部沖下來,並 吸放數次直到均勻沖散細胞以1:3 或 1:4 的比例分盤培養。
(2)冷凍細胞
冷凍細胞前一天需更換成新鮮的培養液。培養好約八分滿的細 胞,將舊的培養液去除,再用3~5 ml 之 1X PBS 沖洗細胞,約 二至三次。用8~10 ml 的培養液,將細胞全部沖下來,再將細 胞液吸取置於50 ml 離心管,以 1000 rpm,10 分鐘之離心條件
將細胞沉澱下來。然後去除上清液,以1 ml 冷凍培養液將細 胞打散,置於冷凍管中,先存放於 4℃冰箱 30 分鐘,再移置 於-80℃冰箱,12 小時之後再存放於液態氮桶。
(3)解凍細胞
由液態氮桶中取出細胞,將細胞溶於適量的RPMI 培養液,離 心2000 rpm,5 分鐘。倒掉舊的 RPMI 培養液,再用 10 ml 的 RPMI 將細胞打散,放置隔天細胞貼附後再換培養液,觀察約 一個禮拜等細胞狀況穩定後再做實驗。
二、收集細胞 lysates:
1. 試劑:
主要溶液Lysis buffer(RIPA buffer),以 4.38 g NaCl、3.0285 g Tris-base、1.25 g Deoxycholate、5 ml IGEPAL CA-630(相當的粘稠,
要緩慢的取出)來配製,以上藥品溶於適量的二次水後,將配成 pH 值為7.4,500 ml 溶液,高溫高壓滅菌後保存於 4℃,用以將細胞裂 解。當要收集細胞,再配置成modified RIPA buffer,內含蛋白質水 解酶抑制劑。若要配製成1 ml,則需要 1 ml RIPA buffer、5 μl 之 200 mM Sodium orthovannadate(final conc.= 1 mM)、5 μl 之 200 mM EGTA(final conc. = 1 mM)、5 μl 之 0.5% Aprotinin(final conc. =
0.0025%)、5 μl 之 200 mM PMSF(final conc. = 1 mM)。Sodium orthovannadate 是 phosphatase inhibitor,EGTA 是鈣離子的螯合劑,
Aprotinin 是 serine protease inhibitor,PMSF 也是 serine protease inhibitor。如實驗需求亦可多追加一倍的抑制劑。
2. 步驟:
先測量整體medium 重量,取 1 ml medium 保存起來,以利往後 對於分泌型蛋白的實驗。去除舊的medium 後,以 1X PBS 清洗兩至 三次,將PBS 吸乾以免影響蛋白質濃度。再加入適量 modified RIPA buffer(視細胞的滿度而定,滿盤的細胞約用 800 至 1000 μl),接著 用刮杓將細胞刮下(冰上操作),後置於eppendorf。再把 eppendorf 在 rack 上刮動,來回 3~5 次以幫助裂解細胞。以 4℃、10000 rpm,離 心10 分鐘後取上清液到新的 eppendorf,接著定量並且 denature 後直 接進行Western blot,若需存放則置於-80℃冰箱。
三、蛋白質濃度測定:
1. 試劑:
BSA 1 μg/μl,Protein Assay Kit
2. 步驟:
首先配製BSA 1 μg/μl 作為製作標準曲線之用,配製含不同濃度 的BSA 溶液 800 μl(例如配置濃度間隔 5,0~30 μg BSA),再加入 200 μl Protein Assay Kit(dye),混合均勻後靜置10 分鐘。各別取 200 μl,加入 96 孔盤中,避免氣泡產生影響吸光值,再以 ELISA reader 來測定 595 nm 波長的吸光值,劃出標準曲線(R-squared>0.99)。
在樣品的測定上,取10 μl 的 cell lysate,加入 790 μl 的二次水與 200 μl 的 protein assay kit(dye),混合均勻後靜置 10 分鐘,取 200 μl 加 入96 孔盤中以 ELISA reader 測定 595 nm 的吸光值。將所得的吸光 值帶入標準曲線求得原樣品的濃度。
四、蛋白質電泳(SDS-PAGE)
