國立臺灣大學生物資源暨農學院 植物病理與微生物學系
碩士論文
Department of Plant Pathology and Microbiology College of Bioresources and Agriculture
National Taiwan University Master Thesis
十字花科根瘤病生物防治資源之開發與應用 Exploration and application of bio-control resources
for the clubroot disease of Brassicaceae
江維翰 Wei-han Chiang
指導教授:沈偉強博士 Advisor: Wei-Chiang Shen, Ph.D.
中華民國 102 年 7 月
July 2013
i
ii
中文摘要
十字花科根瘤病(clubroot disease of Brassicaceae)是由絕對寄生之內部寄生
性黏菌 Plasmodiophora brassicae Woronin 所引起之十字花科重要病害。目前本
病害多以施用石灰提高土壤酸鹼值,進行防治,但也造成土壤鹽鹼化之問題,因
此開發其他防治方法,實為刻不容緩之事。本論文目標有三,一為藉由實際的田
間調查,了解臺灣根瘤病之發病狀況,以及農民耕作習慣與防治施行等;二為嘗
詴探討臺灣 P. brassicae 小種的不同,提供十字花科作物育種及田間品系種植選
擇 之 參 考 資 訊 ; 三 為 開 發 生 物 防 治 資 源 , 藉 由 分 離 田 間 土 壤 中 具芥 子 酶
(myrosinase)活性之微生物,分解硫配醣體(glucosinolate)產生異硫氰化物
(isothiocyanate),進行十字花科根瘤病生物防治燻蒸測詴。針對上述目標,實
際調查田間發病狀況,全省各主要高冷蔬菜產區皆有此病害之發生,發病率於10
至100%不等。在根瘤病菌小種分群方面,我們自臺灣8縣市23處病田採得樣本,
嘗詴進行分子分群詴驗,結果發現臺灣根瘤病菌無法以 rDNA 進行分群,但以
一特殊序列分子標記可發現臺灣帄地和高山的菌株分屬不同的小種,可作為未來
進一步分群之用。在分離具芥子酶活性之微生物方面,自臺灣許多十字花科蔬菜
種植田,分得19株具有芥子酶活性的真菌,並進行其中5株真菌之生長特性及防
治測詴,發現由臺北市北投區分離之Aspergillus japonicus var. aculeatus,不論在
iii
中溫(20-30°C)生長速度,及於植物根部根圈定棲能力,皆有不錯之能力,而
於溫室和田間詴驗中,當該菌和菜籽粕共同施用於土壤時,對根瘤病皆得到穩定
且優良的防治效果。藉由本論文的研究,找到頗具生物防治潛力之真菌,期待未
來能結合耕作防治,應用於十字花科根瘤病之防治,提供有效且對環境無不良影
響的防治新方法。
關鍵詞:十字花科根瘤病,Plasmodiophora brassicae,芥子酶,硫配醣體,異硫
氰酸酯,生物燻蒸,小種
iv
Abstract
Clubroot disease of Brassicaceae, caused by an obligate endo-parasitic slime mold
Plasmodiophora brassicae Woronin, is one of serious problems of cruciferous crop.
Currently, one of major control methods for clubroot disease is to elevate pH value by
adding lime into soil; however, detrimental effects including soil alkalization and
hardening often occur and exhibit impact on plant growth and environment. Thus,
alternative and effective control measures are currently in need. The goal of our study
is to explore bio-control resources and develop new disease control method for
clubroot disease. To achieve this goal, three specific aims include: (i) to investigate
the occurrence of clubroot disease in the high elevation fields of Taiwan; (ii) to collect
and differentiate P. brassicae races; (iii) to isolate myrosinase-producing microbes
and develop a biofumigation control protocol based on degradation products,
isothiocyanate, of glucosinolates. Our field survey revealed that clubroot disease
occurs in the fields of high elevation growth areas with incidence of 10-100%. Twenty
three clubroot samples collected from 8 different counties in Taiwan were subjected to
race differentiation experiments. We failed to differentiate these samples by rDNA
sequence, but samples from plain fields or high mountains can be separated into 2
v
pathotypes based on specific sequence marker. Furthermore, 19 myrosinase-producing
fungal isolates were isolated from field samples. Among these fungi, 5 isolates were
subjected to growth and disease control assays and one isolate from Beitou,
Aspergillus japonicus var. aculeatus showed good growth ability at 20-30°C and
extensive root colonization capability. In both green house and field tests, this isolate
also exhibited good disease control results when combining with canola meal. In this
study, a myrosinase-producing fungal isolate was identified and combination of this
bio-control agent with other culture control measures may potentially provide an
effective and environmental- friendly method to control the clubroot disease.
Keywords: cluberoot disease of Brassicaceae; Plasmodiophora brassicae; myrosinase;
glucosinolate; isothiocyanate; biofumigation; race
vi
目錄
口詴委員會審定書... i
中文摘要... ii
英文摘要... iv
一、前言... 1
二、前人研究... 3
2.1 十字花科根瘤病菌... 3
2.2 硫配醣體(glucosinolate)和芥子酶(myrosinase)系統... 7
2.3 生物燻蒸(bio-fumigation) ... 10
2.4 根瘤病菌小種(race)的分群 ... 11
三、材料與方法... 14
3.1 臺灣地區十字花科根瘤病樣本收集、病害調查與小種分群... 14
3.2 具芥子酶活性之真菌分離、培養與鑑定... 17
3.3 根瘤病菌接種原之製備及接種... 18
3.4 具芥子酶活性真菌特性測詴... 19
3.5 具芥子酶活性真菌之溫室防治測詴... 21
3.6 具芥子酶活性真菌之田間防治測詴... 23
vii
3.7 Aspergillus japonicus var. aculeatus 簡單製劑化及其田間效果初探... 25
3.8 防治機制初探... 27
四、結果... 31
4.1 臺灣十字花科根瘤病之病害調查及病株樣本之採集... 31
4.2 根瘤病菌 DNA 之抽取、增幅與分子分群 ... 33
4.3 具芥子酶活性真菌分離、培養與鑑定... 34
4.4 具芥子酶活性真菌特性測詴... 36
4.5 具芥子酶活性真菌之溫室防治測詴... 37
4.6 田間測詴... 38
4.7 初步製劑化之田間測詴... 39
