國立臺灣大學生命科學院植物科學研究所 博士論文
Graduate Institute of Plant Biology College of Life Science
National Taiwan University Doctoral Dissertation
綠豆發芽過程中子葉粒線體DNA之複雜性
和DNA複製型式相關性之探討
Correlation between mtDNA complexity and mtDNA replication mode in developing cotyledon
mitochondria during mung bean seed germination
研究生﹕成 寧 Advisee:Ning Cheng
指導教授﹕戴 華 博士 (Dr. Hwa Dai)
林納生 博士 (Dr. Na-Sheng Lin) 共同指導﹕鄭石通 博士 (Dr. Shih-Tong Jeng)
中華民國 105 年 9 月 September 2016
目 錄
List of Tables ... iv
List of Figures ... v
中文摘要 ... vii
Abstract ... viii
Abbreviation ... ix
I. Introduction ... 1
1. Mitochondrial genome ... 1
2. Mitochondrial nucleoid ... 2
3. Plant mitochondrial genome ... 3
4. Mitochondrial genome replication ... 5
5. The aim of the study ... 7
II. Materials and Methods ... 8
1. Materials ... 8
2. Mitochondria isolation from mung bean cotyledons ... 8
3. MtDNA purification of mung bean cotyledons ... 9
4. Flow cytometric of isolated mitochondria ... 10
5. Pulsed-field gel electrophoresis ... 10
6. Analytical ultracentrifugation analysis ... 11
7. EM examination of mtDNA ... 11
8. PCR analysis ... 12
9. In vivo mtDNA synthesis ... 12
10. LC-MS/MS ... 13
11. Mung bean nuclease and RNaseH treatment
for mtDNA ... 13
12. DNA purification from PFGE gels ... 14
13. Real-time PCR analysis ... 14
14. Immunoblot Analysis ... 14
III. Results ... 16
1. Altered quantity of mtDNA at different stages of cotyledon development ... 16
2. Physical characters of cotyledon mtDNA during seed germination ... .17
3. Fluctuation of cotyledon mtDNA synthesis in organello during seed germination ... 18
4. The difference between wb and fm mtDNA in terms of buoyant density, ultra-structure and restriction enzyme patterns ... 20
5. MtDNA ultrastructural dynamics during cotyledon development ... …20
6. Structural characteristics of giant mtDNA rosette complex ... 22
7. Unraveling the mtDNA rosette structure by freeze-thaw treatment in
vitro ... 238. RDR recombination assessed by real-time PCR ... 26
9. Mitochondrial nucleoproteins present in cotyledon mitochondria with cotyledon aging during mung bean seed germination ... 26
IV. Discussion ... 28
V. Conclusions ... 38
VI. References... 39
Tables ... 45
Figures ... 50
List of Tables
Table 1 Real-time PCR analyses for recombination activity design ... 45 Table 2 EM of mtDNA structure during cotyledon development ... 46 Table 3 The nucleoproteins present in mung bean cotyledon mitochondria
with progressive mitochondrial development during seed
germination ... 47
List of Figures
Figure 1 Quantitative alteration of mtDNA content per cotyledon
mitochondrion harvested from different cotyledon stages during seed germination ... 50 Figure 2 PFGE fractionation analysis of two mtDNA populations with
different mobilities present in different-aged cotyledons ... 51 Figure 3 The fluctuation of mtDNA with two different buoyant density
sub-populations in different-aged cotyledon mitochondria ... 53 Figure 4 Southern blot hybridization and PCR analysis of purified mtDNA samples for possible contamination with plastid DNA ... 54 Figure 5 DNA synthesis capacity of different-aged cotyledon mitochondria
... 55 Figure 6 AUC, EM and restriction enzyme digestion analysis of wb and fm sub-populations of cotyledon mtDNA purified from PFGE gel ... 56 Figure 7 The progressive alteration of cotyledon mtDNA structural
conformations at early stages of seed germination ... 57 Figure 8 The progressive alterations in structural conformation of
cotyledon mtDNA during seed germination ... 59 Figure 9 Giant rosette mtDNA complex found in cotyledon mitochondria
during stages of high DNA replicative synthesis activity ... 60 Figure 10 EM images of irregular giant rosette structures found in day-1
cotyledon mtDNA ... 61 Figure 11 Structure and conformation alterations of day-1 cotyledon
mtDNA molecules effected by in vitro freeze-thaw treatment ... 63
Figure 12 The small reticular networks around the core structure found in
early and high RDR replicating cotyledon mitochondria ... 65 Figure 13 EtBr, MBN, and RNaseH treatment in mung bean cotyledon
mtDNA ... 64 Figure 14 AUC analysis of cotyledon mtDNA after RNaseH treatment and heat denaturation ... 67 Figure 15 The recombination forms quantification of repeated sequence E
and F in mung bean cotyledon mt-genome during seed germination ... 68 Figure 16 Hypothesized model of RDR in mung bean cotyledon
mitochondria during seed germination ... 69
中文摘要
綠豆子葉粒線體 DNA 結構、型態和複雜性在綠豆種子發芽過程
中呈現高度的動態變化,本研究目的在藉此了解植物粒線體 DNA 複
雜性和複製過程的相關性。研究結果顯示,在早期子葉發育過程中,
粒線體中短的 DNA 片段轉換為長的線性 DNA 以及花瓣狀結構的
DNA。隨著 DNA 在子葉發育過程中複製活力越高,伴隨著越來越複
雜的花瓣狀結構出現。在 DNA 複製過程中,複製叉會出現在這種花
瓣結構核心周圍部分。以低溫處理種子或低溫儲存已純化之子葉粒線
體 DNA,會誘導 DNA 轉成長線狀結構,花瓣結構的核心部分消失。
這種長線結構上可明顯觀察到 D 環 (D-loop)、霍利迪交叉 (Holliday
junction)等複製結構。長線結構型態的 DNA 總長度可能超過花瓣結
構型態的長度 30 倍以上。本研究推測這種花瓣核心結構包含大量聚
集的 DNA,是依賴重組複製模式(Recombination dependent replication, RDR)中 DNA 複製的主要場所以及複製起點位置。在花瓣結構上出 現的衛星核心結構代表另一個重新開始的複製起點,進而形成巨大且
複雜、具有大量複製中間體的粒線體 DNA 分子結構。此研究結果可
證實花瓣狀 DNA 結構是植物粒線體複製之中間產物,並且確定粒線
體 DNA 是依循依賴重組複製模式進行複製。
ABSTRACT
The highly dynamic changes in mitochondrial DNA (mtDNA) conformation, structure and complexity correspond to the activity of mtDNA recombination dependent replication (RDR). This study aimed to elucidate the interrelationship between mtDNA replication and genesis of the multi-genomic, highly complex structure of plant mtDNA in mung bean cotyledons. The results suggest that short DNA fragments convert to longer linear and rosette structures early in the
development of cotyledon mitochondria. Consequently, a large number of rosette structures appear, with simultaneous elevation of mtDNA synthesis. A fork-like structure near the rosette core appears during mtDNA replication. With in vivo prolonged-cold incubation or in vitro freeze-thaw treatment, the rosette structure is converted to a much longer linear DNA structure and the rosette core disappears. This large linear DNA with displacement loops, Holliday junctions and other structures attached may increase in size more than 30 times as compared to the original rosette entity. The rosette core may consist of condensed mtDNA and play an initial and central role in RDR. The satellite cores in the rosette structure may represent the re-initiation sites of mtDNA RDR in the same parental molecule. Consequently, highly complex and giant mitochondrial DNA molecules, representing RDR
intermediates, are formed in vivo. The core-and-rosette structures represent replicating DNA and almost certainly use RDR.
Abbreviation
AUC Analytical ultracentrifugation cpDNA chloroplast DNA
DAPI 4',6-diamidino-2-phenylindole ddCTP 2',3'-Dideoxycytidine-5'-Triphosphate DNA deoxyribonucleic acid
EM electron microscope
fm fast-moving mitochondrial DNA in PFGE gel
LC-MS/MS iquid chromatography coupled with tandem mass spectrometry.
mtDNA mitochondrial DNA NEM N-ethylmaleimide OD optical density
PFGE pulsed-field gel electrophoresis RCR rolling-circle replication
RDR recombination-dependent replication RI refraction index
RITOLS RNA incorporation throughout the lagging strand wb well-bound mitochondrial DNA in PFGE gel
I. Introduction
1. Mitochondrial genome
Mitochondria evolved from a prokaryotic ancestor, so researchers believe that the mitochondria from most organisms including plants contain a circular genome.
