國立臺灣大學生命科學院生化科技學系 碩士論文
Department of Biochemical Science and Technology College of Life Science
National Taiwan University Master Thesis
以重組大腸桿菌外泌生產耐熱型漆酶
Secretory production of thermostable laccase in recombinant Escherichia coli
邱柏凱 Bo-Kai Chiu
指導教授:李昆達 博士 Advisor: Kung-Ta Lee, Ph.D.
中華民國 105 年 7 月
July 2016
口試委員會審定書
謝誌
時光飛逝,在醱酵實驗室的研究也將隨本論文的完成告一段落,感謝李昆達老 師兩年多來在研究上給予的指導與支持,以及蘇遠志老師、劉俊民老師、陳俊任老 師及劉裕國老師對本論文的斧正與編修、研究的各項建議;也謝謝劉啟德老師在 Lab meeting 中給予實用的建議與肯定。
關於醱酵實驗室的夥伴們,感謝敬岳學長從實驗規劃、執行及操作,與論文編 修的各項建議,口試的安排及統籌,平日對實驗室成員的照顧與關心,到天馬行空 談及醱酵產業的過往及未來展望,都讓我得以充滿信心進行研究,也受益良多;謝 謝顥哥、派和宜宏,針對諸多時事與政策,及各種新奇事物的討論與批判,充分體 現了新時代的思維與對未來國家社會的期望,這都將成為我在學術領域外,開拓不 同視野的思考方向與養分;感謝慧嫻學姐及劉啟德老師實驗室的成員們(筱涵、凱 軍、書華、曉玲、孟薇和淑君)在研究挫折時給予的協助和鼓勵,並提供許多實驗 的想法與建議。此外謝謝科錦大廚及宥丞豐富了實驗室每個下午的咖啡香與不定 期的伙食,以及帶來各種新奇的事物與體驗;此外,除了建勳在實驗時的柔情歌聲 依然在耳,也謝謝柏谷、振倫、雨頡和競文這些學弟們輪流守夜看顧醱酵槽的辛勞 付出。
感謝父母親的支持,讓我得以無慮地進行研究並完成學業。在臺大這深具歷史 內涵又充滿活力;富含學術底蘊又不乏思維衝擊的殿堂,似有淘不盡的寶藏。接近 十年的歲月中,雖然也曾難過、苦惱與寂寞,但走過這一遭,收穫良多。謝謝醱酵 實驗室的包容,讓我經歷了在臺大最精彩、最豐富並充滿歡樂的學習之旅,祝福所 有曾經幫助過我的人,願你們一切平安喜樂。然而研究僅是開端,新的人生旅程啟 航在即,邁出校門,迎向前方諸多挑戰,也正是貢獻所學、回饋社會的開始。
柏凱 2016.08.10 於醱酵學實驗室
摘要
漆酶 (EC 1.10.3.2) 是自然界廣泛存在的一種藍色多銅氧化酵素,可氧化分解 木質素及包括雙酚A (bisphenol A)、鄰苯二酚 (ortho-diphenol)、對苯二酚 (para- diphenol)、胺基苯酚 (aminophenol) 等多種酚類化合物物質,同時將氧氣還原為水,
常應用於紙漿工業、工業廢水處理及生質能源開發等。本實驗室先前自納豆菌 Bacillus subtilis natto NTU18中選殖漆酶基因cotA,並轉殖至大腸桿菌表現,發現重 組納豆菌漆酶的熱穩定性較市售雲芝漆酶為佳。為進一步優化重組大腸桿菌生產 納豆菌漆酶之效能與可調控性,減低工業化生產及純化程序成本,本研究利用訊號 序列修飾目標基因,及調整胞膜間區 (periplasmic space) 釋放條件等方式,探討重 組大腸桿菌外泌生產納豆菌漆酶的可行性。經實驗發現,納豆菌漆酶毋需修飾訊號 序列,即能自然轉移至胞膜間區。此外,藉由菌體培養過程中添加 2% glycine及 Triton X-100,並採「誘導」與「釋放」分開處理的方式,可有效提升納豆菌漆酶釋 放至培養基的效率,使用Hinton’s flask振盪培養可生產納豆菌漆酶活性達1.06±
0.007 U/mL ,再配合細胞高密度培養技術,經20小時培養後於胞外釋放之納豆菌 漆酶活性可達5.72±0.38 U/mL,較原先胞內表現並破菌的方式所得活性提高近170 倍,有利於工業化生產效能之提升。
關鍵字:漆酶、納豆菌、大腸桿菌、外泌生產
Abstract
Laccase (EC 1.10.3.2) is one of the blue multicopper oxidases that catalyze the oxidation of lignin and many aromatic substrates including bisphenol A, ortho-diphenol, para-diphenol, and aminophenol with reduction of oxygen to water. It has many applications such as dye removal in textile, industrial wastewater treatment, and biofuel development. The laccase gene cotA was cloned from Bacillus subtilis natto NTU18, and laccase CotA was produced in Escherichia coli expression system by our lab. In previous study, the higher optimum reaction temperature indicated that recombinant CotA was more themostable than commercial fungal (Trametes versicolor) laccase. In this study, 5 kinds of N-terminal and C-terminal signal peptides were added to cotA gene sequence to enhance the translocation of target protein, and the periplasmic protein was released with chemical method to upgrade the efficiency and controllability of recombinant CotA laccase production in E. coli. The results showed that CotA laccase can be transferred to periplasmic space without any signal peptide addition. And using 2% glycine and Triton X-100 after induction would increase CotA laccase release to the culture medium. The released laccase activity reached 1.06±0.007 U/mL in Hinton’s flask, and 5.72±0.38 U/mL in bioreactor with high cell-density culture for 20 hours, which was 170-fold to intracellular production.
Keywords: Laccase, Bacillus subtilis natto, Escherichia coli, Extracellular production
縮寫表
ABTS 2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)
bp Base pair
CFU Colony forming unit
DCW Dry cell weight
DNS Dinitrosalicylic acid
DO Dissolved oxygen
IPTG Isopropyl β-D-1-thiogalactopyranoside
kDa Kilodaltons
LB Luria-Bertani broth
OD Optical density
PCR Polymerase chain reaction
rpm Rotation per minute
SDS-PAGE Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
sp Signal peptide
vvm Volume of air per volume of medium per minute
中英文對照表
Base pair 鹼基對
CFU 菌落形成單位
DCW 細胞乾重
DO 溶氧量
Hinton’s flask Hinton 氏錐形瓶
OD 吸光值
PCR 聚合酶連鎖反應
Plasmid 質體
Primer 引子
Promoter 啟動子
Rotation per minute 每分鐘轉速
Signal peptide 訊號序列
Yeast extract 酵母萃取物
目錄
口試委員會審定書 ... I 謝誌 ... II 摘要 ... III Abstract ... IV 縮寫表 ... V 中英文對照表 ... VI 目錄 ... VII 圖表目錄 ... IX
第1 章 前言 ... 1
1.1. 大腸桿菌 (Escherichia coli) 外泌系統 ... 1
1.1.1. 第一型外泌系統 (Type I secretion system) ... 1
1.1.2. 第二型外泌系統 (Type II secretion system) ... 2
1.2. 異源蛋白質的表現與外泌 ... 2
1.3. 漆酶 (laccase) ... 4
1.3.1. 起源與功能 ... 4
1.3.2. 結構 ... 5
1.3.3. 應用 ... 5
1.4. 漆酶的異源表現 ... 6
1.5. 納豆菌 ... 7
1.6. 研究目的 ... 7
1.7. 研究大綱 ... 8
第2 章 材料與方法 ... 9
2.1. 菌株及基因載體 ... 9
2.1.1. 熱休克 (heat shock) 細胞轉形法 ... 10
2.2. 含訊號序列之納豆菌漆酶重組基因與質體建構 ... 10
2.2.1. 納豆菌漆酶基因 cotA... 10
2.2.2. 訊號序列 (signal peptide, sp) ... 10
2.2.3. E. coli M15/pQE-30 Xa-cotA-hlyA 表現系統建構 ... 12
2.2.4. E. coli M15/pQE-30 Xa-(Type II signal peptide)-cotA 質體建構 .... 12
2.2.5. E. coli BL21 (DE3)/pEXP5-CT/TOPO®-(Type II signal peptide)-cotA 質體建構 ... 13
2.3. 重組大腸桿菌培養與誘導異源漆酶表現 ... 13
2.3.2. 5 L 醱酵槽培養 ... 13
2.4. 冷滲透休克法 (Cold osmotic shock) ... 15
2.5. Glycine /Triton X-100 釋放法 ... 15
2.6. 漆酶活性測定 ... 16
2.7. SDS-PAGE 電泳與活性染色分析 ... 16
第3 章 結果 ... 18
