國立宜蘭大學自然資源學系 碩士論文
Department of Natural Resources National Ilan University
Master Thesis
棲蘭山檜木林冠層下檜木栽植苗之生長表現 Growth Performances of Planting Seedlings of Taiwan Cypress under Canopy of Taiwan Cypress Forest in Chi
lan-Shan
指導教授:林世宗 博士 Dr. Shu-Tzong Lin Ph.D.
研究生:林進龍 Chin-lung Lin
中華民國九十六年七月
中文摘要
棲蘭山區位盛行雲霧帶林是以檜木為優勢種,包括台灣扁柏(Chamaecyparis obtusa var. formosana)及紅檜(Chamaecyparis formosensis),森林保育處曾移除部份之枯倒木進 行檜木保育更新作業。本研究以棲蘭山檜木天然林經移除檜木枯立木及倒木後,所形成 冠層環境下,針對檜木更新苗階段之環境適應能力進行評估。
試驗地設置於棲蘭山 130 線林道檜木林枯倒木整理區,其上層林冠水平結構變化,
在枯倒木整理後,林冠孔隙變動不大,整理後 6-7 年孔隙面積明顯增加,孔隙呈聚集現 象,分散度變化不大,推測颱風為主要干擾因子。以林分視覺景觀系統(Stand Visualization System ;SVS)解析林分垂直結構,上木以台灣扁柏為優勢;中、下層則以闊葉樹為優 勢。在此異質林冠下,以栽植二年生之均質檜木苗模擬檜木更新幼苗階段進行試驗,並 以半球面鬱閉度計(spherical densiometer)調查各樣區上層之林冠鬱閉度,區分為疏、
中、密三級,比較不同冠層環境下紅檜及台灣扁柏栽植苗,於 9 年間苗木發育階段存活 與生長表現,探討冠層環境下檜木初始更新後之適應策略。
栽植初期,在密林冠環境下,台灣扁柏苗木死亡率較紅檜低,而在疏林冠環境下紅 檜苗木死亡率較台灣扁柏苗低,顯示台灣扁柏苗木較紅檜苗木耐蔭。兩種檜木苗於 4 年 生後疏鬱閉林冠環境下死亡率較高,亦發現在此環境易遭受囓齒類動物危害。
在各林冠環境下,9 年間紅檜苗高及基徑表現皆優於台灣扁柏。兩種檜木苗年連生 長量多以密林冠環境下為低,而於林冠間之連年生長量差異,則以台灣扁柏苗木較紅檜 為明顯,顯現台灣扁柏幼樹於密林冠下之忍受性。苗木生長率以初期階段為高,在疏林 冠下者優於密林冠下者,且紅檜苗高及苗徑生長率大於台灣扁柏,後期苗木生長率下降 且於樹種間及林冠環境間差異不大;然在 8 年生後於密林冠環境下台灣扁柏苗高生長量 與生長率卻有高於疏林冠者之趨勢。苗木健壯商數以密林冠下較高,樹種間亦以紅檜為 高,表現紅檜對光需求性較高。
由檜木幼樹於不同林冠鬱閉度下之死亡率、生長性狀及其相關性分析,表現檜木在 林冠環境下之適應策略,台灣扁柏為遮蔭忍受型,紅檜為遮蔭逃避型。
關鍵詞:檜木更新、紅檜、台灣扁柏、林冠異質性、耐蔭性、苗木適應策略
Abstract
Yellow cypress (Chamaecyparis obtusa var. formosana) and red cypress(Chamaecyparis
formosensis)were dominant tree in cloud forest zone located Chilanshan area of Northestern
Taiwan. Partial snag and dead-fall cypress tree were removed in some natural cypress forests by The Forest Conservation Administration for processing cypress resources conservation and regeneration option. This study focused on growth adaptability of cypress saplings under canopy environment after the removal work.The study site was set up in the nature forest after remove treatment on line 130 forest road in Chilanshan. The canopy gap dynamic was stability after treatment in the early phase but the gap patch show more clumpy during 6-7years. The mechanism may be by typhoon disturbance. With stand visualization system program(SVS), the canopy vertical structure of the forest show that main canopy story were composed of yellow cypress dominantly, subcanopy were hardwoods in the site. For exploring the adaptation strategy of these 2 cypress species at sapling stage under diversity of canopy condition, experiment was designed with 2yrth planting seedlings of yellow and red cypress as sapling stage under 3 canopy closure degree (weak, medium, strong, using spherical densitometer) plot site. We measured and compared mortality and growth performance of 2 cypress species stock in 9 years after planted.
Results show mortality rate of red cypress higher than yellow cypress under strong canopy closure at initial plantation stage, but lower under weak canopy, that expressed yellow cypress have more shade tolerant than red cypress. After 4 years, the mortality of the 2 cypress species were higher under weak closure, also, found much seedlings destroyed by rodents under open canopy condition.
The growth performance (height, diameter) of red cypress were better than yellow cypress under all canopy conditions during observation time. Annual growth of both cypress seedlings were litter under closure than open canopy, and growth difference of yellow cypress were significant than red cypress among canopy conditions. Growth rate of seedlings under open canopy were higher at initial time, and red cypress was better than yellow cypress. But growth rate declined as seedling develop, and no significant difference among species and canopy conditions. We also found the tendency that yellow cypress grew well under closure canopy after 8yrth. Seedling sturdiness quotient of red cypress were higher than yellow cypress, especially under closure canopy, that show light requirement of red cypress much than yellow cypress.
