國立臺灣大學理學院地質科學系暨研究所 碩士論文
Department of Geosciences College of Science
National Taiwan University Master Thesis
溫度調控微生物硫酸還原作用之硫同位素分化研究:
以 Thermodesulfobacterium 與 Desulfovibrio 菌株為例 Temperature controls of the sulfur isotope fractionation during sulfate reduction by Thermodesulfobacterium and
Desulfovibrio strains
孫嘉璘 Chia-Lin Sun
指導教授:林立虹 博士 Li-Hung Lin, Ph.D.
王珮玲 博士 Pei-Ling Wang, Ph.D
.中華民國 101 年 1 月
January, 2012
i
口試委員會審定書
ii
致謝
能夠完成一本論文並不容易,開始寫致謝更是千頭萬緒,兩年半的過程中受 到太多協助和鼓勵,感謝之意溢於言表。最感謝的莫過於兩位指導教授王珮玲和 林立虹老師,不論是在實驗或是生活上遇到的難題,總是給我支持與鼓勵,謝謝 老師的耐心與協助,真的很幸運可以成為兩位老師的指導學生。感謝海研所林曉 武老師和朱美妃老師抽空參與碩士論文口試,提供許多寶貴建議使論文更加完 整。
感謝實驗室的大家,謝謝小靜在寫論文這段時間的陪伴,不但幫助我進行硫 同位素的分析,還給我許多論文上的建議,幫我抒發煩悶緊張的心情。謝謝玟錦 作為我的趴呢,陪伴我度過許多做實驗的夜晚和不開心的時刻。謝謝婷文、文瑜、
永欣、季融、小英學姐、興麟學姐、韻如、劭璿、宛玲、369、韋萱、哲維和哲 銘在實驗上和論文過程中給我的幫助和滿滿的關懷。謝謝佳靜、悅婷和光忻在我 一開始進入實驗室時帶領我進行實驗。謝謝 Jeff 幫助我進行硫同位素分析,讓 我有初步的資料。總之,在實驗室的日子謝謝大家的陪伴與幫助,因為有你們做 實驗的時光並不孤單。
感謝在成大認識的大家,謝謝小花做我的室友和在台北奮鬥的戰友,謝謝佩 萱、皓安和佩芳等人總是的給我鼓勵和安慰。感謝由聖總是做我情緒的垃圾桶,
接受我最直接的情緒,但仍不厭其煩的給我鼓勵和建議。最後感謝我最親愛的家 人,謝謝你們不論如何都陪在我身邊,認為我是最好的。因為你們,讓我更加勇 敢和堅強。
碩士過程中的收穫和成長也許未必能完全表現在此本論文中,但卻是我最值 得珍藏的部分。
iii
摘要
硫酸還原菌會將海水中硫酸鹽還原成硫化氫,最終形成黃鐵礦被保存在沈積 物中,而硫酸鹽還原過程中同時伴隨有機物的氧化分解,因此硫酸還原作用被視 為影響碳-硫-鐵循環的重要過程之一。由於硫酸還原菌所進行的硫酸還原作用造 成硫同位素的分化,且分化程度與環境因子有關,因此地質記錄中硫酸鹽類礦物 沈澱與沈積型黃鐵礦的硫同位素成份可提供重要的地球環境變動訊息。
硫酸還原菌造成硫同位素分化程度差異的原因仍然不清楚,可能與物種差異 和細胞硫酸鹽還原速率有關,或是受到溫度、硫酸鹽濃度和電子供應者等因素所 影響。為解釋複雜的分化情形,前人提出細胞內部代謝步驟的概念模型,並加入 質量流的計算,用以推估微生物代謝過程對硫同位素分化程度的影響,其中硫酸 鹽進出細胞的通量與細胞內部硫酸鹽轉換的通量是影響硫同位素分化程度的重 要因子。
本研究利用培養嗜熱菌 Thermodesulfobacterium commune 接近株與嗜溫菌 Desulfovibrio gigas 探討溫度如何影響硫酸還原菌的生理代謝,並進而影響硫同 位素的分化程度。Thermodesulfobacterium commune 接近株的培養溫度範圍為 34-79 oC,最適生長溫度為 72 oC,而最高細胞硫酸還原速率發生在 77 oC,其同 位素分化範圍在8.2-31.6 ‰ 之間,在大多數培養溫度下硫同位素分化在 8.2-
18.9 ‰ 之間上下變動,僅在 68 oC 出現一高值。Desulfovibrio gigas 的培養溫度 範圍為 10-41 oC,最適生長溫度為 33 oC,而最高細胞還原速率發生在 41 oC,其 同位素分化範圍在 10.3-29.7 ‰ 之間,最大分化程度出現在最高溫以及最低溫的 培養,而最小分化程度則出現在中間溫度。兩菌株所產生的分化範圍與前人研究 相近,但與同種屬菌株相比則其最大分化皆大於前人研究結果,顯然菌株培養的 條件不同便會造成同位素分化程度的不同。此外,此二菌株之分化程度隨溫度的 變化趨勢並不相同,其變化並不符合前人根據細胞生理特性隨溫度變化之理論分
iv
化模型。將多重硫同位素分析結果與前人模式所建立的多重硫同位素分化網格比 較,顯示該網格並無法包絡本研究的分析結果,表示前人模式所建立的多重硫同 位素分化網格仍須重新檢討。
綜合上述討論可知,微生物硫酸鹽還原作用造成的硫同位素分化程度雖已有 各種實驗觀察和模型加以解釋,但仍有諸多不一致的分析結果,其中溫度雖為單 一環境因子,但對菌株的生理學特性可能並無系統性的影響,也因此難以預測其 造成硫同位素分化的程度變化。未來若能結合不同菌株之生理學特性對於環境因 子的反應與多重硫同位素分析,將可更進一步探討硫同位素分化程度與硫酸還原 菌之特定生長條件之間的關係。
關鍵字:硫酸還原菌、硫同位素分化、生長溫度
v
Abstract
Microbial sulfate reduction is a major mechanism driving anaerobic mineralization of organic matter in global ocean. While sulfate-reducing prokaryotes are well known to fractionate sulfur isotopes during dissimilatory sulfate reduction, unraveling the isotopic composition of sulfur-bearing minerals preserved in sedimentary records could provide invaluable constraints on the evolution of seawater chemistry and metabolic pathways. Variations in sulfur isotope fractionations are partly due to inherent differences among species and also affected by environmental conditions (e.g. sulfate abundance and temperature). Sulfur isotope fractionations caused by microbial sulfate reduction have been interpreted to be caused by a sequence of enzyme-catalyzed kinetic isotope fractionation steps. The fractionation factor mainly depends on (1) the sulfate flux into and out of the cell, and (2) the flux of sulfur compound transformation between the internal pools.
This study examined the multiple sulfur isotope fractionation patterns catalyzed by a thermophilic Thermodesulfobacterium-related strain and a mesophilic Desulfovibrio gigas over a wide temperature range. The Thermodesulfobacterium-related strain grew between 34 and 79 oC with an optimal temperature at 72 oC and the highest cell-specific sulfate reduction rate at 77 oC.
The isotope fractionation (ε34Ssulfate-sulfide) ranges between 8.2 and 31.6‰ with a maximum at 68 oC. The D. gigas grew between 10 and 45 oC with an optimal temperature at 30 oC and the highest cell-specific sulfate reduction rate at 41 oC.
The isotope fractionation ranges between 10.3 and 29.7 ‰ with higher fractionations at both lower and higher temperatures. The isotope fractionation causing by these two strains is similar to previous reports, but the maximum fractionation is greater than that by the same species. Apparently, the differences in growth conditions may cause the different isotope fractionation. In addition, the change of fractionation with temperature is different for the two strains and cannot be predicted by a standard model considering physiological characteristics of cells. The result of multiple sulfur isotope measurements in this study cannot be described by a sulfate reduction network, which calculated the Δ33S and δ34S values by assuming the equilibrium fractionation among internal steps. Indeed, the sulfate reduction network has to be reevaluated.
Although there are many experiments and several models to study the sulfur isotope fractionation by microbial sulfate reduction, but the result is not conclusive.
Temperature is one of the most important environmental factors, but it may not make systemic influence on the physiology of strains and also the isotope fractionation.
vi
Further studies regarding physiological responses to environmental factors with the multiple sulfur isotope analysis may probably offer a linkage between sulfate isotope fractionation and growth conditions by sulfate reducing microorganisms.