3.1. 納豆菌漆酶 (CotA laccase) 於 E. coli M15/pQE-30 Xa 系統之表現... 18
3.1.1. 含訊號序列 (signal peptide) 之納豆菌漆酶的建構與表現 ... 18
3.1.2. 納豆菌漆酶在胞膜間區 (periplasmic space) 之累積 ... 18
3.2. 納豆菌漆酶於 E. coli BL21 (DE3)/pEXP-5CT/TOPO®系統之表現 ... 19
3.2.1. 含訊號序列納豆菌漆酶的建構與表現 ... 19
3.2.2. 納豆菌漆酶在胞膜間區之累積 ... 19
3.2.3. 胞內納豆菌漆酶的釋放 ... 19
3.3. 漆酶釋放條件的最適化 ... 20
3.3.1. 最適誘導期 ... 21
3.3.2. 最適釋放期 ... 21
3.4. 以細胞高密度培養策略外泌生產納豆菌漆酶 ... 22
第4 章 討論 ... 23
4.1. 訊號序列對 CotA laccase 生產與外泌的影響 ... 23
4.1.1. 訊號序列效能評估 ... 23
4.1.2. CotA laccase 在胞膜間區 (periplasmic space) 的累積 ... 23
4.2. CotA laccase 的釋放策略 ... 24
4.2.1. 釋放策略的比較 ... 24
4.2.2. 誘導與釋放策略的調整 ... 24
4.3. 細胞高密度培養的成效與策進 ... 26
4.3.1. 細胞高密度培養對 CotA laccase 外泌生產的提升 ... 26
4.3.2. 未來策進方向 ... 26
4.4. 結論 ... 27
圖表 ... 28
參考文獻 ... 37
附錄 ... 40
圖表目錄
表 1 聚合酶連鎖反應引子 ... 28 表 2 在 E. coli M15/pQE-30 Xa 系統中,修飾不同訊號序列之 cotA laccase 表現情
形 ... 29 圖 1 在 E. coli M15/pQE 30-Xa 系統修飾不同訊號序列,對納豆菌漆酶於胞膜間
區累積的影響 ... 30 圖 2 在 E. coli BL21 (DE3)/pEXP-5CT/TOPO® 系統中,IPTG 濃度及溫度等誘導
因子,對含不同訊號序列之納豆菌漆酶,於胞膜間區累積的影響 ... 31 圖 3 在 E. coli BL21 (DE3)/pEXP-5CT/TOPO®-C (TOPO-C) 誘導培養時,同時添
加不同濃度Glycine 或 Triton X-100,對於納豆菌漆酶釋放至胞外的影響 .. 32 圖 4 SDS-PAGE 膠體電泳及活性染色分析在 E. coli BL21 (DE3)/ pEXP-
5CT/TOPO®-C (TOPO-C),胞外釋放 CotA laccase 情形 ... 33 圖 5 E. coli BL21 (DE3)/ pEXP-5CT/TOPO®-C (TOPO-C) 誘導時同時添加 2%
Glycine 及 Triton X-100 後,納豆菌漆酶活性隨誘導時間釋放至胞外的變化 情形 ... 34 圖 6 E. coli BL21 (DE3)/ pEXP-5CT/TOPO®-C (TOPO-C) 經誘導並濃縮 10 倍後,
再經釋放處理之胞外漆酶活性,隨釋放時間變化情形 ... 35 圖 7 利用 5 L 醱酵槽以細胞高密度培養 E. coli BL21 (DE3)/ pEXP-5CT/TOPO®-C
(TOPO-C),外泌生產納豆菌漆酶情形 ... 36
第 1 章 前言
1.1. 大腸桿菌 (Escherichia coli) 外泌系統
細菌常主動分泌消化酵素、毒性蛋白質或其它致病物質至胞外,藉以從周 遭環境攝取營養素或對抗其它生物以利生存,此類外泌蛋白質常帶有特殊序 列的胺基酸片段,稱作訊號序列 (signal peptide),可供胞內特定蛋白質辨認,
並協助運送至細胞膜上對應的通道蛋白質,主動運輸目標蛋白質至胞外。
革蘭氏陰性菌因具有包含內膜 (inner or cytoplasmic membrane)、胞膜間區 (periplasmic space) 及外膜 (outer membrane) 等膜結構,胞內物質向外運輸的 調控亦較革蘭氏陽性菌複雜,革蘭氏陰性菌的外泌系統主要分為第一至第五 型外泌系統 (Type I~V secretion system) [1] ,而其中又以常應用於外源基因 表現的大腸桿菌 (Escherichia coli) 第一型及第二型外泌系統相關研究為多,
故以下分別針對此二型外泌系統進一步介紹。
1.1.1. 第一型外泌系統 (Type I secretion system)
第一型外泌系統是藉由一系列橫跨內、外膜的通道蛋白質,於細胞質側辨
認C 端帶有特定訊號序列的蛋白質後,經過胞膜間區,直接主動運輸至胞外,
蛋白質運送到胞外後仍保留其C 端訊號序列。該種外泌途徑最常見於部分致
病型大腸桿菌,如J96、CFT073 等泌尿道感染型大腸桿菌品系(uropathogenic E. coli , UPEC) 於分泌-hemolysin 等 repeats-in-toxin (RTX) 毒素時使用,另 外在其它菌種亦有作為蛋白酶或脂肪酶的分泌途徑 [2]。
以大腸桿菌外泌毒性蛋白-hemolysin (HlyA) 途徑為例,HlyA 蛋白質 C 端帶有glycine 及 aspartate 密集且重複的訊號序列 (G-G-X-G-X-D-X,字母為 胺基酸縮寫,X 為任一胺基酸),該段序列可被細胞內膜上的穿膜蛋白質 HlyB 辨認,於分解ATP 提供能源後,結合 HlyD 及外膜上 TolC 通道蛋白質,形成
橫跨內、外膜的運輸通道,直接將HlyA 連同訊號序列一併運送至胞外。
1.1.2. 第二型外泌系統 (Type II secretion system)
第二型外泌系統係兩階段分泌,即目標蛋白質先由內膜運輸系統運送至 胞膜間區後,進一步穿越外膜分泌至胞外。經由第二型外泌系統運輸的目標
蛋白質,其 N 端帶有特殊訊號序列,可被細胞質中不同的伴護蛋白質
(chaperone) 辨認,協助目標蛋白質折疊 (folding) 並攜帶至內膜附近以利運 輸 [3],目標蛋白質所帶的訊號序列則於穿過內膜時被切除。
前述N 端訊號序列一般約由 18 至 30 個胺基酸組成,該序列通常包含 n- region、h-region 及 c-region 等三部分,n-region 為帶正電的胺基端 (amino terminus),h-region 為疏水性核心 (hydrophobic core),而 c-region 則為具極性 的切割端 (cleavage region) [1],當目標蛋白質穿過內膜,被運送至胞膜間區 時,即由c-region 切除訊號序列。
依據訊號序列特徵、伴護蛋白質及內膜運輸蛋白質種類的不同,第二型外 泌系統可細分為Sec-dependent (Sec) pathway 及 Twin-arginine translocation (Tat) pathway。經由 Sec pathway 運送的蛋白質,在細胞質內多尚未完成折疊,需 由 SecB 伴護蛋白質或其它由蛋白質及 RNA 組成的信號識別顆粒 (signal recognition particle , SRP) 辨認結合及協助運送至胞膜間區方能完成折疊並分
泌出胞外;而在細胞質內已正確結合相關輔酶,且完成折疊者,多經由 Tat
pathway 運輸。此外,經由 Tat pathway 運輸的蛋白質,其訊號序列的 n-region 通常含有S/T-R-R-X-F-L-K 的序列特徵 [4, 5]。
1.2. 異源蛋白質的表現與外泌
大腸桿菌因具有生長快速、易於高密度培養且外源蛋白質負載量高等特 性,且其基因轉殖、表現、調控及培養策略的相關研究皆較完整,故在重組 DNA 技術生產工業、食用酵素或醫療用蛋白質等應用方面已被廣泛利用 [3,
6]。為提升異源蛋白質產量,一般藉由強效啟動子、轉形多倍數量基因或細 胞高密度培養等方式,提高目標基因表現量或總菌量達成目的。但大腸桿菌 大量表現外源基因時,常發生蛋白質折疊不完全而無活性的異源蛋白於細胞 內聚集,又無法外泌而形成包涵體 (inclusion bodies),實務上易增加後續蛋白 質純化與復性 (renaturation) 等處理程序的複雜性及生產成本 [7]。
為了改善異源蛋白於胞內累積的問題,許多研究傾向利用大腸桿菌既有 的外泌系統,於目標蛋白質上修飾特定訊號序列,誘使宿主細胞主動外泌目 標蛋白質至胞外,即可在培養基中回收。例如,Su 等人利用第一型外泌系統 的-hemolysin (HlyA) secretion pathway , 於 目 標 蛋 白 質 修 飾 HlyA signal peptide,成功以大腸桿菌外泌表現嗜高溫放線菌 Thermobifida fusca 的角質酶 (cutinase) [8]。
此外,亦有研究利用第二型外泌系統,誘使宿主細胞將目標蛋白質運送至 胞膜間區,再將目標蛋白質釋放至培養基;釋放的方式包含物理法如冷滲透 休克法 (cold osmotic shock),化學法如添加 lysozyme、glycine、Triton X-100
或EDTA 等,抑或直接使用膜狀結構較為脆弱的突變菌株作為宿主細胞,以
利目標蛋白質直接釋出 [9, 10]。例如,Gerard 等人利用 E. coli Twin-arginine translocation (Tat) pathway TorA 訊號序列 (torAsp),將耐熱溫泉菌 Thermus thermophilus 胞內表現的 -glycosidase 運送至 E. coli 宿主細胞的胞膜間區,
再藉由冷滲透休克法釋放至胞外 [11]。
然而,許多的研究顯示,宿主細胞的品系、訊號序列的種類,及目標蛋白 質本身的特性,都可能影響外泌生產的成功與否及其效率;而訊號序列的選 擇,也未有通用的規則可循。因此,對於不同的目標蛋白質,目前僅能藉由 不斷嘗試,找尋其最適的外泌途徑及生產方法 [3, 9]。
1.3. 漆酶 (laccase)
1.3.1. 起源與功能
漆酶 (EC 1.10.3.2, benzenediol: oxygen oxidoreductase) 係指一類以銅離子 作為輔因子 (cofactor) 的多酚氧化酶 (polyphenol oxidase, PPO),最早於 1883 年由 Yoshida 氏發現漆樹 (lacquer, Toxicodendron vernicifluum, 異名 Rhus vernicifera) 樹液的硬化和其含有可氧化酚類化合物的酵素有關,並稱之為漆 酶 (laccase) [12]。
漆酶於自然界分佈甚廣,在植物、真菌、昆蟲及細菌中皆有類似功能的酵 素或序列之發現。由於漆酶的來源多,不同物種間的漆酶在序列、結構、催 化能力或特性方面差異很大,而在同一來源的漆酶,其反應基質、最適催化 條 件 亦 有 不 同 。 目 前 研 究 較 為 廣 泛 的 漆 酶 , 以 真 菌 中 的 擔 子 菌 (Basidiomycetes) 及子囊菌 (Ascomycetes) 等來源為大宗,而其中白腐真菌 (white rot fungi) 可分別分泌纖維素 (cellulose) 及木質素 (lignin) 的分解酵 素,使木材顏色變淡或成白色,是真菌漆酶相關研究中最為詳細的 [13]。