1. 試劑:
(1)30% Acryamide/0.8% bisacryamide
配 置 方 法 為 150 g Acryamide 與 4 g N’N’- methylene bisacryamide 溶 於 二 次 水 中 , 可 配 置 成 500 ml 的 30%
Acryamide/0.8% bisacryamide,避光保存於 4℃。這是電泳過程 中分離蛋白質的介面之用。
(2)Tris-base(2 M , pH 8.8, pH 6.8)
配置方法為,以121.14 g Tris-base 溶於適量二次水中,再配製 成500 ml pH 8.8 或 6.8 的溶液,保存於 4℃。Tris 在電泳的過 程當中主要扮演調整pH 值的角色。上層 pH 6.8 的中性環境可 讓glycine 帶正電,讓電泳時蛋白移動緩慢以達到 stacking 的功 能,而下層pH 8.8 的鹼性環境有助於分離含 SDS 的負電蛋白。
(3)10% APS 與 TEMED
配置方法為1 g APS,溶於 10 ml 二次水,配製的目標濃度為 10% APS,保存於-20℃。在 acryamide 與 bisacryamide 作用的 過程中,APS 扮演著催化的角色,而 TEMED 則扮演輔酶的角 色,TEMED 則是購買自 Sigma 的藥品,保存於 4℃。
(4)10X sample buffer
配製方法為1 ml 的 1M Tris-HCl pH 6.8、5 g SDS、25 g sucrose、
10 mg bromophenol blue、5 ml 2-mercaptoethanol 溶於二次水,
再加二次水至500 ml,保存於室溫。SDS 的作用在於使蛋白質 帶 負 電 ,sucrose 可 使 整 個 樣 品 溶 液 黏 稠 不 易 飄 散 , β-mercaptoethanol 有助於打斷雙硫鍵幫助變性,而 bromophenol blue 可用來調整溶液的顏色以方便樣品注入。Sample buffer 目 的在於混合樣品後有助於denature,且讓蛋白帶負電性。
(5)5X running buffer
配製方法為7.5 g Tris-base、36 g Glycine、2.5 g SDS,溶於水 配製成 500 ml,保存於室溫,使用前再稀釋成 1X running buffer。主要為電泳過程中通電的介質,其中藥品的比例會影 響到整個電泳過程的穩定度。
(6)Resolving gel(8%)
配製方法為15.8 ml 之二次水、6 ml 之 2 M Tris- base (pH 8.8)、8 ml 之 30% Acryamide/0.8% bisacryamide、150 μl 之 20%
SDS、150 μl 之 10% APS、20 μl 之 TEMED。
(7)Stacking gel
配製方法為 8 ml 之二次水、0.625 ml 之 2 M Tris-HCL(pH 6.8)、1.33 ml 之 30% Acryamide/0.8% bisacryamide、50 μl 之 20%SDS、50 μl 之 10% APS、10 μl 之 TEMED。
2. 步驟:
將兩塊玻璃板、兩條 spacer 及灰色夾組合成一個製膠架,然後 利用黑色旋鈕固定在底座。將下層膠(resolving gel)配方所需的藥 劑依序加入燒杯,混合攪拌均勻,利用 pipette 加進製膠架中至適當 位置。以75%酒精加滿,利用不同極性的溶液來使 gel 平整,待凝膠
(約25 分鐘)。將酒精倒掉,插入間格髮梳(comb),將上層膠(stacking gel)配方所需的藥劑依序加入燒杯。攪拌均勻,利用 pipette 加到製 膠架中至適當位置。等上層膠凝固之後(約25 分鐘),將 comb 拿走,
倒入上、下層各250 ml 1X running buffer。將 sample 定量完和 10X sample buffer(10:1)混和均勻後,以乾式加熱器煮沸 5 分鐘,然後 稍微離心一下,用micropipette 把樣品注入 well 中,進行膠體電泳法