4.8 異硫氰酸酯與生物燻蒸對於休眠孢子的影響... 40
五、討論... 42
六、參考文獻... 50
七、表... 59
八、圖... 75
附錄... 102
1
壹、 前言
十字花科植物為臺灣極重要的蔬菜種類,以甘藍為例,民國101年全國栽植
面積達7,369公頃,總產量達341,279公噸(農委會,2013)。十字花科作物的病害
甚多,而根瘤病(clubroot)為其重要病害之一。本病由一種絕對寄生之內部寄
生性黏菌Plasmodiophora brassicae Woronin所引起。十字花科根瘤病於臺灣夏季
好發於臺中福壽山、南投仁愛鄉等高冷蔬菜生產區,而冬季則多出現於桃園新竹
等北部地區的冬季蔬菜產區,影響作物品質產量頗鉅。若是在氣候、土壤環境適
當,且田間蔬菜連作時,根瘤病極可能會發生大流行。十字花科根瘤病之發生,
受環境及氣候的影響甚大,在溫度20~25°C,濕潤(70~80%土壤含水量),酸性
(pH 5-6左右)的土壤環境最適合根瘤病菌的生長繁殖。傳統上,防治十字花科
根瘤病多以施用石灰、S-H混合物、矽酸爐渣等方式,藉由提高土壤酸鹼度至7.4
以上來防治本病(蔡,1987)。但這些物質的過度使用,將造成土壤因鈣離子過高
而鹽鹼化的副作用,且不利好弱酸性土壤之十字花科作物的生長。本病害雖有推
薦農藥氟硫滅,但效果不佳且價格昂貴,亦不符現今推展無毒農業之趨勢。吾人
已知十字花科作物多會產生硫配醣體(glucosinolate),其會被芥子酶(myrosinase)
分解進而產生異硫氰酸酯(isothiocyanate),該產物已知在較高濃度下對許多植
物病原菌有毒殺效果或抑制其生長,此結果或可應用於十字花科根瘤病之防治。
2
根據前人研究,有許多土壤微生物可產生該酵素並分解硫配醣體,但目前尚未應
用於根瘤病之病害防治,若進一步了解其作用機制,進行防治測詴並將其實用化,
除可有效防治根瘤病,亦符合未來無毒農業之潮流。十字花科作物內的硫配醣體
之分解產物也可能影響休眠孢子的行為。若是能夠更加了解各項影響的因子,或
可以此修正現行之防治方式。除此之外,藉由田間訪查、收集樣本、進而以分子
生物學方法了解臺灣根瘤病菌的小種差異,或可調整栽培習性,以達到耕作防治
的效果。本論文之總目標,在於自土中分離具芥子酶活性之真菌,並以之進行防
治測詴及應用、了解孢子發芽之影響因子與生物燻蒸對休眠孢子之影響等三方面,
搭配田間訪查蒐集樣本和了解根瘤病菌小種及其分布,及了解目前田間實際耕作
及防治情況,期望研究成果可應用於十字花科根瘤病之耕作防治及生物防治上,
並減少傳統化學防治方法對作物生長及環境等方面的不良影響。
3
貳、 前人研究
一、十字花科根瘤病菌 Plasmodiophora brassicae Woronin
十字花科根瘤病菌(Plasmodiophora brassicae Woronin)為一種內部寄生性
黏菌(Plasmodiophoromycetes);其主要特徵為:(一)圓柱狀的細胞分裂;(二)
絕對內寄生的營生方式;(三)游走子著生前生馬鞭狀雙鞭毛;(四)具有高耐受
性的休眠孢子(Dixon, 2006)。根據small subunit ribosomal RNA(SSU rRNA)序
列分析結果,發現P. brassicae和雙鞭毛蟲門(Dinozoa)、纖毛蟲門(Ciliophora)
和頂複合器門(Apicomplexa)之親緣關係較相近,而和真菌界、雜色藻界等親
緣較遠(Castlebury and Domier, 1998)。根瘤病菌目前的分類地位如下:原生生
物 界 ( Protista )、 寄 生 黏 菌 門 ( Plasmodiophoromycota )、 寄 生 黏 菌 綱
( Plasmodiophoromycetes )、 寄 生 黏 菌 目 ( Plasmodiophorales )、 寄 生 黏 菌 科
(Plasmodiophoraceae)、寄生黏菌屬(Plasmodiophora)。
生活史
本病原以絕對寄生方式維生,其生活史複雜:病原於土壤內以休眠孢子殘存,
帄均殘存時間約 3.6 年,最長可達 18 年(Dixon, 2006);休眠孢子直徑約 3.0 至
5.0 μm,在掃描式電子顯微鏡下可見其表面有棘,單套單核,核直徑 1.5μm,俟
4
適當環境或寄主出現時,即發芽生成梨形、兩根前生馬鞭狀鞭毛之初生游走子
(primary zoospore)進行侵染(Macfarlane,1970)。根據前人研究,休眠孢子在
以下的狀況較容易萌發為游走子:(一)腐爛的根瘤;(二)十字花科植物之根分
泌物;(三)pH 6~6.5;(四)溫度 20~25°C;(五)相對濕度 70%以上(Macfarlane,
1970;Suzuki et al, 1992;Takahashi, 1994)。游走子的萌發開啟了該病原的生活
史,此時胞內產生一大液泡及侵入釘(penetration peg),藉由液泡產生的壓力將
侵入釘與胞質注入寄主細胞(Aist and Williams, 1971),侵入後在根毛內進行細
胞核分裂形成多核的初生原生質體(primary plasmodium),之後初生原生質體內
產生膜並將原生質體分割成多核的游走子囊(zoosporangium),游走子囊內的核
再行有絲分裂並產生單核的次生游走子(secondary zoospore),可再進入土中作
為感染原。具親和性的次生游走子進行細胞質融合(plasmogamy)轉為雙核態
(n+n),並侵染根部皮層;游走子於其內進行核分裂,產生多核態的次生原生質
體(secondary plasmodium),之後具親和性的兩核進行核融合(karyogamy),形
成雙套(2n)細胞核並進行減數分裂,此時次生原生質體轉為休眠胞子囊(resting
sporangium)並於其內生成無 數休眠胞子,釋放於土 中開始另一個侵染循環
(Ingram and Tommerup, 1972)。
5
發生生態
本病好發於氣候冷涼(20°C 左右)且濕度高之區域,故於溫帶地區容易大
發生,而臺灣地區則易發生於海拔較高之夏季蔬菜及帄地之冬季蔬菜產區。土壤
條件也是發病的決定性因子,除濕度(70~80%含水力)外,酸鹼度更是重要的
影響因素,通常在土壤呈微酸性(pH 5~6)時容易發病。本病所造成最明顯的病
徵即根部不規則畸形腫大,表面凹击不帄,影響根部對水份及養分的吸收,至後
期病根因細菌或真菌之二次感染而腐爛空洞化並發出惡臭,致使全株死亡;地上
部則因根部失去功能而造成全株萎凋、下位葉黃化,該症狀在溫度高、日照強之
正午時分尤為明顯,另外本病也會造成甘藍類作物無法結球或結球甚小,以致不
能收成。十字花科根瘤病在加拿大每年造成至少約 10%的損失,嚴重時幾乎全無
收成(Faggian et al, 1999;Xue et al, 2008)。
P. brassicae 寄主範圍主要為十字花科的蕓苔屬(Brassica spp.),如甘藍、油
菜、白菜、芥菜等皆是,另外像蘿蔔屬(Raphanus spp.)、阿拉伯芥(Arabidopsis
spp.)、山葵屬(Wasabia spp.)、芥子屬(Sinapis spp.)等十字花科植物亦為該病
原之寄主。此外在某些非十字花科作物中,根瘤病菌雖然可以侵入根部並完成生
活史,但是卻沒有明顯的根瘤及地上部萎凋症狀,如金蓮花(Tropaeolum majus)、
木瓜(Carica papaya)、甜菜(Beta vulgaris)等(Ludwig-Müeller et al, 1999.)。
6
而有些植物雖然不是根瘤病菌的寄主,但是其根分泌物卻有誘導休眠孢子發芽的
效果,如韭蔥(Allium porrum)、黑麥(Secale cereale)、黑麥草(Lolium perenne)
等(Friberg et al, 2006 )。
防治方法
目前植物保護手冊推薦的化學防治方式主要為 0.3% 氟硫滅(flusulfamide)
粉劑(商品名:根必強),其詳細的作用機制目前並不了解,但可能和影響孢子發
芽有關(Tanaka et al, 1999)。現在田間最常使用的防治方式為施用生石灰以提高
土壤酸鹼值至 7.4 以上及土中二價離子濃度,其原因應亦為影響孢子發芽(Niwa
et al, 2008;村上和畔柳,2007),但是土壤的酸鹼值及二價離子濃度過高將導致
土壤鹽鹼化及硬化,使農地不適於耕作,因此亟需研擬新的防治方法。
目前已有許多關於根瘤病生物防治的研究,包括 Trichoderma spp.(Cheah et
al, 1997)、Streptomyces spp.(Cheah et al, 2000)、Acremonium alternatum(Jäscheke
et al, 2010),但作用方式仍莫衷一是,可能為產生抗生物質、二次代謝物影響根
瘤病菌或根圈微生物相等原因,而 A. alternatum 可能以內部共生的方式影響根瘤
病之發病。另外有一種真菌 Phoma glomerata (Corda) Wollenw. & Hochapfel 雖然
不會直接對 P. brassicae 造成影響,但施用其培養液於植株卻可防止根瘤生成,
7
推測其原因可能為該菌產生一種拮抗生長素(IAA)的物質 epoxydon,使植株不
會因生長素過量產生根瘤(Arie et al, 1998)。在加拿大亦有學者以 Gliocladum
catenulateum 與 Bacillus subtilis 之製劑進行防治測詴,可有效防治油菜根瘤病之
發生達 80%(Peng et al, 2011),其中後者已被我國植物保護手冊列為治療本病之
推薦用藥。
二、硫配醣體(glucosinolate)和芥子酶(myrosinase)系統