Especially, a metazoan study found a near-genome-sized circle genome on electron microscopy (EM). Restriction-mapping data revealed mitochondrial DNA (mtDNA) also associated with a circular genome (Lonsdale, 1984; Palmer & Herbon, 1986;
Marechal & Brisson, 2010).
However, most plant mtDNA is found in linear and branched forms on EM
(Backert et al., 1996; Bendich, 1996; Oldenburg & Bendich, 1996; Backert & Borner, 2000). In 1991, pulsed-field gel electrophoresis (PFGE) experiments of
Saccharomyces cerevisiae showed that mtDNA appeared in linear polydisperse
tandem arrays in a genome and ranged from about 75 to 150 kb. This result rejected earlier hypotheses that all mtDNA genomes were circular (Maleszka et al., 1991;
Williamson, 2002). Oldenburg and Bendich (2001) reported mtDNA of liverwort appearing as linear, head-to-tail concatemers and branched forms on PFGE and direct gel image analyses (Oldenburg & Bendich, 2001). Candida albicans mtDNA 2-D gel study also found that mtDNA could form a complex branched network by
recombination replication (Gerhold et al., 2010). Now most authors agree that the plant organelle genome is a mixture of monomers and head-to-tail concatemers of circular and linear molecules and easily forms a highly complex branched structure (Marechal & Brisson, 2010).
Such complex structures in the mitochondrial genome imply that mitochondria may include more than one population. Both plant and animal mitochondria show two main sub-population genomes on PFGE assay. One population stays in a well and the
other migrates to the site that equal or lower than the genome size. Some tests show that well-bound mtDNA was unlikely caused by incompletely removed protein or excessive molecular weight but may due to the complex DNA structure effect from mtDNA replication (Dudareva et al., 1988; Oldenburg & Bendich, 1996; Dai et al., 2005; Gerhold et al., 2010).
2.
Mitochondrial nucleoid
MtDNA is packaged into bacterial chromosome-like structures called
“mitochondrial nucleoids” (mt-nucleoids); mammalian cells contain thousands of copies of mitochondrial genome in hundreds of nucleoids (Chen & Butow, 2005). In human mtDNA, individual mtDNA molecules within a nucleoid may be involved in replication or transcription or may not be active in nucleic acid synthesis at any time (Reyes et al., 2011). Plants and yeast contain more than one nucleoid per
mitochondrion (Chen & Butow, 2005; Dai et al., 2005). The nucleoids in organelles, both in mitochondria and plastids, contain DNA, RNA, and proteins. The functions of DNA maintenance are associated with membrane-attached DNA and include
replication, recombination and repair. Nucleoid protein composition changes during development may result in DNA released from membrane and damage but not
repaired. Then these unbound DNA are degraded by nucleases (Oldenburg & Bendich, 2015).
This negative evidence supports a circular mtDNA genome. A circular DNA genome can replicate indefinitely and leave the membrane and is not digested by exonucleases. However, the circular genome forms of mtDNA show a low proportion of total DNA on EM or from analysis of DNA-moving data (Oldenburg & Bendich, 1996; Oldenburg & Bendich, 2015). Previous studies of mammalian cells suggested
that mtDNA replication and transcription occurred in the central core of the
mt-nucleoid and function with core region protein such as TFAM, mtSSB, DNA pol , mtRNA polymerase, and Twinkle. The peripheral region surrounds the core region and contains abundant metabolic proteins, such as Hsp60, prohibitin, and ATAD3.
These proteins may function in mtDNA maintenance and repair (Bogenhagen et al., 2008; Bogenhagen, 2012).
The model of the yeast mt-nucleoid suggests a segregation apparatus of core components consisting of the membrane proteins Mdm10-Mdm12-Mmm1 in a complex. This complex locates in the outer membrane primarily, and Mmm1 extends into the inner membrane and protrudes into the matrix. Mdm31 and Mdm32 are inner-membrane proteins and were proposed to interact with Mdm10, Mdm12, and Mmm1. This complex mediates mitochondria attaching to actin cables in the cytosol and anchoring mt-nucleoids in the matrix side (Chen & Butow, 2005). For yeast cells in the respiration-repressed condition, mtDNA is packaged by Abf2, and mtDNA transactions are inhibited. When cells shift to the respiration condition, ACO1 and ILV5 levels are increased and substitute for the packaged Abf2 (Chen & Butow, 2005).
Unlike mt-nucleoids in mammalian and yeast cells, with preliminary models, in plants, the mt-nucleoid content and structure remain unclear.
3. Plant mitochondrial genome
Plant mitochondria have an exceedingly complex and peculiar genetic system.
Investigating and understanding the mechanisms of plant mtDNA replication is difficult, mainly because of the large size and multipartite complexity. One
characteristic of plant mitochondrial genomes different from animal genomes is that higher plant mitochondria have a broad range of genome size, from 208 kb to 11.3 mb,
much larger and more complex than mitochondrial genomes for animals (14–42 kb) or fungus (19–235 kb) (Backert et al., 1997b; Cupp & Nielsen, 2014; Losada et al., 2014). However, plant mitochondrial genomes do not encode more proteins and RNAs than animal genomes even with the larger size. Most of these large genomes are caused by repeated sequences, large introns, AT-rich non-coding regions, and other non-coding DNA sequences (Cupp & Nielsen, 2014). Recent studies suggest that some plant mitochondria have a multichromosomal structure. As an example, the cucumber (Cucumis sativus) mitochondrial genome has one large (1556 kb) and two small (84 and 45 kb) circular-mapping chromosomes (Alverson et al., 2011a).
Actually, plant mitochondrial genomes show gene content variation during
evolution, which causes gene loss and functional gene transfer to the nuclear genomes.
Ribosome protein genes and succinate dehydrogenase genes are the most likely to be lost. The cox2 in Vigna radiata is one of few exceptional genes that functions in the electron transport chain and is absent in mitochondria; it transfers from the nuclear genome (Alverson et al., 2011b; Petersen et al., 2015).
Plant mitochondrial genomes often contain large repeat sequences associated with reversible homologous recombination . A number of studies have shown that plant organelle genomes can rapidly change their structures and undergo DNA rearrangement. The striking changes in mtDNA structural characteristics indicate the highly dynamic nature of organellar DNA structural organization during different cell growth cycles (Oldenburg & Bendich, 1996; Backert & Borner, 2000; Dai et al., 2005;
Kumar et al., 2014; Kumar et al., 2015; Oldenburg & Bendich, 2015). In C. albicans, a noncoding region in an inverted repeat is involved in replication initiation via homologous recombination, and large inverted repeats identified in these genomes play a main role in altering molecular architectures (Gerhold et al., 2010; Valach et al.,
2011). This phenomenon leads to the generation of small sub-genomic circles and increasing genomic complexity (Chang et al., 2013; Gualberto et al., 2014). In
soybeans, it is suggested that large amount of repeat sequences mediate the formation of various circular structures. The observed linear or branched DNA structures by EM are the result of DNA degradation, and consequently disrupt the net-like structure of mtDNA. (Takanashi et al., 2006; Chang et al., 2013).
4. Mitochondrial genome replication
Some modes of mtDNA replication to copy the mitochondrial genome have been proposed. The mtDNA genome is considered to be in a circle form in mammalian cells, so the strand-coupled model proposes that replication is initiated from a region near the OriH region bidirectionally by a progression of the two forks around the mitochondrial genome circle. The other mode is asynchronous strand-displacement, with mtDNA replicating asymmetrically and mtDNA synthesis primed at the OriH origin within the non-coding region called the displacement loop (D-loop). The L-stand replication initiates in the opposite direction when OriL is exposed in the circular mitochondrial genome. (Bowmaker et al., 2003; Kazak et al., 2012). In recent study, mammalian mtDNA was proposed to replicate by the model of RNA
incorporated throughout lagging strand (RITOLS). The RITOLS model is like the strand-displacement model, and the lagging strand is initially transcribed as RNA before being converted to DNA (Pohjoismaki et al., 2010; Kasiviswanathan et al., 2012; Reyes et al., 2013). The “bootlace model” is derived from RITOLS; pre-formed L-strand RNA is incorporated at the mtDNA replication fork via 3’ to 5’ hybridization with the displaced H-strand and is replaced by lagging-strand DNA in a subsequent process (Reyes et al., 2013)
Research of the replication of plant mtDNA is mainly impeded by the highly complex structure. Only a few details are known about the mode or origin of DNA replication and recombination in mitochondria. High abundant linear or branch DNA molecules were observed on EM in different studies but were considered degradation products from larger circular molecules. The ϭ-like structures DNA were revealed, which suggested that rolling-circle replication (RCR) is involved in genome maintenance. The D-loops found on organelle DNA also suggested a dual
origin–dependent RCR model. However, RCR models depend on the assumption that mtDNA exists in genome-sized circles in plant mitochondria (Marechal & Brisson, 2010). The other possible model of plant mtDNA replication is recombination- dependent replication (RDR): replication starts at the D-loop formation, and a free 3’-OH single-stranded DNA overhang can intrude into another copy of the genome at a homologous site. New DNA is synthesized on the leading strand, which establishes the replication fork (Oldenburg & Bendich, 1996; Cupp & Nielsen, 2014).