Keyword: microbial sulfate reduction, sulfur isotope fractionation, growth temperature
vii
目錄
口試委員會審定書... i
致謝... ii
摘要... iii
Abstract ... v
目錄... vii
圖目錄... ix
表目錄... xi
附表目錄... xii
第一章 緒論... 1
1.1 硫的地球化學特性... 1
1.2 海洋環境中硫循環與硫酸鹽異化還原作用... 1
1.3 硫同位素之地質記錄與其重要性... 3
1.4 硫酸還原菌造成硫同位素分化程度與其影響因素... 5
1.4.1 硫酸鹽濃度與硫酸鹽還原速率... 5
1.4.2 電子供應者... 6
1.4.3 溫度... 6
1.5 多重硫同位素分化係數... 7
1.6 硫酸鹽異化還原作用與其分化模型... 9
1.6.1 Rees 模型 ... 10
1.6.2 Farquhar 模型 ... 10
1.6.3 Brunner and Bernasconi 模型 ... 12
1.7 研究動機與目的... 13
第二章 研究方法... 14
2.1 培養條件... 14
2.2 溫度梯度反應槽 (thermal gradient block) ... 15
2.3 細胞觀察... 16
2.3.1 血球計數盤 (counting chamber) ... 17
2.3.2 分光光度計 (spectrophotometer) ... 17
2.4 硫酸鹽濃度測量... 18
2.5 同位素分析樣本之前處理... 18
2.5.1 硫酸鋇樣本... 18
2.5.2 硫化銀轉換... 19
2.6 氣相同位素比值質譜儀... 21
2.7 多重硫同位素分析與操作... 22
2.8 計算使用公式... 23
viii
2.8.1 細胞硫酸鹽還原速率 (cell-specific sulfate reduction rate) ... 23
2.8.2 細胞生長速率 (specific growth rate) ... 23
2.8.3 硫同位素分化計算... 23
第三章 結果... 24
3.1 細胞濃度與硫酸鹽濃度變化... 24
3.2 細胞生長速率與細胞硫酸鹽還原速率... 37
3.3 硫同位素分化... 39
第四章 討論... 42
4.1 細胞生理特性與溫度的關係... 42
4.2 硫同位素分化與前人研究比較... 42
4.3 細胞硫酸鹽還原速率與硫同位素分化的關係... 43
4.4 溫度與硫同位素分化的關係... 45
4.5 硫酸鹽還原作用之多重硫同位素分化模型... 47
4.5.1 多重硫同位素分化網格計算方法... 47
4.5.2 多重硫同位素分化網格的限制... 50
4.5.3 多重硫同位素分化網格的擴充... 54
4.6 利用多重硫同位素分化網格探討細胞新陳代謝之質量流... 63
4.7 相關研究與未來工作... 67
第五章 結論... 68
參考文獻... 69
附錄... 72
ix
圖目錄
圖1-1、海洋沈積物中的硫循環 ... 2
圖1-2、不同時期硫酸鹽類蒸發岩與沈積岩中黃鐵礦的硫同位素成份 ... 4
圖2-1、溫度梯度反應槽 ... 15
圖2-2、細胞生長曲線 ... 16
圖2-3、硫化銀轉換之蒸餾系統 ... 20
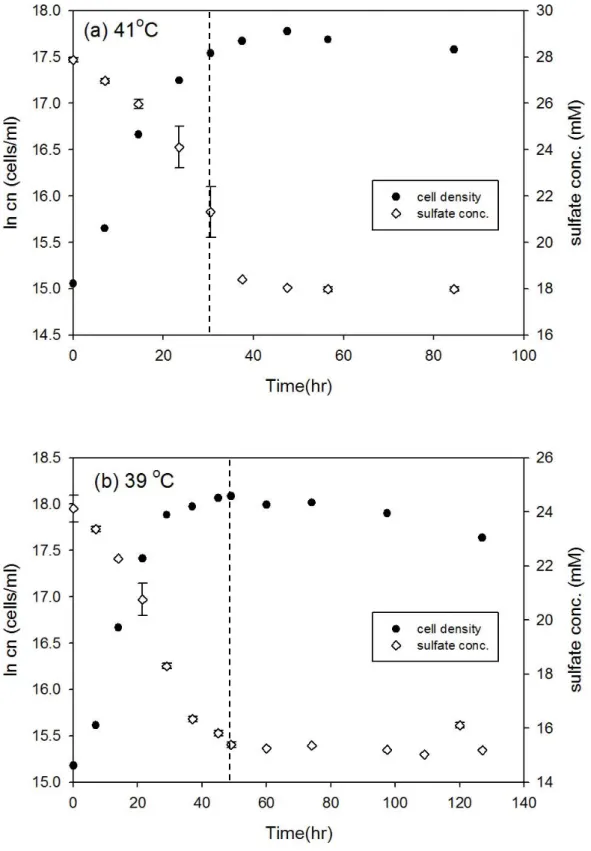
圖3-1、D. gigas 在 41 oC 和 39 oC 之細胞生長曲線與硫酸鹽濃度變化 ... 25
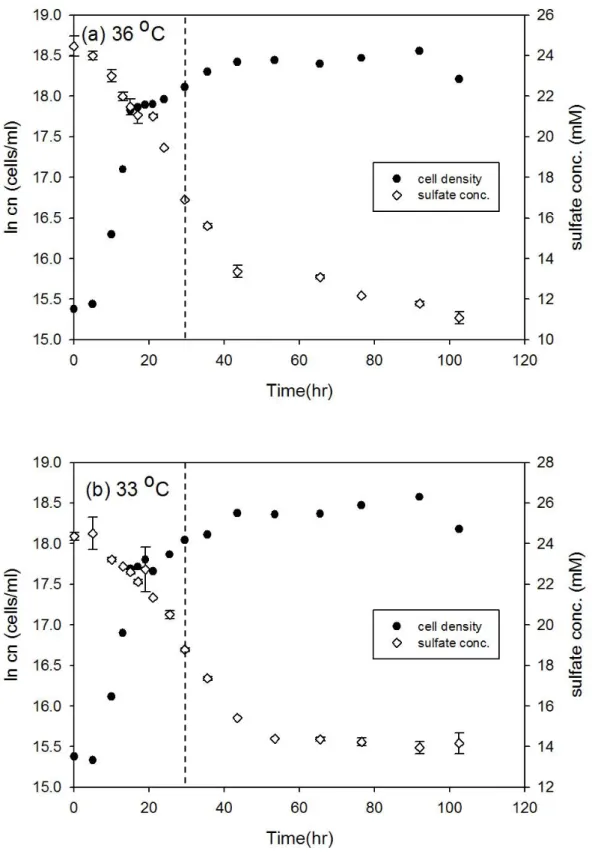
圖3-2、D. gigas 在 36 oC 和 33 oC 之細胞生長曲線與硫酸鹽濃度變化 ... 26
圖3-3、D. gigas 在 30 oC 和 27 oC 之細胞生長曲線與硫酸鹽濃度變化 ... 27
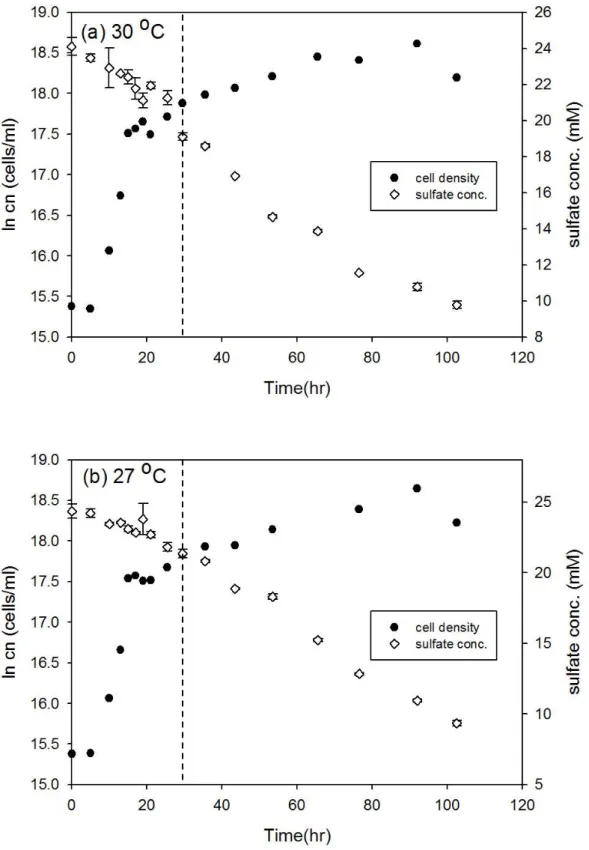
圖3-4、D. gigas 在 24 oC 和 21 oC 之細胞生長曲線與硫酸鹽濃度變化 ... 28
圖3-5、D. gigas 在 19 oC 和 16 oC 之細胞生長曲線與硫酸鹽濃度變化 ... 29
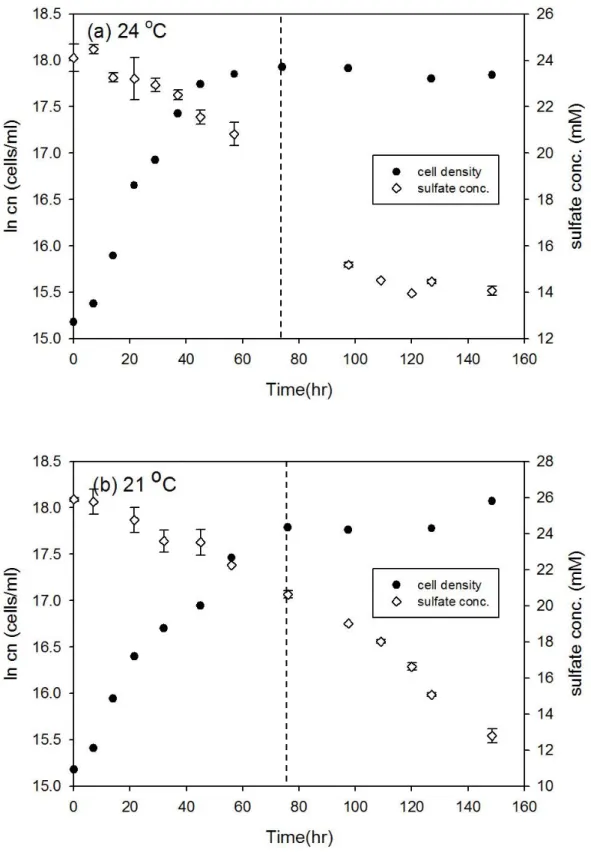
圖3-6、D. gigas 在 13 oC 和 10 oC 之細胞生長曲線與硫酸鹽濃度變化 ... 30
圖3-7、T. commune 接近株在 79 oC 和 77 oC 之細胞生長曲線與硫酸鹽濃度變 化... 31
圖3-8、T. commune 接近株在 72 oC 和 68 oC 之細胞生長曲線與硫酸鹽濃度變 化... 32
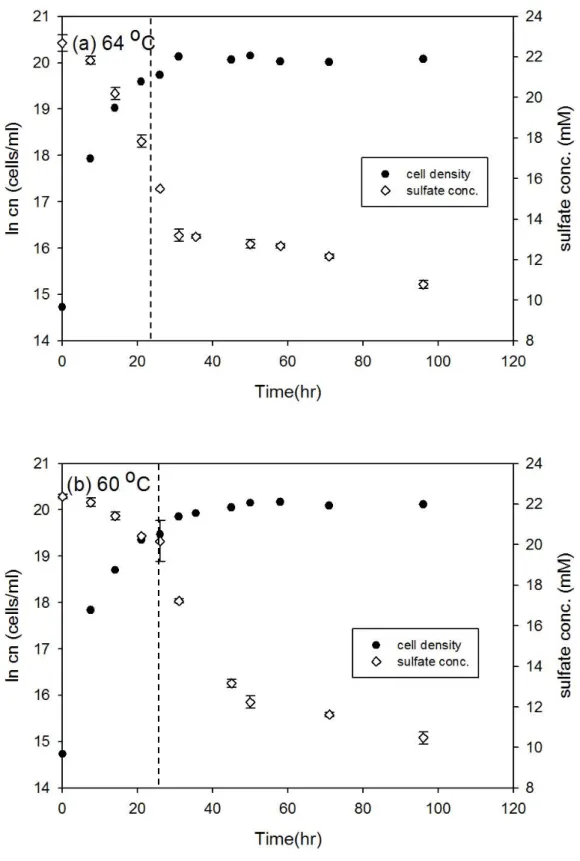
圖3-9、T. commune 接近株在 64 oC 和 60 oC 之細胞生長曲線與硫酸鹽濃度變 化... 33
圖3-10、T. commune 接近株在 55 oC 和 51 oC 之細胞生長曲線與硫酸鹽濃度變 化... 34
圖3-11、T. commune 接近株在 47 oC 和 45 oC 之細胞生長曲線與硫酸鹽濃度變 化... 35
圖3-12、T. commune 接近株在 40 oC 和 35 oC 之細胞生長曲線與硫酸鹽濃度變 化... 36
圖3-13、兩菌株之細胞生長速率與細胞硫酸鹽還原速率隨溫度的變化 ... 38
圖3-14、兩菌株產生之硫同位素分化 (34ε) 隨溫度的變化 ... 40
圖3-15、兩菌株產生之 Δ33S 值隨溫度的變化 ... 41
圖4-1、細胞硫酸鹽還原速率與硫同位素分化之間的關係 ... 44
圖4-2、不同 f3 和 f5 值與相對應分化大小的模型 ... 46
圖4-3、多重硫同位素分化網格 ... 52
圖4-4、不同溫度下之多重硫同位素分化網格 ... 53
圖4-5、33λ 值改變對於網格的影響 ... 58
圖4-6、 亞硫酸鹽轉化硫化物過程之分化大小對多重硫同位素網格的影響 ... 59
圖4-7、硫酸鹽轉化為亞硫酸鹽過程之分化大小對多重硫同位素網格的影響 ... 60
圖4-8、嗜熱菌 T. commune 接近株之多重硫同位素分化網格 ... 61
圖4-9、嗜溫菌 D. gigas 之多重硫同位素分化網格 ... 62
x
圖4-10、嗜溫菌 D. gigas 與嗜熱菌 T. commune 接近株之作圖結果 ... 65 圖4-11、Desulfobacterium autotrophicum 與 Desulfovibrio gigas 之作圖結果 ... 66
xi
表目錄
表4-1、平衡分化作用之分化係數與溫度的關係 ... 51
表4-2、建構分化網格使用之不同溫度的平衡分化係數 ... 51
表4-3、不同 33λ 值對於 33α3 和 33α4 的影響 ... 57
表4-4、分別調整 34α3 和 34α4 建構分化網格使用之分化係數 ... 57
xii
附表目錄
附表一、D. gigas 在不同溫度下之細胞濃度與硫酸鹽濃度隨時間變化 ... 72
附表二、T. commune 接近株在不同溫度下之細胞濃度與硫酸鹽濃度變化 ... 80
附表三、兩菌株之細胞生長速率與硫酸還原速率... 86
附表四、兩菌株之硫同位素分析資料... 86
附表五、兩菌株之硫同位素分化計算結果... 89
1
第一章 緒論
1.1 硫的地球化學特性
硫在自然界扮演重要的角色,不僅是地殼中重要的成分之一,所合成的蛋白 質對所有的植、動物和細菌都相當重要。硫的氧化還原態可從 -2 價的硫化物 (sulfide) 到 +6 價的硫酸鹽 (sulfate),在自然環境中主要以硫酸根離子 (SO42-)、
聚硫化物 (Sn2-)、元素硫 (S0) 和硫化物 (S2-) 的形態存在,可形成多種有機或無 機硫化合物。自然界中硫的儲存庫包括岩石圈、水圈、大氣圈與土壤圈,其中岩 石圈與海水為主要的硫存庫。在海洋環境中的缺氧條件下,海水中的硫酸鹽會經 由硫酸還原菌作用還原為硫化氫,為重要的生物地球化學作用之一。
1.2 海洋環境中硫循環與硫酸鹽異化還原作用