From mortality and growth expression at sapling stage under different canopy condition, We supported the hypothesis of adaptation strategy that red cypress was shade avoidance type and yellow cypress was shade tolerance type.
Key words : Cypress forest, Chamaecyparis obtusa var. formosana, Chamaecyparis
formosensis, forest canopy heterogeneity, shade tolerance, adaptability.
謝誌
本論文承蒙恩師 林世宗教授悉心教誨與殷切督促,研究方向之指 引、觀念之啟迪及生活上的關切,文稿草成,復多次逐字匡正,增添潤飾,
全文始克完成,師恩浩瀚,謹誌於卷首,以表最崇高之謝忱。並感謝郭幸 榮教授、關秉宗教授、邱祈榮教授、陳子英教授細心審閱論文初稿,給予 寶貴意見及斧正指導,使本論文更臻完善,謹致最誠摯之謝意。
感謝森林保育處,給予公假前往進修,王處長成明、李副總技師炎壽 暨各位長官多方鼓勵與提攜,棲蘭山工作區同事們在工作上的體諒及擔 待,而能公餘戮力撰寫論文,始能完成學業,銘感於心。
調查期間感謝棲蘭山工作區同事馮漢章、黃坤生、吳春榮、楊建國、
邱明輝、王銀地等,同窗偉欣、享勳、瑞廷等於野外調查的大力協助;學 妹思佳在林冠量測時不辭辛苦的相助;智昕、亞臻在統計上援助;台大潘 孝隆、王瑞明等在航照圖及野外定點;土壤樣本分析工作承宜大土壤研究 室蔡呈奇老師及瑀芳同學的大力襄助;雅惠在簡報製作時費心幫忙;研究 室助理陳小玲小姐行政上的支援,在此致上由衷之感謝。由於前述師長及 同仁之鼎力協助,本論文方能順利完成,僅此一併致謝。
這篇論文能夠順利完成,實在得助於太多太多人有形無形的幫助與鼓 勵,文成倉促,疏漏謬誤在所難免,尚祈林業先進不吝指正。謹以此論文 獻給我最親愛的爸媽兄姐弟及內人、岳母,以及所有關心我、支持我的諸 位們。
林進龍 謹誌於 育林研究室 2007.07.16
目 錄
中文摘要………i
Abstract………ii
謝誌………iii
目錄 ………....iv
圖目錄 ……….vii
表目錄 ……….viii
壹、前言 ……….….………1
貳、前人研究 ………..……4
一、冠層結構量化………..……4
二、干擾分類與特性 ………...………….….……..…5
三、干擾後森林之發育 ………..…...…6
四、干擾後苗木適應 ……….…..………..10
參 、 材 料 與 方 法 … … … …… . … … … … 1 3 ㄧ、研究區域概述……….…13
(一)研究區位置……….……..13
(二)氣候概況……….………..14
(三)地質與土壤……….………..15
二、研究方法……….…..…..…..…16
(一)研究材料……….…..…..…..…16
(二)苗木量測……….…..…..…..…16
(三)冠層環境量測……….…..…..…..…16
(四)試驗區林分結構分析……….…..…..…..…20
三、資料分析……….….…...…….20
肆、結果………..………….…….………….21
一、試驗區上層林冠結構分析………..………….…….………….21
(一)冠層水平結構異質性變動………..………….…….………...21
(二)試驗林分組成結構分析………..………….…….………...21
(三)試驗區內樣區上層林冠鬱閉度分析……….…….………...22
二、不同冠層環境下檜木苗死亡率之比較…..………….…….………….28
(一)不同鬱閉度下檜木苗之死亡率…..………….…….………..28
(二) 不同鬱閉度下檜木苗受囓齒類動物危害之比率….………...31
三、不同冠層環境下檜木苗高生長之比較…..………….…….………….31
(一)不同冠層環境下檜木苗之苗高…..………….…….………...31
(二)不同冠層環境下檜木栽植苗之苗高連年生長量….………..33
四、不同冠層環境下檜木苗徑生長之比較…..………….…….………….36
(一)不同冠層環境下檜木苗之苗徑…..………….…….………...36
(二)不同冠層環境下檜木苗之苗徑連年生長量….…….………...37
五、不同冠層環境下苗木生長率之比較……….…….………..39
(一)不同冠層環境下檜木苗之苗高生長率……….…….………….40
(二) 不同冠層環境下檜木苗之苗徑生長率………….…….…………...41
六、不同冠層環境下檜木苗形質之表現……….43
(一)不同冠層環境下檜木苗之健壯商數………...43
(二)不同冠層環境下檜木苗之樹冠形態………...46
七、不同林冠鬱閉下檜木苗生長參數之相關性……….…….…………..48
伍、討論………..………….…….………..53
一、檜木林經整理干擾後林冠孔隙結構之變動…….…….………..53
二、不同林冠鬱閉度下檜木苗死亡率……..………….…….………..53
三、不同林冠鬱閉度與檜木苗生長表現…..………….…….………..55
(一)苗高及苗徑…..………….…….………..55
(二)連年生長量…..………….…….………...55
(三)各苗齡生長率之變化….…….………...56
四、不同林冠鬱閉度下檜木苗形質表現…..………….…….………..57
五、紅檜與台灣扁柏更新生長比較………..………….…….………..58
陸、結論………..………….…….………..61
柒、參考文獻………...…...63
捌、附錄………..………….…….………..70
圖目錄
圖1左為冠層覆蓋量測方法,右為冠層鬱閉量測方法。….……….….…..4 圖2研究樣區位置圖……….……….……..…….14 圖3半球面鬱閉度計(spherical densiometer)量測位置...……..……….17 圖4試驗區不同時期上層林冠孔隙變動情形………….……….……..…….24 圖5試驗區上木胸徑分佈圖……….……….……..……….25 圖6試驗區上木樹高分佈圖……….……….……..……….26 圖7林分視覺景觀系統(SVS)試驗區上層林木結構示意圖.……..……...27 圖8不同林冠鬱閉度下各苗齡檜木苗之死亡率…..….……….……....…….30 圖9不同林冠鬱閉度下檜木苗受囓齒類損害情形….……….……..……...31 圖10不同林冠鬱閉度下各苗齡檜木苗之苗高..…….……….……..……33 圖11不同林冠鬱閉度下各苗齡檜木苗之苗高年生長量.……….……..……35 圖12不同林冠鬱閉度下各苗齡檜木苗之苗徑…….……….……...……….37 圖 1 3 不 同 林 冠 鬱 閉 度 下 檜 木 苗 之 苗 徑 連 年 生 長 量 … . … … . . . … … . 3 9 圖14不同林冠鬱閉度下各苗齡檜木苗之苗高生長率…….……...………41 圖15不同林冠鬱閉度下各苗齡檜木苗之苗徑生長率….………42 圖16不同林冠鬱閉度下各苗齡檜木苗之健壯商數……….……..………45 圖17不同林冠鬱閉度下栽植9年生檜木苗之樹冠形態….……….47
表目錄
表 1 試驗區不同時期孔隙及林冠變動情形………...………...23
表 2 試驗區林分上木組成結構………...………...25
表 3 樣區鬱閉度分級………...………...28
表 4 不同林冠鬱閉度下各苗齡檜木苗之死亡率(%)變異分析表………29
表 5 不同林冠鬱閉度下各苗齡檜木苗之苗高(㎝)變異分析表………..32
表 6 不同林冠鬱閉度下檜木苗之苗高連年生長量(㎝ yr-1)變異分析表…..34
表 7 不同林冠鬱閉度下各苗齡檜木苗之苗徑(㎝)變異分析表………..36
表 8 不同林冠鬱閉度下檜木苗之苗徑連年生長量(㎝ yr-1)變異分析表…..38
表 9 不同林冠鬱閉度下各苗齡檜木苗之苗高生長率變異分析表…………..40
表 10 不同鬱閉度下各苗齡檜木苗之苗徑生長率變異分析表………42
表 11 不同林冠鬱閉度下各苗齡檜木苗之健壯商數變異分析表………44
表 12 不同林冠鬱閉度下栽植 9 年生檜木苗之樹冠形態變異分析表……….46
表 13 栽植 9 年後台灣扁柏苗木各生長性狀之相關矩陣……….51
表 14 栽植 9 年後紅檜苗木各生長性狀之相關矩陣……….52
表 15 紅檜與台灣扁柏更新生長比較………59
附表 1 1992-2006 侵襲本研究區颱風資料………70
附表 2 不同林冠鬱閉度下各苗齡檜木苗之死亡率比較………..71