植物體內常會有些二次代謝物(secondary metabolites),雖然這些物質並非
為植物生存所必需,但卻具有各式各樣的生理作用,並可為人類利用;硫配醣體
(glucosinolate)便是一類這樣的物質。硫配醣體又稱為芥子油苷(mustard oil
glucosides),主要分布於十字花科蕓苔屬(Brassica spp.) 及同科之植物,另外非
十字花科植物如旱金蓮科(Tropaeolaceae)、番木瓜科(Caricaceae)、木犀草科
(Resedaceae)等亦有之(Ludwig- Müller et al, 1999)。該物為一群含硫與氮、帶
負電、中性的醣苷(glycoside)分子,結構上可分為三部分:由硫氫基(-SH)和
β-D-glucose形成的葡萄糖苷、磺酸根、及來自各式胺基酸的支鏈,根據支鏈來源
之胺基酸及形式可將硫配醣體分為三群:芳香族硫配醣體(aromatic glucosinolates;
8
來自Phe和Tyr)、脂肪族硫配醣體(aliphatic glucosinolates;來自Ala、Leu,、Ile、
Met或Val)、吲哚族硫配醣體(indolic glucosinolates;來自Trp),上述胺基酸
經過chain elongation,由細胞色素79(CYP79)轉變為醛肟(aldoxime),最後接
上一個葡萄糖分子即成為硫配醣體(Halkier and Gershenzon, 2006)。
硫配醣體在植物中具有許多功能;吲哚族硫配醣體為植物生長素(IAA)的
前驅物(Ludwig-Müller et al, 1999),另外在植物的防禦機制上,該物為合成一
種植物殺菌素(phytoalexin)- camalexin的前驅物(Siemens et al, 2008),除此之
外也與系統性抗病(systemic defense)有關,如癒傷醣(callose)的形成(Clay et
al, 2009)。硫配醣體對於真菌、細菌性病原的直接毒性不高(Doughty et al, 1991),
但是當該物在自然狀況下或經酵素催化分解後所產生的物質卻可抑制微生物生
長或其他的生理反應(Vidhyasekaran, 2008)。因此接下來將敘述硫配醣體之分解
機制與其產物。
植物內所含的硫配醣體除在高溫下可自行分解外,自然界中最主要還是以芥
子酶(myrosinase)催化分解反應。芥子酶為貝塔-葡萄糖苷酶(β- glucosidase)
的一種,分布於植物組織中的特化細胞(myrosin cell)的細胞質中,除植物外,
亦有許多微生物具有該酵素,如Aspergillus sydowi、A. niger、A. oryzae、Fusarium
oxysporum、Enterobacter cloacae等(Ohtsuru et al, 1973;Tani et al, 1974;Smits et
9
al, 1993)。芥子酶在整個反應中將硫配醣體的葡萄糖分子剪除,形成一不穩定的
中間產物thiohydroxamate-O-sulfonates,該物在不同的反應條件下(如酸鹼值、
epithiospecifier protein或亞鐵離子存在與否等),可以被芥子酶重新排列成不同的
產物:異硫氰酸酯(isothiocyanate)、腈類(nitrile;epithionitrile)、硫氰酸酯
(thiocyanate)、唑烷硫酮(Oxazolidine-2-thione)(Halkier and Gershenzon, 2006)。
在這些產物中,最為人所熟知的為異硫氰酸酯。該物為芥子酶在酸鹼為中
性時分解硫配醣體所生的物質,依其所攜帶支鏈的不同可將之分為丙烯基型
(allyl type)和苯基型(phenyl type)兩類,具有揮發性及刺激性味道。如前所
述,硫配醣體對於微生物的直接毒性不高,但當其轉換為異硫氰酸酯時,便可抑
制許多微生物生長,如Aspergillus niger、Rhizopus oryzae(Drobnica et al, 1967)、
Fusarium oxysporum (Davis et al, 1964)、Leptosphaeria maculans(Mithen and
Lewis, 1986 )、 Glomus mosseae ( Vierheilig and Ocampo, 1990 ) 等 ; 而
Plasmodiophora brassicae於5 ppm的丙烯基異硫氰酸酯(allyl isothiocyanate)或2.5
ppm的苯基異硫氰酸酯(phenyl isothiocyanate)下,其休眠孢子的發芽即可被抑
制,但有趣的是,在更低的濃度下(1 ppm),兩種異硫氰酸酯反可促進休眠孢子
發芽(Hooker et al, 1945)。
長期以 來, 以該 物進 行病 害防治 的例 子不 勝枚 舉, 如土 壤燻蒸 劑邁隆
10
(Dazomet)即為一種丙烯基異硫氰酸酯的製品,本藥劑不僅可燻殺土中的真菌
病原外,亦可用以防治線蟲;除化學藥劑外,常用於有機防治的生物燻蒸法
(bio- fumigation)亦為一例。
三、生物燻蒸(bio-fumigation)
燻蒸(fumigation),乃是利用氣態有效殺菌物質滲透進入目標物,以達到除
滅害蟲與有害微生物的方法。吾人所熟知的燻蒸方式不外乎以化學藥劑(如溴化
甲烷等)進行之,但傳統的化學燻蒸劑對包括人在內的非目標生物均具極高毒性,
並且容易破壞環境,因此已漸被捨棄不用。在耕作防治法中也有應用類似方法以
達到防治效果者,此即為生物燻蒸。生物燻蒸(bio- fumigation)即將植物殘體、
組織或其他物質埋於土壤之中,利用微生物的酵素將其中部分物質分解並產生氣
體殺菌物質,以達到病害防治的效果;這其中最著名的例子即為利用十字花科植
物進行的生物燻蒸,本法乃埋入該科植物之前作或綠肥殘體,讓土壤中具芥子酶
活性的微生物分解硫配醣體產生異硫氰酸酯,產生土壤燻蒸的效果(Kumar,
2005)。本法已廣泛應用於土壤病害的防除,例如若在小麥栽植前種植油菜(canola;
Brassica napus L.)或印度芥菜(Indian mustard;B. juncea L. Czern and Coss.),
之後將之殘體犁入土中,可減少小麥全蝕病(wheat take-all disease)的發生(Angus
11
et al, 1994)。除利用栽植十字花科植物之綠肥之方法外,亦有將蕓薹屬(Brassica
spp.)榨油後之種子粉,和一具芥子酶活性之微生物Pseudomonas boreopolis搭配
施用於西瓜帅苗,具有防治因Pythium spp.所造成之帅苗猝倒病的效果(Chung et
al, 2003)。另外於土中施用尿素經微生物產生的尿素酶(urease),將尿素分解
產生氨氣,對於土壤病原也有防治之效果(方,1988),該方法已使用於褐根病
之防治,亦可算是生物燻蒸的一種應用。
四、根瘤病菌小種(race)的分群
許多病原在種以下,尚有小種的分類,不同的小種可影響同一種病原對寄主
的病原性,因此了解一病原的小種在植病研究上有其重要性。Honig(1931)證
明了P. brassicae也有小種之別,而最早被廣為使用的分群方法為Williams於1965
年 所 提出 ,利 用 甘藍 ( Brassica oleracea L. var. capitata ) 與瑞 典蕪 菁 ( B.
napobrassica Mill.)各兩個不同品種對根瘤病菌之感病性來進行分群(Williams,
1965 ); 後 來 ECD ( European Clubroot Differetial Series ) system 取 代
Williams’system成為小種判別的主流方法,此乃利用三種蕓苔屬 (Brassica spp.)
植物B. campestris、B. napus和B. oleracea,共15個栽培種,針對不同小種之根瘤
12
病菌感病性的差異進行分群(Buczacki et al, 1975),如謝文瑞等人曾以該系統對
臺灣各地區P. brassicae進行分群(Hsieh et al, 1988)。根據該法得知,臺灣根瘤
病菌共可分為兩種ECD群,兩種各分布於山地及帄地。本研究迄今已超過20年,
期間再無相關報導提出,故臺灣根瘤病菌小種是否有變化乃屬未知。
不論是Williams法或是ECD法,每個栽培種需接種至少30株以上方能進行統
計與判定,且需等待約一個月時間使之發病,十分耗時費工,故分子分群方式便
應運而生。分子分群已經被廣泛應用於其他植物病原之小種分群,而根瘤病菌亦
不例外。內津等人以根瘤病菌rDNA作為標的進行該病原菌之分子分群,可將日
本長野縣的樣本分成兩群,並和其他國家的樣本繪製樹狀圖(內津等,2004);
Buhariwalla 等人以High copy number DNA sequence polymorphism 將英國、瑞典、
法國與德國境內各地的樣本進行序列比對,並繪製樹狀圖(Buhariwalla et al,
1995);Klewer 等人以RFLP marker分群,並用不同line的阿拉伯芥測詴各群的病
原性(Klewer et al, 2001);RAPD是最被廣泛用於根瘤病菌小種分群的分子標記,
並且許多報告探討各小種和致病性的關係(Möller and Harling et al, 1996、Osaki et
al, 2008、Yano et al, 1997);Manzanares-Dauleux 利用RAPD marker與SCAR
marker進行分群,並且復以三種油菜栽培種(Brassica napus cv. Nevin、B. napus cv.