Increasing data suggest that replication is a phage T4-like replication mechanism for mtDNA in plant cells (Oldenburg & Bendich, 2001; Bendich, 2010; Gerhold et al., 2010; Gualberto et al., 2014). This model indicates that most mtDNA is in branched and linear form, and these structures are variable and due to monomeric and
head-to-tail concatemeric arrays (Oldenburg & Bendich, 2001). Most studies suggest that plant mitochondria use different mechanisms for DNA replication and have several replication initiation sites (Shedge et al., 2007; Gerhold et al., 2010; Cupp &
Nielsen, 2014). However, until now, the current models of RDR and/or RCR do not satisfactorily explain the highly complex mitochondrial genome observed on PFGE, in moving images and on EM. Currently, we cannot rule out that mtDNA replication may involve different processes and factors depending on the plant, plant tissue and
developmental stage.
5. The aim of the study
In mung bean cotyledons grown from 4 °C-12 hr to day 1, mt-nucleoids showed synchronized development in terms of mtDNA structural organization during seed germination (Dai et al., 2005). Here, I aimed to elucidate the progressive alteration of mtDNA in structural conformation, DNA organization and complexity in cotyledon development during mung bean seed germination. I also attempted to determine the association between mtDNA structural complexity and the mtDNA replication process.
From serial mtDNA dynamic analysis at different cell age points under EM
examination, I provide further evidence that RDR is involved in mtDNA replication, and the unique RDR mode in plant mitochondria creates the high complex and dynamic organization of mtDNA in mung bean cotyledons.
II. Materials and Methods
1. Materials
In this study, I studied mtDNA organization in cotyledon mitochondria from mung bean (Vigna radiate [Wilzed] cv. Tainan No. 5). The genus Vigna, containing mung bean, is an important protein source in Africa and Asia. For plant mitochondrial study, mung beans have a rapid growth rate and are etiolated seedling are easy to obtain to avoid plastid contamination.
The mitochondrial genome of mung bean has been assembled and predicted as a single, circular-mapping molecule that is 401,262 nt long with 45.1% GC content, 31 proteins and 3 rRNA and 16 tRNA genes (Alverson et al., 2011b). Different from other plants, the mung bean mitochondrial genome is one of the few that lacks the large (>1 kb) recombining repeats. Most repeats in the mung bean mitochondrial genome are < 100 nt (Alverson et al., 2011b; Naito et al., 2013).
2. Mitochondria isolation from mung bean cotyledons
Mitochondria were harvested from different-aged mung bean cotyledons, including dry seeds; seeds in 4 °C water for 2, 6 and 12 hr; and seeds immersed in water at 27 °C for 12 hr. Seedlings were examined at days 1 (12-hr water immersion at 27 °C, followed by 12-hr growth in vermiculite at 27 °C), 2 and 3. Some
experiments involved two additional stages, 4 °C-12 hr immersed and an additional 2 or 6 hr 27 °C immersion. All cotyledons harvested from seeds and seedlings were kept in the dark. All embryos were removed. The M-system buffer preparation for isolation and removal of plastids/nuclei was as described (Dai et al., 1991): M-1 buffer [pH 7.2, 0.4 M D-mannitol, 25 mM MOPS (3-Morpholinopropane-1-sulfonic acid)
last three added fresh just before use]; M-2 buffer (pH 7.2, 0.4 M mannitol, 5 mM MOPS, 1 mM EGTA, 0.1% (w/v) BSA, 1.8 M sucrose /M-3 buffer (pH 7.2), 1.8 M sucrose, 10 mM Tricine, 1 mM EGTA, 0.1% (w/v) BSA]; M-4 buffer (pH 7.2, 0.2 M D-mannitol, 10 mM Tricine, 1 mM EGTA); and M-5 buffer (pH 7.2, M D-mannitol, mM Tricine, 2 mM EGTA).
Cotyledons were homogenized in M-1 buffer (1 mL/g tissue) in a Waring Blendor, than filtered through four layers of cheesecloth and two layers of Miracloth.
The filtrate was centrifuged to reach 4000 g, then braked under low speed twice. The crude mitochondria in supernatant was pelleted at 14,000 g for 15 min. Crude
mitochondria was resuspended in M-2 buffer and fractionated on a continuous sucrose gradient (0.6-1.8 M in M-3 buffer) at 81,000 g for 1 hr in a swinging bucket rotor.
Purified mitochondria was banded between 1.30 to 1.35 M in the sucrose gradient.
The population was recovered and slowly diluted by adding M-4 buffer to a give a final concentration of 0.6 M. The mitochondria was recovered by centrifuging at 20,000 g for 15 min, then suspended in M-5 buffer. The mitochondria purified from the M-system was used for flow cytomety, pulsed-field gel electrophoresis, and mtDNA synthesis.
3. MtDNA purification of mung bean cotyledons
The B-system isolation buffers for mtDNA were used as described with
modification (Sambrook & Russell, 2001; Dai et al., 2005): B-l buffer [pH 7.2, 0.5 M D-mannitol, 10 mM TES (pH 7.2), 1 mM EGTA, 1% (w/v) PVP-40, 0.1% (w/v) BSA, 0.1 M diethyldithiocarbamate, 10 mM mercaptoethanol, 20 mM sodium ascorbate (the last three added fresh just before use]; B2A buffer [pH 7.2), 0.5 M D-mannitol, 10 mM TES (pH 7.2), 0.1% (w/v) BSA]; B2B buffer [pH 7.2, 0.5 M D-mannitol, 10
mM KH2PO4, 50 mM EDTA, 150 mM NaCl, 0.1% (w/v) BSA]; 1.8 M sucrose/B-3 buffer [pH 7.2, 1.8 M sucrose, 10 mM TES (pH 7.2), 20 mM EDTA, 0.1% (w/v) BSA].
Cotyledons were homogenized and nuclei and plastids were removed in B-1 buffer in the same step as the M-system isolation method. After nuclei and plastids were removed, crude mitochondria were incubated with 27 μg/ml DNase I and 10 mM MgC12 for 30 min at room temperature in B-2A buffer. The DNase-treated mitochondria were washed by adding extraction B2B buffer containing 50 mM EDTA and pelleted at 14000 g for 20 min, then fractionated on a continuous sucrose gradient at 81,000 g for 1 hr. Pure mitochondria was lysed with 1% sarcosyl and incubated with 0.2 mg/ml proteinase K in 50 °C for 1 hr. It was then applied to a 5-μg/ml DAPI (4',6-diamidino-2- phenylindole)-CsCl gradient and centrifuged at 117,000 g 17 hr, then decelerated to 89,000 g 24 hr. The MtDNA-DAPI population in CsCl gradient could be detected under UV light. The mtDNA were recovered by Centricon-100.
4. Flow cytometry of isolated mitochondria
Mitochondria from different stages of cotyledons were stained with 5 μg/ml DAPI for 3 min, then analyzed by use of an Elite ESP flow cytometer (Beckman and Coulter) with a 70-mW 365-nm UV laser for excitation. Fluorescence was measured at 457/50 nm. For each experiment, 50,000 mitochondria were analyzed.
5. Pulsed-field gel electrophoresis
Cotyledon mitochondria at different stages were resuspended in TE buffer (10 mM tris-HCl, 1 mM EDTA, pH 8.0), mixed with an equal volume of 1.5% low
melting-point agarose. The plugs were treated with ESP buffer (1 mg/ml proteinase K,
1% sarkosyl, 0.5M EDTA, pH 9.0) at 50 °C overnight. PFGE was performed at 30–60’ pulse time, 150 V (5 V/cm) on 1% agarose gel for 16 hr in 0.5x TBE buffer (45 mM Tris-borate, 1 mM EDTA) at 13 °C. Gels were then stained with ethidium bromide, de-stained by washing and underwent conventional Southern blotting analysis (Sambrook et al., 1982).