附表 3 不同林冠鬱閉度下各苗齡檜木苗之苗高比較………..72
附表 4 不同林冠鬱閉度下檜木苗之苗高連年生長量比較………..73
附表 5 不同林冠鬱閉度下各苗齡檜木苗之苗徑比較………..74
附表 6 不同林冠鬱閉度下檜木苗之苗徑連年生長量比較………..75
附表 7 不同林冠鬱閉度下各苗齡檜木苗之苗高生長率比較………..76
附表 8 不同林冠鬱閉度下各苗齡檜木苗之苗徑生長率比較………..77
附表 9 不同林冠鬱閉度下各苗齡檜木苗之健壯商數比較………..78
附表 10 不同林冠鬱閉度下栽植 9 年生檜木苗之樹冠形態比較………79
壹、前言
扁 柏 屬 (Chamaecyparis) 的 台 灣 扁 柏 (Chamaecyparis obtusa var.
formosana) 及紅檜 (Chamaecyparis formosensis) 兩為台灣之重要針葉樹樹 種,因材質佳、形質優美、天然木高大通直、直徑大、材段長且有耐腐朽 及抗蟲等優點,是林業重要的上等木材,具有很高的經濟價值,也是造林 的重要樹種(郭寶章,1995),主要分布在台灣中高海拔山區之雲霧林帶。
在分布的海拔高度上,紅檜略低於台灣扁柏,但兩者同時存在的區域,
台灣扁柏略高 200-500 m,唯皆分布於盛行雲霧帶(章樂民,1963;蘇鴻傑,
1984)。有學者提出台灣扁柏在低溫環境下有較強的淨光合速率,所以分布 海拔較紅檜為高(梁亞忠,1990;賴宜鈴,2006)。溫帶雨林在台灣的海拔分 布呈現由南往北遞降的趨勢,其植物組成在海拔梯度及南北分布上有差異 ( 蘇鴻傑, 1984) 。南部的大武山自然保留區,檜木林的分布約在海拔 1,700-2,500 m 之間,以紅檜為主(王鑫等,1988; 1989)。中部的沙里仙溪流 域,檜木林的海拔分布在 2,200-2,500 m 之間(鍾年鈞,1994),以紅檜為主,
常伴隨有台灣扁柏、台灣雲杉(Picea morrisonicola)、台灣粗榧(Cephalotaxus wilsoniana)及台灣鐵杉(Tsuga chinensis)等裸子植物。北部的檜木林,以南湖 大山為例,分布在海拔 1,800-2,300 m 之間,而太平山區的檜木林則分在海 拔 1,300-2,600 m 之間(章樂民,1963;劉棠瑞、應紹舜,1973;蘇鴻傑,1984)。
柳榗(1975)對台灣東西部的檜木天然林進行調查,分析檜木分布的環境 因子。紅檜分布在東北向、東向、南向及西南向之間,而台灣扁柏則在西 南向、西向及北向之間。另就分布坡面的位置而言,台灣扁柏多分布在坡 面之中、上部及山脊,而紅檜則分布在坡面的中、下部位(陳子英等,2002;
魏瑞廷,2006)。柳榗(1975)則認為紅檜好陰濕,台灣扁柏耐乾旱,兩樹種 皆耐瘠薄。根據第三次森林資源調查顯示目前現存的天然檜木林,北限大 致在插天山一帶,南限則是在阿里山、玉山、秀姑巒山一帶,主要分布在
北部及東部,其中以棲蘭山檜木林的面積最為集中。
當森林中的紅檜與台灣扁柏的優勢度超過 50%以上時,可稱為紅檜或 台灣扁柏的純林,此種森林中,樹冠較為稀疏的老林內常見有檜木的幼樹 發生(柳榗,1971)。但紅檜與台灣扁柏二者天然更新能力之差異,在成熟之 台灣扁柏林中之幼樹多見於樹冠鬱閉破裂之下方(柳榗,1975),詹琬婷(2004) 由樹輪分析判釋台灣扁柏可在鬱閉林分樹冠下生長,並經由小尺度干擾進 行更新,說明台灣扁柏具孔隙更新能力而為孔隙種。至於紅檜林之幼樹,
大部皆出現在演替初期的空曠地,其餘在老齡林中所見之少數幼樹皆見之 於大形樹冠鬱閉破裂孔隙之正下方或在外圍 1 m 以內之地區,認為紅檜為 較不耐蔭之樹種(柳榗,1975;賴宜鈴,2006)。林旭宏等(1999)調查瑞岩溪 紅檜林型組成及其徑級分布,研判其兼具孔隙種與先驅種之特性,即其幼 苗可於成熟林下以前生苗(advance growth)的形式存在,以等待孔隙形成後 晉升至冠層,而在新形成的裸露地中也可更新良好,可與其它先驅植物共 同競爭;但在過於老熟之紅檜林分中,由於上層鬱閉破裂導致林床之地被 植物過度繁盛,將會影響包括紅檜等木本植物的更新。
傳統林業經營以永續林木生產為目的,而近年來美國提倡的新林業 (New Forestry)、生態林業(Ecological Forestry)、或生態系經營(Ecosystem Management),則強調永續性森林生態系經營的革新理念。生態系經營就是 用生態方法,融合了人民的需求與環境的價值,以達成森林的多目標利用 經營,使得森林呈現出多樣的、健康的、生產的以及永續的生態系。應用 森林生態系經營理念,以適應性經營模式保育檜木天然資源,即依檜木生 態系統組成維持檜木林結構之完整性,同時亦維護森林生態系之功能。
森林經營者在策劃結合生態學和經濟學目標的育林技術計畫時,需要 瞭解林分自然發育過程的綜合科學,包括干擾控制和生物〝遺骸〞(biological legacies)的評估,例如留存的立木、枯立木以及倒木(Franklina et. al., 2002)。
而森林的干擾可分為天然干擾(natural disturbance),如火災、暴風、洪水、
地滑、地震、山崩、乾旱、動物及病蟲危害等;與人為干擾(anthropogenic disturbance),如伐採林木、放牧、移除林內枯枝落葉、採石開礦、採集果 實及藥材、旅遊及工業污染等(李洪選等,2004;徐化成,2004)。
棲蘭山林區檜木天然林中物種組成多樣,其中以紅檜及台灣扁柏兩種 富有高經濟價值。光復初期為振興經濟,傳統林業經營為達提高收益、降 低成本,多採大面積皆伐方式作業,後為降低環境及生態上的衝擊,作業 方式調整為小面積皆伐,並曾進行單株擇伐及枯立倒木移除之更新試驗。
據林業試驗所洪良斌(1975)調查風倒、枯死及不健全之缺頂或腔腐林木蓄積 量約 39%,林冠多呈破裂,林下雜草、灌木及箭竹叢生,下層幾無檜木幼 林存在,加以檜木根盤甚淺,且該地區灰化土層堅硬,極易發生枯死及風 倒現象,為妥善利用森林資源,森林保育處辦理枯立倒木整理保育計畫,
於整理後採用自生苗保育、天然下種及人工造林三種方法建造複層林。皆 伐作業、單株擇伐、枯立倒木整理等作業皆為檜木天然林人為干擾的方式,
其干擾強度、大小及影響皆有不同。
枯立倒木整理作業對後續檜木森林資源經營,不僅考量木材可供利用 亦可更新形成檜木林,促進森林資源的循環性以維持永續性,同時亦可運 用不同的更新作業方式,形成不同的檜木森林類型,增加檜木森林的多樣 性。在禁採天然林後,森林保育處就棲蘭山林區部份老熟化的檜木林為促 進其更新,實施枯倒木移除以釋放林地之空間,提供檜木的下種與發育環 境。本研究即以整理作業後之檜木林所形成之林冠結構下,以均質之紅檜 與台灣扁柏栽植苗設定為檜木林之幼苗期,比較檜木幼苗在建造期之生長 適應能力的表現,提供檜木天然林永續經營之參考。
貳、前人研究
一、冠層結構量化
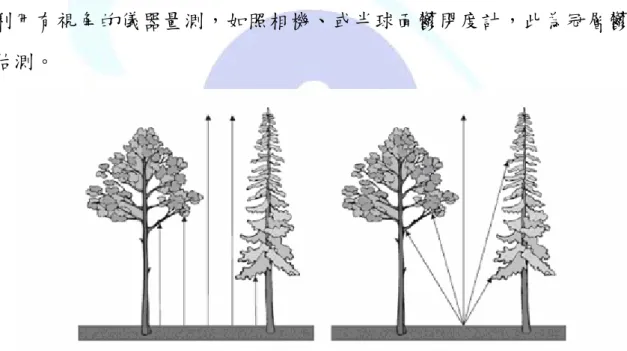
森林冠層覆蓋(forest canopy cover),亦稱為冠層覆蓋度(canopy coverage) 或樹冠覆蓋(crown cover)。冠層覆蓋可提供為多目標之生態指標,例如判釋 植群活力和動物棲息地、森林地表微環境之變化,以及由冠層葉面積指數 (leaf area indes, LAI)及光合能力評估森林生產力(Jennings et al., 1999, 林登 秋和江智民,2002)。有關冠層覆蓋的量測與表現有各種方法,有各種詮釋,
首需認識及區隔冠層覆蓋(Canopy cover)與冠層鬱閉(canopy closure)之概 念。其中冠層覆蓋之定義為林木樹冠垂直投影於森林地表之比率;而冠層 鬱閉之定義為單一點上空被植被遮蓋的比率(圖1)(Korhonen et al., 2006)。如 果利用有視角的儀器量測,如照相機、或半球面鬱閉度計,此為冠層鬱閉 度估測。
圖1左為冠層覆蓋量測方法,右為冠層鬱閉量測方法。(摘自Korhonen et al., 2006)
Fig. 1 Canopy coverage (left) is always measured in vertical direction, whereas canopy closure (right) involves an angle of view.
林登秋和江智民(2002)曾論述以半球面影像技術在森林生態研究之應 用,可供估測穿透林冠的光照及林冠葉面積指數。在生態研究方面,冠層 鬱閉可應用在林下光照(賴宜鈴,2006)、林冠結構變動、林下植物的生長分 布、森林的演替更新等之評估,本方法雖具有移動迅速節省時間及經費優 點,然研究者需對影像特質及分析的原理與流程有充分的了解,才能在事 前影像取樣點的選擇、影像拍攝時間的選定、影像的分析乃至於資料的統 計分析時避免錯誤的發生與誤差之累積。王雪卿和林登秋(2006)以傳統相機 及數位相機搭配魚眼鏡頭,探討相機類型、影像解析度、影像壓縮及不同 分析軟體對估測林冠光穿透之影響,結果顯示影像壓縮與否對分析之結果 並無顯著影響,而解析度則有顯著影響,解析度愈高則估算之林下光照越 高。使用傳統相機及數位相機所推估之林冠光穿透可相差超過 5%,不同影 像分析計算軟體所造成之差異亦可達 5%以上,因此使用時需注意解析度、