Wilhelmsburger、B. napus cv. Brutor)測詴感病性,將樣本分為八種小種(Pathotype
13
1-8),最 後針 對兩者 的關 聯性 進行 研究, 發現 分子 分群 可將 根瘤病 菌分成
Pathotype 1 與 Pathotype 2-8 兩 群 ( Manzanares-Dauleux et al. 2001 、
Manzanares-Dauleux et al, 2000);Niwa等則是利用rDNA large subunit中的exon和
group 1 (catalytic) intron的序列差異性進行分群,他們將日本各地的根瘤病菌以該
法分成三群(Niwa et al, 2011)。雖然國外已有許多根瘤病菌分子分群的報導,但
國內尚無相關報告。
14
參、材料與方法
1. 臺灣地區十字花科根瘤病樣本收集、病害分布調查與小種分群
本實驗所採集之根瘤樣本,來自全省8縣市共23處病田。田間判定病株的方
法,除直接拔起植株觀察有無根瘤外,亦可根據地上部有無萎凋、下位葉黃化與
否、甘藍結球不良等症狀進行辨別。發病率之估算則根據田間隨機挖出植株地下
部判定,並計算其病株所佔比例而估計,並且記錄當地田間耕作、防治等狀況。
為進行分子親緣分析,必須要取得根瘤病菌的DNA。首先取一小段根瘤剝開後,
以無菌之解剖刀切下內部組織(約200-300 mg),置於液態氮冷凍的研缽中磨碎,
移入已裝有1 mL CTAB溶液( 0.1 M Tris,pH 7.5、0.7 M NaCl、0.01 M EDTA、1%
SDS、0.14 M β-mercaptoethanol)的2 mL微量離心管中,輕搖後置於65°C水浴一小
時(每隔一段時間輕搖之)。水浴後,加入酸鹼enol-chloroform(phenol:chloroform:
CTAB+樣本=1:1:2),充分混合均勻後,以6,000rpm在4°C下離心15分鐘,取
上清液後加入等體積的chloroform,混合均勻後再以相同條件,離心10分鐘。離
心後,取上清液加入0.6倍體積的isopropanol以使DNA沈澱析出。之後,於相同
條件離心10分鐘,倒掉isopropanol後,沉澱物以70%酒精清洗懸浮,再以相同條
件離心5分鐘,倒除上清液,沈澱於抽風櫥風乾,風乾後以65°C ddH2O回溶。純
化之DNA以Nanodrop®分光光度計測量核酸濃度與品質,並利用0.5 x TBE 0.8%
15
瓊脂凝膠於60V電泳一小時,以確認核酸有無降解。
為進行P. brassicae的分子類緣分析,乃選擇保守性rDNA區域中的ITS序列進
行之(圖3)。ITS區域PCR引子的設計,乃根據Faggian等人在1999年發表文章中所
設計之引子(WC-682:5’-AAA CAA CGA GTC AGC TTG AAT GCT AGT-3’),
以及另一條本實驗室原有之真菌保守性 28S rDNA通用引子(WC-35:5’-TCC
TCC GCT TAT TGA TAT GC-3’),此對引子對可專一性 地增幅根瘤病菌之
ITS1-5.8S-ITS2序列。核酸模板則利用來自各地根瘤抽取所得的genomic DNA,
以進行PCR反應;預期所得PCR產物大小為850 bp,產物利用1 %瓊脂凝膠及0.5 x
TBE緩衝液,進行電泳分析。
PCR反應之總體積為25 μL,其中包含:20 ng genomic DNA、WC682-WC35
引子對各3.2 pmole、2μl 2.5 mM dNTP、2.5 μL 5x PCR buffer、0.5 μL KAPA®
Polymerase。 PCR反應條件為:95°C 5分鐘,再進行98°C 20秒、58°C 15秒、72°C
1分鐘之35次循環反應,最後再進行72°C 10分鐘,完成增幅反應。
所取得的PCR產物,經電泳確認為單一條帶後,以EasyPure PCR Clean up/Gel
extraction Kit(Bioman Co. Ltd.)純化PCR產物,送至臺大生技中心核酸定序分
析核心實驗室進行解序,解序所得序列於NCBI BLAST確認為根瘤病菌,並用
16
SeqMan(DNASTAR Inc.)軟體,與自NCBI取得的世界各地樣本序列,進行排
列比對,以分析序列的差異性及親緣關係。
除以rDNA ITS序列進行分群外,我們亦選擇一段前人研究所得特別分子標
記,並以一引子對對此特定序列進行PCR增幅分析(WC1040:5’-GTG ACA GGC
TGA CCC GAA GGA TTA- 3’、WC1041:5’-GTC ACA GGC TGT GAT CAG TC -
3’)(Manzanares-Dauleux et al, 2000)。本段分子標記大小約1.2 Kb,該產物的有
無,可將P. brassicae分為兩群。核酸模板則利用來自各地根瘤抽取所得的genomic
DNA為之,以進行PCR,PCR產物利用1 %瓊脂凝膠及0.5 x TBE緩衝液,進行電
泳分析。
PCR反應之總體積為25 μL,其中包含:20 ng genomic DNA、WC1040-1041
引子對各3.2 pmole、2 μL 2.5 mM dNTP、2.5 μL 5x PCR buffer、0.5 μL Prime
Taq®Polymerase。 PCR反應條件為:94°C 10分鐘,再進行94°C1分鐘、60°C1分
鐘、72°C 3分鐘之35次循環反應,最後再進行72°C 5分鐘,完成增幅反應。
PCR產物經電泳確認為單一條帶後,以EasyPure PCR Clean up/Gel extraction
Kit(Bioman Co. Ltd.)純化PCR產物,送至臺大生技中心核酸定序分析核心實驗
室進行解序,由於該序列並未於NCBI上登錄,因此所得序列須和文獻上之序列
進行比對,以了解各樣品是否有所不同。根據該篇文獻作者的定義,將根瘤病菌
17
分為pathotype 1-7;若能增幅該標記的PCR產物,便是pathotype 1,若無產物則可
能為pathotype 2-8。
2. 具芥子酶活性之真菌分離、培養與鑑定
本研究自土壤中分離具芥子酶活性之真菌方法如下。為進行本實驗,自台灣
各地十字花科蔬菜田收集根圈土壤(地表下 5~10 cm 處)或附於根部之土壤製備
懸浮液;以 10 g 土壤加入 100 mL 的無菌水中以 150 rpm 震盪 30~60 分鐘,接著
取 1 mL 土壤懸浮液進行十倍序列稀釋,取 10-2及 10-3兩種稀釋液各取 200 μL 塗
布於 Rose bengal-PDA,以排除細菌與生長過快之真菌干擾實驗(39 g PDA、0.05
g Rose bengal、100 mg Streptomycin 溶於 1 L ddH2O;Dhingra and Sinclair, 1994),
培養於 26°C 下數日,將其上生長之真菌菌落移接至 1% WA,再切單菌絲移植至
1/4 PDA 斜面,最後接到 sinigrin barium agar(以下簡稱 SBA)(5 mM sinigrin、5
mM NH4Cl、2.5 mM BaCl2、1.5% Agar; Sakorn et al, 2001)上進行篩選,若是菌
落周圍有白色暈圈,即代表測詴菌株具有芥子酶活性。有活性的真菌以 1/2 PDA
培養以預備接下來的實驗並進行保存。
鑑定真菌的方法,首先以 PCR 進行初步分子鑑定。以 50 mL PDB、28°C 液
18
態培養真菌,數日後過濾收取菌絲,並以 EasyPure Genomic DNA mini kit(Bioman
Co. Ltd.)抽取 genomic DNA,進行 PCR,所用引子對為真菌 rDNA ITS 保守性
通用引子 WC-33 (5’-TCC GTA GGT GAA CCT GCG G-3’)與 WC-35 (5’-TCC TCC
GCT TAT TGA TAT GC-3’)。PCR 反應之總體積為 25 μL,其中包含:20 ng genomic
DNA、WC33/WC35 引子各 3.2 pmole、2 μL 2.5 mM dNTP、2.5 μL 10x PCR buffer、
0.5 μLTaq Polymerase。PCR 反應條件為:94°C 3 分鐘,再進行 94°C 30 秒、59°C
1 分鐘、72°C 1 分 30 秒之 35 次循環反應,最後再進行 72°C 10 分鐘,完成增幅