不同於真菌漆酶,許多研究顯示漆酶在不同物種當中扮演多元的角色。例 如,Bao 及 Sato 等人在火炬松 (Pinus taeda) 木質部及細胞壁發現的漆酶可參 與木質素的生合成 [14, 15] ;另外,根據 Dittmer 等人的研究,菸草天蛾 (Manduca sexta) 及甘比亞瘧蚊 (Anopheles gambiae) 之漆酶基因 msLac1、
msLac2 及 agLac1 的表現,和昆蟲的消化系統運作或不同時期之外骨骼發育有 關 [16]。
而在細菌漆酶的研究方面,一般認為與細菌的型態發生、防止內孢子遭受 UV、過氧化氫 (H2O2) 破壞或維持細胞內銅離子濃度的恆定有關 [17]。以枯 草桿菌 (Bacillus subtilis) 的 CotA 蛋白質為例,無論是在其胺基酸序列與結構 上 所 具 有 的 4 個 保 守 區 域 , 及 對 於 包 含 Syringaldazine (N,N-bis(3,5-
dimethoxyhydroxy-benzylidenehydrazine)) 、 ABTS (2,2'-azino-bis(3- ethylbenzothiazoline-6-sulphate)) 等常見的漆酶基質,均有催化功能,因此 CotA 被認為是一種漆酶。依據 Hullo 等人研究,CotA 主要表現於枯草桿菌內孢子 表層,並參與內孢子表面褐色色素的生合成,及類似真菌漆酶之下游產物 melanin-like product,以提供內孢子對抗 UV 光的侵害 [18]。
1.3.2. 結構
漆酶包含 4 個銅離子鍵結,依據電子順磁共振 (electron paramagnetic resonance, EPR) 特性進一步分為 3 類:第 1 類 (Type 1 Cu) 為順磁性,且於 610 nm 具有吸收峰,使漆酶結晶成藍色而稱藍銅 (blue copper) 酵素;第 2 類 (Type 2 Cu) 雖具順磁性,惟於可見光譜無吸收,故僅於電子順磁共振光譜中 可偵測;第3 類銅離子 (Type 3 Cu) 於 330 nm 具有吸收峰,但因含有成對電 子而無順磁性,僅可由吸收光譜測得 [13, 19]。
另外,依據Kumar 等人分析 100 餘種漆酶結構,發現位於 4 個銅離子結 合位的12 個胺基酸均含 HXH motif 的保守區域 (L1-L4) ,其中 L2 和 L4 在 不同漆酶間的保守性高,L1 和 L3 的差異與漆酶的多樣性及還原電位差異有 關 [20, 21]。以 Bacillus subtilis 漆酶 CotA 為例,由 Type 1 Cu 與 HCH motif 中的Cysteine 492 結合,另 Histidine 491、Histidine 493 則分別與 Type 3 Cu 結合,發生氧化還原反應時,電子即經由HCH motif 於 Type 1 Cu→Type 2 Cu
→Type 3 Cu 所構成的核心區域傳遞 [20] 。
1.3.3. 應用
漆酶主要藉由對酚類或芳香環類化合物進行氧化反應,使其產生自由基,
由O2接收電子並還原為H2O [17] ,以 benzenediol 為定義基質的反應定義式:
4 benzenediol + O2 → 4 benzosemiquinone +2 H2O
相較於過氧化酶,漆酶的還原電位較低,過去認為僅能氧化酚類化合物,
但 後 續 研 究 指 出 , 若 添 加 適 當 還 原 介 質 如 ABTS (2,2'-azino-bis(3- ethylbenzothiazoline-6-sulphate)),先經漆酶反應後生成較高還原電位的自由基,
除能有效提升木質素分解效率,亦能分解多環芳香化合物及工業染劑等 [22, 23]。
在紙漿工業上,去除木質素是相當重要的加工程序,不同於過去必須使 用強酸、強鹼並在高溫下利用化學法分解,漆酶能在相對溫和的條件下分解木 質素,不僅有效降低成本,也不易對環境產生壓力,是目前主要的應用方向[23]。
由於漆酶對氧化基質種類較無專一性,在含氧氣的環境中即可反應並分 解許多含有生物毒性及污染性的酚類或芳香環類化合物,副產物為水,對環境 影響小,因此,也被應用於工業廢水處理。依Abadulla 等團隊研究,Trametes hirsuta 漆酶能分解 triarylmethane、indigoid、azo 及 anthraquinone 等染劑,並 降低染劑對Pseudomonas putida 的生物毒性達 80% [22]。
1.4. 漆酶的異源表現
生物所自然生合成的漆酶受限於原物種生長特性,漆酶產量低且分離純
化程序繁瑣,難以符合工業應用需求。故利用重組DNA 技術將漆酶基因轉由
較易培養、生長快速的宿主生產表現,可有效提高產量,也能藉由基因工程技 術進一步調控表現機制、提高酵素專一性及反應環境耐受性等。例如,在真菌 漆酶異源表現的研究方面,Hong 等人以醱酵槽培養技術,利用 Pichia pastoris SMD1168 生產雲芝 Trametes versicolor 漆酶 Lcc1,最高活性可達 140 U/mL [24]。
細菌漆酶較真菌漆酶具有反應溫度高、耐受pH 值廣,且細菌較真菌生長
快速等優勢,但會受限於細菌漆酶的表現量較低,不易純化等問題。以Bacillus subtilis 漆酶基因 cotA 為例,該基因主要表現於內孢子表層,細胞表現漆酶 時已進入內孢子形成狀態,難以透過一般蛋白質純化技術取得,遑論進行量
產與應用。因此,藉由異源表現方式大量生產蔚為主流。依據Guan 等團隊研 究,將Bacillus pumilus W3 漆酶 CotA 表現於 Bacillus subtilis WB600 中,該 重組漆酶在 20-90°C 及 pH 6.5-11.5 對 2,6-DMP 皆有活性,以醱酵槽培養並 經酵素純化後最高產量達373.1 U/ml [25]。
1.5. 納豆菌
枯草桿菌 (Bacillus subtilis) 屬於芽孢桿菌屬 (Bacillus),為革蘭氏陽性、
好氣菌,在土壤、生物體表面等自然環境中普遍存在,當外在環境惡劣不利生 長時,會形成內生孢子進入休眠。另,其全基因體序列於1997 年完成解碼 [26]。
目前在生物防治、農業資材及食品添加等方面已被廣為利用。
納豆菌 (Bacillus subtilis natto) 為枯草桿菌的亞種之一,不同於其它枯草 桿菌,納豆菌生長需要生物素 (biotin) [27],為製作日本傳統發酵食品納豆 (な っ と う, natto) 所 使 用 的 菌 種 。 納 豆 菌 發 酵 分 解 大 豆 時 會 產 生 大 量 - polyglutamic acid (-PGA),即納豆表面的黏稠狀物質, -PGA 以及其衍生物 除在化妝品的生產應用,也可作為包覆益生菌或其它食品的無毒抗凍劑,應用 範圍十分廣泛 [28-30]。此外,納豆菌亦能分泌多種蛋白酶且不會產生毒素,
被認為是安全的 (Generally recognized as safe, GRAS) 食用菌株。近年研究也 發現,納豆菌可藉由促進腸道乳酸菌生長,改變腸道菌相而被認為是腸道益生 菌 [31]。此外,也有研究發現,將納豆菌添加於雞隻飼料中餵食 28 天,可促 進其腸道細胞發育,有利雞隻生長 [32]。
1.6. 研究目的
本實驗室先前自納豆菌 Bacillus subtilis natto NTU18 中選殖漆酶基因 cotA ,並轉殖至大腸桿菌表現,發現重組納豆菌漆酶的熱穩定性較真菌漆酶為 佳。為進一步優化重組大腸桿菌生產納豆菌漆酶之效能與可調控性,以減低工 業化生產及純化程序成本,本研究試以訊號序列修飾納豆菌漆酶基因,及調整
胞膜間區蛋白質釋放條件等方式,探討重組大腸桿菌外泌生產納豆菌漆酶的可 行性並優化生產條件,並配合細胞高密度培養技術,以提升納豆菌漆酶單位活 性及產量,俾利相關產程發展。
1.7. 研究大綱
本研究藉由在cotA 基因上分別修飾第一型 (hlyA) 或第二型外泌系統訊 號序列,並試以E. coli M15/pQE 30-Xa 及 E. coli BL21 (DE3) 系統表現,利 用Hinton’s flask 振盪培養篩選適當培養及納豆菌漆酶胞外釋放條件後,轉以 醱酵槽進行細胞高密度培養,並探討納豆菌漆酶外泌生產方式及相關策略之 最適化。
第 2 章 材料與方法
2.1. 菌株及基因載體
本研究選用Escherichia coli DH5作為重組基因質體之保存及複製用菌株,
另分別使用coli M15 (genotype: F–, Φ80ΔlacM15, thi, lac–, mtl–, recA+ , KmR) 及E. coli BL21(DE3) (genotype: F– ompT gal dcm lon hsdSB(rB–mB–) λ(DE3 [lacI lacUV5-T7p07 ind1 sam7 nin5]) [malB+]K-12(λS))等菌株,作為以 pQE-30 Xa (QIAGEN, Hilden, Germany) 及 pEXP5-CT/TOPO® (Invitrogen)質體為載體之重 組基因表現宿主。
E. coli M15/pQE-30 Xa 系統係由 QIAGEN 公司所開發,由於 pQE-30 Xa 質體上建構有T5 promoter/lac operator,可於添加 IPTG 後啟動表現下游插入 基因。另外該質體於Multiple cloning site (MCS) N 端的 Histidine tag,有助於 所表現蛋白質後續利用Ni-NTA 管柱層析法進行純化;FXa recognition site 則 可被FXa protease 辨認切除。另 E. coli M15 因含有 pREP4 質體,搭配 pQE- 30 Xa 質體可有效減少誘導前目標蛋白質先行表現而影響菌體生長,且可利用 kanamycin 篩選,提高正確轉形株比率。
E. coli BL21(DE3)/ pEXP5-CT/TOPO®系統係利用Invitrogen 公司所開發之 pEXP5-CT/TOPO® TA Expression Kit,由於 E. coli BL21(DE3) 含有 T7 RNA polymerase pEXP5-CT/TOPO®質體使用T7 promoter,可於添加 IPTG 後啟動表 現下游插入基因。另外該質體於Multiple cloning site (MCS) N 端的 Histidine tag,有助於所表現蛋白質後續利用 Ni-NTA 管柱層析法進行純化;FXa recognition site 則可被 FXa protease 辨認切除。另 E. coli M15 因含有 pREP4 質體,搭配 pQE-30 Xa 質體可有效減少誘導前目標蛋白質先行表現而影響菌 體生長,且可利用kanamycin 篩選,提高正確轉形株比率。
2.1.1. 熱休克 (heat shock) 細胞轉形法
取200 ng 質體 DNA 加入 20 L E. colicomplement cell,靜置冰上 30 分 鐘後以42°C 水浴 1 分鐘,置於冰上 5 分鐘,加入 LB 培養基至 200L,以 37°C 振盪培養 1 小時,取 100 L 菌液塗抹於適當之篩選培養基。
2.2. 含訊號序列之納豆菌漆酶重組基因與質體建構
2.2.1. 納豆菌漆酶基因 cotA
本研究使用之納豆菌漆酶基因 cotA,係由本實驗室保有之納豆菌株 B.