6. Analytical ultracentrifugation analysis
Different-stage cotyledon mitochondria were lysed by 1% sarcosyl and treated with 0.2 mg/ml proteinase K at 50 °C for 1 hr. The mean CsCl density was adjusted to 1.7284 g/cm3 (RI=1.4020). Mitochondria were centrifuged to equilibrium at 44,000 rpm at 20 °C for 24 hr by using the Beckman XL-A analytical ultracentrifuge (Beckman-Coulter, Brea, CA, USA), and absorbance was measured at 260 nm.
Micrococcus lysodeikticus DNA (buoyant density at 1.731 g/cm3) was used as a marker. The buoyant density is calculated from the fragment’s mean distance r from the axis of the ultracentrifuge at sedimentation equilibrium (Ifft et al., 1961):
m r2- r2m)/(2B)
wherem and rm are the buoyant density and radial position of a marker DNA, respectively, is the angular speed, and B is 1.190 x·109 (cgs units) for Beckman models E and XL-A under standard conditions.
7. EM examination of mtDNA
MtDNA were purified from DAPI-CsCl-gradient. The Kleinschmidt method was used for surface spreading DNA (Thresher & Griffith, 1992). A 50-l aliquot of DNA
in water was mixed with ammonium acetate to a final concentration of 0.25 M.
Cytochrome c was added to 4 μg/ml and then dropped on parafilm for 20 to 40 min. A collodion film covering the EM grid was touched to the drop and washed with 75%
and 90% ethanol, then air-dried and rotary shadowed with platinum at a 15° angle in a JEOL JEE-420T high-vacuum evaporator (JEOL, Peabody, MA, USA). Before
observation, the grid was coated with carbon to support the film. Samples were examined under a Philips CM100 electron microscope (Philips, Eindhoven, Netherlands) at 80 kV.
8. PCR analysis
Three sets of primers were used for organelle DNA amplification:
1. Mitochondria-encoded gene cox 3: cx3-34U, gtagatccaagtccatggcct and cx3-458L, gcatgatgggcccaagttacggc
2. Nucleus-encoded gene cox 2: cx2-527U, tcccacaaaggattgttcatgg and cx2-994L, cctaactcttaccacgttatat
3. Chloroplast-encoded gene rbcL: rbcL-1049A, tggactgatggacttaccagtcttgatcg and rbcL-4S, acttcgcaagcagcagctaa ttca ggact
The template DNA was amplified by PCR with an initial 95 °C-1 min heating period; 20 or 30 cycles of 94 °C for 30 sec, 55 °C for 30 sec. and 72 °C for 1 min; and finally 72 °C for 5 min to terminate the reaction.
9. In vivo mtDNA synthesis
As described, cotyledon mitochondria of various ages were dissolved in IO buffer (50 mM Tris, 20 mM MgCl2, 40 mM KCl, 2 mM DTT, 1 mM ATP, 50 µM dATP, 50 µM dGTP, 50 µM dTTP). Then, 50 µCi of [α-32P]dCTP (3000 Ci/mmol)
was added to each reaction, and the reaction was mixed well and incubated at 30 °C with shaking for 1 hr. The reaction was stopped by adding 100 μM dCTP and 25 mM EDTA. The mixture was then centrifuged at 14,000 g for 10 min, and the pellet was dissolved in 100 μl TE buffer. Next, 0.5% Triton-X100 was added, and the mixture was incubated at 25 °C for 5 min, then centrifuged at 20,000 g for 1 hr to remove bacterial contamination. The supernatant was then applied to PFGE as described.
Additionally, the gel was dried before development of X-ray films (Dai et al., 2005).
10. LC-MS/MS
Cotyledon mitochondria of various stages were treated with 1% NP-40 and 1%
sarcosyl, then underwent PFGE fractionation. The well-bound (wb) and fast-moving (fm) mtDNA–protein complex (wb- and fm-nucleoid, respectively) was spliced from PFGE gel and underwent trypsin digestion, as we previously described (Lo et al., 2011). The pellet of tryptic peptides was dissolved in 10 to 20 μl of 0.1% (w/v) formic acid for LC-MS/MS. The data were analyzed by using an in-house search with
Mascot v2.2.06 (Matrix Science) against the UniProtKB/Swiss-Prot Viridiplantae database (green plants, 28,773 protein entries) as described (Lo et al., 2011).
11. Mung bean nuclease and RNaseH treatment for mtDNA
Day-1 mtDNA purified by CsCl gradient was used for mung bean nuclease treatment. An amount of 0.2-μg mtDNA for each treatment was incubated with 0 to 10 units mung bean nuclease (M2025S, New England BioLabs) at 30 °C for 30 min. In RNaseH treatment, mtDNA was incubated with 0 to 3 units RNaseH (M4281, Promega Corp.) at 37 °C for 1 hr. MtDNA was separated on 0.5% agarose gels at 1 V/cm and 21 °C for 20 hr in 0.5x TBE buffer after nuclease treatment and underwentconventional Southern blotting analysis.
12. DNA purification from PFGE gels
The method of mtDNA purification from PFGE gel for analytical
ultracentrifugation analysis was previously described (Li et al., 2010). A 100-mg gel slice was incubated in tubes with 300 μl 6 M NaI, and the tube was heated at 70 °C 3 min to dissolve the gel. Then 10-μl silica matrix from silicon dioxide-removed fine silica particle was added for incubating for 2 min at room temperature. The matrix was centrifuged at 16,000 g for 10 sec, washed with washing buffer (50% ethanol, 10 mM Tris-HCl, pH 7.5, 100 mM NaCl, 1 mM EDTA), then resuspended in 30 μl sterile water and heated at 70 °C for 2 min and centrifugred at 16,000 g for 2 min and the supernatant was collected as DNA eluate.
13. Real-time PCR analysis
Real-time PCR involved use of FastStart Universal SYBR-Green Master Mix (Rox) (Roche) in a AB QuantStudio 12K Real Time PCR System (Life Technologies) programmed to hold at 50 °C for 2 min, then run at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 sec, then 60 °C for 1 min. Specific primers for Vigna radiata mtDNA were designed and are in Table 1. Cox3 level was an internal control.
Fluorescence signal of SYBR-Green dye and the Ct value for each reaction were determined by using QuantStudio 12K Flex (Life Technologies), setting the threshold fluorescence into the exponential phase of the amplifications.
14. Immunoblot Analysis
Proteins from mitochondria at different cotyledon stages were separated by SDS-PAGE, then transferred to polyvinylidene fluoride membranes (Millipore), which were incubated with antiSSB and antiATPase 1 hr at room temperature to identify MtSSB and ATPase (GTMA). Immunoprotein complexes were detected by using the Amersham ECL Plus-Western Blotting Detection System (GE Healthcare).
III. Results
1. Altered quantity of mtDNA at different stages of cotyledon development
To check the mtDNA quantity, mitochondria were isolated from cotyledons at different stages and stained with DAPI for fluorescence microscopy observation (Zeiss Axio Imager Z1, detected by CoolSnap HQ) (Fig. 1A) and flow cytometry (Fig.
1B) to indicate the amount of DNA from isolated cotyledon mitochondria. MtDNA content increased in cotyledons, from 4 °C-12 hr samples, and peaked in 27 °C-12 hr samples. The high content was sustained in day-1 and -2 mitochondria after
germination and steeply decreased in day -3 mitochondria.
The mtDNA content in cotyledon mitochondria was analyzed by PFGE. The same amount of mitochondria in terms of protein content (0.5 mg for serial aged mitochondria) was used. Mitochondria from different stages of cotyledons were lysed and treated with protease K. MtDNA separated into two sub-populations by PFGE as previously reported (Bendich, 1996; Dai et al., 2005): one stayed in the sample loading well (well-bound, wb) and the other one migrated as a smeared zone to the area equivalent to 50 to 200 kb of linear DNA (the fast-moving component, fm). The mobility of fm mtDNA was the same among different stages of cotyledons (Fig. 2A).
The mtDNA content was detected by Southern blot analysis (right panel of Figs 2A and B). As for flow cytometry, with Southern blot analysis, 4 °C-12 hr immersed cotyledons showed less mtDNA content, especially in the wb fraction. The wb mtDNA appeared in 27 °C-12 hr mitochondria and increased in day-1 mtDNA, followed by some decrease in day-2 mtDNA. A small amount of the wb form was found in day-3 cotyledons. In 4 °C-12 hr immersed cotyledon, a large amount of fm
fluctuations of wb and fm parts indicate these two sub-populations may have a different structure and therefore different functions.