(50 V、20 mA,約時間 18 小時;90 V、22 mA,時間 12 小時)。
五、Western blot analysis:
1. 試劑:
(1)Transfer buffer
配製方法為以1.5 g Tris-base、7.2 g Glycine、0.5 g SDS、100 ml Methanol 溶入二次水配製成 500 ml,保存於室溫。通常一次可 配大量保存,過程中會有許多氣泡以及少許的放熱。
(2)Blotting buffer
配製方法為以5 ml 10X TBS、1.5 g BSA、25 μl Tween 20 補二 次水至 50 ml,保存在 4℃。主要用來將轉印後的 PVDF 膜做 blocking 的動作,以免會有抗體非專一性的辨識。由於 BSA 是
(3)10X TBS and washing buffer
配製方式為30.3 g Tris-base 與 43.83 g NaCl 溶於二次水後,pH 調成7.4,再補水至 500 ml,保存於室溫。TBS 主要用來調整 blotting buffer 與 washing buffer 的 pH 值,Washing buffer 則是 以50 ml 的 10X TBS、500 μl Tween 20(1%)加二次水到 500 ml,主要的功能在洗去非專一性不緊密連接以及未連接的抗 體。
2. 步驟:
剪取適當大小的 PVDF transfer membrane(14.5×12.5 ㎝),以 methanol 浸泡約 30 秒後,再以二次水浸泡 2 分鐘。濾紙使用前浸泡 transfer buffer,依順序三張濾紙、gel、PVDF、三張濾紙,利用 semi-phor transblotter 將下層膠中的蛋白質 transfer 到 PVDF transfer membrane
(25V、300 mA,1.5 小時)。之後將PVDF transfer membrane 取出,
浸泡於適量的blotting buffer 中,在室溫下置於 shaker 上 preblotting 2.5 小時(shaker 擺動速度:5rpm),進行 blocking 的動作。之後將 blotting buffer 倒掉,加入適量含有 primary antibody(依不同抗體有 不同稀釋倍數)的 blotting buffer,於 4℃下 overnight。隔天取出之 後,在室溫下先置於shaker 上作用 30 分鐘(shaker 擺動速度:5rpm),
再以適量 washing buffer 清洗 membrane 三次,5 分鐘、5 分鐘、10 分鐘(shaker 擺動速度: 30 rpm)。加入適量含 secondary antibody 的 blotting buffer(依不同抗體有不同稀釋倍數),室溫 blotting 1.5 小時
(shaker 擺動速度: 5 rpm)。再以適量 washing buffer 清洗 membrane 五次,5 分鐘、5 分鐘、10 分鐘、15 分鐘、15 分鐘(shaker 擺動速 度: 30 rpm)。依 membrane 大小(0.125 ml/cm2),加入適量的ECL kit solution I 與 II 等比例的均勻混和液。在 membrane 與之作用 5 分鐘 後,以X-ray film 感光。
六、細胞移動性分析(cell migration assay)
1. 準備物品:
(1)48-well Boyden chamber,必須以清潔劑清洗過後晾乾。
(2)Polycarbonate filter(8 μm),分有正反兩面,正面光滑而反面 粗糙,正面朝上反面朝下,當細胞穿透後會被吸附在反面上,
再以染色觀察細胞。
(3)Trypan blue,可以將死細胞染色,被染到的死細胞呈現藍色而 活細胞則呈現透明無色。
(4)甲醇(約 20 ml),主要功能為固定細胞用。
(5)Giemsa stain reagent(約 20 ml),細胞染色用,染到的細胞呈
現紫紅色或深紫色。另外還需準備血球計數器來計算細胞量。
2. 步驟:
細胞先分盤至6 cm culture dish,根據實驗組別,依時間處理藥 劑後準備開始實驗。收取1 ml 的培養液,方便做分泌型蛋白的測試。
以1X PBS 清洗細胞 3 次,每次 3 ml,之後以適量的 serum free 的 RPMI medium 將細胞沖下打散。取 20 μl 細胞加入 20 μl 的 trypan blue
(1:1)進行細胞計數,將細胞濃度調整至 2×104個/50 μl。取出 48-well Boyden chamber,在 lower chamber 中,每個 well 放入 30 μl 含有 10 % FBS 的 RPMI medium。之後再依序將橡膠墊片,polycarbonate filter
(8 μm)及 upper chamber,以對稱的方式鎖緊 chamber。polycarbonate filter 為了方便辨識正反面,在膜上右邊做個截角。在 upper chambe 中,每個well 放入 20000 個細胞,最後再以保鮮膜包住整個 48-well Boyden chamber,放置於 37℃,5% CO2 細胞培養箱中,使細胞進行 爬行實驗5 小時。取出 48-well Boyden chamber 之後,依序分解,將 polycarbonate filter 正面朝上浸泡於 20 ml 甲醇中 10 分鐘,進行細胞 固定。固定完的polycarbonate filter 以反面朝上的方式放在乾淨的地 方風乾。之後做細胞染色的步驟,將polycarbonate filter 浸泡於 20 ml 的Giemsa stain reagent 1.5 小時,再以 d.d H2O 將染色退去,待退染
完成後,將polycarbonate filter 固定於載玻片上,利用顯微鏡(400X)
觀察細胞爬行狀況,並將其量化作圖。
七、腹腔巨噬細胞製備及培養
1. 事前準備:
50 ml 離心管,把剪刀、針筒、針頭、塑膠 dropper 用資料袋封好,
照UV overnight。並準備手套、衛生紙、酒精、剃毛刀、感染性垃 圾袋。
2. 步驟:
將4-6 weeks 的 SD rat(約 250~300 g)做腹腔注射,打入 3%之 Brewer thioglycolate medium,每隻 10 ml。C57BL/6 和 C57BL/6 inducible nitric oxide synthesized knockout mice,每隻打 1 ml。誘發腹 腔的巨噬細胞聚集。4 天後,方可實驗。實驗前先將 37℃回溫過的 PBS 分裝到 50 ml 離心管內,並把所有東西準備齊全後,將滅菌後的 物品放入laminar flow。使用 laminar flow 前以酒精消毒桌面,並以 噴過酒精的衛生紙平鋪於桌面上,待實驗物品放入後照 UV 30 分 鐘。之後前往犧牲老鼠,以 CO2處理犧牲老鼠後將腹部的毛剃光,
噴酒精以衛生紙把所有的鼠毛清乾淨避免污染。將老鼠放在 laminar flow 桌面上的衛生紙上。利用 37℃回溫的 PBS 打入腹腔中,按摩約
1 分鐘後,剪開老鼠的腹腔,將剩下的 PBS 以針筒加入。一隻老鼠 以100 ml PBS 將巨噬細胞溶出,一次注入最多 50 ml,分數次注入 與吸出來完成實驗。緩慢將PBS 回抽(此時 PBS 為混濁,並且盡量 避免產生氣泡)。將回抽的 PBS 離心(2000 rpm 20 分鐘),去除上清 液。以適量的RPMI(10%FBS、1% P-S、1% L-Gln)將細胞輕輕打 散。在37℃ 5% CO2下培養在培養皿中,待細胞貼附約 5 小時,去 除舊的 RPMI 培養液,以 37℃之 PBS 沖洗五次,再換上新的 RPMI 培養液,繼續培養24 小時待做實驗。
第三章 結果(Results)
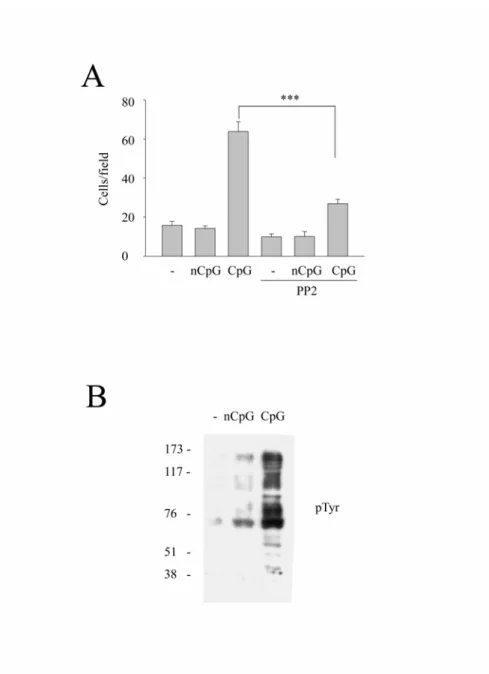
一、Src family kinase 參與在 CpG 誘發的巨噬細胞移行中
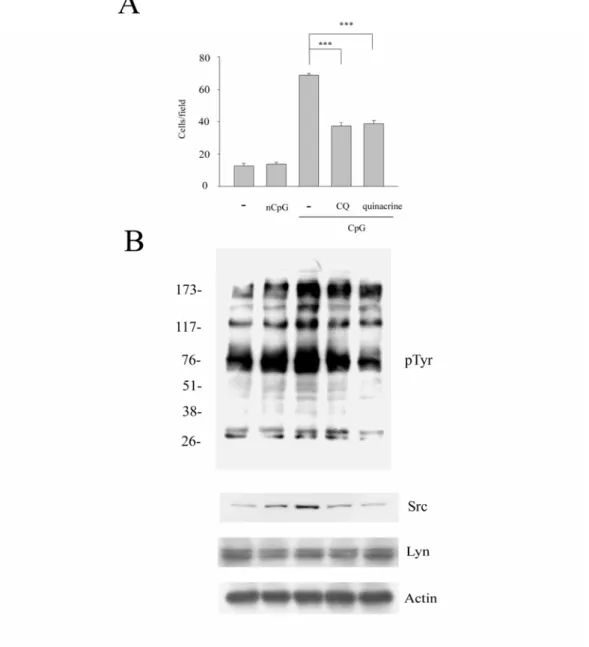