海洋環境為主要的硫儲存庫之一,其中的硫循環主要包含四個步驟:(1) 陸 地上岩石與沈積物中的硫化物和硫酸鹽經過風化和淋溶作用形成溶解態的硫酸 鹽,硫酸鹽經由河流運輸帶入海洋再進入海洋沈積物的孔隙水中,(2) 沈積物孔 隙水中的硫酸鹽經由微生物作用還原形成硫化物,(3) 硫化物可能被氧化形成元 素硫或是硫酸鹽而回到海水中,或是 (4) 與鐵氧化物反應形成黃鐵礦被保存在 海洋沈積物中 (圖 1-1) (Jørgensen and Fenchel, 1974)。海洋沈積物內進行的微生 物硫酸鹽還原作用不僅是轉變海水中硫酸鹽為硫化物的重要方式,海洋沈積物中 超過百分之五十的有機碳也會經由微生物硫酸鹽還原作用氧化分解,提供微生物 生存所需的能量 (Jørgensen, 1982)。黃鐵礦是微生物硫酸還原作用的產物,當保 存在沈積岩中的黃鐵礦經過岩石循環被抬升至地表時,會與氧氣和水作用而被風 化,產生硫酸鹽再度輸入海洋,因此微生物硫酸鹽還原作用不僅會影響硫循環,
對於碳循環與大氣中的氧氣含量也具有一定程度的影響,所以被視為全球生物地 球化學循環中重要的過程之一 (Berner, 1989; Canfield, 2004)。
微生物硫酸鹽還原作用由硫酸還原菌執行,硫酸還原菌廣泛的分布在含有硫
酸鹽 到1 度 還原 此過
同位 之間 變化
圖1 197
鹽的缺氧環 100 oC 以上
(hypersalin 原形成硫化 過程稱為硫
硫酸鹽異 位素分化,因 間硫同位素 化的重要依
1-1、海洋沈 74)。
環境中,對於 上 (Jørgense ne) 的環境 化物並且伴隨 硫酸鹽異化還
CH O
異化還原作用 因此硫酸鹽 素成份的差異 依據。
沈積物中的 Forming pyrite
S
於環境變化 en et al., 19 (Brandt et 隨有機物的 還原作用 (D
NH H 106H 用會造成反 鹽異化還原作
異,留在地質
的硫循環。說 Sulfide oxid
oxidation
2
化有很大的忍 992),而鹽度
al., 2001)。
的氧化分解,
Dissimilato PO 53 HCO 16N 反應物與產物
作用造成海 質記錄中的
說明請見內 Sulfate r bacteria Sulfate ation n
忍耐力,生 度則可從淡
硫酸還原菌 過程中會交 ory sulfate re
SO 14 NH HPO 物之間硫同 海洋中的硫酸 的此一變化被
內文 (修改自 eduction
S S
influx生存的溫度範 淡水 (freshw 菌的新陳代 交換八個電 eduction),
H
O 53H 同位素成分的
酸鹽與沈積 被視為了解
自Jørgensen
Sea water
ediment
範圍可從0 water) 到高 代謝會將硫酸 電子產生能量 反應如下式
S
的差異,產生 積物中的黃鐵 解早期地球環
n and Fench
oC 高鹽
酸鹽 量,
式。
生硫 鐵礦 環境
hel,
3
1.3 硫同位素之地質記錄與其重要性
沈積岩中含硫礦物的硫同位素組成與地球環境的氧化還原狀態有關,因此利 用長時間尺度的硫同位素地質記錄可以推測早期地球環境的變動 (Canfield and Teske, 1996; Habicht et al., 2002)。圖 1-2 為不同時期沈積的硫酸鹽與沈積岩中黃 鐵礦的硫同位素成份紀錄,依照硫同位素的變化情形可以將過去的地球環境分為 三個時期(圖中以虛線將三時期分隔),分別為 24 億年前、24 億年到 9 億年之 間和 9 億年前到現今。
第一時期沈積型黃鐵礦的硫同位素組成變化很小,硫酸鹽和黃鐵礦的硫同位 素成份的最大差異小於 10‰,此時地表環境的硫主要來自火山噴發與熱液作用 所釋出進入大氣與海洋中的硫化氫 (H2S) 和二氧化硫 (SO2),硫同位素的分化可 能受控於無機作用或是微生物硫酸還原作用。若是無機作用導致的同位素分化則 發生溫度需要高於 200 oC,顯然與沈積環境中的溫度條件不同,此外有證據顯 示在 3450 百萬年前硫酸還原菌就已經出現,因此推測此時期的硫同位素分化與 微生物作用有關 (Shen et al., 2001)。經由微生物作用若僅產生 10‰ 的硫同位素 分化,表示當時硫酸鹽異化還原作用是在低硫酸鹽濃度下進行 (硫酸鹽濃度小於 200 μM) (Habicht et al., 2002),因此藉由太古宙記錄的硫同位素變化範圍可推論 當時海洋中的硫酸鹽濃度遠低於現代,也隱含此時大氣中含氧量很低,不利於硫 酸鹽的形成。
第二時期黃鐵礦的硫同位素組成變化程度增加,與同時沈積硫酸鹽之硫同位 素組成的最大差異從 10‰ 增加到 40‰,硫同位素分化程度的增加可能受到當 時全球氧化事件影響 (Holland et al., 2006),當時大氣中的氧氣濃度上升使得保存 在沈積岩中的硫化物經由風化作用氧化形成硫酸鹽進入海水中,提高海水中硫酸 鹽的濃度,硫酸鹽異化還原作用若在硫酸鹽濃度較高時進行,其硫同位素分化程 度較大,因此硫同位素分化程度的增加反映當時海水中硫酸鹽濃度 (>200 μM) 與大氣中氧氣含量的增加。
4
第三時期黃鐵礦的硫同位素組成變化可達 55‰,代表地表環境與現今地球 類似,硫酸鹽異化還原作用仍為硫循環中主要的過程,而微生物對硫的自身氧化 還原作用 (sulfur disproportionation) 也參與其中,因而增大了硫酸鹽與黃鐵礦的 硫同位素組成差異 (Canfield and Thamdrup, 1994)。
沈積岩中含硫礦物的硫同位素組成不僅紀錄著地球環境的變動,從上述資料 我們也可發現即使同時期所沈積的黃鐵礦其分化大小也會有所不同,因此是什麼 因素調控硫酸鹽異化還原作用所造成的分化大小,值得被進一步的探討與研究。
圖1-2、不同時期硫酸鹽類蒸發岩與沈積岩中黃鐵礦的硫同位素成份 (修改自 Shen et al., 2001)。每個菱形表示各時期沈積岩中黃鐵礦的硫同位素成份,上方的 兩條線段表示當時海水中硫酸鹽的硫同位素成份,而下方線段則表示與海水中硫 酸鹽硫同位素成份差距 55‰,以虛線區分三個階段,分別為 24 億年以前、24 億 年前到 9 億年前和 9 億年前到現今地球。
5
1.4 硫酸還原菌造成硫同位素分化程度與其影響因素
關於硫酸還原菌造成的硫同位素分化程度,前人已經進行過相當多的研究,
主要利用嗜溫菌 Desulfovibrio spp. 和 Desulfotomaculum spp. 進行硫同位素分 化實驗 (Canfield et al., 2006; Detmers et al., 2001; Johnston et al., 2005; Kaplan and Rittenberg, 1964; McCready, 1975),發現在純菌培養過程中,硫同位素分化程度 可從3 ‰到 46 ‰。造成不同分化程度的原因仍不清楚,可能受到不同菌種之間 的差異、細胞硫酸鹽還原速率、溫度、硫酸鹽濃度或是電子供應者的類型等因素 影響 (Canfield et al., 2006; Detmers et al., 2001; Habicht et al., 2002; Harrison and Thode, 1958; Hoek et al., 2006; Johnston et al., 2007; Kaplan and Rittenberg, 1964;
McCready, 1975; Mitchell et al., 2009),但是除了硫酸鹽濃度的影響較為明確之外,
其他因素如何系統性的影響硫同位素分化仍不清楚或仍有爭議,以下將分節說明 各種因素對於硫酸還原菌造成硫同位素分化程度的影響研究。
1.4.1 硫酸鹽濃度與硫酸鹽還原速率
研究顯示當硫酸鹽濃度很小時 (<200μM),不論是嗜高溫或中溫的純菌株 或是自然環境的群落,這些硫酸還原菌造成硫同位素的分化程度會因此受到 抑制而變小 (Habicht et al., 2002)。在多數的研究結果中硫酸鹽還原速率與硫 同位素分化程度之間具有反比的關係,當硫酸鹽還原速率低時,會造成較大 的硫同位素分化,相反的若是硫酸鹽還原速率快時,產生的硫同位素分化程 度較小,而此關係通常存在於對於特定菌株的研究,在改變培養溫度、電子 提供者種類或濃度時,同時造成硫酸鹽還原速率的改變,因而產生分化程度 與還原速率之間的負相關性 (Hoek et al., 2006; Kaplan and Rittenberg, 1964;
Sim et al., 2011)。然而此一關係未必存在於所有硫酸鹽還原菌株當中,在部分 研究結果中兩者之間並未具有相關性(Canfield et al., 2006; Detmers et al., 2001;
Johnston et al., 2007) 。
6
1.4.2 電子供應者