Without protease treatment in the same PFGE assay, the mtDNA–protein complex (mt-nucleoids) also separated into two sub-populations. As for protease treatment assay in 4 °C-12 hr immerse cotyledons, the wb form was not detected in dry seeds, or in 4 °C-2, -6 or -12 hr immersed cotyledons by Southern blot analysis (Fig. 2B-b, lanes 1-4). The wb component emerged unambiguously in the 27 °C-12 hr mitochondria and its relative amount increased in day-1 and -2 mtDNA (Fig. 2B-b, lanes 5-7). In addition, the fm component mobility fluctuated during cotyledon development. The retarded fm–protein complex was observed during migration as a smear zone in the area equivalent to 50 to 300 kb linear double-stranded DNA (Fig.
2B). The migration retardation of the fm mtDNA–protein complex became obvious in 27 °C-12 hr mitochondria increased in day-1 mitochondria and remained
approximately the same up to day-2 mitochondria. As a control, proteinase K treatment was applied to an extra day-1 mitochondrial sample (Fig. 2B, lanes 8); the electrophoresis mobility of the fm component was increased in the resulting mtDNA sample (Fig. 2B, lanes 6-8).
2. Physical characteristics of cotyledon mtDNA during seed germination
PFGE analysis revealed two sub-populations in mung bean mtDNA that were presumably caused by differences in DNA conformation and organization (Gerhold et al., 2010). To understand the altered mtDNA conformation/organization, different aged cotyledon mitochondria were lysed and underwent analytical ultracentrifugation (AUC) analysis in a CsCl gradient. The DNA buoyant density estimated by the
marker DNA for Micrococcus lysodeikticus is 1.731 g/cm3. Two sub-populations of mtDNA, peak a, ρ= 1.696 and peak b, ρ= 1.709 g/cm3, were observed by measuring the optical density (OD) at 260 nm after 24-hr centrifugation. Peak b was consistently detected in all mtDNA samples throughout seed germination. Peak a was considered the co-purified chloroplast DNA (cpDNA) as previously described (Wells & Ingle, 1970; Kolodner & Tewari, 1972). It was not detected in 4 ºC- 2 hr and day-3 mitochondria but was seen in other samples (Fig. 3). Southern blot hybridization analysis showed slight chloroplast DNA contamination in 27 °C-12 hr and day-1 mitochondria, but most of the DNA population in 27 °C-12 hr to day 1 mitochondria was mtDNA (Fig. 4A). The mitochondria from day 3 seedlings, as a control, showed peaks at a and b in the AUC analysis, but no chloroplast DNA was detected by Southern blot analysis. The PCR analysis shown in Fig. 4B also shows only a small amount of plastid DNA present in isolated cotyledon mtDNA. These results indicate that peak a may be the mtDNA and cpDNA mixed population. The ratio of peak a to peak b quantity decreased from day-1 to -3 mitochondria, so peak a and b may be two sub-populations of mtDNA with different physical conformation/organization and physiological function.
3. Fluctuation of cotyledon mtDNA synthesis in organello during seed germination
The resuts from PFGE and AUC assay indicated the mtDNA in different growth stages may have different organization for different physiologic functions. To
examine the correlation between mtDNA content/mtDNA complexity and mtDNA synthesis ability during cotyledon development, 32P-dCTP incorporation was used as described (Dai et al., 2005). At the early seed immersion stage, the newly synthesized
mtDNA was clearly in the fm form in isolated 4 °C-12 hr mitochondria after PFGE fractionation of newly synthesized mtDNA (Fig. 5A, lane 1), whereas the relative amount of the newly synthesized wb form increased gradually after an additional 2 and 6 hr incubation at 27 °C (Fig. 5A, lane 2 and lane 3). 27 °C-12 hr to day-2 cotyledon mtDNA showed a much higher DNA synthesis in both fm and wb forms (Figs 5A, lane 4 and 5B, lane 1-3). Day-3 mitochondria showed a sharp decline in mtDNA synthesis activity (Fig. 5B, lane 4). This result suggests that cotyledon mtDNA with 27 °C-12 hr immersed seed and day 1 after germination has high DNA synthesis ability.
MtDNA synthesis was inhibited with three DNA polymerase inhibitors, aphidicolin, N-ethylmaleimide (NEM), and ddCTP, to assess the nature of the polymerase responsible for the observed in organello mtDNA synthesis (Fig. 5C).
After 1-hr pulse-labeling, in organello cotyledon mtDNA synthesis was strongly inhibited by NEM and ddCTP (Fig. 5C, lane 4-8), with no effect of aphidicolin treatment, even at high concentration (100 μM; Fig. 5C lane 2, 3). Thus, the in organello DNA synthesis activity of mung bean mitochondria may be driven by an
organello localized γ-like DNA polymerase rather than a nucleus-localized α-like polymerase (Sala et al., 1980; Heinhorst & Cannon, 1993; Dai et al., 2005; Zhang et al., 2011). Furthermore, during cotyledon development, the fluctuation detected for
the in organello mtDNA synthesis was substantial, especially for the wb form, which suggests that the observed mtDNA synthetic activity reflected a replicative mtDNA synthesis not just a repair/recombination event, which may also be involved. (The DNA synthesis data were generated by Dr. Annamalai and me).
Taken together, the results indicate that cotyledon mtDNA content per
mitochondrion, the structural complexity of mtDNA, and its replication activity all
fluctuated in parallel with each other during cotyledon development. This line of evidence further suggests that the increase in structural complexity of mtDNA is linked to its replication activity as reflected by the increase in mtDNA content per mitochondrion.
4. The difference between wb and fm mtDNA in terms of buoyant density, ultrastructure and restriction enzyme patterns
To clarify the correlation of the populations on PFGE gel and AUC assay data, wb and fm 27 °C-12 hr cotyledon mtDNA was eluted from PFGE gel for AUC assay.
Both wb and fm DNA still showed low density peaks (~1.6981 and 1.7006 g/cm3, respectively) and high density peaks (~1.7094 and 1.71124 g/cm3, respectively). Both low and high density peak profiles showed as very broad peaks (Fig. 6B), which may have been caused by high fragmentation during re-purification of wb/fm DNA from PFGE gel. EM revealed a more complex DNA structure in wb than fm mtDNA (Fig.
6C). The restriction enzyme digestion pattern indicated the similarity between wb and fm mtDNA, except in some minor bands (Fig. 6D, arrows; the data were generated by Yih-Shan Lo ).
5. MtDNA ultrastructural dynamics during cotyledon development
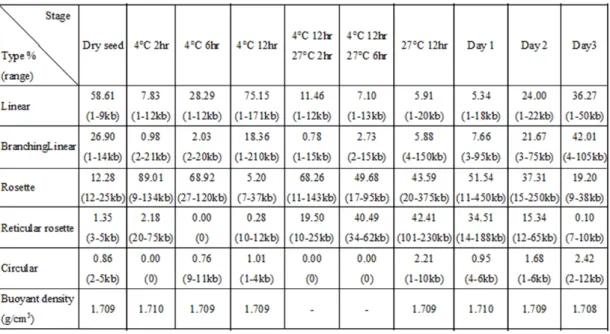
To further elucidate to the issue of mtDNA structural change during different cotyledon stages, mtDNA from different-aged cotyledons purified by CsCl-DAPI gradient ultracentrifugation were examined on EM. Before seed germination, mtDNA was observed as a short linear fragment or branch structure, most just 3-5 kb long in dry seeds (Fig. 7A and Table 2). After 4 °C-2 hr immersion, the linear fragments were slightly longer, and a high proportion was converted to a simple rosette structure incotyledons (89% of total length, Fig. 7B, Table 2). Most of these rosettes were 20 to 40 kb. These rosette structures maintained a similar but looser structural conformation in up to 4 °C-6 hr immersed samples; however, the rosettes plus reticular rosettes (rosettes comprise small reticular network) decreased from 91% to 69% of the total mtDNA length (Fig. 7C and Table 2). After 4 °C-12 hr imbibition, nearly all of the mtDNA was converted to a longer linear form and the rosette core disappeared simultaneously (Fig. 7D). The 4 °C-12 hr long linear mtDNA molecules were much longer than the individual rosettes for 4 °C-2 hr and 4 °C-6 hr mtDNA; only 5% of the total length of mtDNA maintained a rosette-like structure. The large linear mtDNA converted back into rosette structures when seeds were returned to 27 °C for 2 hr and 6 hr, after a 4 °C-12 hr imbibition; nearly 87% and 90% of the total length consisted of simple or regular rosette molecules, respectively (Figs. 7E, F and Table 2).