subtilis natto NTU18,並由林軒立同學由該菌株中選殖而得,DNA 序列全長 1,542 bp,轉換為胺基酸序列並與 NCBI 登錄之標準菌株 B. subtilis str. 168 spore coat protein A 序列比較,僅 1 處胺基酸序列不同 (Tyr400 → Ala400),但變異 位置並非位於銅離子所結合之4 個 HXH domain。重組納豆菌漆酶 CotA laccase 最適反應溫度為85°C,相對市售雲芝 Trametes versicolor 最適反應溫度 65°C 為高。另納豆菌漆酶最適反應酸鹼值為4.5;酵素動力學係數 Km為0.176 mM,
Vmax為1.74 M min-1 [33]。
2.2.2. 訊號序列 (signal peptide, sp)
2.2.2.1. 第一型外泌系統 C 端訊號序列 (C-terminal signal peptide for type I secretion system )
參 照 NCBI 上 經 解 序 之 E. coli J96 hemolysin A 蛋 白 (protein ID:AAA23975.1) 末端第 965-1024 個胺基酸之基因序列,並利用 GeneDesign 工具 (http://54.235.254.95/gd/) 設計引子組 cotA-hlyA-1~6 (如表 1),分別製備 模板引子混合液 (template primer mix, TPM):6 組引子混合並以 ddH2O 調配 至終濃度15 nM ;及外引子混合液 (outer primer mix, OPM) :取 cotA-hlyA- 1 與 cotA-hlyA-6 混合並以 ddH2O 調配至終濃度 1.5 M。再以 Polymerase Chain Assembly Method (PCA) 反應 [34] 合成 C 端訊息序列 DNA 片段 hlyAsp。
PCA 反應流程:取 2.5 L TPM、10 L ddH2O 及 12.5 L Q5® High-Fidelity DNA Polymerase (NEBuilder) 混合,並以下述流程反應:98°C 15 分, 50°C 30 秒,72°C 1 分;98°C 30 秒,50°C 30 秒,72°C 1 分,25 個循環;72°C 3 分鐘後維持於4°C。前述反應產物以 ddH2O 稀釋 5 倍後取 2.5 L,混合 2 L OPM、10 L ddH2O 及 12.5 L Q5® High-Fidelity DNA Polymerase (NEBuilder) , 再經98°C 15 分, 50°C 30 秒,72°C 1 分;98°C 30 秒,50°C 30 秒,72°C 1 分,25 個循環;72°C 3 分鐘後維持於 4°C 完成反應,並取合成之 C 端訊息序 列DNA 片段,以 Zero Blunt® PCR Cloning Kit (Invitrogen) 接合於 pCR®-Blunt 質體後,轉形於E. coli DH5增殖後收取質體DNA 定序確認。
2.2.2.2. 第二型外泌系統 N 端訊號序列 (N-terminal signal peptide for type II secretion system)
本實驗針對第二型外泌系統,分別挑選 Sec-dependent (Sec) pathway:
malEsp、ompAsp,及 Twin-arginine translocation (Tat) pathway:torAsp、amiAsp 等較多誘使異源蛋白質成功外泌的訊號序列 (sp)。malEsp 及 ompAsp 分別取 自E. coli maltose-binding periplasmic protein 及 outer membrane protein A 之 N 端序列,該等蛋白質係藉由Sec pathway 運送,且分別與細胞攝入營養物質及 外膜蛋白質的運送有關;而 torAsp 及 amiAsp 則取自 E. coli Trimethylamine- N-oxide reductase 及 N-acetylmuramoyl-L-alanine amidase 之 N 端序列,該等 蛋白質係藉由Tat pathway 運送,皆與細胞攝入營養物質有關。
利用Wizard Genomic DNA Purification Kit (Promega, Madison,WI, USA) 抽取之coli K12 菌株染色體 DNA 作為模板,並分別以 TorAF1、TorAR1;
AmiAF1、AmiAR1;MalEF1、MalER1 及 OmpAF1、OmpAR1 等引子對進行 聚合酶連鎖反應 (Polymerase chain reaction, PCR),增殖得 N 端訊號序列 DNA 片段:torAsp、amiAsp、malEsp 及 ompAsp。PCR 反應劑量:模板 DNA 1 L、
0.5 M 預混引子對 4 L 及 Q5® High-Fidelity DNA Polymerase (NEBuilder) 5
L。PCR 反應流程:98°C 30 秒;98°C 10 秒,55°C 30 秒,72°C 10 秒,30 個循環;72°C 2 分鐘後維持於 4°C。另以 Zero Blunt® PCR Cloning Kit (Invitrogen) 接合於 pCR®-Blunt 質體後,轉形於 E. coli DH5增殖後收取質體 DNA 定序確認。
2.2.3. E. coli M15/pQE-30 Xa-cotA-hlyA 表現系統建構
以所合成之C 端訊號序列 DNA 片段及 cotA 基因為模板,並使用 pQE→
cotA、cotA←hlyA、cotA→hlyA、hlyA←pQE 等引子進行 PCR,並以經 smaI 限制酶處理後之pQE-30 Xa 直鏈狀質體,利用 HiFi DNA Assembly Master Mix (NEBuilder) 接合成環狀質體 DNA,轉形至 E. coli M15,並以含 50 ppm kanamycin 及 100 ppm ampicillin LB agar plate 篩選具抗性菌落,並抽取其質 體DNA,定序確認插入質體之序列正確,以 3 mL LB 於 37°C 隔夜培養,取 70 L 加入 30 L 50% glycerol 於-80°C 保存。
2.2.4. E. coli M15/pQE-30 Xa-(Type II signal peptide)-cotA 質體建構
本研究利用Gibson assembly 技術將帶有 signal peptide 之 cotA 基因片段 嵌入pQE-30 Xa 質體,Gibson assembly 係由 Daniel Gibson 於 2009 年所發明,
藉由設計具有相互重疊序列的引子,於兩段待接合的 DNA 添加重疊序列後,
再利用PCR 反應接合。
同時以前項所合成之N 端訊號序列 DNA 片段及 cotA 基因為模板,並使 用 A-TorA-cotA、A-AmiA-cotA、A-MalE-cotA、A-OmpA-cotA 與 cotA-6His 等引子進行PCR,合成 torAsp-cotA、amiAsp-cotA、malEsp-cotA 及 ompAsp- cotA 等片段,混合經過 smaI 限制酶處理後之 pQE-30 Xa 直鏈狀質體,利用 HiFi DNA Assembly Master Mix (NEBuilder) 接合成環狀質體 DNA,轉形至 E.