在過去的文獻中指出 CpG 透過 TLR9 的辨識而產生一連串的免 疫反應。細胞移行為巨噬細胞重要功能之一,而 Src 在過去的認知 中,參與在癌症細胞增生及轉移過程。所以我們想知道 Src 在 CpG 誘發的細胞移行能力中所扮演的角色。使用Raw264.7 巨噬細胞,在 加入CpG 或 nCpG 前 20 分鐘先加 PP2(10 μM)處理,經 48 小時後 再比較各組細胞移行的能力。由Fig. 1A,我們發現在加入 CpG(0.3 μg/ml)的組別中,細胞移行的能力明顯較未加藥及 nCpG(0.3 μg/ml)
組別多;但在合併PP2 處理下,可發現原本 CpG 誘發的細胞移行的 能力會被PP2 所抑制,而 PP2 為 Src family kinases 的抑制劑。這樣 的結果也透露Src family kinases 可能參與在 CpG 誘發的細胞移行中。
因為Src family kinases(SFKs)為 nonreceptor tyrosine kinases,
而 酪 氨 酸 (tyrosine ) 磷 酸 化 可 牽 動 肌 動 蛋 白 細 胞 骨 架 ( actin cytoskeleton)重組,導致細胞延伸(cell spreading)、黏附(adhesion)
和移動(motility)等 [48]。所以我們進一步去看在巨噬細胞受 CpG 的刺激下,酪氨酸磷酸化的情形。而由 Fig. 1B 可以得知,在 CpG 刺激下,酪氨酸磷酸化的增加量大於未處理及nCpG 組別。
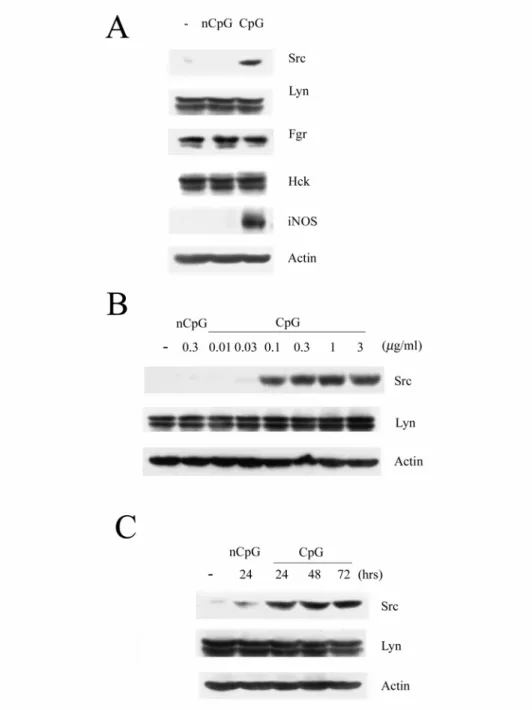
二、Raw264.7 巨噬細胞在 CpG 的刺激下會增加 Src 的表現量 進一步我們想知道到底是哪個 Src 家族成員參與在 CpG 引發的 巨噬細胞移行。根據過去的文獻,我們知道許多細胞會表現Src,而 Lyn、Fgr 和 Hck 是巨噬細胞中表現最多的 SFKs [34]。所以我們使用 Raw264.7 巨噬細胞加入 CpG 或 nCpG(0.3 μg/ml)處理 48 小時後分 析Lyn、Fgr 和 Hck 的蛋白表現量。由 Fig. 2A 可知 CpG 刺激後只有 Src 量表現增加,而 Lyn、Fgr 和 Hck 的蛋白量無論有無加藥處理皆 沒有很大的差異。
另外,我們用不同的 CpG 濃度處理 Raw264.7 細胞,其濃度分 別為0.01、0.03、0.1、0.3、1、3(μg/ml)。由 Fig. 2B 可知隨著濃度 的增加,Src 的表現量也隨之增加;在 CpG 濃度為 0.3 μg/ml 時,Src 表現量增至一個穩定值。以CpG 0.3 μg/ml 處理 Raw264.7 細胞不同 時間,我們同樣可發現 Src 在 CpG 的刺激下其蛋白會隨時間的增加 而增加(Fig. 2C)。由於 CpG 處理 48 小時後的 Src 表現量最大,所 以後續的實驗所選擇CpG 的濃度為 0.3 μg/ml,時間為 48 小時。
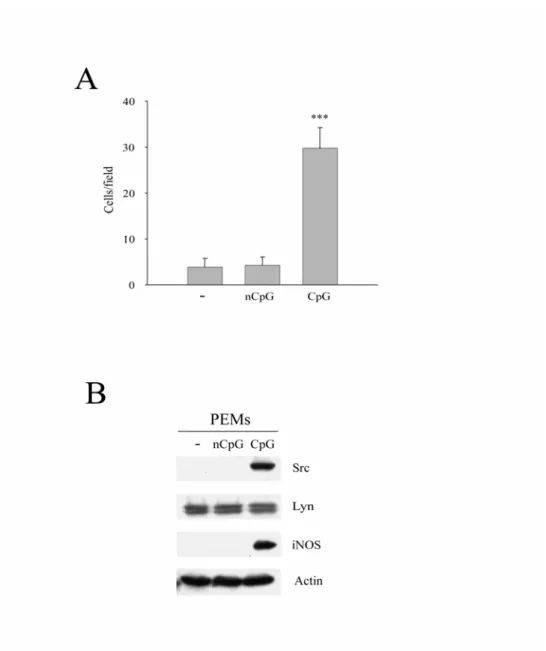
三、老鼠腹腔巨噬細胞在 CpG 的刺激下會增加 Src 的表現量和細胞 移行
我們在巨噬細胞的細胞株中發現 Src 參與在 CpG 所誘發的細胞 移行。同時我們也想知道類似的現象是否也在活體巨噬細胞中發 生,所以我們以腹腔注射的方式將TG(thioglycolate)注入老鼠(rat)
腹腔,使巨噬細胞聚集。四天後,以 CO2將老鼠犧牲所取得的腹腔 巨噬細胞(peritoneal macrophages;PEMs),我們以 CpG 或 nCpG 0.3 μg/ml 處理 48 小時後觀察其 Src 的表現量和細胞移行的能力。相較 於未加藥和nCpG 組別,CpG 會增加 PEM 的移行能力(Fig. 3A),
而在Src 表現增加的同時,Lyn 的表現並無變化(Fig. 3B)。iNOS 在 此做為巨噬細胞活化的 positive control。由上述結果可知 CpG 所誘 發的Src 增加及細胞移行不僅發生在巨噬細胞的細胞株上,同時也可 見於活體的巨噬細胞。
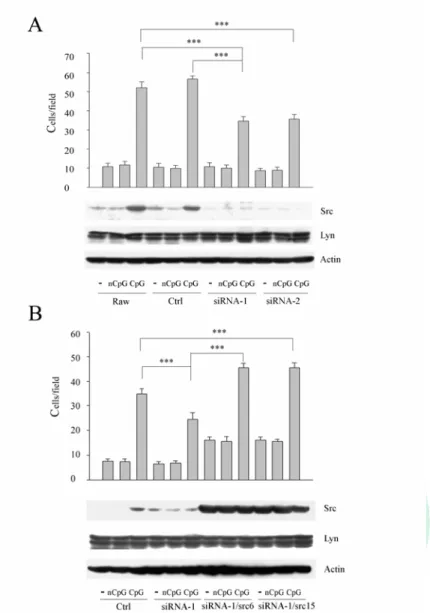
四、Src 參與在 CpG 誘發的巨噬細胞移行中