硫酸還原菌進行硫酸還原作用所使用的電子供應者與其造成硫同位素分 化的關係,可能與該電子供應者是否進行完全氧化有關。前人研究發現當電 子供應者若被完全氧化 (有機碳氧化產生二氧化碳) 產生的硫同位素分化較 大 (>15 ‰),若是進行不完全氧化 (有機碳氧化形成醋酸) 產生的硫同位素分 化則較小 (<18.7 ‰),但其機制為何仍需要進一步的研究 (Detmers et al., 2001)。除此之外,部分電子供應者對於硫同位素分化的影響與硫酸鹽還原速 率有連帶相關性,若電子供應者為較複雜的有機碳則會造成較慢的反應速率,
使得硫同位素分化程度增大 ( Sim et al., 2011)。顯然電子接受者為影響硫同位 素分化重要因素之一,然而大部分研究中所提供的電子供應者多為較易分解 的有機碳,包括乳酸、醋酸、丙酮酸鹽、葡萄糖和酵母萃取物,但是複雜的 有機碳 (如:醇類) 同樣可作為硫酸還原菌的電子供應者 (Laanbroek et al., 1982) 卻很少在實驗中被使用。雖然特定菌種可以使用的電子接受者不同,但 特定環境可提供的電子接收者可能也有所限制,因此分析硫酸還原菌利用不 同種類電子供應者進行的硫同位素分化,可幫助我們進一步了解電子供應者 對於硫同位素分化的影響。
1.4.3 溫度
溫度被視為影響硫同位素分化程度的重要因素之一,對單一菌株而言,
細胞生長速率與硫酸還原速率隨溫度都有特定的變化,細胞生長速率在最適 生長溫度下達到最大值,硫酸鹽還原速率則會大致隨溫度上升而上升,但硫 同位素分化程度隨溫度的變化則無規律性的變化。前人實驗中利用溫度梯度 反應槽進行溫度對於硫同位素分化程度影響,使用的菌種包括嗜溫菌 (Desulfovibrio desulfuricans 和 Desulfobacterium autotrophicum) 與嗜熱菌 (Thermodesulfatator indicus 和 Archaeoglobus fulgidus),電子供應者包括氫氣、
乳酸鹽和醋酸。培養結果顯示硫同位素分化隨溫度變化具有不同的趨勢,
7
Thermodesulfatator indicus 和 Desulfovibrio desulfuricans 產生的較大程度分化 發生在最低溫和最高溫培養,最小則發生在中間溫度 (Canfield et al., 2006;
Hoek et al., 2006),Archaeoglobus fulgidus 和 Desulfobacterium autotrophicum 產 生的硫同位素分化程度雖然會有小幅的上下跳動,但大致上會隨著溫度升高 而下降 (Johnston et al., 2007; Mitchell et al., 2009)。
綜合前人研究可知硫酸還原菌產生的硫同位素分化程度受到許多因素所影 響,因此需要藉由細胞分化模型配合微量硫同位素 (Δ33S) 的測量才能夠進一步 討論分化程度改變的原因,以及這些變化的控因。
1.5 多重硫同位素分化係數
早期硫酸鹽異化還原作用的硫同位素分化的研究中僅使用兩種主要硫同位 素 (32S 和34S) 進行分析討論 (Rees .,1973),後期研究顯示利用多重硫同位素分 析 (32S、33S 和 34S) 配合硫酸鹽異化還原作用的分化模型,可提供更多關於細 胞內部質量流變化的資訊 (Farquhar et al., 2003)。下文將簡介多重硫同位素分析 使用的符號與各符號之間的關係。
硫具有四種穩定同位素 (32S, 33S, 34S, 36S),在自然界的豐度分別為 32S 佔 94.93%、33S 佔 0.76%、34S 佔 4.29% 和36S 佔 0.02%,物質中硫同位素組成 以δ3xS 表示 (如下式),其中 3x 為 33、34 或 36,標準品為美國亞利桑那 州代阿布洛峽谷 (Canyon Diablo) 鐵隕石中的隕硫鐵,簡稱 CDT。
δ S S/ S
S/ S CDT 1 1000
物質間硫同位素分化的程度以分化係數 α (fractionation factor) 表示,為物質之 硫同位素比值的比值,下式為分化係數的定義,其中 3x 亦可為 33、34 或 36。
αA B RA
RB, R XS S 分化大小也會以 ε 表示,ε 與 α 之間的關係式為:
εA B 1000 αA B 1
8
同位素之間質量的差異為產生分化的主要原因,含硫的化合物經歷同位素分 化作用時,會受控於硫同位素之間質量的差異,因此 δ33S、δ34S 和 δ36S 之間具 有一定比值關係。λ (dependency factor) 值用以描述分化反應造成反應物與產物 之 δ33S、δ34S 和 δ36S 之間的相關性(如下式),λ 值會因不同反應過程而改變,
但是變化通常很小 (Johnston et al., 2007)。
λ ln(1 δ SA
1000 ) ln(1 δ SB 1000 ) ln(1 δ SA
1000 ) ln(1 δ SB 1000 )
λ ln(1 δ SA
1000 ) ln(1 δ SB 1000 ) ln(1 δ SA
1000 ) ln(1 δ SB 1000 )
將地球上絕大多數樣品的 δ34S 分別對其 δ33S 和 δ36S 作圖則會呈現良好 的線性關係,關係大約如下式,此線性關係被定義為參考分化線 (RFL, Reference Fractionation Line),而此線性的斜率則被選定為 0.515 和 1.90。
δ S 0.515 δ S δ S 1.90 δ S
由於有些測量樣本中微量硫同位素的分析結果與 RFL 有些微的偏差,此種微小 的偏差來自分化過程中涵蓋了動力學和平衡分化的過程 (如: 微生物作
用)(Farquhar and Wing, 2003),使得 λ 值不恰好為 0.515 或 1.90,或是產生分 化的反應為光化學反應,因此為了方便描述這偏差,以Δ3xS (capital delta) 表示 δ33S 或 δ36S 與 RFL 之間的偏差,之間的關係如下式:
∆ S δ S 0.515 δ S
∆ S δ S 1.90 δ S
也可使用下式表示,其中 33λ 和 36λ 設定為 0.515 和 1.90。
∆ S δ S 1000 ﹝﹛1 δ S
1000﹜ λ 1﹞
∆ S δ S 1000 ﹝﹛1 δ S
1000﹜ λ 1﹞
9
1.6 硫酸鹽異化還原作用與其分化模型
不同的外在條件會影響細胞內部的質量流,藉由硫酸鹽異化還原作用的概念 模型,能夠幫助我們了解外在因素如何影響硫酸還原菌細胞內部質量流與最終產 生的同位素分化大小之間的關係。早期認為硫酸鹽異化還原作用僅包含兩個步驟,
分別為硫酸鹽進入細胞內與硫酸鹽還原成硫化物,兩步驟可產生的分化程度差異 很大,因此藉由兩步驟反應速率的差異可產生不同程度的分化 (Thode et al., 1951)。
Harrison and Thode (1958) 提出硫酸鹽異化還原作用可分為三步驟,每個步 驟的同位素分化程度不同,其中步驟一和步驟二為可逆反應。步驟一為硫酸鹽進 入細胞的過程,可以經由擴散作用或是由細胞主動吸收,皆會造成些微的同位素 分化。當硫酸鹽經由擴散作用進入細胞體內,較輕的硫同位素 (32S) 擴散速度較 快,造成細胞內部的硫酸鹽會富集較輕的硫同位素,產生硫同位素分化。另一方 面,當硫酸鹽不斷進入細胞內部會使得細胞內部電荷轉為負值,造成硫酸鹽無法 再自由通過細胞,因此細胞會開始主動吸收陽離子 (例如:鈉離子),同時會產 生複合體 (complex),複合體形成時偏向使用較重的硫同位素 (34S) 產生硫同位 素的分化。硫酸鹽以不同方式進入細胞會造成細胞內部硫酸鹽硫同位素數值變大 或變小,但不論硫酸鹽以何種方式進入細胞,其所造成硫同位素分化較其他步驟 小。步驟二為細胞內部硫酸鹽還原形成亞硫酸鹽的過程,當硫酸鹽進入細胞當中,
細胞內的硫酸鹽會經由酵素作用還原形成亞硫酸鹽 (SO32-),過程中造成硫同位 素分化的主要原因來自於 32S 與 34S 與氧原子之間鍵結力的不同,氧原子與
32S 之間的鍵結力比起與 34S 之間的弱,因此酵素作用時,32S-O 之間的鍵結 較易被破壞,造成產物亞硫酸鹽富集 32S,反應物硫酸鹽則會富集 34S。步驟三 為亞硫酸鹽還原形成硫化氫並排出細胞體外,細胞體內的亞硫酸鹽和氫分子作用 被還原形成硫化氫,並將硫化氫排出細胞體外。
根據 Harrison and Thode (1958) 所提出概念,陸續有更加詳盡與量化的模型
10
被提出,以下分節敘述之。