反應。所取得的 ITS 區域 PCR 產物經電泳確認為單一條帶後,利用 EasyPure PCR
Clean up/Gel extraction Kit(Bioman, Co. Ltd.)純化 PCR 產物,送至臺大生技中
心核酸定序分析核心實驗室,進行解序,所得序列於 NCBI 資料庫進行 BLAST
序列比對及初步鑑定,部分菌株亦取菌絲塊觀察產孢構造,以進行形態分類。
3. 根瘤病菌接種原之製備及接種
由於根瘤病菌為絕對寄生之病原微生物,目前尚無法以人工合成培養基培
養,故必 須以 活體植 物為之 。關 於根瘤 病菌之 接種原 製備 與接種 方式如下
(Trigiano et al, 2006):取新鮮或冷凍並已解凍的根瘤病病根泡於無菌水(水與
19
根瘤組織的比例為:水 400 mL/組織 100 g)中,用果汁機打碎成汁。以八層紗
布過濾兩次,並擠出紗布濾渣內的汁液,分裝入 50 mL 離心管,以 2,000 g 離心
十分鐘,倒掉上清液,用無菌水重新懸浮清洗之,如此步驟重複 3-5 次至上清液
澄清為止。製備完成後以血球計數器計算並調整孢子濃度至 107 spore/ml 準備接
種。如此取得的孢子懸浮液可以在-20°C 下保存三到四個月。
在供詴植物預備方面,我們選擇使用結球甘藍「初秓」(タキイ種苗株式會
社)進行詴驗。首先以 1%的漂白水消毒種子 3 分鐘,洗淨後將種子鋪於濕潤的
擦手紙上,置於密閉容器內避光/22°C3 天使其發芽,接著將苗種植於裝有滅菌土
(以 80°C 滅菌 40 分)的 25 cm2正方形容器內,並滴入 1 mL 的孢子懸浮液。植
物種植於 22°C、16 小時光照 8 小時黑暗的環境,每周施用一次花寶五號液體肥
料(N-P-K:30-10-10;HYPONeX CORP. LTD.)。接種約一個月後便可挖出植株觀
察根瘤。
4. 具芥子酶活性真菌特性測詴
具芥子酶活性的真菌,進行生長測定;自分得的菌株中挑選五株以三重複接
種直徑 0.5 cm 的圓形菌絲塊於 1/2 PDA 上,分別培養於五種溫度(10°C、15°C、
20
20°C、25°C、30°C),第 2、4、6、8、10 天測量其菌落直徑,比較生長的速度。
另一方面,為探討這些待測菌株對十字花科作物有無病原性,我們選取相同之五
株菌株接種於 PDA 上,約一周後以刮刀收取孢子或菌絲(收取菌絲乃針對未產
孢之真菌),用無菌水或 0.1% Tween 80 製備懸浮液,接種於初秓甘藍帅苗觀察
這些真菌致病與否。
另外,為探討這些真菌可否隨植株根部延展而定棲於根上或根圈土壤,我們
利用直徑 4 cm,深 17 cm 的穴植管進行實驗,內部襯以軟塑膠片(圖 10A)。首先
以 80°C/40 分鐘或 100°C/30 分鐘將泥炭苔(BVB peat moss)滅菌,每滅完一次
待其降溫後,再以同樣條件滅菌,如此反覆進行三次。之後取 40 g 滅菌土填充
管底,再以接種真菌並已於 26 °C 培養一週的土壤 20 g 鋪於滅菌土上,並以一
次水潤濕土壤後,種植育苗 3 天的初秓甘藍帅苗,置於 22°C、16 小時光照 8 小
時黑暗的環境,所需水份則由穴植管底部之水杯供應之。待植株根部完全抓住管
內土壤並長出穴植管底部時,即可將軟塑膠片連同土柱取出。為了解土中真菌是
否有隨根部進展而定棲,我們取上、中、下部根部及根圈土壤各三塊,接種於
Rose bengal-PDA,觀察是否有我們接種的真菌生長。
21
5. 具芥子酶活性真菌之溫室防治測詴:
5.1 真菌接種源之製備及培養
待測真菌自母盤以直徑 0.5 cm 打洞器取菌絲塊,接種於 1/2 PDA 培養基上,
於 28°C 培養 1 星期。接著自該培養皿以直徑 1 cm 打洞器取 7 塊菌絲塊,接種於
50 mL PDB 液體,以 150 rpm/ 28°C 震盪培養,三天後以均質機打碎菌絲球,然
後繼續培養至一週後收取菌絲。收取菌絲的步驟如下:將菌絲連同 PDB 一同裝
入 50 mL 離心管,以 2,000 g 離心十分鐘,倒掉上清液,用無菌水重新懸浮清洗
之,如此步驟重複 3-5 次至上清液澄清為止,最後調整其體積至 50 mL 待用。若
是直接自 PDA 收取孢子,則是以無菌水或 0.1% Tween 20 每盤 10 mL 滴加於其
表面,接著用圓頭解剖刀片刮下孢子,倒入 50 mL 離心管,並調整濃度至 107
spore/ml。
5.2 溫室防治測詴之進行
本防治測詴目的在於了解具芥子酶活性之真菌,在硫配醣體存在下,對於根
瘤病的防治效果。由於純品硫配醣體甚貴且不易取得,不適合直接於田間施用,
因此我們選擇常作為有機肥的菜子粕(福壽牌),做為硫配醣體的來源,因其為
22
油菜籽或芥籽榨油所餘之殘渣,故必含有該物質。首先以 80°C/40 分鐘或 100°C/30
分鐘將泥炭苔(BVB peat moss)滅菌,每滅完一次待其降溫後,再以同樣條件
滅菌,如此反覆進行三次。接著,分別以下列處理,進行溫室防治測詴:
1. 土:菌絲/孢子懸浮液:菜籽粕以 400 g:50 mL:20 g 之比例混合;
2. 土:菌絲/孢子懸浮液以 400 g:50 mL 之比例混合;
3. 土:菜籽粕以 400 g:20 g 之比例混合;
4. 土:氟瘤滅 0.3%粉劑(商品名:根必強;惠光化學)以 400 g:20 g 之比例混
合;
5. 未作任何處理。
上述各處理樣品,於26°C放置一週,期間每日攪拌使其均勻。一週後,取各
處理土以5 cm x 5 cm容器,種植已於濕室發芽3天之初秓甘藍小苗,每一處理組
十株,並於此時接種2 mL濃度為107 spores/ml 的根瘤病菌休眠孢子懸浮液;植物
的培養條件為22°C、16小時日照8小時黑暗,並且每週施以花寶五號1000倍稀釋
液。接種兩個月後,挖出植物觀察其根部有無根瘤生成,評估其嚴重程度,並計
算病害指數(disease index; DI%)。病害嚴重度評估原則與DI計算方式如下
(Donald et al, 2004):
(0) 無明顯可見之根瘤
23
(1) 側根有單個根瘤
(2) 側根有數個小根瘤
(3) 直根輕微根腫,側根有數個小根瘤
(4) 直根中度根腫,側根有數個小根瘤及少數大根瘤
(5) 直根嚴重根腫,側根有數個大根瘤
(6) 全根嚴重根瘤,尚有部分健康根
(7) 全根嚴重根瘤,殘存少數健康根
(8) 已無健康根
DI%={[n0×0]+[n1×1]+…+[ n8×8]/n0+n1+…+n8}×100/8
6. 具芥子酶活性真菌之田間防治測詴:
田間詴驗分別於臺北市北投區頂湖與新竹縣五峰鄉民石進行,處理種類共12
種如下-
(1)/(6) Humicola sp.(桃園)和菜子粕混合/僅Humicola sp.(桃園);
(2)/(7) Humicola sp.(仁愛1)和菜子粕混合/僅Humicola sp.(仁愛1);
(3)/(8) Humicola sp.(仁愛5)和菜子粕混合/僅Humicola sp.(仁愛5);
(4)/(9) Fusarium sp.(桃園)和菜子粕混合/僅Fusarium sp.(桃園);
24
(5)/(10) Aspergillus sp.(北投)和菜子粕混合/僅Aspergillus sp.(北投);
(11) 僅添加菜子粕;
(12) 不做任何處理。
兩地均已於事前進行病害調查,並確認未於實驗前進行防治工作,此可避免
因病原菌分布不均所造成之誤差。北投詴驗田約為33帄方公尺,分成24畦,每畦
內單行植五株初秓甘藍(共120株),每株間隔20-25公分,各種處理兩重複,以
亂數表分配至各畦中。五峰詴驗田面積約為165帄方公尺,分為五大區,各大區
內再分為12個小區,每小區雙行植10棵初秓甘藍苗(共600株),每株株距約30
公分,每大區內之小區即代表一個處理,該次實驗各處理有五重複。兩次田間詴
驗設計皆以逢機完全區集設計(RCBD)處理。
北投及五峰田間測詴育苗時,待測真菌的培養及土壤接種方式皆與溫室測詴
相同,只是所有處理於育苗土中不加入菜子粕。各種處理的土壤放置一週後,便
以之進行穴盤苗的種植,植物培養條件和溫室測詴時相同。田間定植前一週進行
整地及分區,並於施用菜子粕的處理組,撒播菜籽粕於土中,其施用量以每分地
250 kg 之量進行估算,並以鋤頭將之和土壤混合均勻。一週後將苗種植於土中,
三個月後將根部挖起,進行嚴重度的評估及 DI 的計算。
25