接下來我們想確認 Src 在 CpG 誘發的細胞移行過程的上扮演的 角色。首先我們用 src siRNA 細胞去探討在 Src 表現量下降時,CpG 誘發巨噬細胞的移行是否受到影響。Raw 細胞和 control 組別在 CpG 刺激後,細胞移行能力增加。但 CpG 刺激後的 src siRNA 細胞
(siRNA-1,siRNA-2),其移行能力相較於 Raw 細胞和 control 組別 明顯減少。雖然CpG 誘發的 Src 在 siRNA-1 及 siRNA-2 細胞中表現 減少,但Lyn 的表現並無變化(Fig. 4A)。所以 Src 在 CpG 引發的細 胞移行扮演重要的角色。
但我們不能排除 siRNA 非特異性的因素,所以我們再將 Src 放 回去 src siRNA 細胞中,去看當 Src 表現恢復後是否也能恢復原本減 少的細胞移行能力。我們比較siRNA-1 及 siRNA-2 細胞和 Raw 細胞 組別,發現在 siRNA-1 和 siRNA-2 細胞中,當 Src 表現量下降,同 時細胞移行能力減弱。但當將 Src 放回去 siRNA-1 (siRNA-1/Src6,
siRNA-1/Src15) 細胞時,可以發現不僅 Src 表現量增加,同時細胞 移行能力也增加了(Fig. 4B)。所以在此我們證明了Src 在 CpG 誘發 巨噬細胞移行能力中的重要地位。
五、在巨噬細胞中在 CpG 的刺激下,增加的 Src 表現量和細胞移行 移行能力是透過 TLR9-dependent 的機制
根據2006 年 JCB 的文獻指出 CpG 誘發的酪氨酸(tyrosine)磷 酸化可經過 TLR9-independent 的機制 [49]。而 Src 為 nonreceptor tyrosine kinase,所以我們也想知道 CpG 誘發的 Src 和細胞移行能力 到底是為TLR9-independent 或是 TLR9-dependent 的路徑。為回答這 個問題,我們使用 Raw264.7 巨噬細胞,在加入 CpG 或 nCpG 前 30 分鐘先加 chloroquine(10 μM)或 quinacrine(1μM )處理,經 48 小時後再比較各組總體蛋白的酪氨酸磷酸化。我們發現CpG 處理下 的 總 體 蛋 白 酪 氨 酸 磷 酸 化 增 加 , 但 在 合 併 處 理 chloroquine 或 quinacrine 後細胞的總體酪氨酸磷酸化下降(Fig. 5B)。同時在TLR9 抑制劑處理下所 CpG 誘增之 Src 的表現量減少而細胞移行受抑制
(Fig. 5A)。所以我們認為在長時間 CpG 刺激下所增加的細胞移行 與Src 的表現量主要循 TLR9-dependent 的路徑。
六、1400W 和 ODQ 可抑制 Src 表現量和巨噬細胞移行能力
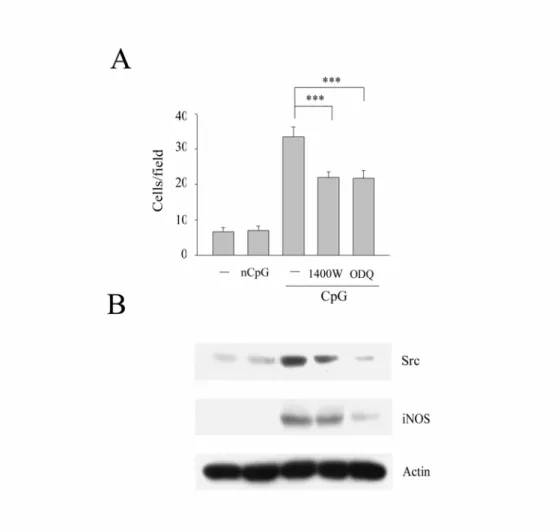
另外,本實驗室發現在 iNOS knockout 老鼠腹腔的巨噬細胞經 LPS 刺激後,Src 的表現量減少,同時細胞移行能力也下降;而在加 入NO donor 和 NO 下游產物 cGMP 的類似物 8-br-cGMP 後,發現可 恢復 Src 的表現量和細胞的移行。所以 iNOS 和其下游產物 NO 及 cGMP 被證實了可以調節 LPS 誘發的 Src 和細胞移行能力。有了這 樣的結果,我們也想知道是否 iNOS 也影響 CpG 誘發的 Src 和細胞 移行能力。為了回答這個問題,我們先以1400W(iNOS 的抑制劑),
和其下游產物sGC 的抑制劑 ODQ 去處理 Raw264.7 細胞,再觀察其 對CpG 所引發的 Src 表現和細胞移行有無影響。我們發現 CpG 所誘 增的Src 的表現量(Fig. 6B)和細胞移行能力(Fig. 6A)在 iNOS 和 sGC 的抑制劑的處理下皆減少了,所以 iNOS 和 sGC 參與在 CpG 誘 發Src 的表現量和巨噬細胞移行能力中。
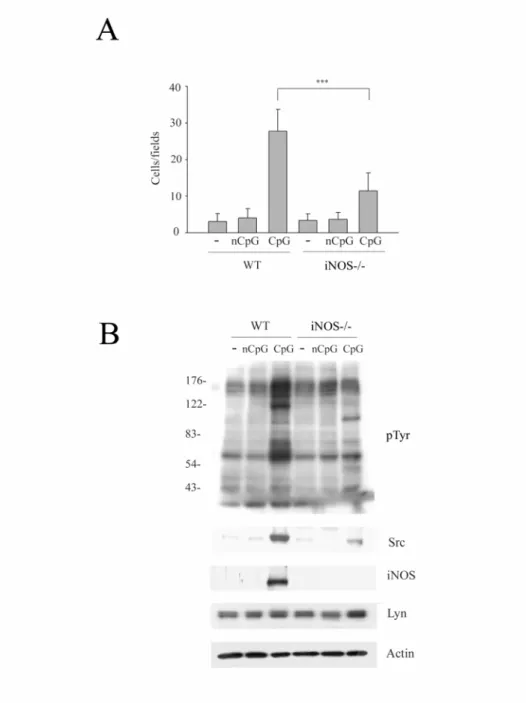
七、CpG 處理巨噬細胞後 Src 表現量和移行能力的增加需要 iNOS