1.6.1 Rees 模型
Rees (1973) 根據 Harrison and Thode (1958) 所提出的模型,經過修改提 出詳細的硫酸鹽異化還原作用在細胞內部的反應路徑及其相對應的分化係數 (34α):
SO (SO APS SO H S 1 括弧表示細胞膜,數字表示不同的步驟。此模型中包含四個主要步驟:(1) 硫 酸鹽穿過細胞膜進入細胞,硫酸鹽進出細胞為可逆反應 (Cypionka, 1995),此 步驟產生的硫同位素分化較小,約 3‰ 到 0‰,(2) 細胞內部的無機硫酸鹽 經由三磷酸腺苷 (adenosine triphosphate,ATP) 活化形成腺苷-5'-磷酸硫酸酐 (adenosine-5'-phosphosulfate,APS),(3) APS 中硫與氧鍵結斷裂,APS 還原 形成亞硫酸鹽,(4) 經由酵素作用使亞硫酸鹽還原形成硫化氫。步驟 (2) 和 (3) 皆為可逆反應,雖然硫酸鹽活化成為腺苷-5'-磷酸硫酸酐的過程沒有硫同位素 分化產生,但仍有 -22‰ 到 -25‰ 的硫同位素分化分別發生在 APS 還原形 成亞硫酸鹽和亞硫酸鹽還原為硫化物的過程中。由Rees 模型可得知最終硫同 位素分化的大小與哪一步驟是限制步驟有關,若硫酸鹽進入細胞成為限制步 驟,則會表現出最小的硫同位素分化,相反的最大同位素分化則發生在限制 步驟為細胞內部硫酸鹽還原的可逆反應 (Canfield et al., 2006; Hoek et al., 2006)。
1.6.2 Farquhar 模型
由於 Rees 模型僅可定性去推測細胞新陳代謝步驟對硫同位素分化的影 響,因此 Farquhar 模型便將 Rees 模型加入了質量流 (mass flow) 與多重硫 同位素的概念,定量考慮反向反應的分流對同位素分化的影響,幫助我們了 解微生物作用對硫同位素分化的影響 (Farquhar et al., 2003)。此模型假設微生 物硫酸鹽還原反應為一連串熱力學平衡的分化反應的組合,33λ 值設定為
11
0.5145,分化係數則為溫度函數。
SO (SO
, , , ,
APS SO
, ,
, , , ,
H S
括弧表示細胞膜,數字表示不同的步驟,φ 為質量流,α 為分化係數,根據此 模型定義了兩個主要影響分化結果的過程,分別為硫酸鹽進出細胞質量流的 通量與細胞內部硫酸鹽轉換的通量 (Canfield et al., 2006)。硫酸鹽進入細胞為 可逆反應,因此硫酸鹽進入細胞質量流的通量等於最後產生硫化氫的通量加 上反向流出細胞外硫酸鹽的通量,關係式如下:
φ φ φ f3為硫酸鹽經由微生物作用形成硫化氫的比例:
f φ
φ φ
當f 趨近於 1 時,表示進入細胞當中的硫酸鹽大部分還原形成亞硫酸鹽,僅有 少部分再以硫酸鹽的形式離開細胞 (φ2 ≒ 0),當f 相當小時,表示大部分進 入細胞當中的硫酸鹽都以硫酸鹽的形式反向離開細胞 (φ2 很大),或是最終產 生硫化氫的通量很小 (φ3 很小)。由於硫酸鹽形成 APS 不會產生硫同位素分化,
因此將APS 省略直接以硫酸鹽還原形成亞硫酸鹽代表細胞內部硫酸鹽轉換的 通量,此為第二個主要影響硫同位素分化的過程,這過程同樣為可逆反應,
因此形成亞硫酸鹽的通量會等於亞硫酸鹽再氧化為硫酸鹽的通量加上最後產 生硫化氫的通量,關係式如下:
φ φ φ
f5表示在細胞內部的亞硫酸鹽形成硫化氫的比例:
f φ
φ φ
當f5相當大時,表示亞硫酸鹽大部分還原形成硫化氫 (φ5 ≒ 0),當 f5趨近於 0 時,表示大部分的亞硫酸鹽又再氧化形成硫酸鹽,僅少量形成硫化氫 (φ3 很 小)。設定 f3和f5並採用適當的分化係數,即可建立多重硫同位素分化網格,
12
測量分化作用產生的多重硫同位素組成,便可依此網格推測微生物硫酸鹽還 原作用中不同微生物內部步驟作用如何表現出不同微量硫同位素組成的特 色。
1.6.3 Brunner and Bernasconi 模型
部分研究發現硫酸鹽異化還原作用產生的分化可大於前人模型中所預估 的最大值46‰ (Rudnicki et al., 2001; Sim et al., 2011),而較大的硫同位素分化 被認為與三硫磺酸鹽的產生有關,因此Brunner and Bernasconi (2005) 再次修 正模型,考慮亞硫酸鹽還原時的其他複雜路徑。當亞硫酸鹽還原時並非直接 形成硫化氫,而是經由許多中間物質 (S3O62-, S2O32-) 變化所產生,這些反應 被稱為「trithionate pathway」(Akagi, 1995),trithionate pathway 會使得硫同位 素分化增大,但亞硫酸鹽還原過程究竟是單一反應或是由多個反應組合形成,
仍無法確定 (Rabus et al., 2006)。Trithionate pathway 共有三個反應式:
3SO H 4H S O 3H O S O H S O SO 2H
S O H H S SO
雖然反應更加複雜,但整體的微生物硫酸鹽還原模型仍與前人相近,模型如 下:
SO (SO APS SO S H S H S
牽涉 trithionate pathway 的步驟皆假設為可逆反應,硫化氫可同時存在於細胞 內外,在細胞內部的硫化氫可再氧化形成中間物質或是運輸出細胞體外 (Brunner et al., 2005)。上述不同步驟具有不同的分化係數:34α1=1.003、34α4b
=0.975、34α6=0.947 和34α7=1.005,α7 定義為亞硫酸鹽轉變為中間物質 (S2O32-、S3O62-) 與中間物質氧化為亞硫酸鹽過程中所產生的硫同位素分化 (Brunner and Bernasconi, 2005)。根據此模型推導最大硫同位素分化可達 75‰,
超過純菌培養得到的同位素分化結果,能夠解釋環境和地質記錄中所觀察到 的變化 (Johnston et al., 2007),但由於生物體內的中間物質是否存在或者是否
13
每種硫酸鹽還原菌都含有此中間物質仍無法確定,因此此模型仍需要更多的 驗證。
1.7 研究動機與目的
溫度不僅會影響硫酸鹽還原中酵素的反應速率,也能夠影響硫酸鹽在細胞內 外的交換速率,因此被認為扮演硫同位素分化的重要角色,但以往研究對於溫度 如何系統性的影響硫同位素分化仍無一致的看法,因此本研究利用培養嗜熱菌 Thermodesulfobacterium commune 接近株與嗜溫菌 Desulfovibrio gigas 進行代謝 反應物的多重硫同位素測量,並配合硫酸鹽異化還原的分化模型和質量流計算,
希望建立溫度與硫同位素分化之間的關係,以了解硫酸鹽還原菌對硫同位素分化 程度影響的機制。
14
第二章 研究方法
2.1 培養條件
本研究進行培養的兩菌株為生長在不同溫度範圍下的硫酸鹽還原菌,分別為 Desulfovibrio gigas 和 Thermodesulfobacterium commune 接近株,皆為絕對厭氧 菌。Desulfovibrio gigas 由潟湖環境中純化出來,最適生長溫度在 30-35 oC (Gall, 1963),Thermodesulfobacterium commune 接近株特性與 Thermodesulfobacterium commune 相似,後者生長的溫度範圍在 45-85 oC,最適生長溫度為 70 oC (Zeikus et al., 1983)。
兩者使用相同的培養基,培養基的配方為每公升溶液中加入:1.17 g 氯化鈉 (NaCl)、0.4 g 氯化鎂 (MgCl2×6H2O) 、0.3 g 氯化鉀 (KCl)、0.15 g 氯化鈣 (CaCl2×2H2O)、0.27 g 氯化銨 (NH4Cl)、0.1 g 磷酸二氫鉀 (KH2PO4)、3 g 硫酸 鈉 (Na2SO4)、1 g 酵母萃取 (yeast extract)、10 ml 微量元素 (TES) 溶液和 10 ml 1 M 乳酸 (Lactic acid)。將其混合均勻以 121 oC 滅菌 20 分鐘,冷卻前放入厭氧 箱靜置一夜除去氧氣,最後加入20 ml 2%硫化鈉 (Na2S) 和 20 ml 1 M 碳酸氫鈉 (NaHCO3) 並且利用經由 0.22 μm 過濾後的鹽酸與氫氧化鈉調整培養基的酸鹼 值 (pH) 至 7.5。
每次進行溫度與硫同位素分化大小實驗前,會先將兩菌株分別以 60 oC 和 30 oC 培養至指數生長期使細胞具有最大活性,再以 1:10 的比例將菌液混入全 新培養基中,最後將其放入溫度梯度反應槽內開始進行培養。進行培養時,監測 菌株的細胞數目和硫酸鹽濃度隨時間的變化,當細胞進入指數生長期時,終止細 胞生長並進行同位素分析樣品取樣。