coli M15,並以含 50 ppm kanamycin 及 100 ppm ampicillin LB agar plate 篩選 具抗性菌落,並抽取其質體 DNA,定序確認插入質體之序列正確,以 3 mL
LB 於 37°C 隔夜培養,取 70 L 加入 30 L 50% glycerol 於-80°C 保存。
2.2.5. E. coli BL21 (DE3)/pEXP5-CT/TOPO®-(Type II signal peptide)-cotA 質
體建構
以前項所合成種 signal peptide-cotA 片段,利用 pEXP5-CT/TOPO® TA Expression Kit (Invitrogen),接合成環狀質體 DNA,轉形至 E. coli BL21 (DE3) , 並以含100 ppm ampicillin LB agar plate 篩選具抗性菌落,並抽取其質體 DNA,
定序確認插入質體之序列正確,以3 mL LB 於 37°C 隔夜培養,取 70 L 加 入30 L 50% glycerol 於-80°C 保存。
2.3. 重組大腸桿菌培養與誘導異源漆酶表現
2.3.1. 500 mL Hinton’s flask 振盪培養
種菌培養以30 L 菌液置於 3 mL LB broth 中, 37°C 隔夜培養後,取 1 mL 種菌菌液,加入含 100 mL LB broth 及 50 ppm kanamycin、100 ppm ampicillin (E. coli M15/pQE-30 Xa) 或 100 ppm ampicillin (E. coli BL21 (DE3)/pEXP5- CT/TOPO®) 之 500 mL Hinton’s flask,於 37°C 振盪培養至 OD600=0.8-1.0,分 別 添 加 0.05 mM IPTG (Isopropyl β-D-1-thiogalactopyranoside) 及 0.25 mM CuSO4,並移至25°C 振盪誘導培養。為最適化誘導及胞膜間區釋放條件,另 分別測試 25°C 和 37°C 誘導溫度,及在 IPTG 濃度分別為 0.05、0.1、0.5、
1、5、10 mM 等誘導條件下之漆酶活性表現。
2.3.2. 5 L 醱酵槽培養 2.3.2.1. pH-stat 饋料培養
本研究使用Biostat B (B. Braun, Sartorius, Germany) 醱酵槽,以 pH-stat 饋料策略進行細胞高密度培養提升重組大腸桿菌菌量,提高納豆菌漆酶產 量及單位活性。培養起始溫度為37°C,轉速 300 rpm,以 4N NaOH 及饋料
培養基 (Nutrient solution) 控制 pH 值於 7.0,通氣量為 1 vvm,當起泡嚴重 時並添加消泡劑 (Sigma antiform-204)。以溫度計、 pH 計及溶氧電極 (Dissolved Oxygen probe, DO probe) 監控培養全程並記錄相關參數之變化。
E. coli 生長初期優先代謝主培養基中的葡萄糖 (glucose) 獲取能量而 產酸,使環境中pH 值下降,系統藉由饋入 4N NaOH 平衡之。當主培養基 碳源耗盡,氮源使用效率提升,致環境pH 值上升高於 7.05 時,系統自動饋 入饋料培養基,由於饋料培養基內葡萄糖含量極高 (75%),菌體在碳源匱乏
狀態會快速耗氧代謝葡萄糖產能,使槽內溶氧度及pH 值降低;當饋入的饋
料培養基葡萄糖再次用罄,槽內溶氧度及pH 值升高,進而啟動下一次饋料。
藉由上述饋料培養方式,逐步累積細胞密度。
2.3.2.2. 培養流程
種菌培養先以3 mL LB broth 於 37°C 隔夜預培養,再取 1 mL 菌液接種 於含有 100 mL LB 培養基 (內含 100 ppm ampicillin) 之 500 mL Hinton’s flask,置於 37°C、120 rpm 培養 12 小時。接種時,取 15 mL 種菌接種於配 置 1.5 L modified R medium (含 6 mL/L Trace metal solution 及 100 ppm ampicillin) 之 5 L 醱酵槽,開始批次培養並同步記錄相關參數。
接種後培養12 小時,加入終濃度 0.05 mM IPTG 進行 CotA laccase 誘導 表現,每24 小時並添加終濃度 0.25 mM CuSO4以提供銅離子作為漆酶輔因 子,IPTG 誘導開始時,培養溫度降低至 25°C。培養期間於固定時間點取 10 mL 菌液稀釋塗抹於 LB agar plate 監測生菌數;並從中取 1 mL 菌液以 14,000
×g 離心 10 分鐘,上清液以 dinitrosalicylic acid (DNS) 法 [35] 測量培養基 殘糖值;菌體置於 -80°C 冷凍後以冷凍乾燥機凍乾,測量細胞乾重。另外,
於誘導開始後,於固定時間點取出15 mL 菌液樣品,以 10,000 ×g 離心 10 分鐘後去除上清液,並以同體積 LB broth 回溶後取 10 mL 移入 50 mL Hinton’s flask,並添加 2% Glycine 及 Triton X-100,置於 25°C,120 rpm 振
盪培養並釋放目標蛋白質24 小時後,以 14,000 ×g 離心 5 分鐘,取上清液 測定漆酶活性。
2.4. 冷滲透休克法 (Cold osmotic shock)
大腸桿菌為格蘭氏陰性菌,具有雙層膜狀結構,包含外層膜 (outer membrane) 及胞膜間區 (periplasmic space) 等構造。本研究為分離胞膜間區 蛋白質,將菌體分離後,在低溫下,先後置於高滲透壓溶液及純水中,藉由 滲透壓快速改變,水分快速進出胞膜間區,終使外膜破裂剝離,以達分離胞 膜間區蛋白質的目的。
取1 mL 經誘導培養之菌液,以 10,000 ×g 離心 5 分鐘,去除上清液後,
加入1 mL 100 mM Tris–HCl,20% sucrose 及 1 mM EDTA (0°C,pH 8.0) 振 盪回溶 10 分鐘,以 8,000 ×g 離心 5 分鐘後去除上清液,加入 500L 0°C QH2O 回溶,並靜置於冰上 10 分鐘;加入 500L 2 mM MgCl2後,靜置於冰 上10 分鐘後,以 8,000 ×g 離心 5 分鐘,取上清液測定漆酶活性。
2.5. Glycine /Triton X-100 釋放法
在誘導表現同時釋放至胞膜間區實驗中,則於誘導時另添加 2% Glycine 及Triton X-100,置於 25°C 振盪培養 16 小時後以 10,000 ×g 離心 5 分鐘,取 上清液測定漆酶活性。
為提高漆酶純度,另測試先以100 mL LB 液態培養基及抗生素於 500 mL Hinton’s flask 於 37°C 振盪培養重組大腸桿菌至 OD600=0.8-1.0,分別添加 IPTG 及 CuSO4至0.05 mM 及 0.25 mM,並移至 25°C 振盪誘導培養 16 小時 後以10,000 ×g 離心 10 分鐘,去除上清液,並以 10 mL 滅菌 LB 液態培養基 回溶菌體,移至50 mL Hinton’s flask 後加入 2% Glycine 及 Triton X-100,繼 續於25°C 振盪培養,並於固定時間取樣測定漆酶活性。
2.6. 漆酶活性測定
本研究以2,2'-azino-bis(3-ethylbenzothiazoline-6-sulphate).(NH4)2 (ABTS) 為漆酶反應基質,ABTS 經漆酶氧化後形成含穩定自由基的 (ABTS.),其在 波長420 nm 光譜有最大吸收峰值。
將待測溶液經適當稀釋後,取50 L 加入 900 L Laccase reaction buffer (55.6 mM NaOAC、0.25 mM CuSO4) 及 50 L 10 mM ABTS,於 30°C 反應 10 分鐘後,測定420 nm 吸光值。漆酶 1 活性單位 (Unit) 定義為 30°C、1 分鐘 氧化1 mole ABTS 所需酵素量,又 ABTS 吸光常數=36.0 mM-1cm-1,依據 Beer’s law:(A:吸光值,C:濃度,t:反應時間,b:吸光直徑=1 cm)
∆A420= εbC
故漆酶活性計算如下:
Laccase activity (U mL-1):
= C
t × dilution factor
= (∆A420
εbt ) × dilution factor
= ( ∆A420
36.0 × 1 × 10) × 20
= ∆A420 18
2.7. SDS-PAGE 電泳與活性染色分析
配製 10 mL 12.5% 分離膠體 (separating gel) 溶液及 5 mL 4%焦集膠體 (stacking gel) 溶液,並鑄成 SDS-PAGE 電泳膠片 2 片,以 TGS buffer (BIOMAN) 加入電泳槽,並將 20 L 蛋白質樣品加入 5 L 5× non-reducing SDS sample buffer 混合後,注入膠片樣品槽中。以 80 V 電泳至樣品越過焦集膠體後,提高
電壓至120V 電泳至追蹤染劑脫離分離膠體後停止。取其中一片電泳膠體,置
振盪隔夜退染。另一片電泳膠體,置於含50 mM ABTS 的 Laccase reaction buffer,
反應5 分鐘後以 QH2O 退染 1 小時。
第 3 章 結果
3.1. 納豆菌漆酶 (CotA laccase) 於 E. coli M15/pQE-30 Xa 系統之表
現3.1.1. 含訊號序列 (signal peptide) 之納豆菌漆酶的建構與表現
利用自納豆菌株B. subtilis natto NTU18 選殖的 cotA 基因序列,分別在 其C 端增加第一型外泌系統訊號序列 E. coli J96 hlyAsp;及其 N 端增加第二 型外泌系統訊號序列:E. coli K12 torAsp、amiAsp、malEsp 及 ompAsp 等,