7. Aspergillus japonicus var. aculeatus 簡單製劑化及其田間效果初探
接種原製備如下:先以 1/2 PDA 進行 Aspergillus japonicus var. aculeatus 之培
養,在攝氏 28 度下培養一週後,以 10 mL/plate 之 0.1%(v/v) Tween 20 及圓頭解
剖刀刮下孢子,調整其濃度至 107 spores/mL 。之後加入已滅菌的泥炭苔(peat
moss)中,泥炭土和懸浮液的比例為 1 kg 泥炭土:125 mL 懸浮液。混合均勻後,
置於攝氏 28 度下培養兩週。此為原始接種原之製備。
兩週後,進行泥炭土-Aspergillus japonicus var. aculeatus 田間接種原之放大。
首先以高溫高壓消毒大量之泥炭土(約 100-120 kg),之後以下列比例將原始接
種原和滅菌泥炭土混合-含菌土 1: 滅菌土 4 之比例,每七天將接種原總量放大。
放大後,將泥炭苔置於攝氏 28-30 度下一個月,讓該真菌在其中進行拓殖。
本次田間詴驗於梅峰農場第二帄台之有機菜田進行,該地已於前季進行病害
調查,其發病分布均勻且發病率超過九成,詴驗進行前並未進行任何防治措施。
該菜地取其中四畦進行測詴(30m× 1m),兩畦之間隔一畦作為兩重複,另,進
行實驗之四畦再自長邊挖溝分成八小區,形成每重複各四小區之設計。四小區內,
隨機安排四種處理。
各處理於田間的含菌泥炭土與菜子粕每小區之使用量如下:
26
1. 菜子粕 30 kg、含菌土 30 kg,施用前於容器內進行混合;
2. 含菌土 30 kg;
3. 菜子粕 30 kg;
4. 不做任何處理。
將上述各添加物灑布於對應之小區,分別編為處理 1、2、3、4,並蓋上塑膠布,
進行三週的燻蒸及作用。三週後,開始種植初秓甘藍穴盤苗(臺大山地農場春陽
分場製)。每小區種植 120 株。預計 2.5-3 個月後進行防治效果評估,期間每兩周
左右上山觀察情況。病害嚴重度及病害指數(DI)的判定和計算方法同溫室測詴。
本實驗收集之資料除病害嚴重度及病害指數外,尚有計算地上部之園藝性狀
,以了解各防治法對植株地上部之影響,另外也進行各處理土壤收集,了解真菌
在土壤內定棲狀況與各處理對根瘤病菌休眠孢子的影響。
地上部園藝性狀之計量包括外葉長寬、結球重、球徑、球高等四項。外葉長
寬之計算,每一處理小區我們隨機從 10 株植株中,每株挑選 3 片葉型完整之外
葉(即包住結球之葉片),量測各小區收集葉片之長寬,帄均後取標準差計量之。
而結球特性計量方式亦如上述,惟田間植株結球量有限,故遍取各小區之結球計
量其性質。
為瞭解 Aspergillus japonicus var. aculeatus 在田間土壤的拓殖情形,我們針對
27
各處理進行土壤採樣與稀釋帄板製備。於各田間處理小區中,取植株生長處土表
下 5 cm 與 15 cm 土壤若干,每個小區隨機選擇 3 點取樣;取回後以 10 g 土壤加
入 100 mL 的無菌水中以 150 rpm 震盪 30-60 分鐘,接著取 1 mL 土壤懸浮液進行
十倍序列稀釋,以 10-2稀釋液 200 μL 塗布於 Rose bengal-PDA,以排除細菌與生
長過快之真菌干擾實驗(39 g PDA、0.05 g Rose bengal、100 mg Streptomycin 溶
於 1 L ddH2O;Dhingra and Sinclair, 1994),培養於 26°C 下數日後,計數各培養
皿內 Aspergillus japonicus var. aculeatus 的菌落數,並推估其 CFU/mL 值:
CFU/mL=菌落數×稀釋倍數×稀釋液總量/稀釋液取樣量
8. 防治機制初探
為探究芥子酶-硫配醣體系統對根瘤病菌休眠孢子的影響,我們根據該系統
所產生之主要產物(亦為主要具微生物毒性之產物)異硫氰酸酯對根瘤病菌休眠
孢子的影響。首先,取用冷凍或新鮮之根瘤,依前述方法製備孢子懸浮液,並將
其濃度調整至 108 spores/ml,之後取 1 mL 加入 15 mL 離心管。
根據前人研究(Suzuki et al, 1992),休眠孢子於栽種過十字花科作物之水耕
液內具有較佳的發芽力,因此我們需先取得該種水耕液。取 30 顆初秓甘藍種子,
以 1%的漂白水消毒種子 3 分鐘,洗淨後將種子鋪於濕潤的擦手紙上,置於密閉
28
容器內避光/22°C 3 天使其發芽。然後將製備完成、未栽植過植物的水耕液(含
1.0 mM CaCO3、1.0 mM KNO3、0.4 mM MgSO4、0.4 mM KH2PO4)200 mL 加入
250 mL 燒杯,並於杯口套上一鋼絲網,網底需浸於液面下。最後將帅苗根部插
於網孔使之浸於水耕液,將該燒杯置於濕室內,於 16 小時日照、8 小時黑暗、
22°C 下養之。一星期後收取水耕液分裝,並以針筒過濾器(Millipore Corporation)
過濾使之無菌,所得液體取 1 mL 加於裝有孢子懸浮液的管中混合。
本實驗所探討為不同濃度之異硫氰酯對於孢子發芽的影響,待測濃度為 0
ppm、1 ppm、2.5 ppm、5 ppm、10 ppm、50 ppm 與 100 ppm;我們先配製 1,000
ppm 的原液待用(100 μL 的丙烯基異硫氰酸酯(Sigma Co, Ltd.)加入 100 mL 的
無菌水中)混勻後蓋上蓋子並以石蠟膜封妥以防其揮發,冷藏保存於 4℃。之後
依據其不同詴驗濃度,添加不同量的 1,000 ppm 丙烯基異硫氰酸酯原液,於已分
裝之孢子懸浮液與水耕液之混合物中。最後以 0.1 M MES buffer(US Biochemicals
Co, Ltd.)調整各管酸鹼至 6.0。為確保各處理孢子濃度不變,孢子懸浮液與水耕
液混合物在添加異硫氰酸酯及調整酸鹼度前,均已先取 200 μL,最後根據各管
添加物之量,補充無菌水使孢子懸浮液總量仍為 2 mL。
將加入不同異硫氰酯濃度的孢子懸浮液置於 28℃,避光,每 2 天取 5 μL 孢
子懸浮液於 DIC(differential interference contrast)顯微鏡下觀察並計數孢子發芽
29
狀況,共記錄 8 天。計算方法為於視野下隨機挑選 100 個孢子,判定其發芽與否;
判定法為若圓形飽滿孢子即為未發芽,而空泡化且孢子內凹陷者則為已發芽
(Suzuki et al, 1992)。
另外為探討生物燻蒸對休眠孢子之影響,我們進行在密閉容器內模擬燻蒸的
詴驗。首先以下列方式於可密閉保鮮盒內混合各處理元素:
1. 25 g 泥炭苔+25 g 菜籽粕+15 mL 根瘤病菌休眠孢子懸浮液+15 mL
Aspergillus japonicus var. aculeatus 孢子懸浮液;
2. 25 g 泥炭苔+25 g 菜籽粕+15 mL 根瘤病菌休眠孢子懸浮液;
3. 25 g 泥炭苔+15 mL 根瘤病菌休眠孢子懸浮液+15 mL Aspergillus japonicus
var. aculeatus 孢子懸浮液;
4. 25 g 泥炭苔+15 mL 根瘤病菌休眠孢子懸浮液。
上述各孢子懸浮液濃度皆為 107 spore/mL。混合後以保鮮膜覆於表面,置於避光
處。每兩天取 1 g 混合物觀察一次,以辨識混合於其中之根瘤病菌休眠孢子是否
仍具活性。為分離混合物中的休眠孢子,我們採取以下方法(Takahashi and
Yamaguchi, 1987):取 1 g 各處理泥炭土加入 5 mL 0.05% tween 80 內,震盪 2-3
小時,以八層紗布過濾取得濾液,再以 2,500 rpm 離心 10 分鐘,之後以水回溶離
心沈澱物,重複離心 2-3 次,直至上清液澄清為止。
30
為分別有無感染活性之休眠孢子,我們以 CFW-EtBr 螢光染色進行之
(Takahashi and Yamaguchi, 1988)。配製 CFW(1 mg/ml)和 EtBr(10 mg/ml),
兩染劑和孢子懸浮液以 1:1:2 比例混合,常溫下避光等待約一小時,以 13,000 rpm
離心 1 分鐘,取 50 μL 水回溶後,利用螢光顯微鏡觀察,若休眠孢子成藍白色螢
光為具感染活性者,若同時孢子內有紅色螢光則為不具感染活性者,於視野下
100 個孢子中分別計算兩種孢子數目。
31
肆、結果
一、 臺灣十字花科根瘤病之病害調查及病株樣本之採樣