15
2.2 溫度梯度反應槽 (thermal gradient block)
溫度梯度反應槽為長方形鋁塊所製成,反應槽兩端分別為加熱與冷卻系統,
藉此產生溫度梯度,冷卻系統所使用的冷凝機可設定溫度為 -2 到 20 oC,若設 定溫度過高,會使冷凝機水管破裂,造成系統故障,加熱系統則無條件限制 (圖 2-1)。溫度梯度反應槽共有 65 個孔洞 (橫向 13 孔、縱向 5 孔),縱向的 5 個孔 洞為相同溫度, 5 管菌株培養,5 管培養中包含管用來監測細胞與硫酸鹽濃度、
同位素取樣的二重覆和備用取樣,橫向的每個孔洞之間溫度差距約 4-5 oC,因此 利用溫度梯度反應槽進行不同溫度下的菌株培養,可提高對溫度的解析度。
Thermodesulfobacterium commune 接近株培養溫度範圍為 35-83 oC,進行實 驗時加熱器與冷卻器分別設定為 88 oC 與 15 oC,Desulfovibrio gigas 培養溫度 範圍為10-43 oC,進行實驗時加熱器與冷卻器分別設定為 47 oC 和 3 oC。使用溫 度梯度反應槽前,需將反應槽提前一天開啟,使溫度達到平衡,使用前需再次確 認各孔洞溫度。
圖2-1、溫度梯度反應槽。為長方形鋁塊所製成,反應槽兩端分別為加熱 (圖中 左側) 與冷卻 (圖中右側) 系統藉此產生溫度梯度,提高對於溫度的解析度。
16
2.3 細胞觀察
細胞數量計算的目的在於建立菌株的生長曲線,當菌株被移到新鮮培養基時,
細胞濃度隨時間的變化可被畫成一曲線,稱為生長曲線 (圖 2-2),生長曲線包括 延滯期 (lag phase)、指數生長期 (log phase)、停滯期 (stationary phase) 和死亡期 (death phase)。本研究建立生長曲線的目的除了要計算菌株在不同溫度下的生長 速度之外,也便於決定何時要終止細胞生長,進行同位素分析。
圖2-2、細胞生長曲線。細胞生長曲線共分為四期,分別為延滯期、指數生長期、
停滯期和死亡期,表示細胞生長狀況。延滯期 (Lag phase) 時細胞尚未進行分 裂,細胞數沒有增加。指數生長期 (Log phase) 時大部分之細胞都已做好分 裂之準備,開始進行分裂,因此產生細胞濃度快速增加之情形。停滯期
(Stationary phase) 時進行分裂之細胞與死亡之細胞數目達成平衡,或大部分 之細胞為維持本身之新陳代謝而停止分裂,故整體看來,細胞數目並無增加。死 亡期 (Death phase) 時環境變成對細胞有害,例如培養基的養分用盡,或因細 胞本身之代謝廢物排出而使環境出現毒素時,細胞便會大量死亡,細胞數目減少,
進入死亡期。
17
2.3.1 血球計數盤 (counting chamber)
利用血球計數盤配合光學相位差顯微鏡可在固定體積下直接計算細胞數 目。血球計數盤由厚玻璃所製成,通常血球計數盤有二個計數槽 (chamber),
每個計數槽中細刻 9 個 1 mm2之大正方形, 其中位於 4 個角落之正方形再 細刻 16 個小格,深度均為 0.1 mm。利用血球計數盤進行細胞數目的計算,
所觀察的菌液細胞濃度不可過高或過低,若細胞濃度過低則計數統計時的誤 差加大,若細胞濃度過高則會造成數菌不易,因此進行細胞計數前需將菌液 稀釋至適當濃度。
將適當濃度的細胞菌液滴在血球計數盤上,覆上蓋玻片並輕微按壓蓋玻片 四周,使多餘的溶液能流到方格之外,靜置 1-2 分鐘待細胞不隨著溶液流動,
就可用光學相位差顯微鏡計算方格範圍內細胞數目。細胞濃度 (cn,單位為 cells/ml) 的計算為 方格範圍內細胞數目 (Xcell/square),乘以稀釋倍率 (DF),除 以計數範圍方格體積(Vsquare),算式如下:
cn X / DF V
2.3.2 分光光度計 (spectrophotometer)
分光光度計可產生特定波長的光源,光源透過測試樣品會有部分被吸收,
即為樣本的吸光值,由於樣本的濃度會與吸光值成正比,因此藉由分光光度 計測量樣品的吸光值就可得知樣本的濃度。實驗過程中,分光光度計為觀察 細胞濃度最快速的方法,因此本實驗進行菌株培養時以分光光度計監測細胞 濃度隨時間的變化,最初需要先建立吸光值與細胞數目之間的檢量線,利用 血球計數盤計算不同細胞濃度下確切的細胞數目與分光光度計所測得的吸光 值對照,可得知細胞數目與吸光值之間的關係。
分光光度計開機前需暖機 15 分鐘,待光源穩定後,波長設定為 660 nm,
取 1 ml 的樣本加入 0.1 ml 的 6 N 鹽酸裝入比色管中,操作時手握比色管非 透明處,測量前需先將比色管中的氣泡除去,避免影響測量值,每次測量為
18
每 2 秒測一次重複 5 次後取平均值,最後將測量到的吸光值換算為細胞數 目。
2.4 硫酸鹽濃度測量
測量樣品中硫酸鹽濃度,需先將樣品中的微生物與硫化氫去除,再使用離子 層析儀進行分析。培養終止時樣本中加入 20% 醋酸鋅,使樣本中的硫化物沈澱 形成硫化鋅,如此可以避免硫化物再氧化形成硫酸鹽而影響測量值,待硫化鋅完 全沉澱後,離心並過濾除去硫化鋅與細胞,將上清液稀釋至適當濃度,利用離子 層析儀進行分析,求得硫酸鹽的濃度。離子層析儀 (ion chromatography) 的原理 為當樣本通過離子交換管柱時,利用不同離子和管柱之間的正負交換吸引力不同,
來達到各離子間分離的效果,將各離子經由交換管柱所分離出來的時間差和得到 的訊號值與標準品比對,藉此可得知樣本中所含的離子種類與其濃度大小。
2.5 同位素分析樣本之前處理
本研究利用氣相同位素比值質譜儀 (gas isotope ratio mass spectrometer) 進 行多重硫同位素分析,由於氣相同位素比值質譜儀分析使用六氟化硫作為進樣氣 體,因此我們需先將硫同位素樣本進行前處理轉換為硫化銀,以便與氟氣反應產 生六氟化硫。硫同位素樣本的前處理,以下分節進行詳細的介紹。
2.5.1 硫酸鋇樣本
進行同位素樣本取樣時,將 20% 醋酸鋅以 1:10 的比例加入其中,使樣 本中的硫化氫沈澱形成硫化鋅,接著離心過濾除去硫化鋅與細胞,離心過濾 後的樣本再加入鹽酸酸化使酸鹼值低於 4,以確保無硫化氫殘留,最後加入 過量的氯化鋇與樣本中的硫酸鹽進行反應形成硫酸鋇沈澱,清洗烘乾後的硫 酸鋇再轉化為硫化銀。
19
2.5.2 硫化銀轉換
依照 Thode et al (1961) 所提出的操作方法將硫酸鋇轉換為硫化銀,其原 理是利用蒸餾系統 (圖 2-3) 在高溫下使還原溶液 (Thode 溶液) 與硫酸鹽反 應將硫酸鹽還原形成氣態硫化氫,再將硫化氫導入醋酸鋅溶液當中產生硫化 鋅沈澱,最後硫化鋅再與硝酸銀反應製成硫化銀。實驗步驟如下:
1. 秤重約 20-30 毫克的乾燥硫酸鋇樣本,將其顆粒磨細後放入圓底反應瓶中,
加入20 ml 的 Thode 溶液,開啟排煙櫃。
2. 組裝蒸餾系統,磨砂頭塗矽凝膠 (silicon gel) 並加上磨砂接頭夾,旋轉接 頭確定其密合度。玻璃管 (b) 和 (c) 中分別加入適量的去離子水和 5 ml 1 M 的醋酸鋅,經由反應所產生的氣態硫化氫用外徑 1/8 英吋之鐵氟龍 (PTFE) 管通氮氣導入玻璃管 (c) 中,使其與醋酸鋅反應產生沈澱。
3. 連接氮氣瓶,開啟氮氣並以較高流速通氣至少 20 分鐘,通氣 20 分鐘後,
將氣流量調整至每秒產生 2-3 顆氣泡。開啟加熱包和冷卻水,觀察並調整 氣泡速度,直到溶液開始沸騰冒泡 (約 100 oC),繼續觀察直到確定通氣的 速度穩定沒有變化。
4. 反應完畢約需 12-14 小時,確定圓底反應瓶內沒有殘餘任何硫酸鋇粉末。
結束反應前,增加通氣速度持續 20 分鐘,讓殘留在系統中的硫化氫能夠 全部帶出。
5. 通氣 20 分鐘後,利用原有醋酸鋅溶液沖洗吸附在玻璃管壁上的硫化鋅,
加入 0.1 N 硝酸銀數毫升產生硫化銀,所需體積視反應的硫酸鋇重量而定 (1 mg 硫酸鋇需要 0.1 mL 1 N 硝酸銀,再增加 1-2 倍),關緊蓋子使用強 力震盪器 (votex) 幫助反應完全,此時白色沈澱物變成黑色,溶液變成淡 棕色。
6. 將玻璃管蓋子打開,放進乾浴槽加熱至沸騰,維持五分鐘,此時硫化銀顆 粒會增大,溶液變的透明,代表加熱已經足夠。冷卻後待硫化銀顆粒沈澱,
20
用塑膠滴管盡量吸出多餘的澄清液。