再以pQE-30 Xa 質體轉形至 E. coli M15 表現。
以100 mL LB 培養基於 500 mL Hinton’s flask 振盪培養轉形完成之 E.
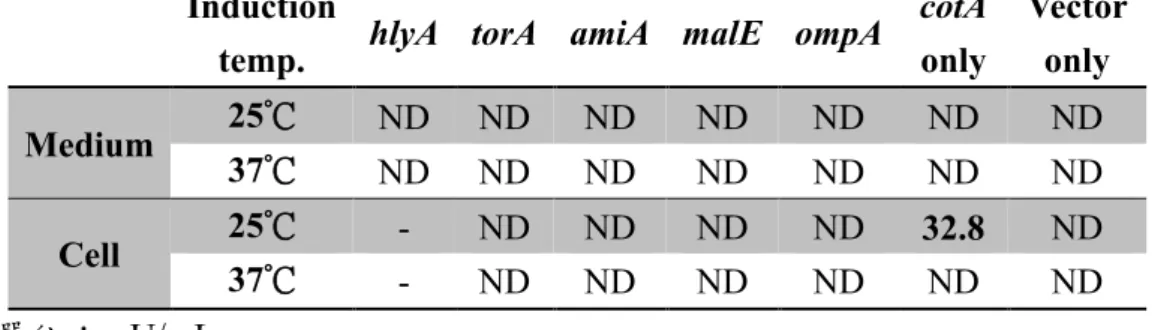
coli M15 並於 25°C 及 37°C、以 0.05 mM IPTG 誘導 CotA laccase 表現 18 小 時後,於培養基中均無法測得漆酶活性。進一步利用超音波破菌後取可溶蛋 白質測試,僅可於25°C 誘導且未修飾訊號序列的對照組 (cotA only) 測得 漆酶活性32.8 mU/mL (如表 2)。
3.1.2. 納豆菌漆酶在胞膜間區 (periplasmic space) 之累積
為了解經修飾具有第二型外泌系統訊號序列的納豆菌漆酶,是否因聚集 於胞膜間區以致無法成功外泌至胞外,故以含100 mL LB 培養基於 500 mL Hinton’s flask 振盪培養各重組 E. coli 並於 25°C 誘導後,使用冷滲透休克法 (cold osmotic shock) 來分離 E coli M15 胞膜間區可溶蛋白質,並測量其漆酶 活性。結果顯示,無論是否修飾具有第二型外泌系統訊號序列,納豆菌漆酶 均有累積在胞膜間區的現象,但未修飾帶有此訊號序列的對照組,其在胞膜 間區所測得漆酶活性仍高於具有修飾訊號序列組別,達93.4±1.09 mU/mL (如
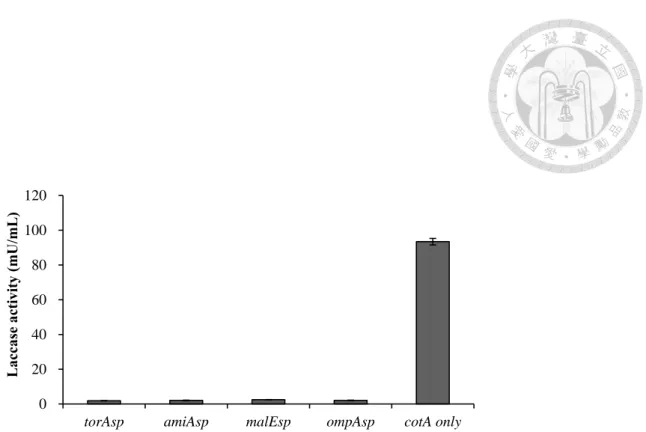
圖1)。顯示訊號序列的存在,未能提升納豆菌漆酶的外泌輸送。
3.2. 納豆菌漆酶於 E. coli BL21 (DE3)/pEXP-5CT/TOPO
®系統之表現3.2.1. 含訊號序列納豆菌漆酶的建構與表現
前述在 E. coli M15/pQE-30 Xa 系統表現含第二型外泌系統訊號序列納 豆菌漆酶,推測可能受到pQE-30 Xa 質體所設計的 Histidine tag 及 Factor Xa (FXa) recognition site 於目標基因序列 N 端的遮蔽,影響了訊號序列功能,
故將含第二型外泌系統訊號序列的納豆菌漆酶改建構於 pEXP-5CT/TOPO® 質體,並轉形至E. coli BL21 (DE3) 表現。
以100 mL LB 培養基於 Hinton’s flask 振盪培養轉形完成之 E. coli BL21 (DE3),並分別於 25°C 及 37°C、以 0.05、0.5、1、5、10 mM IPTG 誘導 CotA laccase 表現後,在培養基中仍無法測得漆酶活性。顯示利用上列第二型外 泌系統訊號序列無法外泌生產CotA laccase。
3.2.2. 納豆菌漆酶在胞膜間區之累積
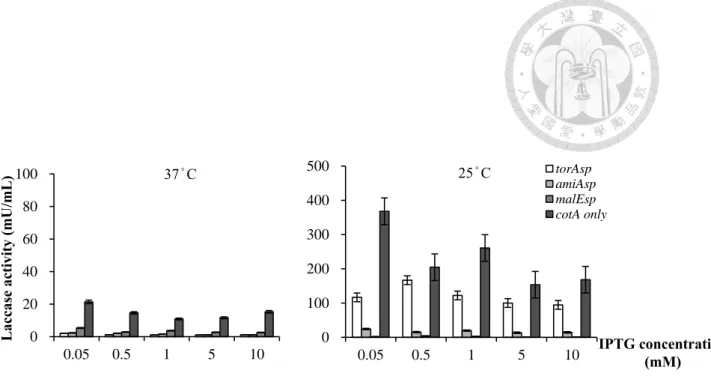
進一步利用冷滲透休克法分離E coli BL21 (DE3) 胞膜間區可溶蛋白質並 測量其漆酶活性。結果如圖2 所示,在 25°C 誘導下,於胞膜間區納豆菌漆 酶的累積,多較37°C 誘導環境為佳。此外,在 0.05-10 mM 等不同濃度 IPTG 誘導下,未含訊號序列的納豆菌漆酶活性表現均高於含有訊號序列的組別。
綜合本階段研究結果,在E. coli BL21 (DE3)/pEXP-5CT/TOPO® 系統中,以 0.05 mM IPTG 於 25°C 誘導表現未含有訊號序列的納豆菌漆酶,於胞膜間 區測得漆酶活性較高,達 367.9±18.3 mU/mL,亦顯著高於表現在 E. coli M15/pQE-30 Xa 系統所得活性,故挑選未含訊號序列組別 (E. coli BL21 (DE3)/pEXP-5CT/TOPO®-CotA only,後簡稱 TOPO-C) 進行後續外泌試驗。
3.2.3. 胞內納豆菌漆酶的釋放
前述試驗顯示,以大腸桿菌異源表現的納豆菌漆酶,即使未予修飾訊號
序列處理,仍可藉由冷滲透休克法分離大腸桿菌胞膜間區蛋白質,而測得目 標蛋白質活性;惟冷滲透休克法雖改善了傳統以超音波破菌等耗能方式取 得胞內蛋白質,卻必須維持在0°C 環境下進行分離,且過程中需多次離心、
振盪、回溶菌體及置換緩衝液,對於工業化生產有所不利。
依據Yang 等人的研究,倘於誘導時同時添加 glycine 及 Triton X-100,可 提升胞膜間區蛋白質的釋放 [10]。為使納豆菌漆酶得以直接釋放至胞外,
及考量未來以醱酵槽進行細胞高密度培養時的菌體穩定性,我們參考 Yang 等人的研究,在誘導表現TOPO-C 時,同時於培養基中添加 0.5-5% (w/v) 不 同濃度的glycine 或 Triton X-100,置於 25°C 下振盪誘導表現 18 小時,將 菌液離心後取上清液測量漆酶活性。結果如圖3 所示,添加 glycine 或 Triton X-100 於培養基,確實能將胞內納豆菌漆酶釋放至胞外,而添加濃度之影響,
以2% glycine 或 Triton X-100 的釋放效果最佳,所釋放至胞外的漆酶活性 分別為38.44±1.85 及 9.85±0.418 mU/mL。
以 SDS-PAGE 蛋白質電泳及活性染色分析法,檢視重組大腸桿菌胞外釋 放CotA laccase 如圖 4。帶有 cotA 基因質體的 TOPO-C 重組大腸桿菌,於 IPTG 誘導時同時添加 2% glycine 及 Triton X-100,取其胞外培養基進行 SDS- PAGE 蛋白質電泳及活性染色,對照未添加及未含有 cotA 基因質體的組別,
經 glycine 及 Triton X-100 處理的組別,在約 40 kDa 處呈現一明顯色帶反 應,並與使用冷滲透休克法分離之胞膜間區蛋白質反應一致,說明了經 glycine 及 Triton X-100 處理後,確能有效釋放累積於胞膜間區的 CotA laccase。
3.3. 漆酶釋放條件的最適化
根據前項結果,利用添加glycine 及 Triton X-100,可將原本僅能於大腸桿 菌胞內表現的納豆菌漆酶釋放至胞外。因此,希望進一步探討外泌生產納豆 菌漆酶的最佳條件,以下試驗之設計是針對生產過程中的最適誘導期及釋放
期分別探討。
3.3.1. 最適誘導期
為了解使用IPTG 誘導及 glycine、Triton X-100 釋放處理的最佳處理期間,
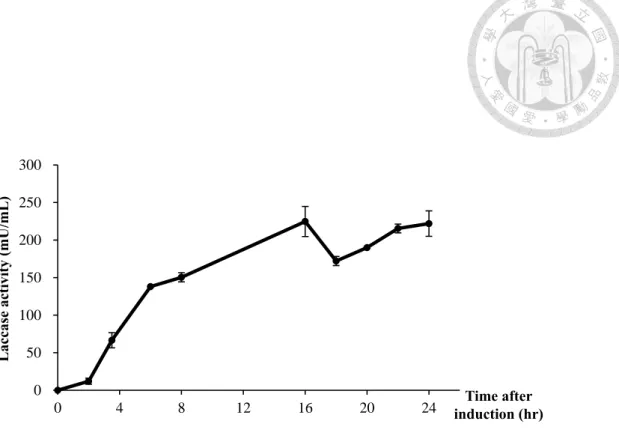
故根據前項結果,於誘導TOPO-C 表現 CotA laccase 時,同時添加 2% (w/v) glycine 及 Triton X-100,並於誘導開始後固定時間點取樣偵測釋放至培養基 內的漆酶活性。結果如圖 5 所示,在加入 0.05 mM IPTG 及 2% glycine、
Triton X-100 進行誘導和釋放後 16 小時,於培養基中測得最高漆酶活性 224.7
±11.6 mU/mL。
3.3.2. 最適釋放期
由前項試驗得知,以0.05 mM IPTG 誘導並以 2% glycine 及 Triton X-100 釋放TOPO-C 胞內納豆菌漆酶,最佳誘導期間為 16 小時。考量大腸桿菌於 誘導過程同時進行胞內物質釋放,會影響菌體正常生長,不利後續細胞高密 度培養時的菌體累積。因此,依據前述所得最佳誘導期間,改良目標蛋白質 釋放策略,先培養並以0.05 mM IPTG 誘導 TOPO-C 於胞內表現、累積 CotA laccase 16 小時,再以 2% glycine 及 Triton X-100 進行釋放。另考量工業化 生產時,倘先收集菌體集中釋放及縮小釋放體積,可提高所得目標蛋白質純 度、減少污染,及降低釋放劑 (glycine、Triton X-100) 使用量等優勢。因此,
本階段實驗調整於16 小時誘導後,先離心收集菌體,並以十分之一體積 (10 mL) LB 培養基回溶,再加入 2% glycine 及 Triton X-100 釋放,並於釋放後
固定時間取樣,觀察釋放至胞外的納豆漆酶活性變化。結果如圖6 所示,同
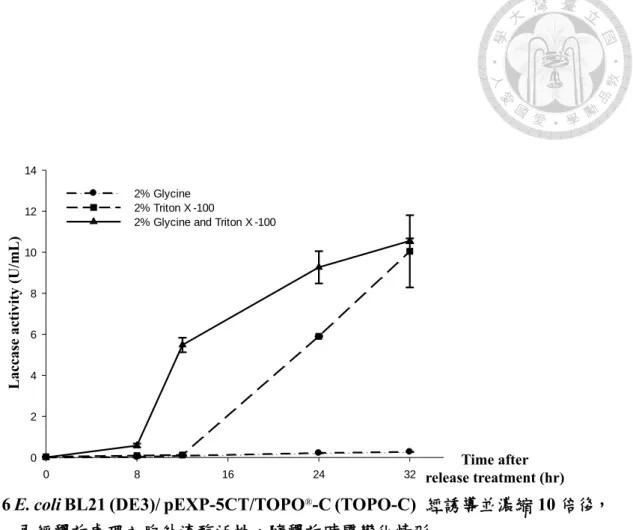
時以2% glycine 及 Triton X-100 處理的組別,其釋放至胞外的漆酶活性於處 理8 小時後較單以 glycine 或 Triton X-100 的組別明顯提升,並於處理 24 小 時後趨緩,32 小時後單位活性達 10.56±0.068 U/mL,換算 10 倍濃縮前單位 活性約1 U/mL。比較前項試驗結果,胞外漆酶產量確有提高。
3.4. 以細胞高密度培養策略外泌生產納豆菌漆酶
上述試驗已逐步最適化 TOPO-C 外泌生產納豆菌漆酶 CotA laccase 的相 關培養、誘導及胞外釋放條件。在此,進一步利用5 L 醱酵槽以細胞高密度 培養策略來提高重組大腸桿菌細胞數量,再以glycine 及 Triton X-100 釋放 CotA laccase 至胞外,以提升目標蛋白質生產效率及產量。
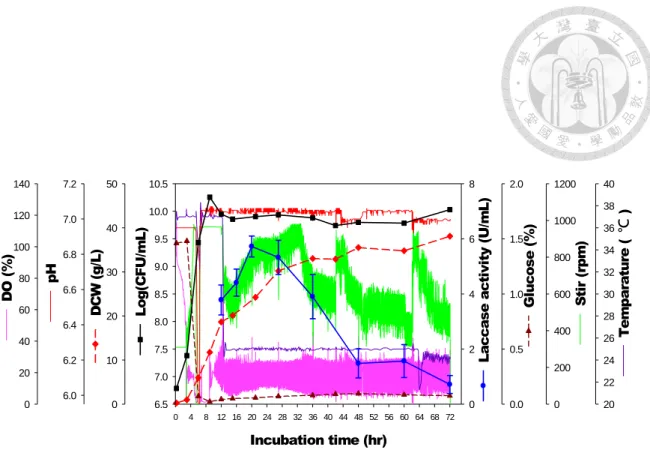
本階段試驗使用Biostat B (B. Braun, Sartorius, Germany) 5 L 醱酵槽,利用 pH-stat 批次饋料培養 (分別以 4N NaOH 及饋料培養基 (Nutrient solution) 控制 pH 值於 7.0 ),於 37°C 培養 TOPO-C 12 小時後,降至 25°C 並添加 IPTG 及 CuSO4進行CotA laccase 誘導生產,期間於固定時間點取樣並轉置 於50 mL Hinton’s flask,測試添加 2% glycine 及 Triton X-100 於 25°C 振盪 釋放處理24 小時後,測量培養基漆酶活性。結果如圖 7 所示,接種 9 小時 後,生菌數已提高至1.8×1010 CFU/mL,且隨著主培養基內碳源耗盡,pH 值