進一步我們用iNOS knockout 的老鼠證實 iNOS 的角色。在“野 生型"組別(wild type),CpG 處理後整體蛋白的酪氨酸磷酸化增加,
但是在iNOS 不存在下,這個現象就不見了(Fig. 7B)。經 CpG 處理 後 iNOS-/-巨噬細胞其 Src 的表現量減少的同時,細胞移行能力也明 顯下降(Fig. 7A)。因此我們認為巨噬細胞中 Src 表現量及細胞移行 的增加,不獨見於 LPS 刺激,也見於 CpG 處理。所以在 CpG 誘發 的Src 的表現量和巨噬細胞移行能力中是需要 iNOS 的。
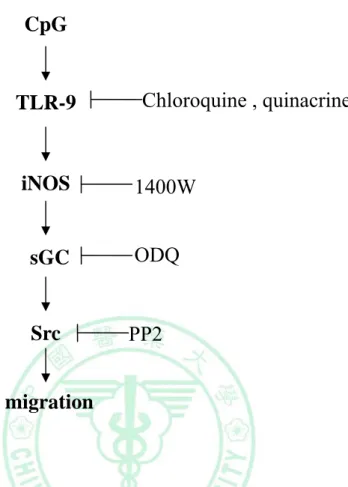
綜合上述實驗結果,我們認為巨噬細胞在長時間 CpG 的刺激 下,Src 表現量的增加是透過 TLR9-dependent 的機制,而影響巨噬 細胞移行。我們也證明了 iNOS 是 TLR9-dependent 的路徑中的關鍵 蛋白,其活性牽動Src 的表現與巨噬細胞的移行能力。所以 iNOS 對 於Src 的表現和巨噬細胞的移行能力是不可或缺的。
第四章 討論(Discussion)
一般認為 CpG DNA 是由位於細胞內 endosome 的 TLR9 所辨 識,而具有引發一連串免疫反應的能力。過去有研究者在表達 hemaggutinin-tagged TLR(HA-TLR9)的 HEK293 細胞加入 CpG ODN 刺激並和Cy3 plus FITC-labeled TLR 抗體一起培養,再利用共軛焦顯 微鏡(confocal microscopy)觀察細胞內狀況。在沒有 CpG DNA 刺 激下,帶有TLR9 分子的 endocytic vesicles 只在細胞膜觀察到 [50]。
在CpG ODN 刺激下可觀察到 Cy3 螢光標記上的 TLR9 vesicles 在 10 分鐘內就移動至接近細胞核。在二小時後CpG ODN 在 endosome 上 的大小和量都明顯增加,但在表達缺乏intracellular domain (ICD)的 TLR9 之 HEK293 細胞則看不到上述現象。此外有文獻使用帶有 GFP-tagged MyD88 的 Raw264.7 巨噬細胞做實驗。在 CpG ODN 刺激 45 分 鐘 後 , GFP-tagged MyD88 會 聚 集 到 lysosomal-associated membrane protein 1 (Lamp-1)-positive endosomal compartment [51]
中。TLR9 的訊息透過 MyD88 的連接蛋白(adaptor protein)聚集
(recruited ) 到 TIR ( toll/IL-1R homologous region) 之 後 活 化 IRAK1-TRAF6-TAK1 路徑 [8]。但其不像 TLR4 調節路徑包含 TIR domain-containing adaptor protein/MyD88-adaptor like(TIRAP/MAL)
[14]。近年來有不少和 MyD88-dependent 和 MyD88-independent 相關
連接蛋白的探討 [52]。在前言中也提到一般 TLR9 訊息傳遞包含 MAPK 路徑像 p38,c-Jun,ERK 和 NIK-IKK-IκB 路徑。在巨噬細胞 中 CpG DNA 會活化 ERK 去引發 IL-10 的產生,但在樹狀細胞
(dendritic cells)和 B 細胞卻不會。另外在巨噬細胞中 CpG DNA 也 會誘發IFN-β 的產生,藉由利用 IFN-α/β receptor 使 STAT1 磷酸化並 產生 IL-10 [53]。CpG DNA 也會活化一些 transcription factors 包含 NF-κB、AP-1、C/EBP、CREB,進而影響細胞激素和化學激素的表 現 [23, 25, 26]。
過去的研究指出,單核球(monocytes)受到CpG DNA的刺激後 其酪氨酸磷酸化增加,而主要參與的SFKs為Hck和Lyn [49]。但其CpG DNA刺激細胞時間很短(<10分鐘)。相較於本文,我們以CpG DNA 刺激細胞48小時,酪氨酸磷酸化也有增加的現象。在合併處理PP2