A much more complex rosette structure appeared with 27 °C-12 hr culture for day-1 and -2 mtDNA (Figs. 8-10). These complex rosette structures were often attached to linear, branched and circular mtDNA structures. Obvious core(s) were visible at the centers of rosettes or sub-rosettes. Nearly 86% of the total mtDNA length in 27 °C-12 hr and day-1 mitochondria were complex rosette plus reticular rosette structures. More reticular networks were found around the core structure, and most appeared in the early mtDNA synthesis stages (Figs. 8, 9, long dotted arrow).
The remaining parts were simple long linear or branchlinear forms. An independent circular structure was nearly absent in our observations. In day-2 cotyledon mtDNA, more than 50% of mtDNA remained as complex rosette structures (Fig. 8C). The giant rosettes were found in highly mtDNA-replicating mitochondria (Figs. 9 and 10).
In day-3 mtDNA, the compact rosette molecules mostly disappeared. Instead, linear DNA with/without attachments to looser networks and branch structures appeared;
only 19% of mtDNA maintained a very loose rosette structure in day-3 cotyledon mitochondria (Fig. 8D). These results indicate that the extent of mtDNA structural complexity is closely associated with the magnitude of synthesis activity during cotyledon development.
6. Structural characteristics of giant mtDNA rosette complex
On EM, a rosette-like structure of mtDNA was obvious in different stages. These individual rosette genome sizes were about 20 to 200 kb, much smaller than the 401 kb reported by other investigators (Alverson et al., 2011b). A giant rosette-like
complex was observed occasionally, especially at the height of the replicative mtDNA synthesis period during cotyledon development. EM revealed giant rosette mtDNA present in 27 °C-12 hr (Fig. 9A) and day-1 (Fig. 9B) mitochondria. Various-sized petal-like structures extended from the central core (arrows in Fig. 9). Around the core region, concentric-like petals were affiliated with putative “RDR forks”
(Oldenburg & Bendich, 1996; Backert & Borner, 2000; Oldenburg & Bendich, 2001;
Gerhold et al., 2010). The RDR forks are represented by the red sketches drawn in the upper left box of panels A and B in Figure 9, respectively. The satellite cores are indicated with arrowheads in panels A and B. Both the central core and satellite core presented as a single or multiple form. Petal-like structures and/or linear DNA structures extended outward from the central or satellite cores. The small reticular networks, marked by long dotted arrows, were often found in giant rosettes. A Holliday junction is indicated by a short dotted arrow in the upper left box of the figure. Generally, the size of the giant rosette structural molecule was larger than the single mtDNA genome size and could be as large as 2-6 genome units per molecule.
The DNA connection between rosettes could be visualized on EM (Fig. 10),
showing mtDNA rosette structures with similar (panel B-a) or varied (panel B-b) complexities in structure and organization linked to each other by DNA junctions.
The DNA junction connecting the rosette structure clearly shows that the connection passes from core to core between the rosettes (Fig. 10A, box b, and 10B). A putative RDR fork (red sketch), Holliday junction (short dotted arrow) and D-loop (short arrow) are also visible in Figure 10.
A complicated giant rosette molecule in the form of relaxed multi-rosette units was found in day-1 mtDNA (Fig. 10A). This irregular giant rosette structure differs from that in Figure 9 and rarely appeared in the investigation. This extremely large and complicated mtDNA molecule may be relaxed or converted from the RDR intermediate shown in Figure 9 during mtDNA purification. The enlarged boxed area A of Figure 10A in the upper right corner exhibits multi-cores, replication forks, a Holliday junction (short dotted arrow), and a D-loop structure (short arrow). All features point to the RDR, for example, branching linear mtDNA (double arrowhead), and a looping circular structure (arrowhead). Some reticular networks were also found near the core (long dotted arrow). The box b of Figure 10A indicates the linkage of three sub-rosettes, from core to core by DNA junctions, similar to Figure 10B. The large complex mtDNA molecule in Figure 10A is > 1000 kb.
7. Unraveling the mtDNA rosette structure by freeze-thaw treatment
in vitroThe expansion and relaxation of the day-1 cotyledon DNA molecule was induced by a long freeze-thaw treatment in water. Day-1 rosette mtDNA (40~200 kb in general) is shown in Figure 11A-a. As compared with the regular rosette-like structure, the rosette components converted into a huge, complicated and loose
mtDNA molecule (panel A-b) after the freeze-thaw treatment in water. The b1, b2 and b3 images at the bottom of Panel A-b represent enlarged images of the corresponding boxed areas displayed in panel A-b (the red sketch presents the mtDNA EM image of Fig. 11A-b). Decomposed mtDNA rosette-like structures appear as a large linear form and are attached with various decomposed replicating structures. The sizes of these large linear structures were > 2000 kb and could not be estimated correctly because they are beyond the range of photography. Moreover, the “core” center had
disappeared. A large number of reticular small networks (~0.5–2 kb per entity, indicated by long dotted arrows in Figs. 11A-b2 and -b3) appeared in this relaxed mtDNA structure. Some large square networks (~10–20 kb per square entity, indicated by the open arrowhead) were also found (Fig. 11A-b2, 11B). Holliday
junctions were present at the corner of each large square structure (short dotted arrow), which suggests the close relationship of these large square structures with RDR.
Interestingly, the core structure also disappeared after the freeze-thaw treatment.
A large number of small reticular networks (~0.5–2 kb) formed and surrounded the core structure without the emergence of a regular/typical rosette structure at the beginning of the mtDNA synthesis (Fig. 12). This reticular network around the core appeared most frequently in the 27 °C-6 hr, 27 °C-12 hr and day-1 mitochondria. In Figure 12A, the mtDNA was purified from 27 °C-6 hr immersed cotyledons; Figures 12B, C, E, and F are from the 27 °C-12 hr immersed cotyledons, and Figure 12D is from day-1 cotyledons. These reticular small networks show a tendency to expand in size, form RDR forks and concentric petal-like structures around the core and, finally, build up a rosette structure as shown in Figure 12F.
In the foregoing study, I proposed that the “core” may represent highly
condensed mtDNA with or without the binding of RNA/protein that converted into a
long linear form with various attached sub-DNA structures in vitro. This form may comprise the complete or partial mitochondrial genome and play an initial and central role in mtDNA RDR. EtBr treatment (10-50 μg/ml) to relax the DNA helix structure before DNA spreading (Fig. 13A) resulted in incomplete decomposition of rosette structures. Therefore, the core could be loosened by EtBr.
In the RDR model, replication initiates at the D-loop formation; new DNA is synthesized on the leading strand and establishes the replication fork (Oldenburg &
Bendich, 2001; Bendich, 2010). Previous study of tobacco mitochondria showed that the wb DNA contained single-stranded mtDNA (Oldenburg & Bendich, 1996).
Single-stranded DNA was also found in the EM analysis here and was evidenced by nuclease digestion (Fig. 13B). Studies of mammalian mtDNA replication suggest that RNA is transcribed initially at the lagging strand before conversion by DNA and produces an RNA-DNA intermediate (Pohjoismaki et al., 2010). The possibility of different buoyant densities of RNA-DNA hybrid and DNA-DNA double strands resulting in the sub-populations in the AUC assay was ruled out. RNaseH treatment analysis revealed no RNA-DNA hybrid present in the mtDNA population (Fig. 14).
Heat denaturation treatment of 27 °C-12 hr mtDNA resulted in a single peak with a high buoyant density of 1.7133 g/cm3.
Some current studies support that the core consists of a DNA–protein complex (Dudareva et al., 1988; Backert & Borner, 2000; Kaufman et al., 2007; Junier et al., 2010; Rebelo et al., 2011). However, mtDNA from CsCl gradient LC-MS/MS did not reveal a significant candidate for the possible proteins bound with the mtDNA rosette core, perhaps because of the low protein level in the DNA–protein complex at the core region in CsCl gradient purified DNA.