上升超過7.5,系統自動饋入饋料培養基平衡酸鹼值。此時,監測槽內殘糖
值自饋料後皆維持於 0.1%以下,顯示菌體可接續利用饋料培養基內葡萄糖
生長,使細胞乾重持續累積,細胞乾重最終 (接種後 72 小時) 累積至 38.1 g/L。
納豆菌漆酶外泌方面,經釋放處理後測得漆酶活性於加入 IPTG 誘導後 (接種後 12 小時) 開始累積,並以誘導後 8 小時 (接種後 20 小時) 最高,達 5.72±0.36 U/mL。之後,逐漸下降,誘導後 60 小時 (接種後 72 小時) 為 0.71
±0.19 U/mL。
第 4 章 討論
4.1. 訊號序列對 CotA laccase 生產與外泌的影響
4.1.1. 訊號序列效能評估
本研究選用第一型外泌系統訊號序列 E. coli J96 hlyAsp 與第二型外泌系 統訊號序列E. coli K12 torAsp、amiAsp、malEsp 及 ompAsp 等 5 種不同的訊 號序列,分別建構於 cotA 基因,藉以評估該等訊號序列對於 CotA laccase 在重組大腸桿菌系統的外泌生產效能。實驗結果顯示,Type I secretion system hlyAsp 訊號序列未能有效促使 CotA laccase 外泌至胞外,可能與本實驗所使 用的重組大腸桿菌本身缺乏-hemolysin (HlyA) pathway 相關的穿膜蛋白質 家族基因有關,而HlyA pathway 較常見於泌尿道感染型等致病大腸桿菌,
亦不適合進行重組蛋白質生產,故我們認為 Type I secretion system 並非為 用於外泌生產CotA laccase 的良好選擇。
Type II secretion system 方面,torAsp、amiAsp、malEsp 及 ompAsp 等 4 種 不同的訊號序列在 E. coli M15/pQE-30 Xa 與 E. coli BL21 (DE3)/pEXP- 5CT/TOPO® 系統中均未能促使目標蛋白質外泌,甚至影響 CotA laccase 在 胞膜間區的活性表現。顯示所選用的訊號序列並非最適,未來可朝嘗試其它 訊號序列方向努力。
4.1.2. CotA laccase 在胞膜間區 (periplasmic space) 的累積
在上述試驗中也發現,CotA laccase 在不修飾訊號序列的情況下,仍然能 在胞膜間區累積,推測可能藉由其它的途徑運送,或CotA 本身即含有未知 的訊號序列促使其運送至胞膜間區,相關機制與原因雖然尚不清楚,但卻也 開啟外泌生產的契機。
我 們 也 利 用 由 National Biomedical Research Foundation (NBRF) 及
University of Delaware (UD) 開發之 Protein Information Resource (PIR) 線上 系統 (http://pir.georgetown.edu/pirwww/search/comp_mw.shtml),預測 CotA 蛋 白質分子量為58,482.54 Da,與本實驗室先前以 SDS-PAGE 分析所得分子量 約70 kDa 相近,惟因本實驗利用活性染色方式分析,蛋白質樣品並未變性 處理,活性條帶位置為40 kDa。
4.2. CotA laccase 的釋放策略
4.2.1. 釋放策略的比較
本研究確認CotA laccase 能自然累積於胞膜間區,我們進一步利用相關策 略釋放胞內的納豆菌漆酶,達成外泌生產的目的。據Choi 等人的研究及整 理,欲將胞膜間區內的蛋白質釋放或運送至胞外,可利用冷滲透休克、添加 化學物質破壞膜狀結構、使用L-form cell、增加表現通道蛋白質或利用表面 系統等方式 [9]。考量未來以醱酵槽進行細胞高密度培養時的菌體穩定性,
我們分別比較了冷滲透休克法及glycine/Triton X-100 釋放法。
從結果看來,以冷滲透休克法及 glycine/Triton X-100 釋放法所釋出的漆 酶活性相近,但 glycine/Triton X-100 釋放法是利用細胞培養過程中添加促 進釋放物質的方式,可改善傳統以超音波破菌的耗能問題。在釋放時機的調 整上,亦較冷滲透休克法彈性,且沒有後者需要維持低溫、反覆置換溶劑及 回溶等程序,相當符合大腸桿菌異源表現並量化生產CotA laccase 的相關需 求,具有良好的可發展性。
4.2.2. 誘導與釋放策略的調整
本研究亦同時針對 glycine/Triton X-100 釋放法的最適化作細部探討,比 較了單獨使用glycine、Triton X-100,及將其合併使用的差異。發現,合併 兩者使用可大幅提升CotA laccase 的釋放,這個實驗結果大致與 Yang 等人
fusion protein sFV/TNF-(single chain FV fragment-tumor necrosis factor alpha fusion protein) 的研究結果相近。
然而,Yang 等人的研究是以 IPTG 誘導 E. coli 表現外源基因的同時,加 入glycine/Triton X-100 進行釋放,但研究中亦指出 glycine 和 Triton X-100 可能藉由增加膜通透性及破壞外膜結構的方式達到釋放效果 [10] 。但這樣 的處理也同時影響了細胞正常生長及菌體累積,不利工業化生產。因此,本 研究嘗試以「誘導」與「釋放」分開處理的改良方式,取代同步處理。經過 分開處理後,發現釋放至胞外的最高漆酶單位活性近1 U/mL,明顯高於同 步處理的最高漆酶活性224.7 mU/mL。顯示本研究的策略:先誘導 E. coli 生 產目標蛋白質並於胞內累積至一定量後,再利用 glycine/Triton X-100 進行 釋放,可在較不影響細胞生長的前提下,增加外源基因的表現與目標蛋白的 累積,進而提升整體產量。
另外,本研究也比較了「誘導」與「釋放」同步處理與分開處理下,單獨 使用glycine 和 Triton X-100 的釋放情形。單獨以 2% glycine 進行同步處理 時,所釋放漆酶活性為38.44 mU/mL,較單獨使用 2% Triton X-100 進行同 步處理所釋放的漆酶活性9.85 mU/mL 為高。但單獨以 2% glycine 進行分開 處理時,其釋放漆酶活性卻遠不及使用2% Triton X-100 進行分開處理的結 果。推測可能和兩者的作用機制差異有關。由於,glycine 是細菌細胞壁中 肽聚醣 (peptidoglycan) 生合成的前驅物,過量的 glycine 可能會影響細菌細 胞壁肽聚醣正常鍵結而使菌體型態異常造成細胞膜系統通透性失衡;而 Triton X-100 是界面活性劑 (detergent),可藉由直接破壞細胞膜結構達到釋 放胞內物質的作用。然而,在「誘導」與「釋放」同步處理下,E. coli 正處 於細胞快速生長期,在受IPTG 誘導生產 CotA laccase 的同時,glycine 也較 容易被代謝,進而加速CotA laccase 通透過細胞膜並釋放;但在細胞快速生
CotA laccase 產量不高。
反之,在「誘導」與「釋放」分開處理的情形,E. coli 經過 16 小時誘導 生產,已進入生長停滯期,對於glycine 的代謝能力有限,故較 Triton X-100 以直接破壞外膜,釋放胞膜間區累積的CotA laccase 效果為差。
綜合以上,本研究結合Yang 等人的研究成果,同時添加 glycine 和 Triton X-100 促進胞膜間區的釋放,並改採「誘導」與「釋放」分開處理的方式進 行,對於CotA laccase 釋放至胞外的效率及總量皆能有效提升。
4.3. 細胞高密度培養的成效與策進
4.3.1. 細胞高密度培養對 CotA laccase 外泌生產的提升
除了對於細胞的釋放效率外,重組 E. coli 的數量直接關係到外泌生產的 效能。因此,我們利用醱酵槽,配合以pH-stat 批次饋料策略進行細胞高密 度培養,藉由總菌量的提升,來拉高CotA laccase 產量。
結果顯示,以醱酵槽進行細胞高密度培養,從培養過程中菌體乾重持續的 累積可知其確實有效提高單位體積內菌體之含量。在外泌生產漆酶活性方 面,最高生產活性達5.72 U/mL,較 Hinton’s flask 振盪培養的結果為高。但 於培養後期,外泌生產的漆酶活性未能持續累積,反而逐漸遞減。進一步檢 視發現,培養後期 (接種 36 小時後) 槽內總生菌數雖未有明顯減少趨勢,
但經含抗生素 (100 ppm ampicillin) 培養基篩選後,具抗性的菌落數僅剩總
菌量約1%。推測可能原因為,培養過程中質體不穩定性所致。細胞分裂過
程中帶有 cotA 基因的質體並未平均分配於子代細胞,導致培養後期具有
CotA laccase 生產能力的菌體比例降低,加上培養最初添加的 ampicillin 受 溫度或光照分解破壞而失去篩選力,以致漆酶生產效能降低。
4.3.2. 未來策進方向
添加ampicillin 等抗生素,維持槽內環境的篩選力,避免未帶有目標基因的 菌體族群擴大,此問題應可獲得改善。此外,對於漆酶的異源生產,研究顯 示,在培養過程當中增加銅離子濃度,可提升漆酶活性 [36],但過量的銅離 子亦可能引起細胞毒性,故此部分尚待更詳細的研究與探討。
4.4. 結論
本研究針對納豆菌漆酶CotA laccase 在大腸桿菌異源表現系統內僅能以胞 內生產的限制予以改良,並探討利用訊號序列 (signal peptide) 修飾目標基因 及調整胞膜間區 (periplasmic space) 釋放條件等方式,外泌生產納豆菌漆酶的 可行性。研究結果顯示,納豆菌漆酶CotA laccase 在重組大腸桿菌內,毋需額 外修飾其它訊號序列,即能自然轉移至胞膜間區累積,此一現象有助於外泌生 產技術的開發。因此,進一步利用 glycine/Triton X-100 釋放法,並調整採用