相機系統以及分析軟體是否相同。
Korhonen et al. (2006)比較幾種量測冠層覆蓋方法之精確度,利用 Cajanus tube、數位照相或半球面鬱閉度計(spherical densiometer)、目測法 (ocular estimation)。其結果以 Cajanus tube 配合穿越線取樣法(line intersect sampling,LIS)最為精確但較費時;以半球面鬱閉度計量測鬱閉度,其平均 值與 Cajanus tube 法差異不大,但量測較快而節省時間;而數位照片或目測 法則視研究者之經驗,且因主觀因素所得結果差異甚大。
二、干擾分類與特性
就干擾之作用方式,可分為外在干擾(exogenous disturbance)和內在干擾 (endogenous disturbance)。如火災干擾為外在的,而小規模的林木倒伏干擾 則是內在的。同一種干擾因素,既可能是較大的干擾,也可能是較小的干 擾 , 既 可 以 是 外 在 干 擾 , 也 可 以 是 內 在 干 擾 。 Sousa(1984) 指 出 干 擾 (disturbance)為「一種獨立事件,它會造成生物體的死亡、取代或傷害,進
而直接或間接的製造了其他生物建立之機會」。Oliver and Larson (1990)以干 擾的頻度(frequency)、林木移除的數量(amount of removed forest)和受到干擾 範圍大小(size and shape of the area disturbed)來描述干擾的特徵。干擾對森林 樹冠層疏開之程度,可分為重大干擾(major disturbance),為林冠層之林木完 全 被 破 壞 後 , 整 個 林 分 毀 滅 而 全 面 更 新 , 或 稱 為 林 分 替 代 性 干 擾 (stand-replacing disturbance);輕度的干擾(minor disturbance),為林冠層之林 木只受到部分破壞的干擾,或稱為林木替代性干擾(tree-replacing disturbance) (Oliver and Larson 1990)。規模較小的干擾如林木的老朽枯死,造成樹冠疏 開(canopy opening),即一般所稱之樹冠孔隙,或以孔隙(gap)稱之(Bray, 1956)。
森林孔隙的形成對該森林的組成、結構及功能具有重要的影響(Brokaw et al., 1989),促使非耐蔭性的先驅樹種及孔隙種(gap species)的天然更新,
形 成 森 林 內 具 有 族 群 動 態 、 樹 種 組 成 及 生 長 率 等 特 性 都 不 同 的 區 集 (patch)(Brokaw, 1985)。孔隙產生以後對於苗木的生長都有幫助,然樹種不 同最適合的生長區集也有差異,使孔隙的大小與區集對於苗木的生長產生 篩選的作用(Duncan et al., 2002;林謙佑等,2004)。森林冠層上層林木死亡 後產生孔隙,這些冠層孔隙對森林有正面影響,包括增加物種多樣性。相 對的孔隙內下層植物的生長和在孔隙邊緣之上層林木枝條的擴張造成冠層 孔隙鬱閉,將決定未來森林之結構和物種的組成(Brian et al., 2004)。
三、干擾後森林之發育
干擾可影響到森林各個層次的結構動態,從個體角度,干擾效應為造 成全部或部份個體的傷害或死亡。從種群來說,干擾影響種群發生過程、
年齡結構、垂直結構、遺傳結構及單株的空間分布。干擾所造成環境上的 異質性,其影響是表現於二個不同性質的梯度上,即是時間和空間上的異 質性。時間梯度即造成所謂的區集動態(patch dynamic),空間上之表現則是
區集的鑲嵌分布(mosaic of patches)。
以地景生態學而言,干擾造成的孔隙更新會使林分在水平空間上形成 鑲嵌狀的區集。因林木死亡或消失而釋出可供生存及生長的空間與資源,
苗木進駐孔隙形成集落,或是由孔隙邊緣的上層優勢木以側枝生長,逐漸 填補破碎的樹冠層,隨著時間的推進,這些孔隙漸漸形成了異質鑲嵌體 (mosaic)而迥異於原林分,改變了原來的地景結構。透過孔隙在不同區域不 同時間的發生,整個地景持續地進行小區集的更新。經過長時間多次干擾、
更新作用下,在最後演替階段,林分結構在水平與垂直兩個層次上發生了 多次的分化,因此林分結構的異質性(heterogeneity)為老林林分結構的重要 特微。
森林內林木的死亡造成樹冠疏開,孔隙將由其他林木生長、填補而逐 漸閉合。Oliver and Larson(1990)將孔隙內族群的更新動態,區分為四個時 期;(一)孔隙期(gap phase):侵入孔隙的幼苗數逐漸增加;(二)建造期(building phase):幼樹密度逐漸達到飽和,且產生激烈的競爭和自然疏伐;(三)成熟 期 (mature phase):更新林木已達樹冠層,競爭趨於緩和;(四)退化期 (regressive phase):冠層林木漸漸老杇,或因外力作用而形成倒木,產生新 的孔隙。上述四個時期週而復始,致使在林中形成小的區集。同時期的區 集在林分外貌上則呈現鑲嵌組合(mosaic of patchs)(劉棠瑞、蘇鴻傑,1983;
Franklina et al., 2002)。
干擾後形成孔隙後最直接、最重要的效應是提高孔隙內的光照條件及 水分變動(Duncan et al., 2002;林謙佑,2004;宋新章等,2006)。因上層林 木死亡所產生的冠層孔隙,可能增加林下之光度、土壤水分(Denslow et a1., 1990,林謙佑,2004)及養分,而這些因子全都可能限制幼苗生長和存活。
在森林上層及下層之間微環境的異質性的作用,會影響幼苗出現及後來的 群落組成和物種共存的潛能(Beckage et al., 2000;Brian et al., 2004)。孔隙下
層之幼苗對環境競爭與自然資源限制間的交互作用,影響幼苗生長與存 活。例如,在冠層孔隙下光度增加,下層草本、蕨類、灌木可能隨之增加,
而與林木幼苗競爭。相反地,冠層孔隙高光下微環境的特徵為礦質養分偏 高或土壤水分不均勻,影響幼苗出現。
林謙佑(2004)研究觀測南投縣和社神木溪天然闊葉樹保護林中,當冠層 出現孔隙後,其內微環境因子的動態變化及對苗木生長的影響。以兩個不 同大小的冠層孔隙,栽植 7 種闊葉樹之ㄧ年生苗木,監測光量與土壤水勢 的變化,觀察其適應情形。結果顯示冠層孔隙形成以後,將使林地相對光 度增加,且於每年 3-9 月生長季節最為明顯,顯示孔隙的環境條件對苗木生 長有所幫助,然樹種不同,其最適合的生長區集也有差異,使孔隙的大小 與區集對於樹種苗木的生長產生篩選的作用,認為光度為決定孔隙內苗木 生長的主要因子而非土壤濕度。
Beckage et al. (2003)研究孔隙與冠層下空間之異質性(以不同處理移除 下層植被、挖溝,移除下層植被及挖溝、施肥,以及對照組)對三樹種之存 活及生長所扮演的角色,發現紅楓(Acer rubrum)、北美鵝掌楸(Liriodendron tulipifera)、北美紅櫟(Quercus rubra)其幼苗生長與存活率有明顯差異,在所 有的微生育地環境中北美紅櫟之幼苗存活率皆較紅楓、北美鵝掌楸兩種 高,但在三個較高資源環境中北美鵝掌楸有較高的生長率。發現在空間異 質性內之微環境,3 個樹種中僅有 2 樹種(北美鵝掌楸及北美紅櫟)能夠共 存,表示較高環境資源能夠促進更新棲地潛能。
種子發芽及幼苗出現之成功與否,將決定植物的分布及植群的結構,
然種子是否得以發芽成為幼苗,常受到許多外在條件所影響,如光、溫度、
水分及氧氣等均為種子發芽所必需(王子定,1978)。對檜木種子發芽的研 究,如溫度有最高溫、最低溫及適溫與變溫,光照有強度與光期(胡大維等,
1978),發現紅檜或台灣扁柏種子發芽率受溫度影響,但光度對檜木的發芽
沒有影響;而檜木種子發芽受水分影響之程度以台灣扁柏較為劇烈,紅檜 較為緩和,亦即紅檜可適應的水分範圍較台灣扁柏寬廣,天然更新之能力 較佳(林湘玲,2001)。因此檜木種子能否成功發芽,主要取決於溫度及水分,
只要有適當的溫度、濕潤的環境檜木種子即能容易發芽。
檜木是以下種方式進行天然更新,因此檜木天然更新能力,視(一)種子 材料供應,包括種子量與下種方式。(二)種子發芽效性,如種子品質、下種 環境對種子活力的影響,及發芽環境。(三)苗木生長適應能力。棲蘭山檜木 之種子成熟期紅檜為11-12月,台灣扁柏為10-11月,下種期紅檜為12月初至 翌年6月底,台灣扁柏為11月初至翌年3月中旬,其中以12月至翌年5月為主 要下種時間。張乃航等(2001)調查三個不同時期枯立倒木整理區之檜木 下種量最大為20 million 粒/ha,最少為6.5 million 粒/ha。林世宗(2005,未 發表)亦調查年下種量可達19.4 milllion 粒/ha。檜木種子發芽率及發芽苗成 活率,於下種前整理之林分較高,因為有較低的地表覆蓋,且經由中耕培 土擾動土表有利於種子發芽及存活。Zobel(1980) 在美國奧勒崗州西南四個 天然林分進行森林地表擾動處理包括地表末擾動處理、移除枯枝落葉層、