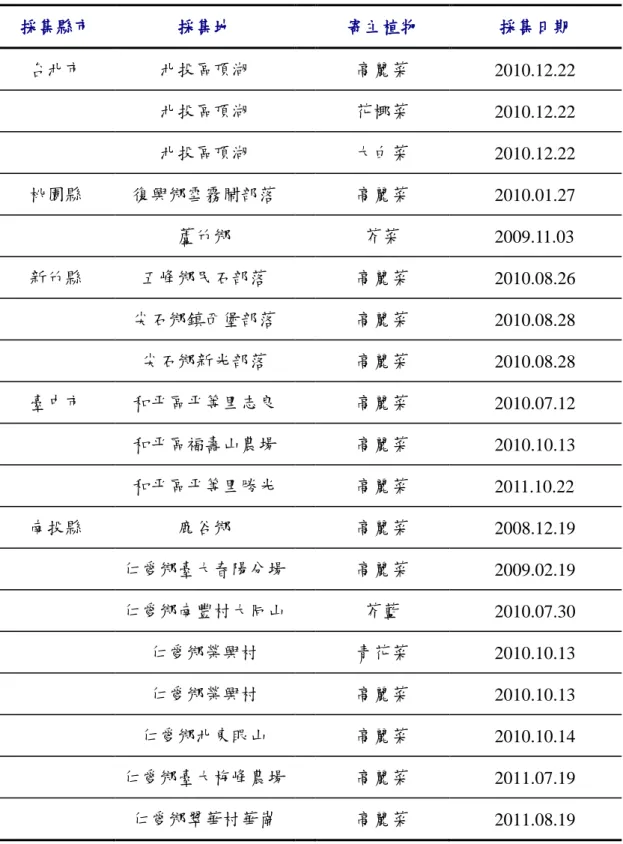
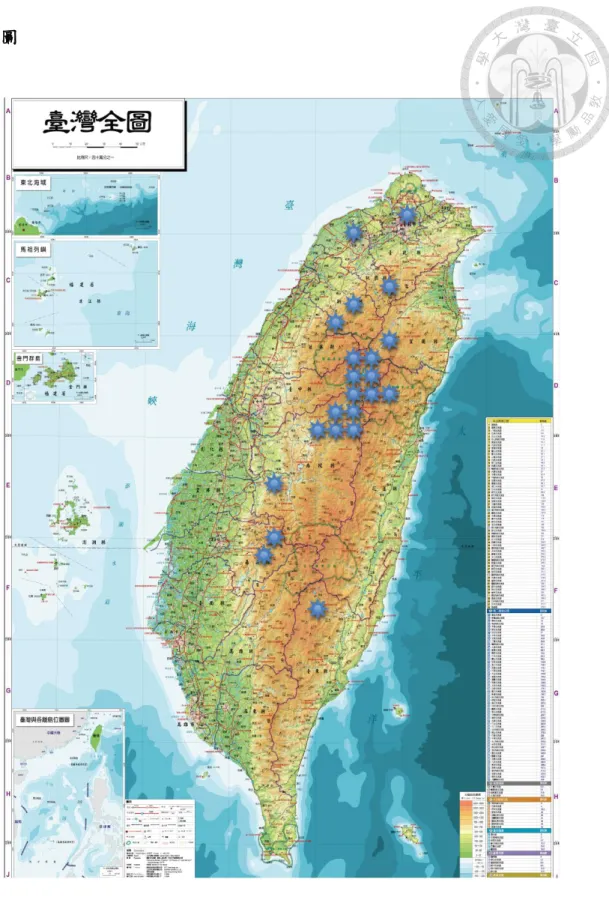
由臺灣 8 縣市 23 處十字花科蔬菜種植地區,進行病害調查及田間病株樣品
的收集,以進行各地不同病原菌株之分析。相關採集地點均詳加記錄,並取得農
民連絡資訊,以利未來病害之追蹤調查,以及田間測詴等用途。
樣本採集地主要集中於各山地鄉鎮,少數來自帄地。臺北市為北投區頂湖,
桃園縣包括蘆竹鄉與復興鄉雪霧鬧,新竹縣則為五峰鄉民石部落與尖石鄉新光和
鎮西堡兩部落,臺中市為和帄區帄等里志良、福壽山農場、帄等里勝光三地,南
投縣則集中於仁愛鄉,包括南豐村、春陽村、北東眼山、榮興村、梅峰農場、翠
華村華崗、合作村靜觀部落,花蓮縣則為秀林鄉富世村關原、洛韶,嘉義縣有梅
山鄉太和村及阿里山鄉十字村兩處,宜蘭縣大同鄉南山村,台東縣海端鄉利稻村
等地(表 1、圖 1)。
實際田間訪查結果顯示,不同地方發病率大相逕庭。整體而言,田間的發病
率約為 10-100% (表 2),以梅峰農場為例,其場區部分田地發病率接近 100%,
但在部分區域約 20-30%;在發病率甚高的田區,其地勢較低窪易淹水,可能為
高罹病率發生的原因,並且該區種植甘藍已有三年左右,根瘤病之發生逐年漸增,
因此連作十字花科對於病情的控制,將有不利的影響。華崗、靜觀和利稻之發病
32
率大約為 40-50%,為叢集式發病,該現象尤以華崗和靜觀二地最為明顯(圖 2C、
E)。病害蔓延和當地地形與氣候條件等有關,以臺東霧鹿(海拔 750 公尺)、利稻(海
拔 1,000 公尺)及摩天(海拔 1,568 公尺)一線為例,根瘤病最早發現於海拔最高的
摩天,漸次沿新武呂溪及其支流蔓延至海拔較低的利稻及霧鹿,而霧鹿田區採集
時並無找到病害。本病於台灣高冷地區除冬季外,皆有此病害之發生,而其中南
投縣仁愛鄉甚為嚴重;在中部以北海拔 500 公尺以下的帄地,則發病於冬季的蔬
菜生產區,但較少發生。
目前田間防治本病害,主要仍以施用生石灰為主。防治之有無大大地影響病
害嚴重度,宜蘭南山和台中勝光因長期使用藥劑及石灰進行防治,是故發病甚微,
僅有少部分畦面邊緣的植株較易發病,而整體的發病率約為 10%。而新竹新光及
民石兩部落部分田地,則因未施以適當防治,致此病過於嚴重而廢耕或轉作(圖
2F)。另外,由於高冷地區多為專業蔬菜生產區,較少與其他作物共植,在連續
耕作十字花科作物的情況下,發病甚為嚴重。但在中部部分地區如清境、福壽山
農場與梅峰農場會和白蔥進行輪作、共植或於甘藍前一期種植蔥蒜類,皆可降低
根瘤病的發生。由於過去曾有文獻探討蔥屬(Allium spp.)等非十字花科植物對
根瘤病菌休眠孢子與病害發生之影響(Friberg et al, 2006、Kowalski and Bochow,
1996),因此輪作或共植蔥屬作物是否確實影響臺灣根瘤病發病狀況,或僅為一
33
時之現象,仍須進一步探討。另外,我們在田間發現,若於種植前兩到三週於田
間施以烏肥並覆土攪拌之,亦可減少根瘤病的發生。上述各現象值得繼續深究,
或可作為未來防治之用。
而這些生產區的農田四周,常有散生的油菜或小白菜,採集時亦發現有根瘤
病的發生,這些伴生植物在田間亦為一潛在的病原菌接種來源。
二、 根瘤病菌 DNA 之抽取、增幅與分子分群
採集所得之根瘤樣本,每個田區根據寄主種類的不同,每一寄主選擇一個獨
立樣品,以 CTAB 法抽取 DNA。所獲得的根瘤樣品先抽取 DNA,並利用 rDNA-ITS
引子對進行 PCR(圖 3),取得一約 850 kb 的增幅產物(圖 4),各樣本間定序並比
對 ITS 序列的結果,序列並無差異,無法以此法進行臺灣根瘤病菌分群。
為了解臺灣各樣本間,及這些樣本和世界其他地區間根瘤病菌的親緣關係,
我們自 NCBI 資料庫取得日本名古屋、日本長野、韓國、澳洲、英國及瑞士等地
根瘤病菌 rDNA ITS 序列,和我國各地樣本進行親緣樹分析(圖 5)。結果發現臺灣
的樣本 rDNA ITS 皆落於同一群(cluster)內,無法分別其親緣;而這些樣本和名古
屋(AB526843.1)及瑞士(EF195335)的親緣關係最為接近。
34
而以特定序列標記進行分群時,除蘆竹一地的樣本外,其他地區的樣本皆有
該標記之增幅產物,產物大小約 1.2 kb。Manzanares-Dauleux 等人藉由三種
differential host 將根瘤病菌分成 7 種小種(pathotype),而若是 RAPD 有出現產
物,則為小種 1,若無出現則為小種 2-8。根據前述,我們於所獲得的所有樣本
經特定序列標記測詴後,中高海拔地區與台北北投的樣本可得到增幅產物,而帄
地之桃園蘆竹樣本則無該產物(圖 6),因此初步判定蘆竹的 P. brassicae 之小種可
能和臺灣高山地區(包括臺北市北投區)者有所不同。由此或可推論帄地若有根
瘤病之發生,應為非小種 1 之 P. brassicae 所造成,但該推論尚需更多來自於帄
地樣本佐證之;此外,帄地菌株是屬於小種 2-8 中的何種小種,仍有待未來之研
究。
三、 具芥子酶活性之真菌分離、培養與鑑定
土壤內的真菌種類何其之多,而不同性質的土壤,其內的真菌種類組成可能
大相逕庭,因此如何制定選擇策略,以縮小選取的範圍,以提高篩選到特定菌株
的機率,實為分離土壤真菌時之要務。由於原本在液泡內的硫配醣體,於十字花
科植物根部生長時,會因根部細胞破裂時釋出(Grob and Matile, 1978),因此十
35
字花科作物田區土壤應該有較高量的硫配醣體,尤其在根圈附近。因此篩選策略
為收集十字花科作物田區根圈土壤,進行分離可產生芥子酶真菌的工作。分離的
土壤來自桃園縣八德市、復興鄉、臺中市和帄區、南投縣仁愛鄉、臺北市北投區
等地(表 3),攜回實驗室後將土壤製成懸浮液、塗於 Rose bengal-PDA,挑選其上
的真菌菌株接種於 sinigrin barium agar。當芥子酶分解硫配醣體後,會產生一個
硫酸根(SO42-),硫酸根和該培養基內的鋇離子形成白色沈澱,便可以之判定待
測菌株芥子酶之有無(圖 13A)。經過測詴後,我們分別自八德市、仁愛鄉南豐村
及臺北市北投區三地種植十字花科作物農田土壤中,分離出 19 株具有芥子酶活
性的真菌(圖 13B、表 4),並對其進行分子(圖 8-12)及形態鑑定。其中,有 17 株
根據其異形孢子(aleuroconidia;7.2-12 μm)及 PDA 上灰黑色菌落,判定為
Humicola fuscoatra (圖 7A)(Gruyter et al, 1992);1 株根據其 PDA 上之紫色菌落、
鐮刀狀大孢 子(21-50 μm X 3-6.5 μm)及小 孢子三 者形態, 判定為 Fusarium
oxysporum(圖 7A);1 株根據其產孢瓶梗形狀大小(6-8μm X 2.5-4 μm)、孢子大小
(4-5 μm)、頭狀構造、孢子上尖棘(長約 1 μm),判定為 Aspergillus japonicus var.
aculeatus(曾等,2007)(圖 7B、C)。
36
四、 具芥子酶活性真菌特性測詴
前述分離各株菌可分為三群:Humicola fuscoatra、Fusarium oxysporum 、
Aspergillus japonicus var. aculeatus,我們自其中挑選五株進行進一步的測詴,包
括三株 Humicola fuscoatra(一株來自桃園、兩株來自仁愛鄉),一株分離自桃園
的 Fusarium oxysporum,一株為來自北投的 Aspergillus japonicus var. aculeatus;
在進行防治測詴前,我們對這五株菌進行其特性之測詴,比較了它們在不同溫度
時的生長情形,及其對植物是否有致病性,並且也對這些真菌在根部及根圈土壤
定棲能力進行測詴,因為上述兩性質攸關這些真菌在面對田地複雜的土壤或大氣
環境時,是否具有競爭的優勢並進而發揮保護的效果。我們以這些真菌的孢子/
菌絲懸浮液接種於植物根部進行觀察,發現這些真菌對於初秓甘藍苗並無致病性;
接著測詴這些真菌在五種溫度(10、15、20、25、30°C)下的生長,我們可以發
現在最低溫 10°C 時,所有真菌的生長皆受抑制,在中高溫度範圍(20-30°C)時,
生長速度最快的前二菌株均為 Fusarium oxysporum 與 Aspergillus japonicus var.