8. RDR recombination assessed by real-time PCR
To assess the extent of recombination activity at different stages, two
representative repeats, “E” and “F”, in the mitochondrial genome of mung bean used in recombination activity testing by real-time PCR were selected according to
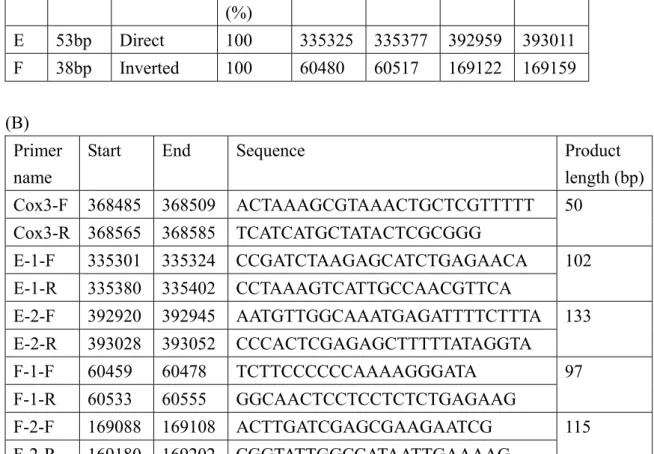
previous study of the mung bean mitochondrial genome (Table 1A) (Alverson et al., 2011b). The diagram of the main genome real-time PCR product E1/E2 and F1/F2 and corresponding recombination forms E1’/E2’ and F1’/ F2’ is in Figure 15A. The analysis of the repeat’s flanking sequences (a, b, c and d), direct repeat (E), and invert repeat (e, f, g and h) (F) are in Table 1B and Figure 15A. Primers for real-time PCR assay are in Table 1B (cox3 was a reference sequence).
The relative quantification of the real-time PCR products for the main genome are in Figures 15B-a (E1 and E2) and C-a (F1 and F2). The recombination products are in Figures 15B-b (E1’ and E2’) and C-b (F1’ and F2’). The relative recombination activity between E1 and E2/F1 and F2 is shown in Figures 15B-c and C-c,
respectively. The short repeat regions E1 and E2/F1 and F2 may undergo low recombination activity (~3.02-4.79x 10 -4 between E1 and E2; 1.22-1.79- x10 -4 between F1 and F2). Short repeats in cotyledon mtDNA with recombination activity may be involved in RDR. Recombination activity did not differ among different-aged mtDNA even if RDR activity differed among different-aged mitochondria.
9. Mitochondrial nucleoproteins present in cotyledon mitochondria with cotyledon aging during mung bean seed germination
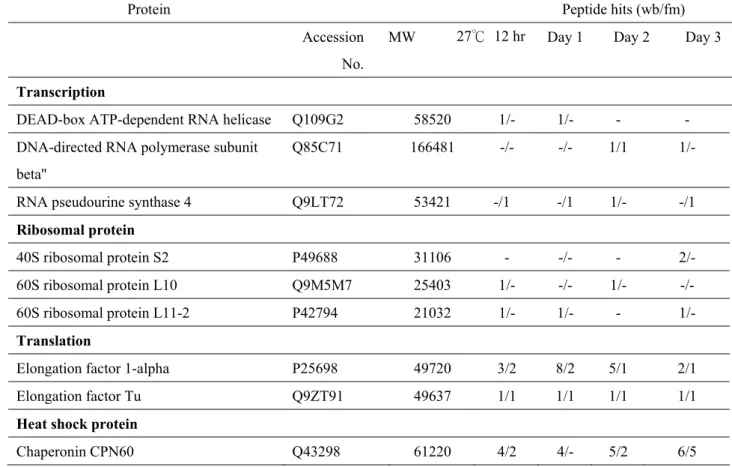
To determine the possible function in different-stage mtDNA, native mt-nucleoids were purified by PFGE fractionation to obtain wb and fm
mitochondria moved in front of fm mtDNA–protein complexes (supporting figure in Table 3B-a).
Elongation factor 1- appeared in the wb DNA–protein complex of 27 °C-12 hr to day-3 cotyledon, but only 27 °C-12 hr and day-1 fm DNA–protein complex showed the same elongation factor. Chaperonin protein CPN 60 and heat shock protein 70 kDa were visible in both wb and fm DNA–protein complexes harvested from 27 °C-12 hr to day-3 cotyledon mitochondria. Some membrane binding proteins, such as ATPase , ATPase, and ANT1, appeared in fm and wb DNA–protein
complexes. The outer membrane protein Porin was also found in every stage. Actin is associated with wb DNA–protein complex of cotyledon mitochondria, from our lab’s previous data (Lo et al., 2011). As the marker protein of mt-nucleoid and joined with mtDNA replication (Edmondson et al., 2005), single-stranded DNA binding protein (mtSSB) was not found in either fm or wb DNA–protein complexes. According to our lab’s unpublished data, mtSSB can be found in day-3 seedling mt-nucleoids isolated by sucrose gradient on LC-MS/MS.
I also detected mtSSB protein in cotyledon mitochondria from all stages by western blot analysis (Table 3B-b). The absence of ssDNA binding protein and the other proteins associated with mtDNA replication may be due to low recovery of the wb/fm protein complex from PFGE gel, and hence, functional proteins associated with mtDNA replication with cotyledon aging cannot be shown (Table 3A).
IV. Discussion
In this thesis, I studied mtDNA replication in mung bean cotyledons to understand the association with the dynamics and changes of physical
structure/conformation during cotyledon development under specific experimental conditions. Combining flow cytometry, PFGE, EM, real-time PCR and biochemical analyses, the results support the following conclusions. (1) MtDNA replication in cotyledon proceeds by a multiple recombination-dependent initiation process generating an organizationally complex physical structure that mediates
recombination and replication. (2) The “rosette core” might contain highly condensed mtDNA with a replication origin and plays an initial and central role in mtDNA replication. (3) A freeze-thaw treatment on mtDNA opened up the unusually large and complex physical structure observed during the active phase of mtDNA replication to reveal a collection of conventional recombination markers, which further confirmed its functional role as an RDR intermediate. (4) Short repeats are actively involved in mtDNA recombination-dependent initiation and likely also in formation of
recombination-mediated structures. Our findings support the earlier interpretation that RDR is the principal mechanism for plant mtDNA replication (Bendich, 1996;
Oldenburg & Bendich, 1996; Backert et al., 1997a; Backert & Borner, 2000;
Oldenburg & Bendich, 2001) and provide certain new insights into plant mtDNA replication and its association with the changes of structure complexity during plant development.
Throughout mung bean cotyledon development, a large fraction of mtDNA migrated as a smear without bands into the PFGE gel with size equivalent to approximately 50 to 200 kb linear DNA markers; most of the remaining mtDNA remained in the well, with a minor fraction falling in between the fast-moving (fm)
and well-bound (wb) zones. The relative amount of fm and wb forms varied significantly during cotyledon development (Fig. 2). This result agrees with earlier findings indicating that plant mtDNA consists of a mixture of monomers and head-to-tail concatermers of circular and linear molecules together with highly complex branched structures (Bendich, 1993; Backert et al., 1996; Bendich, 1996;
Oldenburg & Bendich, 1996; Backert & Borner, 2000; Oldenburg & Bendich, 2001).
The mobility of the fm form of the 27 ºC-12 hr cotyledon mtDNA was significantly retarded and the degree of this retardation was augmented further in day-1 and -2 samples so that most of the fm constituents moved slower than did the 200-kb linear marker (Figs 2B-a,b, lanes 5 -7). A proteinase pre-treatment regressed this retardation in day-1 samples (Fig. 2B-a,b, lane 8). These results suggest that a large fraction of the 27 ºC-12 hr, day-1 and -2 cotyledon mtDNA contains a proteinase K-sensitive structure that migrated slower than the 50- to 200-kb linear DNA marker in PFGE (Fig. 2). Protein(s) associated with this fast-moving component are likely involved in the DNA replication process or in mitochondrial gene function.
To slow the development process, our germination condition included a
prolonged low temperature (4 ºC) imbibition at the onset to enhance the synchrony for capturing the detailed mtDNA replication with accompanying changes to its physical structure. At this time point (4 ºC-12 hr), EM data indicated that 93.5% of total mtDNA length was taken up by linear plus branching linear molecules, with 5.5% by simple rosette plus simple reticular rosette-like structures (Table 2). Also, many long linear molecules (150~210 kb; most > 50 kb) was found (Fig. 7D). Flow cytometry also revealed a very low amount of DAPI-stained DNA present in individual mitochondria at this time point (Fig. 1A-a). Throughout all cotyledon development stages, the relative amount of circular molecules was low. The highest value, 2.2%,
with size < 3 kb was found in the 27 ºC-12 hr sample, with mtDNA synthesis at the onset of its most active phase (Table 2 and Fig. 5).