焚燒枯枝落葉層、翻土擾動4種處理,對羅生扁柏(Chamaecyparis lawsoniana) 播種後種子發芽及存活之效應,以擾動土壤之種子發芽率及存活率最高。
四、干擾後苗木適應
森林冠層與太陽輻射量之關係,對於植群生理生態、野生動物及森林 演替等之研究,具有重要價值(梁亞忠等,1999)。光照也是限制森林林下植 物生長與分布的重要因子,因此了解林下光照環境的時空變異,有助於深 入探究林下植物的生長分布及生產力。由於林下小苗的生長關係著未來林 分的發展及冠層樹種的組成,林下光照因其對樹苗生長的影響也關係著森 林的更新,所以定量林下光照環境為研究森林動態的重要ㄧ環(林登秋和江 智民,2002)。
在冠層干擾之後,有許多的因子影響幼苗的建立。鄰近母樹與母樹數 量(郭婉君,2001;賴宜鈴,2006)、苗床基質(廖啟政,2003)、種子消粍者 出現與傳播(蔡若詩,2000;Manuel et al., 2002)、與氣候和微氣候,全都 導致幼苗淨殖在時空上的變化。
在不同量的光資源環境下,下層林木可藉修改其樹冠形態及葉的解剖 性狀,以利其有最佳的存活機會,而達到上層冠層位置。一般光限制環境 下,多數耐蔭性樹種較非耐蔭性樹種有較高的存活率(Oliver and Larson, 1990; 賴宜鈴,2006)。Robin et al. (2001)研究林下光度(鬱閉冠層、小孔隙、
大孔隙)、幼樹大小(幼木50-100 ㎝及101-200 ㎝)和種內競爭對冷杉幼樹生 長與樹冠形態之重要性,發現林下光度增加則冷杉幼樹之相對苗高及苗徑 生長率增加。林下光度增加達25%或以上時,種內競爭對幼樹生長影響才 具重要性,競爭指數愈大,幼樹相對苗高及苗徑生長率減低。
Coates (2000)進行北方溫帶林孔隙對美西側柏(Thuja plicata)、西部鐵杉 (Tsuga heterophylla)、亞高山冷杉(Abies lasiocarpa)、白雲杉(Picea glauca)、
柱松(Pinus contorta)等五種針葉樹幼苗之影響研究。結果顯示所有樹種在孔 隙大小增加時表現出漸進性生長。所有樹種在孔隙從單一林木孔隙至 1,000-2,000 ㎡時其生長增加最快,但孔隙持續增大之後,對平均生長率之
促進幫助不大。當孔隙大小在小型(20-300 ㎡)與中型(301-1,000 ㎡)時各樹種 之生長表現最顯著。在森林下層各樹種間除了死亡率差異大之外,其生長 率亦低。在中孔隙最陰暗地區(孔隙南緣)、小孔隙樹種間存活亦有差異,表 示孔隙內位置及孔隙大小對苗木存活率及生長表現有顯著影響。
Hoshino et al. (2001, 2002, 2003) 進行日本中部赤澤森林保護區日本扁 柏老齡林冠層狀態下 6 種主要針葉和落葉闊葉林樹種動態學之研究。在 10 年(1988-1998)研究期間,發現日本扁柏(Chamaecyparis obtusa) 和日本花柏 (Chamaecyparis pisifera)可以在鬱閉落葉闊葉樹冠層下更新,日本羅漢柏 (Thujopsis dolabrata)在鄰近冠層孔隙下更新,麻櫟(Quercus mongolica)、厚 朴(Magnolia obovata)和日本櫻樺(Betula grossa)在冠層孔隙下更新。其中針 葉樹淨增殖率低於死亡率,而落葉闊葉種則淨增殖率大於死亡率,針葉樹 淨增殖率明顯低於落葉闊葉樹種。所有主要樹種林木死亡率取決於更新 苗,苗木愈小死亡率愈高,所有樹種幼苗總增殖率皆集中在孔隙環境下。
國內有關苗木之生長適應研究,多於苗圃以人工方式營造不同光度環 境下,觀察苗木之生長型態及生理生態特性之反應,探討各樹種生長發育 的最佳條件(王相華,1995;林科言,2001;沈介文等,2004;林怡芳,2005)。
或將苗木栽植於不同光度環境下,調查新植苗木之存活、光合作用產物分 配比例之調整情形(郭耀綸等,2003;劉興旺,2001)。林渭訪等(1958)、方 榮坤等(1991)觀察苗木於不同光度下之生長情形,結果顯示台灣扁柏及紅檜 之苗高及苗徑生長與光度具正相關。洪良斌等(1979)於林下栽植之紅檜幼林 分中選設 30%、50%、70%、90%四種不同光度林地,進行紅檜幼林需光度 之研究;結果顯示,表示各光度區之生長量差異甚為顯著,年份間生長量 之差異則不顯著。該幼林分直徑與樹高之生長量,均示以 90%之光度區為 最優,70%及 50%同為次之,而以 30%之光度區之生長量最差。
陳岫女(1990)研究發現施用氮磷鉀肥對紅檜苗木生長較佳,而僅施磷、
鉀或磷鉀者形態生長較差,但對台灣扁柏的影響較小。方榮坤等(1990)指出 未施肥紅檜苗木之活力及根系生長潛能較差,造林成活率及生長較劣。陳 秋萍(2003)發現紅檜與台灣扁柏的生長及生物量主要受氮肥影響。王儷儒 (2004)研究紅檜及台灣扁柏苗高及苗徑生長與施用氮磷鉀成正相關;台灣扁 柏苗木在低水勢環境下,其光合作用能力潛勢高於紅檜苗木;於水分充分 供應時紅檜的光合作用能力與台灣扁柏苗木無顯著差異,但苗高明顯較台 灣扁柏高,顯示在水分充足時紅檜幼苗的競爭能力比台灣扁柏苗木高。
賴宜鈴(2006)研究棲蘭山區林下光環境對兩種檜木小苗生長與建立之 影響,以棲蘭山區之氣候有非常潮濕和光量低的特徵,且具明顯的季節性 變化,調查發現台灣扁柏小苗適存光量較紅檜小苗低。在較高光量環境下,
紅檜小苗比台灣扁柏小苗具有較高之存活率、光合作用率、葉相對生長率,
而使生物量明顯比台灣扁柏小苗高,因此比台灣扁柏具有競爭力。但在持 續高光下,兩種檜木小苗的光合作用效率和葉相對生長率都被明顯的抑 制,顯示紅檜和台灣扁柏小苗都非極端陽性樹種。
前述研究多僅限於苗圃育苗或苗木出栽新植階段生長表現,對於苗木 栽植後,隨時間改變其面對種內與種間環境資源及不同干擾程度之情況 下,林木生長狀況的持續監測仍待更多的論證與研究。
參、材料與方法
一、研究區域概述 (一)研究區位置
棲蘭山林區位於雪山山脈之北稜,海拔最高者達 3,000 m 以上,以海 拔 1,000-1,500 m 所佔比例最大,約佔 36﹪,其次為海拔 1,500-2,600 m,
約佔 27﹪。以桃山山脈至喀拉業山(海拔 3,132 m)向北遞減,依序為馬惱 山、東保津寒山、唐穗山、馬望來山、至棲蘭山為稜脊,稜線以東屬蘭 陽溪集水區,稜線以西屬淡水河集水區。
在棲蘭山林區檜木分布依地形、坡度及微環境因子之影響而有所差 異,紅檜大多自海拔 800 m 即有分布,可至 2,800 m,以 1,200-2,200 m 生長較佳;台灣扁柏大多自海拔 1,200 m 才開始分布,常呈純林狀態,
以 1,500-2,600 m 生長較佳。就地形而言,稜線為台灣扁柏、台灣二葉松 (Pinus taiwanensis)及台灣鐵杉等針葉樹為主,形成山地上層針葉林型;
中 坡 及 上 坡 為 台 灣 扁 柏 、 台 灣 鐵 杉 、 台 灣 杜 鵑 (Rhododendron formosanum)、森氏櫟(Cyclobalanopsis morrii)、毽子櫟(Cyclobalanopsis sessilifolia) 、 高 山 新 木 薑 子 (Neolitsea acuminatissima) 、 木 荷 (Schima superba)、白花八角(Illicium anisatum)、霧社木薑子(Litsea elongata var.
mushaensis)等混生,為櫟林帶上層山地針闊葉混洧林型之台灣扁柏型;
溪 谷 為 紅 檜 、 墨 點 櫻 桃 (Prunus phaeosticta) 、 假 長 葉 楠 (Machilus japonica)、錐果櫟(Cyclobalanopsis longinux)等混生,為櫟林帶上層山地 針闊葉混淆林型之紅檜林型(郭寶章,1995;陳子英等,2002;王震哲,
2000;朱恩良,2004;魏瑞廷,2006)。
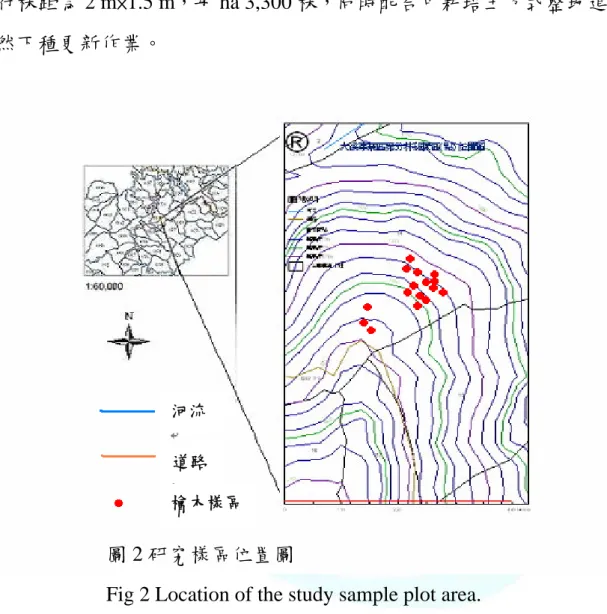
試驗地位於森林保育處轄管棲蘭山林區 130 線林道(圖 2),海拔範圍 約為 1,500-1,650 m。坡度約 20-40∘,坡向為東南—東北,其前身為檜木 天然林,森林保育處於 1996-1997 年實施枯立倒木移除作業,面積約 30
餘 ha,1998 年 2-3 月實施復育更新作業,將基徑 10 ㎝以下之雜木清除,
依水平橫坡帶狀方式整地,於林下人工栽植檜木苗(2 年生裸根苗),栽植 行株距為 2 m×1.5 m,每 ha 3,300 株,同時配合中耕培土方式整地進行天 然下種更新作業。
圖 2 研究樣區位置圖