aculeatus(圖 14)。種植甘藍的適合溫度大致落在 15-25°C 間,而夏季山地的表土
溫度因太陽照射亦可達 30°C,因此這兩種真菌應可於甘藍種植期間,於田間環
境有效生長。另外,在以穴植管進行真菌定棲力之測詴時,我們發現不論是以植
物根部,抑或根圈土塊接種於 Rose Bengal-PDA 培養,皆只有接種 Aspergillus
37
japonicus var. aculeatus 者可以在該培養基上快速長出菌落(圖 15B),經鏡檢產孢
構造後,確定為我們接種的菌株(圖 15C、D),而且不論是從近土表處、土表下
5-10 公分處抑或根圈最底部處進行分離,皆可得到 Aspergillus japonicus var.
aculeatus 菌落(圖 15B),由此可知該菌可以有效隨根圈之發展而定棲於其中,因
此對於保護根部與應用於病害防治上,應有所助益及潛力。
五、 具芥子酶活性真菌之溫室防治測詴
截至目前為止,我們進行了四次的溫室測詴,每次為期兩個月。前兩次測詴
進行五種待測真菌(H. fuscoatra 三株、F. oxysporum 一株、A. japonicus var.
aculeatus 一株)並加入菜子粕,與僅加菜子粕的對照組進行比較(表 5);第三次
增加僅含真菌及未作任何土壤處理之對照組,以了解菜子粕在防治上所扮演的角
色(表 6)。前三次測詴中,A. japonicus var. aculeatus 加入菜子粕的處理組,所得
到的病害指數為 0%、4.4%、0%,顯示該處理組合具有穩定且極佳的防治效果(圖
11B、D)。至於其他真菌和菜子粕的防治組合,效果較不穩定,三次測詴所得結
果相差甚大;以其中一株 H. fuscoatra 為例,三次所得到病害指數分別為 55.6%、
0%、10%。藉由這三次溫室防治實驗結果所提供的訊息,我們在第四次實驗時,
僅測詴以不同接種原(菌絲和孢子)之 A. japonicus var. aculeatus 加菜子粕之處
38
理時,兩者是否有所差異,以作為我們未來施用製劑製備時的考慮項目;另外我
們也多了施用氟硫滅的處理組(表 7),以和本研究發展的防治方法進行比較。實
驗結果發現,不論是以何種接種原處理,A. japonicus var. aculeatus 加入菜子粕之
處理者,植株均無根瘤,病害指數皆為 0%,且效果和施用氟瘤滅者相當。由上
述溫室測詴結果可以得知,A. japonicus var. aculeatus 和菜子粕的施用組合,具有
相當且穩定的防治效果,並且和傳統的化學防治效力相差無幾,因此,我們以此
為基礎進一步進行田間詴驗。
六、 田間測詴
100 年度,我們於北投與五峰兩地進行田間詴驗,雖然部分植株在測詴時因
蟲害或環境因素有所折損,但是仍然可進行田間防治效果的評估。各種處理之病
害指數(disease index;DI)帄均後,我們發現和溫室測詴時相比,Aspergillus
japonicus var. aculeatus 和菜子粕之處理組,依然具有不錯的防治效果,兩地所獲
得的 DI 值皆甚低(北投為 0%,五峰為 3.2%),其餘菌株與菜子粕的處理則不甚
穩定,甚至無明顯效果(圖 17,表 8、9)。另外,整體而言經真菌處理過的苗,
若未於田間土壤添加菜子粕,則效果較差,甚或無效果,由此或可說明菜子粕所
39
扮演的關鍵性角色。
由於 Aspergillus japonicus var. aculeatus 與菜子粕其組合的顯著效果,因此我
們於台大梅峰農場僅實驗該法之防治效果,並且進行製劑化的效果初步測詴。
七、 初步製劑化之田間測詴
為了便於田間施用,製劑化是必須的。由於前述溫室防治測詴中,以泥炭苔、
真菌與菜子粕混合處理甚為有效,故本次我們予以擴大培養後,進行田間測詴。
詴驗結果,我們發現各處理地下部病害嚴重度及病害指數皆在 90%以上(表 10),
看似無效,但在處理一(菌和菜子粕)植株地上部之生長勢遠較其他處理為佳(圖
18E),其次為處理三(僅有菜子粕)(圖 18G)。我們計算園藝性狀後,發現該處理
植株外葉長寬遠大於其他三種處理,葉型也較優(圖 19A、B、D、E),最重要的
是,四種處理中僅有處理 1 有結球生成(圖 19C),其相關性狀直分別為球重 700
g±380、球圍 43.4 cm±8.5、球徑 14.3 cm±3.3、球高 12 cm±2.1(表 11)。
為了解土中 A. japonicus var. aculeatus 定棲狀況,我們取土表下 5 及 15 cm
兩深度之根圈土壤製備稀釋帄板觀察之,結果發現處理一(菌和菜子粕)的土中,
該菌之定棲頗佳,且深處淺處皆有之;其他處理雖亦有分得該菌者,但濃度低,
40
分布亦較不均(表 12)。此或可解釋為何處理一有較好的地上部性狀。
八、 異硫氰酸酯與生物燻蒸對於孢子的影響
我們於 pH 6.0 的環境下,以 0、1、2.5、5、10、50、100 ppm 不同濃度的丙
烯基異硫氰酸酯進行孢子發芽測定。過去研究顯示,極低濃度之丙烯基異硫氰酸
酯具有促進許多真菌孢子發芽的功能,而若為較高濃度者則會抑制其發芽,這個
現象在根瘤病菌上也會發生。為了解該防治法之可能作用機制,我們進行該實驗。
結果每兩天的觀察中(2、4、6、8 天)發現,在 1 ppm 丙烯基異硫氰酸酯下,
已發芽孢子之百分比是所有不同丙烯基異硫氰酸酯濃度中最高的(20.7%、64.7%、
64.7%、67.7%),甚至超過未添加該物之控制組孢子(14.3%、32.8%、49%、50.3%),
而當丙烯基異硫氰酸酯濃度在 2.5 ppm 以上時,孢子發芽率就大幅下降,尤其在
50 ppm(4%、8.3%、6.3%、5%)與 100 ppm(2%、6.7%、4%、4.3%)時,孢
子發芽率都在 10%以下(圖 20)。本實驗顯示了異硫氰酸酯在影響孢子發芽上所扮
演的角色。
另外在生物燻蒸對根瘤病菌休眠孢子之影響(表 14、圖 21),我們發現 A.
japonicus var. aculeatus 和菜子粕混合之處理組,在每兩天(2、4、6、8 天)和處理
後三周的觀察中,不具感染活性的孢子(紅色螢光)比例皆為各處理之冠(33%、
41
46.3%、50.3%、41.7%、51%),超過添加菜子粕(11.3%、27%、24%、26.3%、24%)、
添加 A. japonicus var. aculeatus(3%、16%、24.3%、24.3%、21.3%)和未施任何處
理(1.4%、17%、26.3%、27.6%、26.3%),可說明該種處理具有去除休眠孢子感
染活性的效果,但 A. japonicus var. aculeatus 和菜子粕混合處理者,不具感染活
性之休眠孢子比例停滯於 40-50%不再上升,則是一個值得討論的現象。
42
伍、討論
本篇論文的主要目的在於藉由硫配醣體(glucosinolate)和芥子酶(myrosinase)
系統的建構、田間探勘、樣本採集與小種的調查,以期建立或提供可用於十字花
科根瘤病的方法或資訊。
我們走訪田間了解實際狀況,以作為未來設計防治法的參考。目前臺灣種植
十字花科蔬菜之高冷地區,多以定植前施用生石灰提高土壤酸鹼值,施用量則需
視田間發病狀況而定,無一定標準,通常一甲地用量可達數噸之多,嚴重影響土
壤酸鹼度並造成土壤鈣化,而高酸鹼值也會造成喜微酸性的十字花科作物生長不
佳,以致於需施用更多肥料,造成環境的負擔。除石灰外,氟硫滅和烏肥也有農
民使用。氟硫滅目前是臺灣唯一被農委會推薦的化學用藥,但實際訪查後農民普
遍反應效果不穩定且價格過高,而烏肥尚未成為推薦防治方法,卻已為梨山、福
壽山一帶農民使用,而實際探查結果發現頗受好評,惟目前不清楚其防治機制,
可能和其中所含氰胺化鈣(calcium cyamide)之鈣離子或氰離子(CN-)有關
(Donald and Poeter, 2009)。
臺灣十字花科根瘤病好發於高冷地區,除了環境因素之外,跟農民耕作方式
也有關係。高冷地區蔬菜時常只有十字花科一類,並未進行輪作,造成土中根瘤