(SFKs的抑制劑)後,巨噬細胞的移行減少。由於CpG DNA刺激的 Raw264.7和老鼠腹腔巨噬細胞皆有Src表現及移行增加,故Src被認為 參與了CpG誘發的細胞移行。的確,在Src-attenuated細胞,CpG誘發 的細胞移行減少。但將Src重新置入siRNA-1細胞,在Src表現量恢復 的同時其移行能力也增加了。所以我們認為Src在CpG誘發的細胞移 行中扮演不可或缺的角色。
Williams和Ridley研究中特別提到可為SFKs磷酸化的paxillin可
能 可 以 直 接 參 與 在LPS誘 發 的 巨 噬 細 胞 黏 附 和 延 伸 。 Paxillin為 cytoskeletal adaptor protein,其功能為支撐(scaffold)一些特別的蛋 白像是Crk, c-Src, PKL, FAK和Pyk2去促使細胞黏附 [48]。在其後的 文獻也發現CpG DNA誘發的巨噬細胞黏附是需要SFKs的,藉由SFKs 中的Hck去磷酸化Paxillin,再使Crk磷酸化和Paxillin結合,這樣的 Paxillin-Crk complexes 改 變 細 胞 骨 架 的 排 列 促 使 巨 噬 細 胞 延 伸 [54]。而Src和FAK及Pyk2磷酸化可以調節Paxillin使細胞骨架重組而 使細胞能進行黏附、延伸和lamellipodia的形成 [55]。CpG也可以誘 發Pyk2, Cbl, Vav的磷酸化 [56],而這些蛋白質在細胞黏附和移行上 是必須的 [57]。Cbl又可以和Pyk2、Vav和SFKs作用增加巨噬細胞的 移行能力 [58]。其中Vav影響了內含PI3K的細胞骨架,也調節了細 胞激素及NF-κB活化的路徑 [59]。而在我們的實驗室發現CpG誘發的 巨噬細胞移行是需要Src的,這和過去的研究不謀而合。
在過去的文獻中也提到Src會透過影響NF-κB的活性而調控細胞 激素的分泌,splenocytes在前處理SFKs抑制劑下會使CpG誘發的IL-6 減少,而在人類plasmacytoid dendritic cells(pDCs)中IFN-α製造也 減少 [60]。在MyD88-/-的老鼠中仍見SFKs的活性,CpG刺激下的路 徑同時具備透過MyD88和MyD88-independent的路徑 [49]。在2006 年Miguel等人提出在CpG刺激下,巨噬細胞的蛋白就有快速酪氨酸磷
酸化增加的現象。由於TLR9抑制劑並無法抑制酪氨酸磷酸化,故暗 示著CpG在細胞膜上就能引發反應,而不無需藉助位於endosome的 TLR9 [49]。有趣的是TLR9-/-和MyD88-/-的單核球在其細胞膜上,CpG 可誘發SFKs活性增加並使細胞骨架重組進而增加細胞黏附和移行能 力。這些是TLR9和MyD88 independent的路徑,特別是這樣的路徑卻 無 法 誘 發IL-6的 分 泌 。 另 外 一 篇 利 用 病 毒 DNA 去 感 染 TLR9-/-和 MyD88-/-的老鼠發現leukocyte聚集,但在細胞激素分泌上卻是減少的 [61]。
在我們實驗室使用TLR9抑制劑chloroquine或quinacrine後發現 細胞的總體酪氨酸磷酸化下降,同時CpG誘增之Src的表現量和細胞 移行皆減少。這樣的結果暗示著在長時間CpG的刺激下增加的Src和 巨噬細胞移行是受到TLR9的影響,但似乎和Miguel等人於2006年 [49] 所認為CpG誘發的酪氨酸磷酸化是透過TLR9 independent的路 徑是有出入的,這有兩個解釋,第一為在JCB文獻中使用CpG刺激為 短時間,TLR9抑制劑和TLR9-/-的老鼠其酪氨酸磷酸化並無明顯減 少,反而是SFKs的抑制劑PP2處理下會影響細胞黏附能力。其中Hck 和Lyn扮演影響細胞黏附的主導角色,在其兩個蛋白的siRNA細胞中 黏附能力和TNFα分泌皆減少。所以作者認為CpG DNA的活化有兩 個 階 段 , 首 先 由SFKs 調 節 細 胞 黏 附 、 伸 展 和 移 行 , 此 為 TLR9