Fig 2 Location of the study sample plot area.
於 1998 年實施檜木保育更新作業,整地後未造林前,在面積約 2.4 ha 內以目測法判釋冠層環境,分密、中、疏、空曠四種林冠環境之樣區,
栽植紅檜及台灣扁柏 2 年生裸根苗,以羅盤儀劃設 16 個 20m×20m 的樣 區,各重複 2 個樣區,苗木均編號。
(二)氣候概況
棲蘭山林區地勢西南高東北低,山脈多成東北走向,東北季風常得 以順谷長驅直入,故氣候濕潤。本區氣候 7、8 月氣溫最高平均 27 ℃左
檜木樣區 道路 河流
右,最低 1 至 2 月平均在 13 ℃左右,年雨量在 1,700-2,500 ㎜,全年各 月份的相對濕度在 90﹪以上,屬重濕型氣候,常年雲霧瀰漫 (森林保育 處,2003)。過午後即見雲霧瀰漫,霧滴受樹冠攔截而增加局部降水量,
大氣中相對濕度極高,林區內樹幹上常見蘚苔類植物著生,依台灣地理 氣候區之劃分本研究區域屬於東北以及西北內陸區,以山地植群帶的區 分本研究區域屬於櫟林帶上,亦是一般所稱之盛行雲霧帶(prevalent cloud zone),氣候屬恆濕型氣候,濕度大,全年無乾季(蘇鴻傑,1984;1992)。
惟本區處於颱風經常行進路徑上,每年 6 至 9 月受颱風侵襲機率高,自 1992 年至 2006 年共計有 87 個颱風侵襲台灣,侵襲本研究區颱風計有 23 個(附表 1),每年平均約 1.6 個。於冬季偶有降雪。
(三)地質與土壤
棲蘭山檜木林地質大部分是經過變硬或變質的第三紀巨厚泥質沈積 岩所組成,包括深灰或灰黑色劈理良好的硬頁岩(argillite)、板岩(slate)以 及千枚岩(phyllite)等(王鑫,2000);63﹪之表土層深度在 20-50 ㎝,餘皆 在 20 ㎝以下,表土富鬆脆腐質層(邱志明等,1993)。以腐質壤土分布最 廣,土壤成灰暗色或黃褐色,團粒構造疏鬆通氣良好,適宜林木生長,
其次為砂質壤土、石礫土、腐質土等,土壤多呈偏酸性的反應,pH 值常 在 3.5~4.5。而在急斜地及山陵線上之表土,因土壤淺薄易使林木不易生 長,多為箭竹地、草生地或岩石地等。
本研究樣區之土壤調查,於 2005 年 6 月時,各將樣區畫分成 4 個小 樣區,依土壤深度的變化,在每個小樣區內採集 0-10 與 10-20 ㎝的土壤 樣本與土環,攜回宜蘭大學土壤實驗室分析。土壤分析項目包括土壤物 理及化學性質,分析方法參照張瑀芳(2006)。經調查結果本研究試驗區 土壤物理性質,台灣扁柏樣區土壤質地多為坋質粘土,其次為坋質粘壤 土、粘土、砂質壤土;而紅檜樣區土壤質地多為坋質粘土,其次為坋質
壤土、粘土、坋質粘壤土。台灣扁柏樣區土壤化學性質,表層(0-10 ㎝) pH 為 4.1,有機碳含量為 141.0 g/kg,陽離子交換容量(CEC)為 65.0 coml(+)/kg soil;裏層(10-20 ㎝),表層(0-10 ㎝) pH 為 4.4,有機碳含 量為 84.4 g/kg,陽離子交換容量(CEC)為 48.2 coml(+)/kg soil。紅檜 樣區土壤化學性質,表層(0-10 ㎝) pH 為 4.1,有機碳含量為 160.0 g/kg,
陽離子交換容量(CEC)為 88.7 coml(+)/kg soil;裏層(10-20 ㎝),表層 (0-10 ㎝) pH 為 4.4,有機碳含量為 67.0 g/kg,陽離子交換容量(CEC)為 63.8 coml(+)/kg soil。
二、研究方法 (一)研究材料
本研究之台灣扁柏與紅檜苗木,其來源為 1995 年採自棲蘭山林區大 溪事業區第 94 林班之天然生母樹種子,1996 年於森林保育處明池苗圃 播種育苗,1998 年選擇 30 ㎝以上 2 年生生長健壯之裸根苗出栽至試驗 地。
(二)苗木量測
栽植試驗設置後之最初 2 年每季量測苗木株數、苗徑及苗高基本資 料。苗高為苗木主幹之生長點頂部至地面根與莖交接之長度,苗徑為小 苗主幹接近地面和根交接處之直徑。栽植 3 年後年每半年量測乙次。2005 年 12 月調查的苗木性狀增加胸徑、枝下高、囓齒類動物危害情形等。
2006 年 12 月調查每一樣區內未受人為傷害及樹勢較好之幼木 20 株,量 測林木上、下、左、右之冠幅。
(三)冠層環境量測
本研究樣區最初於 1998 年以目測方式依不同冠層覆蓋度設立,為量
化樣區冠層環境,於 2004 年利用半球面鬱閉度計(spherical densiometer) 方法重新測定 16 個樣區之上方冠層環境,量測方法為:
1.樣區鬱閉度量測
於陰天時手持半球面鬱閉度計離地面高 2.5 m 保持水平,每一樣 區量測 21 個測點(圖 3),每一測點量測左右兩邊(面對上坡)鬱閉度,
計算其平均值,量測時間為 2004 年 10 月。
圖 3 半球面鬱閉度計(spherical densiometer)量測位置圖( 20m×20m 中心 點, 10m×10m 中心點, 5m×5m 中心點)
Fig 3 The diagram of measurement positions of spherical densitometer in the study area. ( Center point of plot (20m×20m ); Center point of plot (10m×10m ); Center point of plot (5m×5 m ))
2.試驗區林冠水平結構分析
為瞭解研究試驗區(約 2.4 ha)經人為干擾(枯立倒木移除及人工 造林作業)、及天然干擾(颱風),對冠層環境是否有影響,因此以不 同時期拍攝之航空照片(購自農林航空測量所),比較枯立倒木整理 前及後之差異。
(1)單點定位
於試驗區外較空曠處(林道旁),利用 GPS 測定控制點座標,
GPS 為萊卡公司(Leica)生產的 SmartRover,天線型號為 AX1230,
訊號接收器型號為 RX1250X。
(2)樣區邊點定位
利用經緯儀與已知控制點座標,以導線測量的方式求得試驗區 及各樣區四個角的座標。經緯儀為 TOPCON 公司製造的全站式光 波測距經緯儀(GPT2000)。
(3)試驗區上層林冠水平結構分析方法
為瞭解試驗區檜木林林冠水平結構是否受人工干擾(枯倒木移 除及人工造林作業)及天然干擾(颱風)影響,利用航空照片量化試 驗區林冠水平結構變化,由於多數航空照片因失真或有雲遮擋無 法利用,經挑選後本研究利用枯倒木整理前(1992)、整理後初期 (1997)、整理後 6-7 年(2003、2004)棲蘭山地區之航空照片,經與 地面控制點之定位與糾正後取得試驗區林冠覆蓋之影像。利用前 述地面控制點將地面樣區境界測量之結果旋轉平移至 97 二度分帶 座標系統。在 ArcGIS 下利用樣區邊界,數化檜木樹冠與孔隙以及 柳杉林覆蓋之區域影像將各類型之區集儲存成*.shp 格式。在 ArcGIS 下,利用向量與網格模式轉換之模組,採用 0.86 m×0.86 m 解析度將前述 shp 格式轉換成 ArcGrid 之格式,帶入 Fragstats 下計 算孔隙水平異質性之相關指標。
Fragstats 常用來計算描述地景(landscape level)、類別(class level)與區集(patch level)等層級之組成(composition)與空間配置 (Configuration)之指標。本研究以試驗區之單一孔隙或樹冠即為一 個區集;所有孔隙或所有樹冠各構成一個類別,即分孔隙類別、
樹冠類別以及柳杉類別即構成本研究試驗區之地景。此即構成不 同地景層級。
以 Fragstats 分析試驗區內孔隙水平異質性之相關指標,
CA(total class area ):表示該類別佔整個試驗區之面積,單位為 ha。
LPI(largest patch index):表示該類別中最大區集(patch)佔試驗區之 面積百分比。PLADJ(percentage of like adjacencies):同一類別內各 區集聚集的程度,值域在 0-100,當值越近 0 表示越分散,越近 100 表示越聚集。 CLUMPY(clumpiness index):用以量測該類型在空 間分散的程度,值域在-1 與 1 之間,越靠近-1 表示該類型區集越 分散,0 表示該類型分散為隨機,越靠近 1 表示該類型分布越聚集。
PLADJ=
( )
1001
⎟⎟
⎟⎟
⎠
⎞
⎜⎜
⎜⎜
⎝
⎛
∑
= mk ik ii
g
g
PLADJ 計算的概念為,將構成地景之所有網格(cells),以兩兩 網格成對(以下稱網格對)為計算基礎,計算所有同屬於某類別之同 類別網格對 (like-adjacencies) 對數(即上式中分子的部份)佔所 有類別網格對之比例(即上式中分母的部份)再乘以 100。
Given Gi =
⎟⎟
⎟⎟
⎟
⎠
⎞
⎜⎜
⎜⎜
⎜
⎝
⎛
−
⎟⎠
⎜ ⎞
⎝
⎛
∑
= i
m
k u
u
e g
g
min1
CLUMPY =
⎥⎥
⎥⎥
⎦