7. 待多餘澄清液移除後,加入 1-2 mL 的濃氨水,關緊蓋子使用強力震盪器 讓氨水與硫化銀顆粒混和均勻,靜置一晚後,在排煙櫃內盡量吸出多餘氨 水,加入去離子水離心至少清洗三次 (3000 rpm、5 分鐘),使氨水氣味完 全清除。最後將硫化銀放入烘箱烘乾,秤重並計算回收率。
8. Thode 溶液的配製:取 816 mL 的濃鹽酸、48% 的氫碘酸 500 mL 和 50%
的次磷酸 245 mL 混合,放入攪拌子與蓋上裱玻璃後置於加熱板上加熱至 沸騰,持續沸騰 1.5 個小時使任何含硫物質揮發,待冷卻後儲存於暗色玻 璃瓶中。
圖2-3、硫化銀轉換之蒸餾系統。蒸餾系統包括:圓底反應瓶 (a)、分別裝去離 子水和醋酸鋅的玻璃管 (b 和 c)、ㄇ型管 (d)、冷凝管 (e)和 Y 型管 (f)。
d
e
f
21
2.6 氣相同位素比值質譜儀
質譜儀的基本原理是將樣本離子化後通過一磁場,帶電離子運動方向會受磁 場的影響而產生偏轉,偏轉的程度受離子運動的速度、離子質量和磁場強度這三 種因素所影響,如下列公式:
r 2Vm B q
上式中r 為離子偏轉軌跡的半徑,V 為離子加速器之電壓,B 為磁場強度,m 為 離子質量,q 為帶電數目。進行同位素分析時,將加速電壓與磁場強度固定,所 以若離子的質量越輕,偏轉會越大,相反的若質量越重則會偏轉越小,利用不同 質量具有不同的偏轉程度並設定接收器就可以測出樣品中同位素之間的比例。
本研究中所使用的為氣相同位素質譜儀,待測樣本需先轉換成氣體再進入質 譜儀進行分析,目前硫同位素測量所使用的氣體有六氟化硫 (SF6) 與二氧化硫 (SO2) 兩種,其中六氟化硫中的氟氣僅有一個穩定同位素 (19F),不同於二氧化 硫中的氧具有三個穩定同位素 (16O、17O 和 18O),因此使用六氟化硫可以直接 測得 33S/32S、34S/32S 和 36S/32S 等比值而免去校正的困難與誤差,因此本研究 使用六氟化硫做為進樣氣體,以便進一步分析微生物硫酸鹽還原作用對多重硫同 位素分化的影響。
將待測樣本轉化為六氟化硫需要經過高溫氟化系統,使樣本與氟氣反應,反 應方程式如下:
Ag S 4F 2AgF SF
接著使用氣相層析儀將六氟化硫氣體純化,收集後利用氣相同位素質譜儀進行多 重硫同位素分析。在同位素質譜儀分析方面,本研究所使用的氣相質譜儀 (MAT253) 為雙進樣系統 (dual-inlet system),重複分析國際標準品以求得參考氣 體 (reference gas) 的同位素組成,未知組成樣品的同位素組成便可透過與參考氣 體比較求得,儀器對於 δ34S 的分析誤差為 0.01‰。
22
2.7 多重硫同位素分析與操作
多重硫同位素分析的實驗步驟如下:
1. 取出 2.5-3 毫克的硫化銀樣本包入錫囊當中,確認硫化銀樣本不會從錫囊中 掉出,防止樣本丟入錫管中時會有掉落的硫化銀吸附在錫管壁中,以及避免 氟氣剛進入錫管中時與硫化銀樣本反應過於激烈而分散,導致反應不完全。
2. 將包好的硫化銀樣本放入乾淨的鎳管中,鎖回反應系統上,開始抽真空並進 行測漏,待真空度降至0.002 torr,加熱鎳管至 100 oC,放置隔夜,此步驟 是為了將樣本中的水氣移除。
3. 反應隔夜後,關掉加熱包,待溫度降為室溫,進行測漏後將鎳管抽至真空。
達到真空後,使用液態氮將鎳管冷凝,接著放氟氣進入鎳管中,關閉反應管 待壓力穩定,加熱鎳管至 200 oC,反應放置隔夜,使氟氣與硫化銀樣本能 夠反應完全。
4. 反應隔夜後,關掉加熱包,待溫度降為室溫,清除多餘的氟氣,將產生的六 氟化硫氣體通過氣相層析儀 (gas chromatography),利用分離管柱將六氟化 硫與氟化過程中產生的雜氣分離,並將六氟化硫氣體濃縮收集在玻璃管中,
以便進行硫同位素分析。
5. 六氟化硫進樣前需先進行質譜儀系統穩定度測試,並將質譜儀內管路抽至真 空,之後將裝有六氟化硫的玻璃管連接上質譜儀進樣端,開啟閥門,靜置 10 分鐘,待玻璃管連接質譜儀的管路抽至真空,再開啟玻璃管的開關。
6. 六氟化硫進樣至質譜儀內時,每個閥門的開啟與關閉,都需靜置 3-5 分鐘,
讓氣體能夠達到平衡,避免產生分化。進行分析前需先利用可壓縮的風箱 (bellow) 將樣本氣體與參考氣體的氣壓調整至相似,才開始進行分析。最後 經過校正計算得到樣本相較於國際標準品的硫同位素比值。
23
2.8 計算使用公式
2.8.1 細胞硫酸鹽還原速率 (cell-specific sulfate reduction rate)
本實驗計算的細胞硫酸還原速率為細胞在生長指數期時,兩時間點 (t1和 t2) 之間每個細胞消耗硫酸鹽的速率。計算公式如下:
csSRR fmol · day/cell SO SO cn cn
2 · T T
〔SO42-〕為硫酸鹽濃度,cn 為細胞濃度,t 為時間 2.8.2 細胞生長速率 (specific growth rate)
細胞在指數生長期的生長速率為複製時間的倒數,此數值越大表示細胞生 長速率越快,此環境越適合生長。計算公式如下:
μ ln cn ln cn t
cn0 為指數生長期開始時的細胞濃度 (cells/ml),cn 為指數生長期任一點的細 胞濃度 (cells/ml),t 為時間 (hr)。
2.8.3 硫同位素分化計算
微生物在密封的血清瓶 (serum vial) 中進行硫酸鹽還原作用,產物不會流 失減少,可以視為一封閉系統,因此利用瑞利分化模型 (Rayleigh fractionation model) 計算微生物硫酸鹽還原過程中所產生的硫同位素分化 (34εsulfate-sulfide):
δ SSOT εlnf δ SSOT f SO T
SO T
δ SSOT 為進行取樣時硫酸鹽的硫同位素組成,δ SSOT 為初始硫酸鹽的硫 同位素組成, SO T 為進行取樣時硫酸鹽的濃度, SO T 為初始硫酸鹽 的濃度。
24
第三章 結果
3.1 細胞濃度與硫酸鹽濃度變化
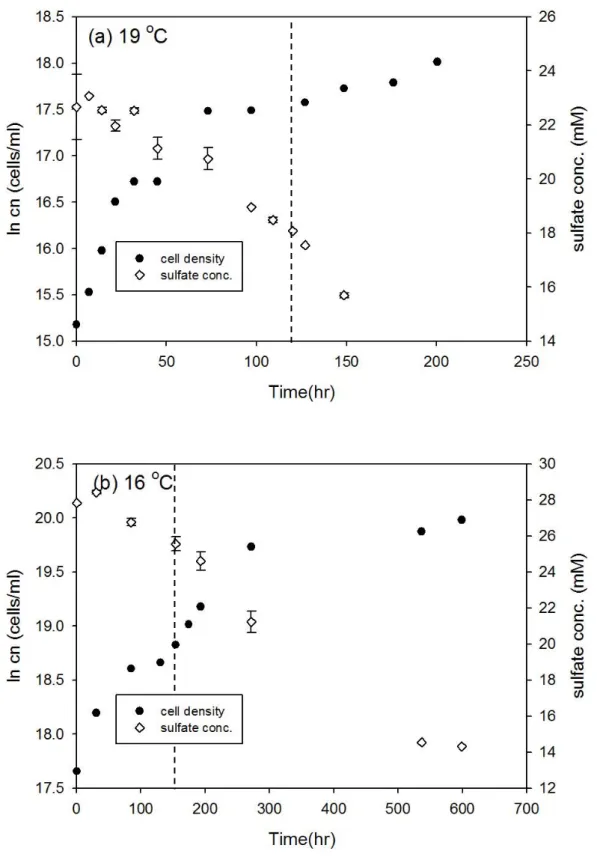
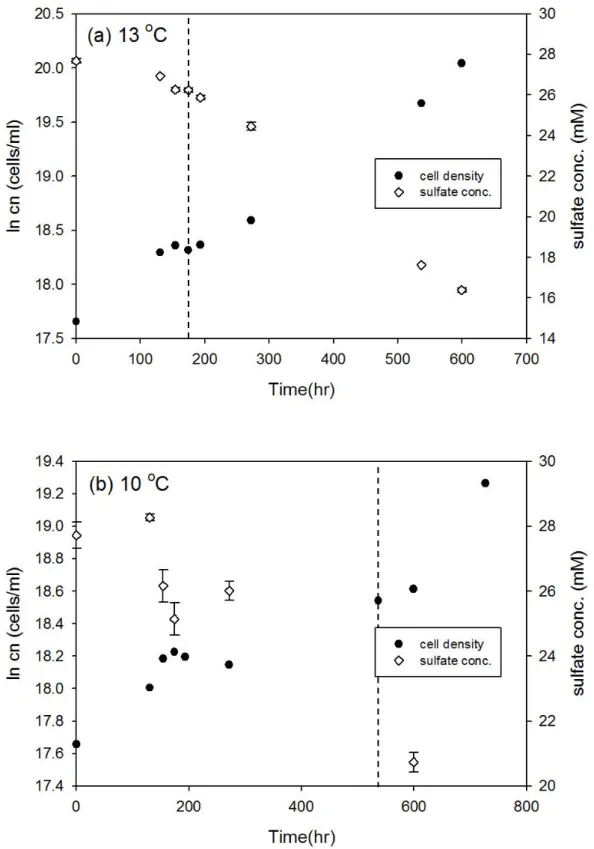
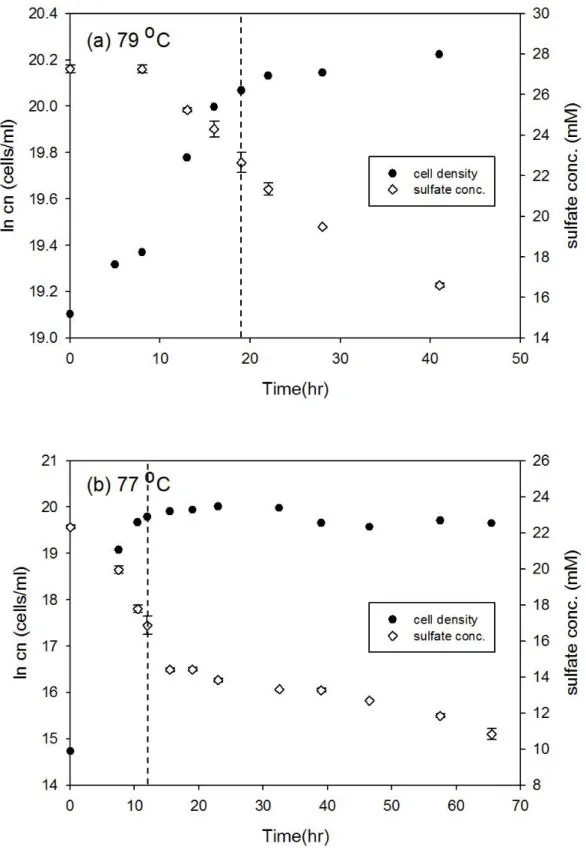
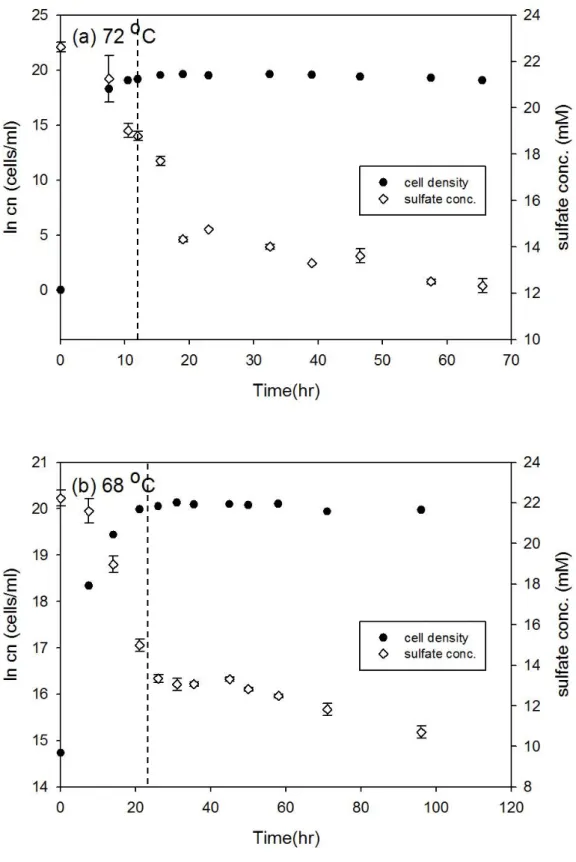
Desulfovibrio gigas 和 Thermodesulfobacterium commune 接近株在大部分培 養溫度下細胞濃度隨時間的變化符合典型生長曲線,但延滯期與指數期的時間長 短卻不相同 (圖 3-1 至圖 3-12),此外由於本研究探討指數生長期硫酸還原菌所造 成的硫同位素分化,因此並無持續紀錄細胞生長進入死亡期的細胞濃度變化。以 Desulfovibrio gigas 在 39 oC 的培養為例,細胞濃度的增加會伴隨硫酸鹽濃度減 少,當細胞生長由延滯期進入指數生長期時,硫酸鹽濃度會大幅的減少,而當細 胞生長進入停滯期時,硫酸鹽濃度變化的幅度也會減小 (圖 3-1b)。本研究結果 中在大部分培養溫度之間的細胞濃度與硫酸鹽濃度變化與上述舉例相同,然而在 Desulfovibrio gigas 的 10 oC 和 13 oC 培養雖然細胞濃度有隨時間增加 (圖 3-6),
但是生長曲線缺乏明確的指數期,Thermodesulfobacterium commune 接近株在 35 oC 和 40 oC 培養則不見細胞數量的增加和硫酸鹽濃度的明顯下降 (圖 3-12)。
25
圖3-1、D. gigas 在 41 oC 和 39 oC 之細胞生長曲線與硫酸鹽濃度變化。實驗中 細胞濃度的誤差為 10%,在 41 oC 和 39 oC 的同位素分析取樣時間分別為 30.5 和 49 小時 (以虛線表示)。
26
圖3-2、D. gigas 在 36 oC 和 33 oC 之細胞生長曲線與硫酸鹽濃度變化。實驗中 細胞濃度的誤差為 10%,在 36 oC 和 33 oC 的同位素分析取樣時間分別為 24 和 29.5 小時 (以虛線表示)。
27
圖3-3、D. gigas 在 30 oC 和 27 oC 之細胞生長曲線與硫酸鹽濃度變化。實驗中 細胞濃度的誤差為 10%,在 30 oC 和 27 oC 的同位素分析取樣時間分別為 29.5 和 29.5 小時 (以虛線表示)。
28
圖3-4、D. gigas 在 24 oC 和 21 oC 之細胞生長曲線與硫酸鹽濃度變化。實驗中 細胞濃度的誤差為 10%,在 24 oC 和 21 oC 的同位素分析取樣時間分別為 74 和 76.5 小時 (以虛線表示)。
29