「誘導」與「釋放」分開處理的方式,再搭配以醱酵槽進行細胞高密度培養策 略,建構了一個兼顧異源蛋白質「大量表現」及「高效釋放」的大腸桿菌外泌 生產模式,以此方式培養後於胞外釋放之納豆菌漆酶活性可達5.72 U/mL。較 原先胞內表現、並以超音波破菌的方式所得活性,提高近170 倍。故本研究所 建構的大腸桿菌外泌生產模式,除有效改善傳統破菌方式造成的能源耗費,對 於酵素工業化製程的簡化及生產成本的降低亦有所助益。
圖表
表 1 聚合酶連鎖反應引子
引子名稱 序列(5’-3’)
cotA-hlyA-1 TCCCCATAAAGCCTATGGAAGTCAGGGTAATCTTAATCCATTAATTAATGAAATCAGC
cotA-hlyA-2 AAATTACCTGCAGCTGAAATGATTTTGCTGATTTCATTAATTAATGGATTAAGAT
cotA-hlyA-3 AAATCATTTCAGCTGCAGGTAATTTTGATGTTAAAGAGGAAAGAGCTGCAGC
cotA-hlyA-4 GCATTACCGGACAACTGCAATAAAGAAGCTGCAGCTCTTTCCTCTTTAA
cotA-hlyA-5 TGCAGTTGTCCGGTAATGCCAGTGATTTTTCATATGGACGGAACTCAATAAC
cotA-hlyA-6 TTATTATGCTGATGCTGTCAAAGTTATTGAGTTCCGTCCATATG
pQE→cotA GCATGCGAGCTCGGTACCCCAATGACACTTGAAAAATTTGTGG
hlyA←pQE GCTTGGCTGCAGGTCGACCCTTATTATGCTGATGCTGTCAAAG
cotA→hlyA TCCCCATAAAGCCTATGGAAGTC
cotA←hlyA TTCCATAGGCTTTATGGGGATCAGTTATATCC
TorAF1 ATGAACAATAACGATCTCTTTCAGGCATCAC
TorAR1 CCCAGCAGATACGGCAGGAAC
AmiAF1 ATGAGCACTTTTAAACCACTAAAAACACTC
AmiAR1 AGTGGAAATAACTGATCACGCCTTC
MalEF1 ATGAAAATAAAAACAGGTGCACG
MalER1 ACTTCAGCGCTACGGCACCC
OmpAF1 ATGAAAAAGACAGCTATCGCGATT
OmpAF2 TCGATCTCTACGCGACGATCC
A-TorA-cotA CGAGCTCGGTACCCCAATGAACAATAACGATCTCTTTC
A-AmiA-cotA CGAGCTCGGTACCCCAATGAGCACTTTTAAACCAC
A-MalE-cotA CGAGCTCGGTACCCCAATGAAAATAAAAACAGGTGCAC
A-OmpA-cotA CGAGCTCGGTACCCCAATGAAAAAGACAGCTATCGC
cotA-6His GCTGCAGGTCGACCCTTAGTGATGGTGATGGTGATGTTTATGGGGATCAGTTATATCC
表 2 在 E. coli M15/pQE-30 Xa 系統中,修飾不同訊號序列之 cotA laccase 表現情 形
Induction
temp. hlyA torA amiA malE ompA cotA only
Vector only
Medium 25°C ND ND ND ND ND ND ND
37°C ND ND ND ND ND ND ND
Cell 25°C - ND ND ND ND 32.8 ND
37°C - ND ND ND ND ND ND
單位:mU/mL ND:Not detected
分別以hlyAsp、torAsp、amiAsp、malEsp 及 ompAsp 修飾之 cotA 基因於 E. coli M15/pQE-30 Xa 系統表現,並偵測不同誘導條件下,胞內 (超音波破菌) 及胞 外漆酶活性表現情形。
僅未修飾訊號序列的組別,經超音波破菌後測得胞內漆酶活性32.8 mU/mL。
圖 1 在 E. coli M15/pQE 30-Xa 系統修飾不同訊號序列,對納豆菌漆酶於胞膜間 區累積的影響
分別以torAsp、amiAsp、malEsp 及 ompAsp 修飾之 cotA 基因於 E. coli M15/pQE- 30 Xa 系統表現,並以冷滲透休克法分離胞膜間區蛋白質,並檢測該區域漆酶 活性表現情形。
未修飾訊號序列的組別於胞膜間區累積活性均較有修飾組別為高,為 93.4±
1.09 mU/mL。
0 20 40 60 80 100 120
torAsp amiAsp malEsp ompAsp cotA only
Laccase activity (mU/mL)
圖 2 在 E. coli BL21 (DE3)/pEXP-5CT/TOPO® 系統中,IPTG 濃度及溫度等誘導 因子,對含不同訊號序列之納豆菌漆酶,於胞膜間區累積的影響
分別以torAsp、amiAsp 及 malEsp 修飾之 cotA 基因於 E. coli BL21 (DE3)/pEXP- 5CT/TOPO®系統表現,並以冷滲透休克法分離胞膜間區蛋白質,並檢測該區 域漆酶活性表現情形。
以25°C 誘導效果普遍較 37°C 佳,且未修飾訊號序列組別活性亦較有修飾組 別高,其中活性最高者為0.05 mM IPTG 誘導組別,為 367.9±18.3 mU/mL。
0 20 40 60 80 100
0.05 0.5 1 5 10
0 100 200 300 400 500
0.05 0.5 1 5 10
torAsp amiAsp malEsp cotA only
Laccase activity (mU/mL)
37°C 25°C
IPTG concentration (mM)
圖 3 在 E. coli BL21 (DE3)/pEXP-5CT/TOPO®-C (TOPO-C) 誘導培養時,同時添 加不同濃度Glycine 或 Triton X-100,對於納豆菌漆酶釋放至胞外的影響 TOPO-C 於 500 mL Hinton’s flask 以 37°C 振盪培養至 OD600=0.8-1.0 後加入 0.05 mM IPTG 及 0.25 mM CuSO4以25°C 誘導,同時添加不同濃度 Glycine 或Triton X-100 進行釋放,並觀察胞外漆酶活性表現情形。
Glycine 或 Triton X-100 的最適添加濃度均為 2% (w/v),所釋放至胞外的漆酶 活性分別為38.44±1.85 及 9.85±0.418 mU/mL。
0 5 10 15 20 25 30 35 40 45
0.5 1 2 5
Glycine Triton X-100
Laccase activity (mU/mL)
Glycine/Triton X-100 concentration (%, w/v)
cotA vector - + + + - + + +
2% Glycine&
Triton X-100 - - + - - - + -
Cold osmotic
shock - - - + - - - +
圖 4 SDS-PAGE 膠體電泳及活性染色分析在 E. coli BL21 (DE3)/ pEXP- 5CT/TOPO®-C (TOPO-C),胞外釋放 CotA laccase 情形
無cotA vector 者為未轉形質體之 E. coli BL21(DE3)對照組。經 Glycine/Triton X-100 釋放 (編號 3),或以冷滲透休克法處理 (編號 4) 組別,於 40 kDa 可見 活性反應條帶。
M 1 2 3 4 1 2 3 4 M (kDa)
圖 5 E. coli BL21 (DE3)/ pEXP-5CT/TOPO®-C (TOPO-C) 誘導時同時添加 2%
Glycine 及 Triton X-100 後,納豆菌漆酶活性隨誘導時間釋放至胞外的變化 情形
TOPO-C 於 500 mL Hinton’s flask 以 37°C 振盪培養至 OD600=0.8-1.0 後加入 0.05 mM IPTG 及 0.25 mM CuSO4以25°C 誘導,同時添加 2% Glycine 及 Triton X-100 進行釋放,並以此為 0 點,定點取樣觀察胞外漆酶活性隨誘導及釋放 處理時間變化情形。
胞外漆酶活性於誘導、釋放後16 小時最高,為 224.7±11.6 mU/mL。
0 50 100 150 200 250 300
0 4 8 12 16 20 24
Laccase activity (mU/mL)
Time after induction (hr)
0 8 16 24 32 0
2 4 6 8 10 12 14
2% Glycine 2% Triton X -100
2% Glycine and Triton X -100
圖 6 E. coli BL21 (DE3)/ pEXP-5CT/TOPO®-C (TOPO-C) 經誘導並濃縮 10 倍後,
再經釋放處理之胞外漆酶活性,隨釋放時間變化情形
100 mL TOPO-C 於 500 mL Hinton’s flask 以 37°C 振盪培養至 OD600=0.8-1.0 後 加入0.05 mM IPTG 及 0.25 mM CuSO4以25°C 誘導 16 小時後離心並以 10 mL LB 培養基回溶菌體,再添加 2% Glycine、2% Triton X-100 以及 2% Glycine 和 Triton X-100 進行釋放,並以此為 0 點,定點取樣觀察胞外漆酶活性隨釋放處 理時間變化情形。
以2% Glycine 和 Triton X-100 共同處理所釋放漆酶效率最高,處理 32 小時後 活性達10.56±0.068 U/mL。
Laccase activity (U/mL)
Time after release treatment (hr)