⎤
⎢⎢
⎢⎢
⎣
⎡
−
−
<
− <
Pi Pi Gi
else P
P P forG
P G
i i i i
i i
1
, 5
&
CLUMPY 的概念與 PLADJ 相似,惟做了一個類別最小周長 的校正項(min-ei)。
(四)試驗區林分結構分析
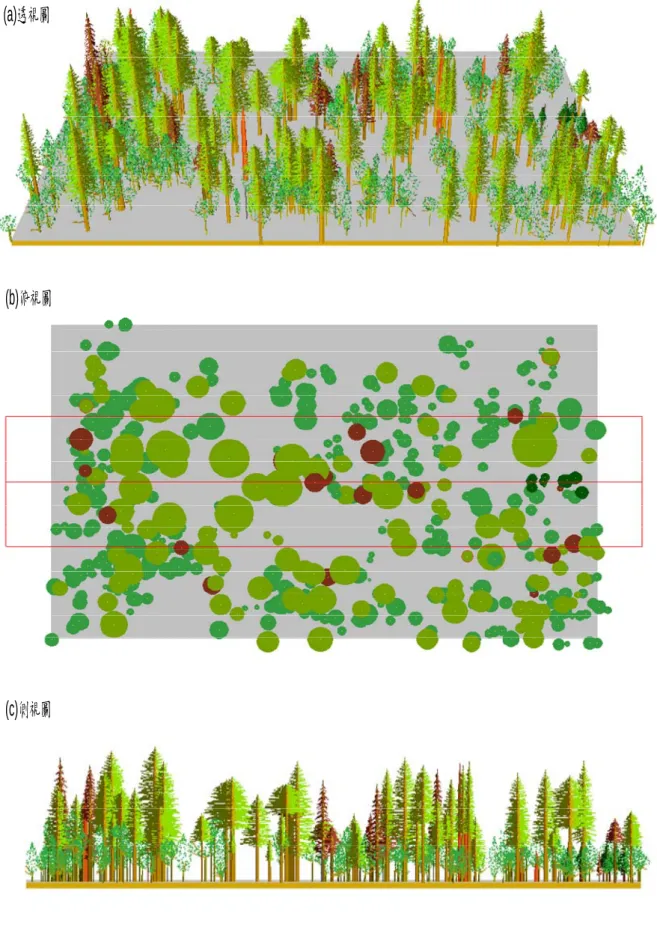
量測每一樣區內 DBH>10 ㎝上木之樹高、胸徑、枝下高、冠幅(以 林木為中心依橫坡及縱坡分上、下、左、右四邊),並定位量測上木之相 對位置(X、Y 座標),利用美國林務署林分視覺景觀系統軟體(SVS 3.30 version)繪製樣區林分結構示意圖。
林分視覺景觀系統(SVS)係將單株林木或倒木屬性量化繪製成圖,其 應用範圍包括:1.真實顯示單株林木和倒木之組成。2.可顯示目前整體林 分內之林分特性。3.使用不同的植物形式、顏色或者標示其他類型,以 區別不同之林分組成。4.林分顯示方式有俯視圖、側視圖、透視圖三種。
5.提供定義冠層內植物種類、外形、顏色、發育形式及林木位置之機制。
6.以簡易列表和圖表方式提供林分資訊(Robert,1997)。
本研究為量化冠層環境狀況以半球面鬱閉度計量測各樣區冠層鬱閉 度;另以 SVS 軟體繪製林分結構圖,以瞭解林分組成結構與環境特性,
提供森林經營者更多元的資訊。
三、資料分析
利用 SAS 統計軟體,以雙因子變異數分析法檢定林冠鬱閉度和檜木樹 種之生長表現。各樹種在不同鬱閉度下苗木生長特差異,則以單因子變異 數分析法檢定比較,若處理間具顯著差異(p<0.05),則進一步以 Duncan’s 檢定法進行事後分析。並以相關係數方法檢定紅檜及台灣扁柏苗生長參數 間之相關性。分析項目包括苗木死亡率:(樣區之苗木死亡株數/苗木栽植株 數)×100。健壯商數(sturdiness quotient,SQ):苗高(㎝)/苗徑(㎜)比值,表現 苗木生長形質,數值越大表示越纖弱。苗徑生長率(growth rate):(當年苗徑 -前一年苗徑)/前一年苗徑。苗高生長率:(當年苗高-前一年苗高)/前一年苗 高。冠長比(live-crown ratio):冠長/苗高;冠長=苗高-枝下高。冠形比(crown shape ratio):樹冠幅/冠長,樹冠幅為半徑。
肆、結果
一、試驗區上層林冠結構分析 (一)冠層水平結構異質性變動
以 1992、1997、2003、2004 年拍攝之航空照片,利用 FRAGSTATS 數化研究區冠層狀況(圖 4),比較不同時期冠層環境之變動,結果如表 1。孔隙總面積比例(CA%)在 1997 年稍微降低,隨後又逐漸上升。孔隙 最大區集的面積(LPI)在 1992 年到 1997 年之 5 年內上升近 9 %,而 在 1997 到 2003 年之 6 年內上升近 28 %,2003 到 2004 年上升近 6 %,
如以平均速率來看第一時期每年上升 1.8%;第二時期每年上升 4.6%;
第三時期則為近 6%。因此可瞭解孔隙逐年增加,且最大之孔隙有擴大 之趨勢。孔隙在試驗區中聚集程度(PLADJ),顯示孔隙在試驗區中空間 分布各年均呈聚集分布。孔隙在試驗區中空間分散程度(CLUMPY),
各年分散程度的變化則較小。由此可知試驗區之孔隙擴大之狀況應多 在前期孔隙之位置鄰近部份增加擴大。
(二)試驗林分組成結構分析
冠層結構之異質性,並非僅有水平之差異,尚有垂直方面之差異。
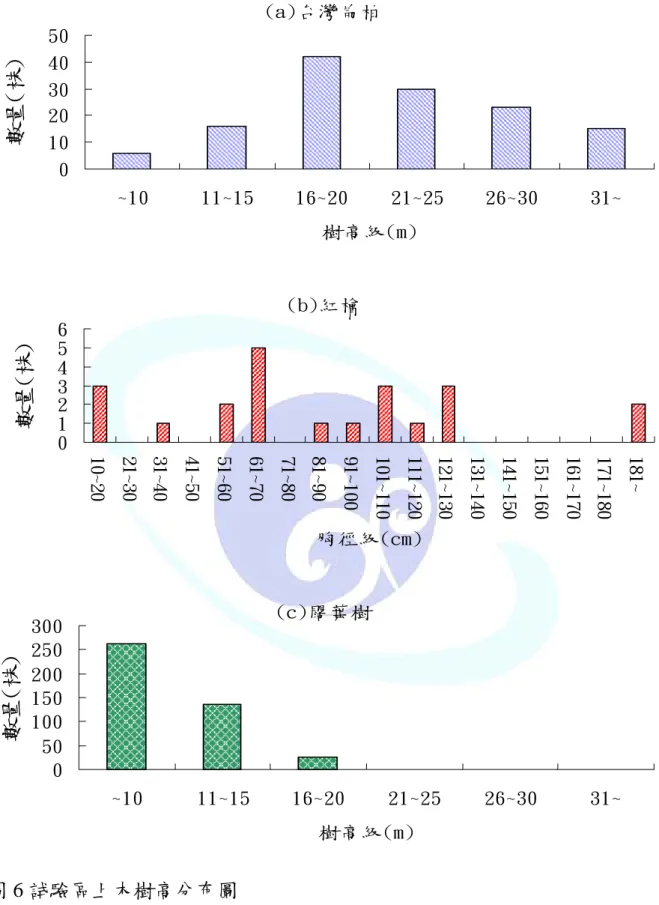
經調查試驗區中 DBH 大於 10 ㎝上木,結果如表 2,闊葉樹佔 71.74%
最多,平均胸徑 25.1 ㎝,平均樹高 10.4 m;台灣扁柏佔 23.24%,平均 胸徑 91.7 ㎝,平均樹高 21.5 m;紅檜佔 3.72%,平均胸徑 91.3 ㎝,平 均樹高 21.3 m。由圖 5 試驗區胸徑級分布顯示闊葉樹呈現反 J 型,偏 態係數為 2.51 呈右偏型,表示小徑級林木較多;台灣扁柏則呈連續分 布,偏態係數為-0.11 呈左偏型,表示中大徑級林木較多;紅檜則呈不 連續分布而且株數不多,偏態係數為 1.50 呈右偏型,表示中大徑級林 木較多。由圖 6 顯示試驗區冠層優勢林木為台灣扁柏呈現連續分布,
偏態係數為-0.69 呈左偏型,表示冠層及次冠層林木較多;紅檜則上、
中、下層皆有呈連續分布,但數量不多,偏態係數 0.85 呈右偏,表示 次及下層林木較多。中、下層林木則為闊葉樹為主數量最多呈反 J 型,
偏態係數為 2.27 呈右偏,表示下層林木較多。以 SVS 繪製研究樣區林 分組成結構圖(圖 7),顯示本區上層以台灣扁柏為優勢。
(三)試驗區內樣區上層林冠鬱閉度分析
樣區設置之初(1998 年)以目測方式判釋其林冠鬱閉度分為空曠、
稀、中、密 4 級,為進一步量化各樣區之林冠鬱閉度,於 2004 年以半 球面鬱閉度計(spherical densiometer),每個樣區量測 21 個測點資料,計 算樣區內之鬱閉度。樣區之林冠鬱閉度平均為 60±10.6%,屆於 37~77%
間,配合原目測之分級,分為 3 級即疏、中、密林冠鬱閉度級(表 3)。
各樣區於二次測量間略有變化,其原因可能為人為量測差異,及歷年 經颱風干擾而使現況與先前林冠略有變動。
表 1 試驗區不同時期孔隙及林冠變動情形
Table 1 Canopy dynamic pattern in experimental site.
年度
類別 1992 年 1997 年 2003 年 2004 年 CA(Total Class Area) (ha)
孔隙 0.85 0.84 1.07 1.22
樹冠 1.23 1.24 1.01 0.86
柳杉造林地 0.25 0.25 0.25 0.25
合計 2.33 2.33 2.33 2.33
CA%
孔隙 36.41 36.17 46.02 52.43
樹冠 52.97 53.21 43.35 36.95
柳杉造林地 10.62 10.62 10.62 10.62 LPI(Largest Patch Index)
孔隙 (%) 5.33 13.86 40.59 46.74 樹冠 (%) 50.49 40.72 24.42 13.41 柳杉造林地(%) 10.62 10.62 10.62 10.62 CLUMPY(Clumpiness Index)
孔隙 0.82 0.84 0.84 0.86
樹冠 0.84 0.84 0.85 0.85
柳杉造林地 0.98 0.98 0.98 0.98
PLADJ(Percentage of Like Adjacenices)
孔隙 87.93 88.49 91.03 92.73
樹冠 91.57 91.87 90.65 89.86
柳杉造林地 96.34 96.34 96.34 96.34
圖 4 試驗區不同時期上層林冠孔隙變動情形
Fig. 4 The main canopy gap dynamic in experimental site.
1992 1997
2003 2004
表 2 試驗區林分上木組成結構
Table 2 The structure of canopy trees in experimental site.
樹種
類別 台灣扁柏 柳杉 紅檜 闊葉樹
株數 132 13 22 424
平均胸徑(㎝) 91.7±41.4# 21.7±6.0 91.3±55.0 25.1±11.1 平均樹高(m) 21.5±6.5 9.0±1.8 21.3±7.1 10.4±2.7
株數(%) 22.34 2.20 3.72 71.74
胸徑分布峰度係數 1.63 2.1 1.42 6.81
胸徑分布偏態係數 -0.11 2.45 1.50 2.51
樹高分布峰度係數 2.63 2.24 2.15 2.06
樹高分布偏態係數 -0.69 1.58 0.85 2.27
#Mean±SD
(a)台灣扁柏
02 46 108 1214 1618
10~20 21~30 31~40 41~50 51~60 61~70 71~80 81~90 91~100 101~110 111~120 121~130 131~140 141~150 151~160 161~170 171~180 181~
胸徑級(cm)
數量(株)
(b)紅檜
0 2 4 6 8
~10 11~15 16~20 21~25 26~30 31~
樹高級(m)
數量(株)
(c)闊葉樹
0 50 100 150 200
10~20 21~30 31~40 41~50 51~60 61~70 71~80 81~90 91~100 101~110 111~120 121~130 131~140 141~150 151~160 161~170 171~180 181~
胸徑級(cm)
數量(株)
圖 5 試驗區上木胸徑分布圖
Fig. 5 The DBH distribution of canopy trees in experimental site.