圖3-5、D. gigas 在 19 oC 和 16 oC 之細胞生長曲線與硫酸鹽濃度變化。實驗中 細胞濃度的誤差為 10%,在 19 oC 和 16 oC 的同位素分析取樣時間分別為 121 和 130.5 小時 (以虛線表示)。
30
圖3-6、D. gigas 在 13 oC 和 10 oC 之細胞生長曲線與硫酸鹽濃度變化。實驗中 細胞濃度的誤差為 10%,在 13 oC 和 10 oC 的同位素分析取樣時間分別為 174.5 和 536.5 小時 (以虛線表示)。
31
圖3-7、T. commune 接近株在 79 oC 和 77 oC 之細胞生長曲線與硫酸鹽濃度變 化。實驗中細胞濃度的誤差為 10%,在 79 oC 和 77 oC 的同位素分析取樣時間 分別為 19 和 12 小時 (以虛線表示)。
32
圖3-8、T. commune 接近株在 72 oC 和 68 oC 之細胞生長曲線與硫酸鹽濃度變 化。實驗中細胞濃度的誤差為 10%,在 72 oC 和 68 oC 的同位素分析取樣時間 分別為 12 和 23.5 小時 (以虛線表示)。
33
圖3-9、T. commune 接近株在 64 oC 和 60 oC 之細胞生長曲線與硫酸鹽濃度變 化。實驗中細胞濃度的誤差為 10%,在 64 oC 和 60 oC 的同位素分析取樣時間 分別為 23.5 和 26 小時 (以虛線表示)。
34
圖3-10、T. commune 接近株在 55 oC 和 51 oC 之細胞生長曲線與硫酸鹽濃度變 化。實驗中細胞濃度的誤差為 10%,在 55 oC 和 51 oC 的同位素分析取樣時間 分別為 36 和 39.5 小時 (以虛線表示)。
35
圖3-11、T. commune 接近株在 47 oC 和 45 oC 之細胞生長曲線與硫酸鹽濃度變 化。實驗中細胞濃度的誤差為 10%,在 47 oC 和 45 oC 的同位素分析取樣時間 分別為 39.5 和 575 小時 (以虛線表示)。
36
圖3-12、T. commune 接近株在 40 oC 和 35 oC 之細胞生長曲線與硫酸鹽濃度變 化。實驗中細胞濃度的誤差為 10%,在 40 oC 和 35 oC 的同位素分析取樣時間 分別為 957 和 1098 小時 (以虛線表示)。
37
3.2 細胞生長速率與細胞硫酸鹽還原速率
藉由觀察細胞濃度與硫酸鹽濃度隨時間的變化可以求得細胞生長速率與細 胞硫酸鹽還原速率。Desulfovibrio gigas 在 30-36 oC 的細胞生長速率相近 (μ=0.20-0.22 h-1),最大值出現在 33 oC,即為 Desulfovibrio gigas 的最適生長溫 度,高於或低於 33 oC 的培養溫度,其細胞生長速率則會隨著溫度的變化而下 降。Desulfovibrio gigas 細胞硫酸鹽還原速率則隨溫度升高而增加,10 oC 和 13 oC 培養的還原速度非常緩慢 (圖 3-13a)。
Thermodesulfobacterium commune 接近株的細胞生長速率隨溫度變化的趨 勢與 Desulfovibrio gigas 相似,最大值出現在 72 oC (μ=0.15 h-1),即為
Thermodesulfobacterium commune 接近株的
最適生長溫度,當溫度大於72 oC 時,細胞生長速率則會隨著溫度上升而下 降,在 45-72 oC 之間會隨溫度上升而上升,而當培養溫度在 40 oC 和 35 oC 時,
生長十分緩慢而無法辨識。細胞硫酸鹽還原速率則在 45-72 oC 之間隨溫度上升 而上升,最大值出現在77 oC (56.1 fmol‧day/cell),但在 79 oC 硫酸鹽還原速率 下降至 29.9 fmol‧day/cell (圖 3-13b)。
圖3 Des (b) 速率
3-13、兩菌 sulfovibrio g
Thermodesu 率大致會隨
(
菌株之細胞生 gigas 之最適 ulfobacteriu 隨著溫度上升
(a)D.giga
生長速率與 適溫度為3 um commun 升而上升,
as
38
與細胞硫酸鹽 33 oC,硫酸
ne 接近株之 但在79 oC
鹽還原速率 酸還原速率會 之最適溫度為 C 時下降。
率隨溫度的變 會隨著溫度 為72 oC,細
變化。(a) 度上升而上
細胞硫酸還 上升。
還原
39
3.3 硫同位素分化
同位素的分化為二重覆平均的結果,Desulfovibrio gigas 產生的硫同位素分
化 (34εsulfate-sulfide) 範圍從 10.3‰ 到 29.7‰,隨溫度的變化可分為兩個部分,以
27 oC 作為分界點,當溫度低於 27 oC 時,除了在 13 oC 出現一個相對稍高值 (29.7‰) 外,其餘的溫度產生的硫同位素分化隨溫度上升而下降,當溫度在 27
oC 以上時,硫同位素分化則會隨溫度上升而上升 (圖 3-14a)。
Thermodesulfobacterium commune 接近株產生的硫同位素分化範圍從 8.2 ‰ 到 31.6 ‰,硫同位素分化大小隨溫度沒有明顯的變化趨勢,大部分培養溫度產生的 硫同位素分化在 8.2 和 18.9 ‰ 之間變動,僅在 68 oC 出現一特殊高值 (31.6 ‰) (圖 3-14b)。
多重硫同位素分析的分析結果顯示 Desulfovibrio gigas 和
Thermodesulfobacterium commune 接近株產生的 Δ33Ssulfate-sulfide 值範圍分別為 0.05‰ 到 0.31‰ 和 0.00‰ 到 0.29‰ 之間,隨溫度的變化趨勢與硫同位素分 化相近 (圖 3-15)
40
圖3-14、兩菌株產生之硫同位素分化 (34ε) 隨溫度的變化。(a) Desulfovibrio gigas 硫同位素分化隨溫度大致呈 V 字型的變化。(b) Thermodesulfobacterium
commune 接近株產生之硫同位素分化隨溫度無明顯規律性變化,僅在 68 oC 出 現一高值。
41
圖3-15、兩菌株產生之 Δ33S 值隨溫度的變化。兩菌株產生之 Δ33S 值隨溫度的 變化趨勢與硫同位素分化相近。
42
第四章 討論
4.1 細胞生理特性與溫度的關係
溫度是影響微生物代謝反應的重要因素之一,細胞生長速率隨溫度的變化反 應溫度對於細胞生理特性的影響,細胞生長速率最快的溫度即為最適生長溫度,
當溫度高於或低於最適生長溫度時,細胞內部營養物質的運輸與蛋白質的運作會 開始無法正常運作使得細胞生長速率下降。硫酸鹽還原速率隨溫度增高有增加的 趨勢,反應當溫度升高反應速率也會隨著增加。根據兩菌株在不同溫度下的生長 曲線,可以計算出細胞生長速率,得知 Desulfovibrio gigas 和
Thermodesulfobacterium commune 接近株的最適生長溫度分別為 33 oC 和 72
oC,前者與前人研究結果相同,後者則稍高於同屬菌株 (Rabus, 2006)。比較細 胞生長速率與細胞硫酸鹽還原速率隨溫度的變化,發現兩菌株之細胞硫酸鹽還原 速率最快的溫度會高於細胞最適生長溫度,許多前人研究結果中也可發現此現象 (Canfield et al., 2006; Hoek et al., 2006; Mitchell et al., 2009),表示當溫度高於最適 生長溫度時,大部分經由硫酸鹽還原所產生能量會用於修補細胞而不是繼續增加 細胞數目。
4.2 硫同位素分化與前人研究比較
Desulfovibrio gigas 和 Thermodesulfobacterium commune 接近株在不同溫度 培養產生之硫同位素分化的變化範圍分別為 19.3‰ 和 23.4‰,此變化範圍未超 過前人同樣以溫度反應槽進行純菌株之硫同位素分化的實驗結果 (Canfield et al., 2006; Hoek et al., 2006; Johnston et al., 2007; Mitchell et al., 2009)。兩菌株產生的最 大硫同位素分化分別為 29.7 ‰ 和 31.6 ‰,符合前人研究硫酸還原菌純菌株的 硫同位素分化大小的範圍 (Canfield, 2001a)。雖然本研究符合純菌培養之硫同位 素分化情形,但經過仔細比較也發現本研究培養的菌株與前人研究之同種菌株所 產生的分化大小與分化範圍並不相同。Desulfovibrio 在最適生長溫度產生的同位 素分化介於 2.0‰ 和 5.4‰ 之間,使用溫度梯度反應槽的多溫度培養則可產生