At the early cotyledon developmental stage, 4 ºC-12 hr, P32-dCTP was
incorporated into the fm form, with very little incorporation into the wb form during a short 60-min pulse under our in organello experimental conditions. This finding is in contrast to that detected for the 27 ºC-12 hr sample, in which the wb fraction was clearly labeled. The two intervening samples, 4 ºC-12 hr + 27 ºC-2 hr and 4 ºC-12 hr + 27 ºC-6 hr, exhibited a gradual increase in wb labeling (Fig. 5a). In day-1 and -2 samples, considerable more P32-dCTP labeling was found in the newly synthesized fm than wb fraction (Fig. 5b). During early cotyledon development stages, the
newly-synthesized fm DNA predominated over the wb component, which constituted only a minor fraction. This situation lasted until the onset of the most active synthesis phase at 27 °C-12 hr, when the newly-synthesized wb fraction became sizable. Thus, formation of highly complex mtDNA (wb form) was closely related to high mtDNA synthesis activity. The quantity of mtDNA per mitochondrion was also coordinately increased and decreased with mtDNA synthesis activity (Figs. 1 and 5). This high fluctuation and exceedingly dynamic characteristics of the organelle DNA content and complexity was found previously (Backert & Borner, 2000; Bendich, 2004;
Oldenburg & Bendich, 2004; Bogenhagen, 2010).
As seen by EM, all mtDNA samples analyzed during mung bean cotyledon development invariably contained mixtures of different structures and sizes (linear, branching linear, rosette, reticular rosette and a few circular, of < 12 kb, see Table 2).
The summation of each samples' composition in terms of percentage of overall length of each structure type over the total sum of length was measured for all types
regardless of the size of each individual molecule (Table 2). Giant rosettes > 500 kb
were scarce in number: 8, 10 and 2 found in 27 ºC-12 hr, day-1 and day-2 cotyledon mtDNA, respectively, on EM. The data for giant rosettes were not included in Table 2.
The scarce amount of giant rosettes may not represent what exists in vivo, because the structure of such complexes and huge mtDNA may easily change conformation and integration during mitochondria isolation and mtDNA purification. An image from each time was chosen to emphasize the representativeness and uniqueness (Figs. 7 and 8).
Our findings øf a rosette-like structure during mtDNA replication agrees well with that shown previously in Chenopodium album (Backert & Borner, 2000). From the architecture and timing of the appearance of regular rosette structure, including giant rosettes, corresponds well with the multi-fibered, comet-like forms of ethidium- bromide-stained mtDNA seen in moving pictures during electrophoresis of several plants (Bendich, 1996). Our results are consistent with and support the earlier interpretations that those large complex structures with size often greater than the genome size (presented in Figs. 9 to 11) represent the RDR intermediates and their formation has been attributed to multiple 3' overhang invasion of homologous regions spanning the repeats to initiate mtDNA replication. After multiple rounds of RDR, the newly replicated DNA results in a very complex branched network (Backert & Borner, 2000; Oldenburg & Bendich, 2001; Bendich, 2010; Gerhold et al., 2010; Gualberto et al., 2014).
The steepest fluctuation in structure composition was observed with the
transition from dry seed to the 4 ºC-2 hr sample, showing a decrease in proportion of linear + branching linear molecules from 85.8% to 8.8% and a corresponding increase in proportion of simple rosette + simple reticular rosette molecules from 13.6% to 91%. This fluctuation pattern closely resembled that observed for the transition from
4 ºC-12 hr to 4 ºC-12hr + 27 ºC-2 hr, in which linear + branching linear molecules decreased in proportion from 93% to 12% and simple rosette + simple reticular rosette molecules increased from 5% to 87.7%. This parallel trend of changing mtDNA composition (Table 2 and Fig. 7) suggesting prolonged low temperature imbibition may have rendered the imbibing seeds to a state somewhat resembling that of dry seeds in terms of the physical structure. The active mtDNA synthesis with
regular/giant rosettes but not simple rosettes is shown in Figures 5 and 7. These results clearly show that the linear and rosette mtDNA structures can be converted in vivo during early cotyledon development, which is affected by the temperature and
duration applied to the seed incubation.
The highly complex giant rosette mtDNA molecule in Figure 10A contains some unique sub-units. Several mtDNA rosettes, presumably replication units that are surrounded by a large circular DNA, are connected to each other. Individual rosette structures with similar (Fig. 10B-a) or various (Fig. 10B-b) complexities in their structures are linked to each other (core to core) by DNA junctions. Figure 10B-a presents three rosette structures with more similar structural complexity, which
indicates their similarity in terms of the re-initiation time (see also box b of Fig. 10A).
The rosettes with different RDR re-initiation times may result in rosettes with various complexities (Fig. 10B-b). This result confirms that the connected rosette structures are part of one giant mitochondrial molecule (Fig. 10A). Putative replication forks are often found near the center of the rosette core and appear as components that extend out from the center core (Figs. 9 and 10). The stretched-out forks and layers of concentric, petal-like structures develop during mtDNA RDR. The satellite-rosette cores represent re-initiation sites of mtDNA replication that surround the parental replicating core and form satellite architecture.
In the freeze-thaw treatment study, the regular-sized rosette structure (~40-200 kb) lost its core composition and converted to linear DNA with a size that was much longer than genome sizes plus various sub-structure attachments (>2000 kb) (Fig. 8).
The core in the rosette-like structure contained condensed plastid DNA, which was clearly demonstrated by Oldenburg and Bendich (2004). These exceedingly long DNA molecules may be expanded from the relaxation of rosette core, which probably contain the whole or partial mitochondrial genome and replication origin. The center core (arrow in Figs. 8 and 9) might be the initial site for mtDNA replication, and the satellite-cores (arrowheads in Fig. 9) might be the subsequent re-initiation mtDNA replication sites on the same mtDNA molecule when the original mtDNA replication process is not yet complete. The continuous serial re-initiation of mtDNA replication on the same molecule may result in a complex branched structure and expand a single mtDNA into many genome-equivalent mtDNAs and form a giant complex mtDNA compartment (Figs. 9-11). These recombination replication intermediates that contain putative replication forks, D-loops and Holliday junctions all indicate that RDR occurs in mung bean cotyledon mitochondria. Because freeze-thaw treatment is effective in opening up the tightly-folded rosette structures, single-stranded DNA regions, with or without associated protein(s), may be involved in holding the tightly folded state of the rosette structures.
After freeze-thaw treatment, the rosette structure converted to a much larger molecule; also an abundance of small networks appeared on the same DNA molecule (~0.5-2 kb for each entity, long dotted arrow in Fig. 11A-b). Moreover, some large square networks (~10-20 kb per square entity, open arrowhead in Figs. 11A-b, 11B) were also found. The large networks arranged in rows showed Holliday junctions (short dotted arrows). Interestingly, similar small reticular networks around the core
structure were present in the very early mtDNA replication stages, especially in the 27 °C-6 hr, 27 °C-12 hr mitochondria and, to a lesser extent, day-1 mitochondria (Fig.
12). The small reticular networks might be produced by short repeat DNA sequences and may be associated with the initiation of RDR. According to real-time PCR analysis (Fig. 15), short repeats in cotyledon mtDNA showed recombination activity in vitro, which supports that short repeats may be involved in RDR initiation. The
proposed progressive development of small networks around the core is shown in Figure 12. Small networks gradually convert to loose petal-like structures and then form the rosette-like structure. The RDR forks are formed during this progressive process (Fig. 12F). The small networks, which appear around the core in large numbers, may be related to the short repeat recombination in mung bean mtDNA for initiation of RDR.
Recombination occurs in shorter repeats, and the production of rearranged mtDNA molecules was reported previously (Andre et al., 1992; Shedge et al., 2007;
Woloszynska & Trojanowski, 2009; Cappadocia et al., 2010; Marechal & Brisson, 2010; Woloszynska, 2010; Sloan, 2013). Some short repeats in various plant
mitochondrial genomes were suggested as hot spots for recombination (Shedge et al., 2007; Cappadocia et al., 2010). Although the mung bean mitochondrial genome lacks the large repeat, a PCR survey of 6 short (38 to 175 bp) repeats revealed evidence for recombination across all of them (Alverson et al., 2011b). In agreement with these data, my findings also indicate that mung bean short repeats are active in
recombination (see Fig. 15). Moreover, the recombination activity detected by quantitative PCR did not vary throughout cotyledon development. Whether recombination activity of short repeats in vivo could be accurately inferred from real-time PCR results alone is questionable; a recombinant clone involving a 175-bp