(a)台灣扁柏
0 10 20 30 40 50
~10 11~15 16~20 21~25 26~30 31~
樹高級(m)
數量(株)
(b)紅檜
0 1 2 3 4 5 6
10~20 21~30 31~40 41~50 51~60 61~70 71~80 81~90 91~100 101~110 111~120 121~130 131~140 141~150 151~160 161~170 171~180 181~
胸徑級(cm)
數量(株)
(c)闊葉樹
0 50 100 150 200 250 300
~10 11~15 16~20 21~25 26~30 31~
樹高級(m)
數量(株)
圖 6 試驗區上木樹高分布圖
Fig. 6 The height classes distribution of canopy trees in experimental site.
(a)透視圖
(b)俯視圖
(c)側視圖
圖 7 林分視覺景觀系統(SVS)試驗區上層林木結構示意圖
Fig. 7 Main canopy structure, view demonstrated with stand visualization system (SVS).
表 3 樣區鬱閉度分級
Table 3 The closure degree classes of sample plots.
樹種 樣區
鬱閉度 分級#
鬱閉度 (%)&
原目測
覆蓋度 樹種 樣區
鬱閉度 分級#
鬱閉度 (%)&
原目測 覆蓋度
台灣扁柏 C Ⅰ 47±9.4 空曠 紅檜 T Ⅰ 37±13.1 空曠
台灣扁柏 J Ⅰ 53±3.4 稀 紅檜 U Ⅰ 46±7.5 空曠
台灣扁柏 A Ⅱ 57±10.3 空曠 紅檜 E Ⅱ 57±7.0 稀
台灣扁柏 K Ⅱ 57±9.0 中 紅檜 L Ⅱ 59±4.5 稀
台灣扁柏 M Ⅱ 59±6.8 中 紅檜 N Ⅱ 60±4.8 中
台灣扁柏 Q Ⅲ 70±8.3 密 紅檜 O Ⅱ 68±1.8 中
台灣扁柏 P Ⅲ 71±3.6 密 紅檜 S Ⅲ 71±2.3 密 紅檜 R Ⅲ 77±2.2 密
#: Ⅰ(疏):鬱閉度<53%;Ⅱ(中):鬱閉度>54-69%;Ⅲ(密):鬱閉度>69%。
&
: Mean±SD
#:I (weak): closure ratio<53% II (medium) closure ratio>54-69% III (strong):
closure ratio >69%. &: Mean±SD
二、不同冠層環境下檜木苗死亡率之比較 (一)不同鬱閉度下檜木苗之死亡率
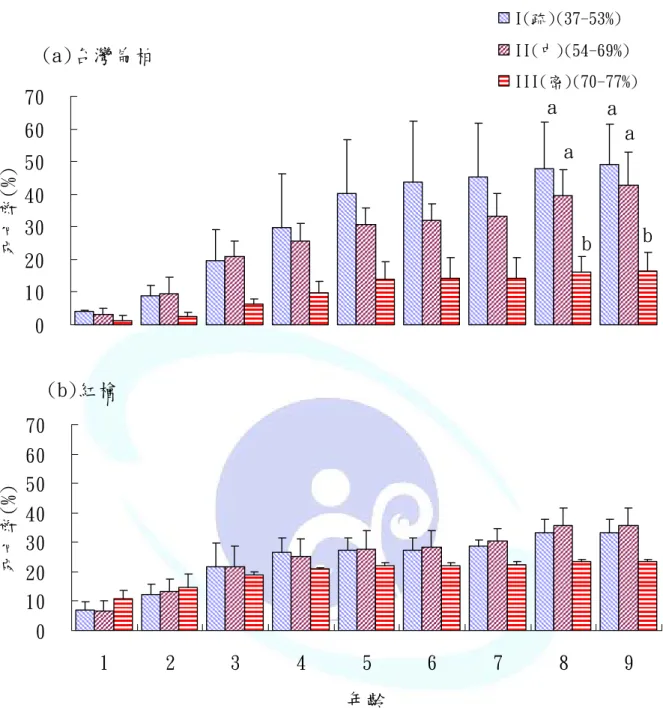
檜木苗栽植後 9 年間,各苗齡之死亡率經雙因子變異分析檢定結 果如表 4 及附表 2,以栽植初期即 1 年生及 2 年生時紅檜及台灣扁柏呈 顯著性差異,以紅檜苗之死亡率高於台灣扁柏,至 3 年生後並無顯著 差異。檜木苗栽植 4 年後之死亡率於林冠鬱閉度間有顯著差異,以鬱 閉度密環境下死亡率較低。而林冠鬱閉度與樹種間於 1 年生及 2 年生 時有交感現象,在密鬱閉林冠下紅檜苗木死亡率較高,而台灣扁柏苗 則相對較低,由初期檜木苗死亡率之表現,顯示台灣扁柏較紅檜耐蔭。
由單因子變異分析分別檢定紅檜及台灣扁柏於不同林冠鬱閉度下 之存活表現(圖 8),僅台灣扁柏於 8 年生及 9 年生時呈現顯著性差異,
於密鬱閉林冠下,死亡率較低僅為 16%,而於中、疏鬱閉度下死亡率 則大於 40%。台灣扁柏於栽植初期苗木死亡率隨年齡逐漸增加,尤其 於中、疏林冠環境下,至 5 年生時始趨於緩和。紅檜苗死亡率於林冠 鬱閉度間無顯著差異。但就平均值表現於 1 年生及 2 年生時,密鬱閉 林冠下之死亡率卻高於中、疏林冠環境下,表示紅檜幼苗生長初期較 不適於密閉林冠環境下,而至 3 年生後則以林冠環境中、疏環境下死 亡率較高。
表 4 不同林冠鬱閉度下各苗齡檜木苗之死亡率(%)變異分析表
Table 4 ANOVA of mortality rate (%) of cypress seedlings at each age class under canopy closure degrees.
來源 1 年生 2 年生 3 年生 4 年生 5 年生 6 年生 7 年生 8 年生 9 年生 樹種 20.94*
(1,9)
15.22*
(1,9)
3.79 (1,9)
0.81 (1,9)
0.35 (1,9)
0.91 (1,9)
0.91 (1,9)
0.99 (1,9)
1.82 (1,9) 鬱閉度級 0.08
(2,9)
0.55 (2,9)
3.43 (2,9)
4.42*
(2,9)
5.56*
(2,9)
5.92*
(2,9)
8.08*
(2,9)
9.58*
(2,9)
9.74*
(2,9) 樹種×
鬱閉度級
4.56*
(2,9)
4.67*
(2,9)
2.67 (2,9)
1.94 (2,9)
2.44 (2,9)
2.76 (2,9)
3.14 (2,9)
2.18 (2,9)
2.35 (2,9)
*: p<0.05;(NDF,DDF ):NDF:為 F test 中分子的自由度;DDF:為 F test 中分母的自由度。
*: p<0.05; (NDF,DDF ):NDF:F test show degree freedom of numerator ; DDF:F test show degree freedom of denominator.
(a)台灣扁柏
0 10 20 30 40 50 60 70
死亡率(%)
I(疏)(37-53%) II(中)(54-69%) III(密)(70-77%)
a
b a a
b a
(b)紅檜
0 10 20 30 40 50 60 70
1 2 3 4 5 6 7 8 9
年齡
死亡率(%)
圖 8 不同林冠鬱閉度下各苗齡檜木苗之死亡率(直條圖上方未標字母及字母 相同者表示檢定後各樹種於林冠鬱閉度間無顯著差異,字母不相同者 表示有顯著差異(P<0.05))。
Fig. 8 Mortality rate of cypress seedlings at each age class under canopy closure degrees. (Vertical bar chart diagram upper unmark letter or same mark letter means non-significance, the distinct letter means
significance(p<0.05))
(二) 不同鬱閉度下檜木苗受囓齒類動物危害之比率
9 年生檜木栽植苗受囓齒類動物危害之比率,經以雙因子變異分析 檢定結果,在不同林冠鬱閉度下有顯著差異(F=10.14,p<0.05),以林 冠鬱閉度中、疏環境下受害比率較大。兩種檜木苗間無顯著差異,且 鬱閉度與樹種間無交感現象。
由單因子變異分析分別檢定紅檜及台灣扁柏於不同林冠鬱閉度下 之受害比率(圖 9),台灣扁柏呈現顯著性差異,於中、疏鬱閉度下受害 比率較大,而紅檜苗則呈不顯著性差異,然仍以中、疏鬱閉度下受害 比率較大。
0.0 10.0 20.0 30.0 40.0 50.0 60.0 70.0
I(疏) II(中) III(密)
冠層鬱閉度級 I(疏)(37-53%) II(中)(54-69%) III(密)(70-77%)
株數(%)
台灣扁柏 紅檜 a
b a
圖 9 不同林冠鬱閉度下檜木苗受囓齒類動物危害情形(直條圖上方未標字母 及字母相同者表示檢定後各樹種於林冠鬱閉度間無顯著差異,字母不 相同者表示有顯著差異(P<0.05))。
Fig. 9 The damage symptom of cypress seedlings by rodent under canopy closure degrees. (Vertical bar chart diagram upper unmark letter or same mark letter means non-significance, the distinct letter means significance (p<0.05))
三、不同冠層環境下檜木苗高生長之比較 (一)不同冠層環境下檜木苗之苗高
檜木苗栽植後 9 年間,各苗齡之苗高經雙因子變異分析檢定結果如 表 5 及附表 3,栽植 9 年間兩樹種間呈顯著性差異,以紅檜苗之苗高大